프테리고투스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
프테리고투스는 실루리아기부터 데본기까지 존재했던 바다전갈의 일종이다. 몸길이가 최대 1.75m에 달하는 대형 바다전갈로, 굽은 발톱과 넓적한 꼬리를 가지고 있으며, 다른 바다전갈과 달리 걷는 다리가 작고 가시가 없었다. 프테리고투스는 뛰어난 시력과 강력한 협각을 이용해 활동적으로 먹이를 사냥하는 포식자였으며, 얕은 바다, 호수, 강 등 다양한 환경에서 서식했다. 프테리고투스는 전 세계적으로 발견되며, 특히 유럽, 북아메리카, 남아메리카 등에서 화석이 발견된다. 프테리고투스는 프테리고투스과에 속하며, 아쿠티라무스, 자켈로프테루스와 유사한 특징을 공유한다.
더 읽어볼만한 페이지
- 1839년 기재된 화석 분류군 - 글립토돈
남아메리카에 서식했던 글립토돈은 갑옷 같은 등갑과 꼬리 갑옷을 가진 거대한 멸종 포유류로, 리처드 오웬에 의해 명명되었으며 기후 변화와 인간의 영향으로 멸종했을 것이라는 가설이 있다. - 데본기의 동물 - 이크티오스테가
이크티오스테가는 데본기 후기에 살았던 사지동물로, 어류와 육상 동물의 특징을 모두 가지며, 튼튼한 사지와 척추를 가지고 물가에서 육상으로의 진출을 시도했던 초기 사지동물의 생활 방식을 보여준다. - 데본기의 동물 - 야이켈롭테루스
야이켈롭테루스는 삼각형 미부와 경사진 협각 지절을 가진 멸종된 유해류 속으로, 한때 프테리고투스 속으로 분류되었으나 재분류되었으며, 거대한 크기와 발달된 시력을 가진 최상위 포식자로 추정된다. - 실루리아기의 동물 - 믹소프테루스
믹소프테루스는 중생대 실루리아기에 살았던 최대 75cm 크기의 육식성 바다전갈로, 단단한 외골격과 헤엄다리를 가지며 매복 사냥을 했을 것으로 추정된다. - 실루리아기의 동물 - 에우립테루스
에우립테루스는 고생대 실루리아기에 번성했던 멸종된 절지동물로, 바다전갈강에 속하며 몸은 전신과 후신으로 나뉘고 겹눈과 다리를 가진 육식성 또는 청소부 역할을 했을 것으로 추정되는 라우라시아 대륙에 분포한 거대한 종이다.
| 프테리고투스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Pterygotus |
| 명명자 | 아가시, 1839년 |
| 속의 유형 종 | †Pterygotus anglicus |
| 유형 종 명명자 | 아가시, 1849년 |
| 속 동의어 | Curviramus Reudemann, 1935년 |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 절지동물문 |
| 아문 | 협각아문 |
| 강 | 퇴구강 |
| 목 | †우미사소리목 |
| 아목 | †우미사소리아목 |
| 상과 | †프테리고투스상과 |
| 과 | †다이오우미사소리과 |
| 화석 기록 | |
| 시기 | 실루리아기 중기 - 데본기 후기, 약 4억 2800만 년 전 - 3억 7220만 년 전 |
| 미확실한 종 | |
| 동종이명 | |
| P. anglicus의 동종이명 | P. atlanticus (클라크 & 루데만, 1912년) P. minor (우드워드, 1864년) |
| P. barrandei의 동종이명 | P. beraunensis (셈퍼, 1898년) |
2. 형태
가장 큰 종인 ''P. grandidentatus''는 몸길이가 1.75m에 달했는데, 이는 ''프테리고투스''가 존재했던 가장 큰 바다전갈 중 하나임을 보여주지만, 일부 근연종(예: ''Acutiramus''와 ''Jaekelopterus'')은 길이 면에서 이를 능가했다.[2] 거대한 종도 있었지만, 많은 종들은 크기가 상당히 작았다. 가장 작은 종인 ''P. kopaninensis''는 길이가 50cm에 불과했다.[3]
''프테리고투스''는 바다전갈의 프테리고투스과에 속하며, 이 과는 실루리아기부터 데본기에 걸쳐 존재했던 고도로 파생된 바다전갈 집단으로, 협각 (첫 번째 다리 쌍)과 미부 (몸의 가장 뒷부분)를 포함한 여러 특징에서 다른 집단과 차별화된다. 프테리고투스과의 협각은 확대되고 튼튼하여, 일부 현대 갑각류의 발톱과 유사하게, 활발한 먹이 포획에 사용하도록 분명히 적응되었으며 다른 바다전갈 그룹의 협각보다 발톱에 잘 발달된 이빨이 있었다.[4] 다른 프테리고투스과 바다전갈과 같이 비늘 모양의 장식으로 덮여 있던 몸의 나머지 부분과 달리, 발톱에는 어떤 종류의 장식도 없었다.[5] 또한, 발톱의 끝 부분은 관련 종인 ''Erettopterus''의 발톱 끝에 있는 날카로운 점과 달리 둥글고 구부러졌다.[6] 프테리고투스과의 미부는 납작하고 넓어졌으며, 헤엄칠 때 방향타로 사용되었을 것이다.[7] 이들의 걷는 다리는 작고 가늘며 가시가 없어 육지에서 걸을 수 없었을 것이다.[8][2]
''프테리고투스''는 주두 (발톱)의 굽은 말단 가장자리를 통해 다른 프테리고투스과와 구별된다. 전체절 (머리)은 부사다리꼴 (둥근 모서리가 있는 사다리꼴)이며, 겹눈은 앞쪽 모서리 가장자리에 위치한다. 미부는 가운데를 따라 뚜렷한 등쪽 용골(또는 키)이 있으며, 짧은 가시로 끝난다.[8]
다른 바다전갈과 마찬가지로 몸은 전체 (prosoma, 머리 가슴 부위)와 후체 (opisthosoma, 복부)의 두 부분으로 나뉜다.[56] . 대왕바다전갈과의 공유 형질로서, 편평하고 세로로 긴 몸의 표면에는 비늘 모양의 돌기가 나 있고, 이빨이 있는 발달된 협각, 단순한 제2-4 다리, 분절되지 않은 생식지, 그리고 주걱 모양의 미절을 특징으로 하는 바다전갈이다[57][58][69] .
전체는 둥근 사다리꼴[59][57][58]의 등갑 (carapace, prosomal dorsal shield)으로 덮여 있으며, 각각 한 쌍의 홑눈 (중안)과 큰 겹눈 (측안)은 그 중앙과 양쪽 앞쪽에 배치된다. 각 겹눈은 4,000개 이상의 홑눈으로 구성된다[60] . 동과의 다른 속과 마찬가지로, 6쌍의 전체 부속지 (관절지) 중 첫 번째 한 쌍인 협각 (chelicerae)은 팔처럼 뻗어 있으며, 끝의 집게는 견고하고, 가동지와 부동지 내측에는 크고 작은 이빨이 배치된다. 가동지는 끝이 부드럽게 굽어 있고, 각 이빨은 뒤쪽으로 구부러져 있으며, 가장자리에는 톱니가 없다[57][58] . 아크티람스나 제켈로프테루스 정도는 아니지만, 부동지, 가동지 모두 수 개의 큰 이빨 중 중앙의 한 개가 가장 발달되어 있다[57][58] . 나머지 5쌍의 부속지는 기절에 턱기부가 있는 다리이며, 그중 제2-4 다리는 가늘고 돌기가 없는 보각 모양이며[59], 제5 다리는 다른 대부분의 바다전갈아목 (Eurypterina)의 종류와 마찬가지로, 패들 모양의 유영각으로 특화되었다[59][57] . 유영각의 턱기부에는 12개 정도의 이빨이 늘어서 있다[57] . 제1 다리(촉지, pedipalp)의 자세한 내용은 불분명하지만, 아마 엘레토프테루스처럼 단축화된 것으로 생각된다[61][62] .
12개의 등판과 1개의 미절 (telson)이 보이는 후체는 세로로 길고, 제5 다리 사이에 끼워 넣은 복면의 하층판 (metastoma)은 끝이 안쪽으로 움푹 들어간 타원형이다[57][58] . 동과의 다른 속과 마찬가지로, 하층판과 제5 다리 바로 뒤에 있는 생식 구개(genital operculum)는 마디가 유합하고, 중앙의 생식지(genital appendage)는 분절되지 않는다[69] . 생식지 중 type-A는 주걱 모양 또는 곤봉 모양, type-B는 단순한 마름모 모양[57][58] . 주걱 모양의 미절은 둥글고 뒷부분이 뾰족하며, 배쪽 정중선은 융기한다[57][58] . 이 정중선은 한 개의 수직 주걱으로 발달한 경우가 있다[55] . 미절 바로 앞의 최종 체절(pretelson)도 배쪽 정중선이 융기하고, 미절에 맞춰 어느 정도 옆으로 넓어진다[57][58] .
본 속은 대형 바다전갈로 알려져 있지만, 체장은 종에 따라 크게 다르다고 한다[64] . 기지 최대급의 ''P. grandidentatus''는 전체 길이가 1.75m에 달한다고 추정되며[64], 이는 동과에서 최대급의 바다전갈로 여겨지는 제켈로프테루스나 아크티람스에 필적한다[63] . 기지 최소 종은 ''P. kopaninensis''로, 전체 길이는 약 50cm 전후로 추정된다[64] .
2. 1. 머리가슴
프테리고투스는 다른 바다전갈과 마찬가지로 몸이 전체(머리가슴 부위)와 후체(복부)의 두 부분으로 나뉜다.[56] 전체는 둥근 사다리꼴[59][57][58]의 등갑으로 덮여 있으며, 한 쌍의 홑눈과 큰 겹눈이 각각 중앙과 양쪽 앞쪽에 배치된다. 각 겹눈은 4,000개 이상의 홑눈으로 구성된다.[60]6쌍의 전체 부속지(관절지) 중 첫 번째 쌍인 협각은 팔처럼 뻗어 있으며, 끝의 집게는 견고하고 가동지와 부동지 내측에는 크고 작은 이빨이 배치된다. 가동지는 끝이 부드럽게 굽어 있고, 각 이빨은 뒤쪽으로 구부러져 있으며, 가장자리에는 톱니가 없다.[57][58] 아크티람스나 제켈로프테루스처럼 부동지, 가동지 모두 수 개의 큰 이빨 중 중앙의 한 개가 가장 발달되어 있다.[57][58]
나머지 5쌍의 부속지는 기절에 턱기부가 있는 다리이며, 그중 제2-4 다리는 가늘고 돌기가 없는 보각 모양이다.[59] 제5 다리는 바다전갈아목(Eurypterina])의 종류와 마찬가지로, 패들 모양의 유영각으로 특화되었으며,[59][57] 턱기부에는 12개 정도의 이빨이 늘어서 있다.[57] 제1 다리(촉지)의 자세한 내용은 불분명하지만, 엘레토프테루스처럼 단축화된 것으로 추정된다.[61][62]
2. 2. 배
다른 바다전갈과 마찬가지로 몸은 전체(머리 가슴 부위)와 후체(복부)의 두 부분으로 나뉜다.[56] 대왕바다전갈과의 공유 형질로서, 편평하고 세로로 긴 몸의 표면에는 비늘 모양의 돌기가 나 있고, 이빨이 있는 발달된 협각, 단순한 제2-4 다리, 분절되지 않은 생식지, 그리고 주걱 모양의 미절을 특징으로 하는 바다전갈이다.[57][58][69]
12개의 등판과 1개의 미절(telson)이 보이는 후체는 세로로 길고, 제5 다리 사이에 끼워 넣은 복면의 하층판(metastoma)은 끝이 안쪽으로 움푹 들어간 타원형이다.[57][58] 동과의 다른 속과 마찬가지로, 하층판과 제5 다리 바로 뒤에 있는 생식 구개(genital operculum)는 마디가 유합하고, 중앙의 생식지(genital appendage)는 분절되지 않는다.[69] 생식지 중 type-A는 주걱 모양 또는 곤봉 모양, type-B는 단순한 마름모 모양이다.[57][58] 주걱 모양의 미절은 둥글고 뒷부분이 뾰족하며, 배쪽 정중선은 융기한다.[57][58] 이 정중선은 한 개의 수직 주걱으로 발달한 경우가 있다.[55] 미절 바로 앞의 최종 체절(pretelson)도 배쪽 정중선이 융기하고, 미절에 맞춰 어느 정도 옆으로 넓어진다.[57][58]
2. 3. 크기
프테리고티드과는 지금까지 살았던 절지동물 중 가장 큰 종을 포함하며, 몇몇 종은 길이가 2미터가 넘는다.[2] ''프테리고투스''가 프테리고티드 중에서 가장 큰 종은 아니었지만, 몇몇 종은 길이가 1미터를 넘을 정도로 컸다.[3]가장 큰 종은 ''P. grandidentatus''로 알려져 있으며, 가장 큰 고립된 협각 조각은 길이가 1.75m에 달하는 것으로 추정된다.[3] 에스토니아의 ''P. impacatus''는 두 번째로 큰 종으로, 가장 큰 조각화된 잔해는 길이가 1.65m에 달하는 것으로 추정된다.[3] 모식 종인 ''P. anglicus''는 헨리 우드워드가 1866년과 1878년 사이에 발견한 큰 등판을 바탕으로 길이가 1.6m까지 자랐다. 길이가 40cm가 조금 넘고 너비가 10.5cm인 이 등판은 머리 가슴의 시작부터 꼬리 자루의 끝까지의 전체 길이가 1.6m인, 확장된 협각을 포함하면 전체 길이가 2m를 초과하는 유르프테리드를 시사한다.[9] 오하이오의 데본기에 살았던 ''P. carmani''는 길이가 1.5m를 넘었을 가능성이 크다.[5]
''P. cobbi''(1.4m), ''P. barrandei''(1.26m) 및 ''P. denticulatus''(1.2m) 종도 길이가 1미터를 넘었다. 더 작은 종으로는 ''P. floridanus''(90cm), ''P. lightbodyi''(75cm), ''P. arcuatus''(60cm), ''P. bolivianus''(55cm)가 있으며, 가장 작은 종인 ''P. kopaninensis''는 길이가 50cm이다.[3]
3. 생태
프테리고투스류 협각류의 협각 형태와 시력은 이들을 뚜렷한 생태학적 그룹으로 구분한다. 절지동물의 시력을 결정하는 주요 방법은 겹눈의 렌즈 수와 눈 사이 각도(IOA, 인접한 렌즈의 광학 축 사이의 각도)를 결정하는 것이다. IOA는 절지동물의 서로 다른 생태적 역할을 구별하는 데 사용될 수 있기 때문에 특히 중요한데, 현대의 활동적인 절지동물 포식자에서는 낮다.[36]
''Pterygotus anglicus''와 ''자켈로프테루스 레나니아에''는 모두 시력이 매우 높았는데, 연구자들은 낮은 IOA와 겹눈의 많은 렌즈 수를 관찰하여 이를 확인할 수 있었다. ''프테리고투스''의 협각은 확대되고 튼튼했으며, 굽은 자유 가지와 서로 다른 길이와 크기의 치아를 가지고 있었는데, 이는 현존하는 전갈과 갑각류에서 강력한 찌르고 잡는 능력에 해당하는 적응이다. ''프테리고투스''와 ''자켈로프테루스''의 IOA 값은 수준 높고 활동적인 현대 포식성 절지동물의 값과 일치하며, 이는 이들이 시각적이고 활동적인 포식자임을 나타낸다.[36]

알려진 모든 프테리고투스류는 협각 발톱을 가지고 있었다. 협각의 첫 번째 관절은 프로소마 또는 "머리"에 위치한 판인 에피스토마에 연결되어 있으며, 전체 부속지를 비틀어 회전시킬 수 있었을 것이며, 이로 인해 연구자들은 협각의 기능이 방어만을 위한 것이거나 심지어 주요 기능이 아닐 것이고, 먹이를 잡아서 입으로 전달하는 것이었을 것이라고 결론 내렸다. 잡힌 먹이는 입에 들어갈 수 있도록 더 작은 조각으로 부서져야 할 것이며, 협각류의 입은 현대 게의 입보다 큰 조각을 삼키는 데 덜 적응되어 있었다. 협각류의 보행 부속지는 어떤 것도 자르거나 운반하거나 잡을 수 없었으므로, 이러한 작업 역시 협각의 역할이었을 것이다. 게의 경우, 발톱은 음식을 찢어 작게 만들고 나서 조각들을 입으로 운반한다. 협각을 가진 현대 절지동물에서 보이는 먹이 섭취 과정을 바탕으로, 한 발톱은 먹이를 잡고 다른 발톱은 조각을 잘라 지속적이고 단순한 움직임으로 입으로 운반했을 것이다.[9]
''프테리고투스''는 완전한 해양 환경뿐만 아니라 호수 및 강 시스템에서도 서식했으며, 오늘날 이들의 화석은 그러한 환경을 나타내는 흔하고 다양한 화석과 함께 발견되고 있다.[37] 특히, ''P. barrandei''와 ''P. cobbi''는 ''아쿠티라무스''(Acutiramus) 화석과 관련된 해양 환경에서 발견된다. ''P. anglicus''는 올드 레드 사암[39]과 캠벨턴 지층[38]에서 발견되는데, 두 곳 모두 담수 호수와 강 시스템의 예시이다.[39]
''프테리고투스''는 ''아쿠티라무스''보다 더 널리 분포했으며, 따라서 영국의 ''P. lightbodyi''와 같이 ''아쿠티라무스''가 없는 많은 종도 존재한다.[14] 또한 해양 종인 ''P. australis''는 특히 깊은 물에서 서식했다.[40] ''P. australis''와 같은 시대, 같은 장소에서 발견되는 유일한 화석 생물은 ''데드로크리누스''(Dendrocrinus)와 ''코프토노크리누스''(Kooptoonocrinus) 속의 바다나리와 ''프로타스터''(Protaster) 속의 거미불가사리이다.[41]
''P. kopaninensis''의 모식 표본이 발견된 퇴적층은 다른 많은 동물 화석도 보존하고 있다. 그중에는 ''레오나스피스''(Leonaspis), ''라피오포루스''(Raphiophorus)와 파편적인 하페티드 삼엽충이 있다. 또한 ''카르디올라''(Cardiola)와 같은 이매패류, ''모노그라프투스''(Monograptus)와 같은 필석도 보존되어 있다.[24]
''프테리고투스''는 일반적으로 다양한 유르프테리드 동물군이 있는 생태계에서 발견되며, 스코틀랜드의 킵 번 층의 ''P. lanarkensis''는 다른 여러 유르프테리드 속과 함께 발견된다. ''슬리모니아''(Slimonia), ''에우사르카나''(Eusarcana), ''나나휴밀러리아''(Nanahughmilleria), ''파라스타일로누루스''(Parastylonurus), ''에레토프테루스''(Erettopterus) 및 ''카르키노소마''(Carcinosoma)가 있다.[42] 에스토니아의 ''P. impacatus''는 ''에레토프테루스'', ''에리에오프테루스''(Erieopterus), 카르키노소마, ''믹소프테루스''(Mixopterus), ''돌리코프테루스''(Dolichopterus) 및 ''에이시슬롭테루스''(Eysyslopterus) 속과 두미류 어류와 함께 발견된다.[43] ''P. monroensis''는 ''믹소프테루스'', ''휴밀러리아''(Hughmilleria), ''유리프테루스''(Eurypterus) 및 카르키노소마와 함께 서식했다.[44] ''P. nobilis''는 ''아쿠티라무스'', ''에레토프테루스'' 및 ''에우사르카나''의 대표 종과 다양한 코노돈트, 복족류, 두족류, 요각류, 연갑류, 삼엽충 및 이매패류와 연골어류 ''온쿠스''(Onchus)와 함께 서식했다.[45]
데본기 올드 레드 사암 지층에서 발견된 ''P. anglicus''의 화석은 ''에리에오프테루스'', ''파게아''(Pagea), ''타르소프테렐라''(Tarsopterella) 및 ''스타일로누루스''(Stylonurus)를 포함한 다양한 유르프테리드와 극어강 어류 ''이슈나칸투스''(Ischnacanthus) 및 ''메사칸투스''(Mesacanthus), 상어 ''클리마티우스''(Climatius) 및 골갑어류 어류 ''세팔라스피스''(Cephalaspis)와 함께 발견된다.[39] 오하이오의 홀랜드 채석장 셰일 지층에서 발견된 유사한 시대의 ''P. carmani''의 화석은 ''돌리코프테루스'', ''스트로빌롭테루스''(Strobilopterus) 및 에레토프테루스와 같은 다양한 유르프테리드와 프테라스피스강 어류 ''알로크리타스피스''(Allocrytaspis) 및 판피어류 어류 ''에타프시스''(Aethapsis)의 다양한 데본기 동물군을 보존한다.[42] 관련 프테리고투스과의 ''자이켈롭테루스(Jaekelopterus)''의 것으로 추정되는 화석 포라스피드 어류의 펑크 상처는 프테리고투스과가 프테라스피스강과 판피어류와 같은 갑옷을 가진 물고기를 잡아먹을 수 있었음을 보여준다.[36] ''P. siemiradzkii''의 화석이 산출된 퇴적층에서는 ''유리프테루스''와 여러 속의 프테라스피스강 어류 화석이 발견되었다. ''라르노바스피스''(Larnovaspis), ''피알라스피스''(Phialaspis), ''코르바스피스''(Corvaspis) 및 ''포돌라스피스''(Podolaspis)가 있다.[46][47]
웨일스 변경 구역 단층 계통의 실루리아기 퇴적층에서 5종의 ''프테리고투스''가 발견되었다. ''P. ludensis'', ''P. arcuatus'', ''P. grandidentatus'', ''P. lightbodyi'' 및 ''P. denticulatus''이다. 웨일스 변경 구역은 알려진 유르프테리드 동물군 중 가장 광범위한 동물군을 보존하며, 정확한 속과 종은 정확한 시기에 따라 다릅니다. 웬록 시대의 ''P. grandidentatus''는 ''믹소프테루스''와 함께 발견되었고, 중부 루드 로우 시대의 ''P. arcuatus''는 ''살테로프테루스''(Salteropterus), 에레토프테루스 및 카르키노소마와 함께 발견되었으며, ''P. lightbodyi''와 ''P. denticulatus''는 서로 동시대에 살았으며 ''에레토프테루스''와 ''카르키노소마'' 속의 종과 함께 발견되었다. 다운토니안 시대의 ''P. ludensis''는 ''카르키노소마'', ''돌리코프테루스'', ''에레토프테루스'', ''휴밀러리아'', ''파라휴밀러리아''(Parahughmilleria), ''유리프테루스'', ''나나휴밀러리아'', ''마르수피프테루스''(Marsupipterus), 스타일로누루스, 타르소프테렐라, 슬리모니아 및 살테로프테루스로 구성된 다양한 유르프테리드와 함께 발견되었다.[14]
때로는 ''프테리고투스''와 함께 발견되는 알려진 유일한 다른 화석 유르프테리드는 또한 프테리고투스과이다. ''P. marylandicus''는 ''에레토프테루스''와 함께만 발견된다.[48] 잉글랜드에서 발견된 미심쩍은 ''P. taurinus''는 극어강 어류, 전거상목 어류뿐만 아니라 두미류 ''헤미키클라스피스''(Hemicyclaspis) 및 ''티에스테스''(Thyestes)와 같은 다양한 물고기의 유해가 산출된 퇴적층에서 발견되었다.[49]
어떤 경우에는 ''프테리고투스''가 그 서식지에서 알려진 유일한 유르프테리드를 나타냅니다. ''P. bolivianus''는 기준 산지에서 ''메타크리파에우스''(Metacryphaeus) 속의 삼엽충과만 관련하여 발견되었다.[9] ''P. floridanus''는 다른 환경에서는 복족류 (''플렉토노투스''(Plectonotus) 등), 요각류, 부족류, 텐타쿨리타 (''텐타쿨리테스''(Tentaculites) 등) 및 다양한 이매패류 (''에오시조두스''(Eoschizodus), ''플레우로다피스''(Pleurodapis), ''아리사이아''(Arisaiga), ''프테리노펙텐''(Pterinopecten) 및 ''모디오모르파''(Modiomorpha) 포함)만 포함되어 있는 것으로 알려진 환경에서 발견되었다.[50] 캐나다의 ''P. gaspesiensis''는 다양한 이매패류와 복족류와 삼엽충 ''파콥스''(Phacops) 및 연갑류 ''트로피도카리스''(Tropidocaris)가 서식하는 환경에서 발견되었지만 다른 유르프테리드는 발견되지 않았다.[51]
바다전갈과 마찬가지로, 프테리고투스 역시 대부분 바다전갈과에 속하는 유영성 포식자였을 것으로 추정된다.[54][60] 유영각으로 해중을 헤엄치며, 수평과 수직의 날을 가진 미절을 키처럼 사용했을 것으로 추정된다.[65] 협각은 뿌리 부분이 넓고 가동 범위가 넓어 포식과 방어 역할을 동시에 수행했을 것으로 생각된다.[55]
바다전갈과 내에서 본 속은 야켈롭테루스(Jaekelopterus)와 마찬가지로 뛰어난 시력을 가진 활동적인 포식자였을 것으로 여겨진다.[66][60] 동과의 아쿠티라무스(Acutiramus)와 에레토프테루스(Erettopterus)에 비해, 이 두 속의 협각은 튼튼하고 중앙의 이가 강력하여, 먹이를 포획하고 부수는 기능에 더욱 특화되어 있었을 것으로 추정된다.[60] 또한, 본 속은 협각으로 퇴적물을 파내 환형동물을 포식했다는 설도 있으며, 그 포식 행동으로 남겨졌을 것으로 보이는 생흔 화석(Monomorphichnus)도 발견된다.[62]
4. 분포
''프테리고투스''는 완전한 해양 환경뿐만 아니라 호수 및 강 시스템에서도 서식했으며, 오늘날 이들의 화석은 그러한 환경을 나타내는 흔하고 다양한 화석과 함께 발견되고 있다.[37] 특히, ''P. barrandei''와 ''P. cobbi''는 ''아쿠티라무스''(Acutiramus) 화석과 관련된 해양 환경에서 발견된다. ''P. anglicus''는 올드 레드 사암[39]과 캠벨턴 지층[38]에서 발견되는데, 두 곳 모두 담수 호수와 강 시스템의 예시이다.[39] ''프테리고투스''는 ''아쿠티라무스''보다 더 널리 분포했으며, 따라서 영국의 ''P. lightbodyi''와 같이 ''아쿠티라무스''가 없는 많은 종도 존재한다.[14] 또한 해양 종인 ''P. australis''는 특히 깊은 물에서 서식했다.[40] ''P. australis''와 같은 시대, 같은 장소에서 발견되는 유일한 화석 생물은 ''데드로크리누스''(Dendrocrinus)와 ''코프토노크리누스''(Kooptoonocrinus) 속의 바다나리와 ''프로타스터''(Protaster) 속의 거미불가사리이다.[41]
''P. kopaninensis''의 모식 표본이 발견된 퇴적층은 다른 많은 동물 화석도 보존하고 있다. 그중에는 ''레오나스피스''(Leonaspis), ''라피오포루스''(Raphiophorus)와 파편적인 하페티드 삼엽충이 있다. 또한 ''카르디올라''(Cardiola)와 같은 이매패류, ''모노그라프투스''(Monograptus)와 같은 필석도 보존되어 있다.[24] ''프테리고투스''는 일반적으로 다양한 유르프테리드 동물군이 있는 생태계에서 발견되며, 스코틀랜드의 킵 번 층의 ''P. lanarkensis''는 다른 여러 유르프테리드 속과 함께 발견된다. ''슬리모니아''(Slimonia), ''에우사르카나''(Eusarcana), ''나나휴밀러리아''(Nanahughmilleria), ''파라스타일로누루스''(Parastylonurus), ''에레토프테루스''(Erettopterus) 및 ''카르키노소마''(Carcinosoma)가 있다.[42] 에스토니아의 ''P. impacatus''는 ''에레토프테루스'', ''에리에오프테루스''(Erieopterus), 카르키노소마, ''믹소프테루스''(Mixopterus), ''돌리코프테루스''(Dolichopterus) 및 ''에이시슬롭테루스''(Eysyslopterus) 속과 두미류 어류와 함께 발견된다.[43] ''P. monroensis''는 ''믹소프테루스'', ''휴밀러리아''(Hughmilleria), ''유립테루스''(Eurypterus) 및 카르키노소마와 함께 서식했다.[44] ''P. nobilis''는 ''아쿠티라무스'', ''에레토프테루스'' 및 ''에우사르카나''의 대표 종과 다양한 코노돈트, 복족류, 두족류, 요각류, 연갑류, 삼엽충 및 이매패류와 연골어류 ''온쿠스''(Onchus)와 함께 서식했다.[45]
데본기 올드 레드 사암 지층에서 발견된 ''P. anglicus''의 화석은 ''에리에오프테루스'', ''파게아''(Pagea), ''타르소프테렐라''(Tarsopterella) 및 ''스타일로누루스''(Stylonurus)를 포함한 다양한 유르프테리드와 극어강 어류 ''이슈나칸투스''(Ischnacanthus) 및 ''메사칸투스''(Mesacanthus), 상어 ''클리마티우스''(Climatius) 및 골갑어류 어류 ''세팔라스피스''(Cephalaspis)와 함께 발견된다.[39] 오하이오주의 홀랜드 채석장 셰일 지층에서 발견된 유사한 시대의 ''P. carmani''의 화석은 ''돌리코프테루스'', ''스트로빌롭테루스''(Strobilopterus) 및 에레토프테루스와 같은 다양한 유르프테리드와 프테라스피스강 어류 ''알로크리타스피스''(Allocrytaspis) 및 판피어류 어류 ''에타프시스''(Aethapsis)의 다양한 데본기 동물군을 보존한다.[42] 관련 프테리고투스과의 ''자이켈롭테루스''(Jaekelopterus)의 것으로 추정되는 화석 포라스피드 어류의 펑크 상처는 프테리고투스과가 프테라스피스강과 판피어류와 같은 갑옷을 가진 물고기를 잡아먹을 수 있었음을 보여준다.[36] ''P. siemiradzkii''의 화석이 산출된 퇴적층에서는 ''유립테루스''와 여러 속의 프테라스피스강 어류 화석이 발견되었다. ''라르노바스피스''(Larnovaspis), ''피알라스피스''(Phialaspis), ''코르바스피스''(Corvaspis) 및 ''포돌라스피스''(Podolaspis)가 있다.[46][47]
웨일스 변경 구역 단층 계통의 실루리아기 퇴적층에서 5종의 ''프테리고투스''가 발견되었다. ''P. ludensis'', ''P. arcuatus'', ''P. grandidentatus'', ''P. lightbodyi'' 및 ''P. denticulatus''이다. 웨일스 변경 구역은 알려진 유르프테리드 동물군 중 가장 광범위한 동물군을 보존하며, 정확한 속과 종은 정확한 시기에 따라 다릅니다. 웬록 시대의 ''P. grandidentatus''는 ''믹소프테루스''와 함께 발견되었고, 중부 루드 로우 시대의 ''P. arcuatus''는 ''살테로프테루스''(Salteropterus), 에레토프테루스 및 카르키노소마와 함께 발견되었으며, ''P. lightbodyi''와 ''P. denticulatus''는 서로 동시대에 살았으며 ''에레토프테루스''와 ''카르키노소마'' 속의 종과 함께 발견되었다. 다운토니안 시대의 ''P. ludensis''는 ''카르키노소마'', ''돌리코프테루스'', ''에레토프테루스'', ''휴밀러리아'', ''파라휴밀러리아''(Parahughmilleria), ''유리프테루스'', ''나나휴밀러리아'', ''마르수피프테루스''(Marsupipterus), 스타일로누루스, 타르소프테렐라, 슬리모니아 및 살테로프테루스로 구성된 다양한 유르프테리드와 함께 발견되었다.[14]
때로는 ''프테리고투스''와 함께 발견되는 알려진 유일한 다른 화석 유르프테리드는 또한 프테리고투스과이다. ''P. marylandicus''는 ''에레토프테루스''와 함께만 발견된다.[48] 잉글랜드에서 발견된 미심쩍은 ''P. taurinus''는 극어강 어류, 전거상목 어류뿐만 아니라 두미류 ''헤미키클라스피스''(Hemicyclaspis) 및 ''티에스테스''(Thyestes)와 같은 다양한 물고기의 유해가 산출된 퇴적층에서 발견되었다.[49]
어떤 경우에는 ''프테리고투스''가 그 서식지에서 알려진 유일한 유르프테리드를 나타낸다. ''P. bolivianus''는 기준 산지에서 ''메타크리파에우스''(Metacryphaeus) 속의 삼엽충과만 관련하여 발견되었다.[9] ''P. floridanus''는 다른 환경에서는 복족류 (''플렉토노투스''(Plectonotus) 등), 요각류, 부족류, 텐타쿨리타 (''텐타쿨리테스''(Tentaculites) 등) 및 다양한 이매패류 (''에오시조두스''(Eoschizodus), ''플레우로다피스''(Pleurodapis), ''아리사이아''(Arisaiga), ''프테리노펙텐''(Pterinopecten) 및 ''모디오모르파''(Modiomorpha) 포함)만 포함되어 있는 것으로 알려진 환경에서 발견되었다.[50] 캐나다의 ''P. gaspesiensis''는 다양한 이매패류와 복족류와 삼엽충 ''파콥스''(Phacops) 및 연갑류 ''트로피도카리스''(Tropidocaris)가 서식하는 환경에서 발견되었지만 다른 유르프테리드는 발견되지 않았다.[51]
프테리고투스의 분포는 적어도 3개의 대륙에 걸쳐 있으며, 화석은 종에 따라 유럽, 북아메리카, 남아메리카의 퇴적층에서 발견되고 있다.[74][67] 본 속의 종일 가능성이 있는 ?''Pterygotus australis''는 오스트레일리아에서 발견되었다.[74][67]
5. 분류
프테리고투스(''Pterygotus'')는 프테리고티오이데아(Pterygotioidea) 상과 프테리고투스과(Pterygotidae)에 속하며, 과와 상과 모두에 이름을 부여했다.[16] 가장 분화된 세 종의 프테리고투스과 유립테리드인 아쿠티라무스(Acutiramus), 자켈로프테루스(Jaekelopterus), 프테리고투스(''Pterygotus'')는 서로 매우 유사하며, 특히 ''자켈로프테루스''(Jaekelopterus)와는 생식 부속기와 미절의 특징에서만 사실상 구별된다.[33]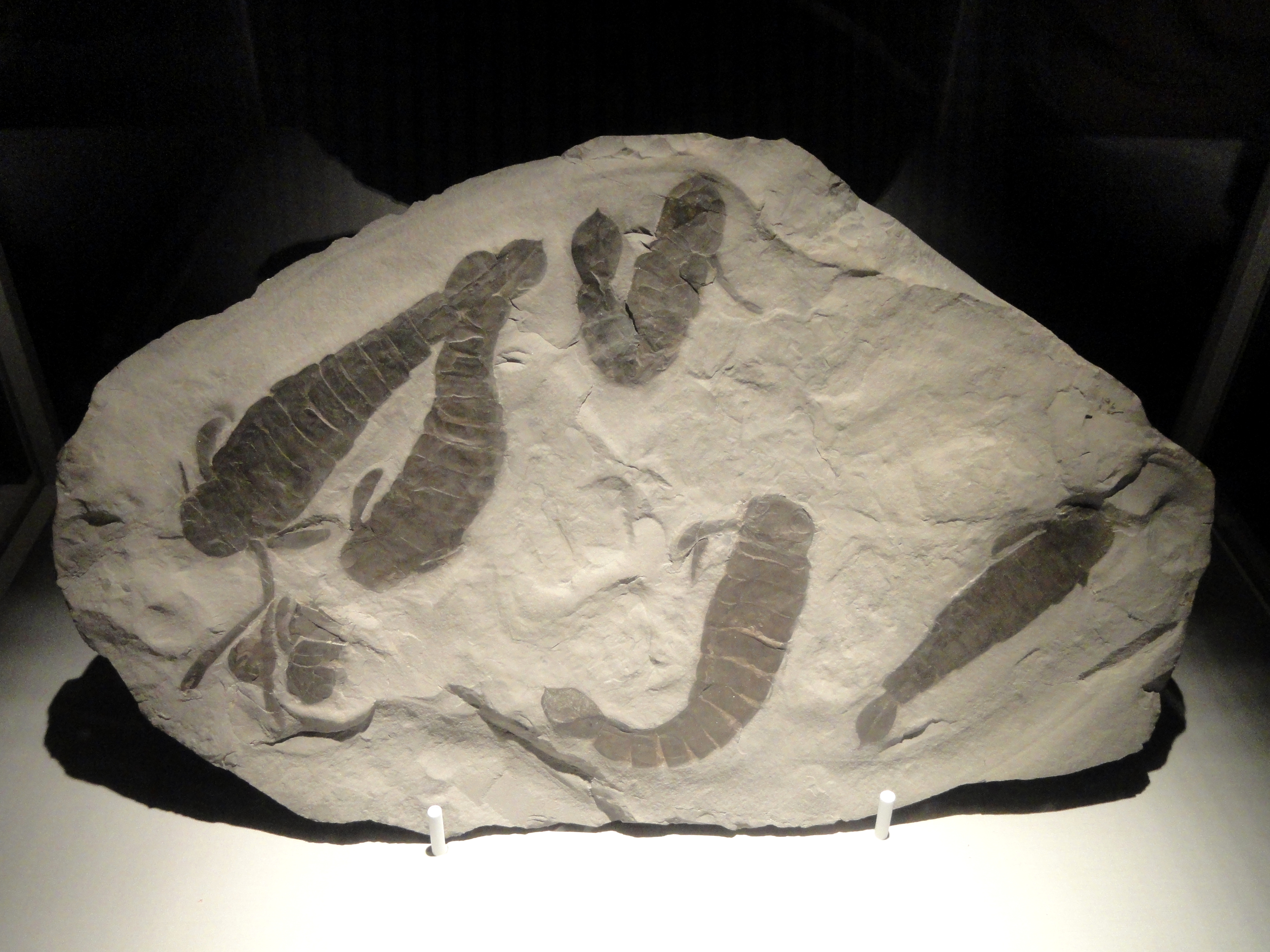
생식 부속기의 유사성은 세 속이 과거에 분류되었던 것처럼 서로 동의어일 수 있음을 시사한다. 집게발의 형태는 생활 방식에 따라 달라지고 개체 발생 전반에 걸쳐 변화하는 것으로 관찰되었기 때문에 속을 구분하는 기준으로 의문이 제기되었다. 미절 형태는 때때로 구별 특징으로 사용되지만, 세 속의 미절은 모두 노 모양이다(''자켈로프테루스''(Jaekelopterus)의 미절은 삼각형이지만, 다른 속의 형태적 범위에 속할 수 있다).[34] 이러한 속들이 서로 동의어인지 여부를 확인하기 위해서는 ''아쿠티라무스''(Acutiramus), ''프테리고투스''(Pterygotus), ''자켈로프테루스''(Jaekelopterus)의 여러 종을 포함하는 포괄적인 계통 발생 분석이 필요하다.[34]
프테리고투스(''Pterygotus'')는 프테리고투스과(Pterygotidae)[68] 중에서 처음으로 발견·기재된 모식 속이다. 시울코프테루스(Ciurcopterus)와 에레토프테루스(Erettopterus)보다 파생적이며, 자켈로프테루스(Jaekelopterus)와 아쿠티라무스(Acutiramus)보다 먼저 분기된 것으로 여겨진다.[63][69][60][70] 20세기 중반까지는 본 과의 모든 종이 본 속에만 분류되었으며, 그 중에서도 에레토프테루스속과 아쿠티라무스속은 각각 "''Pterygotus'' (''Erettopterus'')"와 "''Pterygotus'' (''Acutiramus'')"[55][72]로 본 속의 아속으로 취급되기도 했다.[77]
프테리고투스는 미절이 둥글고 후단이 뾰족하며, 협각은 끝이 매끄럽게 굽어 부동지가 현저하게 특화된 이빨을 결여하는 것으로 동과의 다른 속과 구분된다.[57][63] 그러나 이 분류 체계는 일부 문헌에서 의문시되고 있으며, 이 기준으로는 자켈로프테루스와 아쿠티라무스의 종류도 본 속에 포함될 수 있다.[73] 본 속 중에서 계통 분석이 이루어질 정도로 발견과 연구가 진행된 것은 모식 종인 ''P. anglicus'' 뿐이다.[63][69][70]
2020년 현재, 불확실한 종을 포함하여 18종이 알려져 있으며[74], 최대 체장은 Lamsdell & Braddy (2009)에 근거한다.[69]
- ''Pterygotus anglicus'' Agassiz, 1849(데본기) - 160cm
- 본 속의 모식 종. 협각의 가동지·부동지 모두 끝이 굽어 수직에서 비스듬히 기울어져 있으며, 집게를 닫은 상태에서 가동지의 끝은 부동지의 끝에 의해 둘러싸인다. 가동지의 이빨은 뒤쪽으로 기울어져 있고, 부동지의 이빨은 수직이거나 약간 뒤쪽으로 기울어져 있다. 이빨의 표면은 줄무늬가 있으며, 부동지의 최대 이빨은 가동지의 최대 이빨의 앞쪽 가장자리에 맞물리도록 기울어지는 경우가 있다[57][58] .
- = ''Pterygotus atlanticus'' Clarke & Ruedemann, 1912
- = ''Pterygotus minor'' Woodward, 1864
- ''Pterygotus arcuatus'' Salter, 1859(실루리아기) - 60cm
- ?''Pterygotus australis'' McCoy, 1899(실루리아기) - 체장 불명
- ''Pterygotus barrandei'' Semper, 1898(실루리아기) - 126cm(협각의 단편에 의한 추정)
- = ''Pterygotus beraunensis'' Semper, 1898
- ''Pterygotus bolivianus'' Kjellesvig-Waering, 1964(데본기) - 55cm
- ''Pterygotus carmani'' Kjellesvig-Waering, 1961(데본기) - 체장 불명
- ''Pterygotus cobbi'' Hall, 1859(실루리아기) - 140cm
- ''Pterygotus denticulatus'' Kjellesvig-Waering, 1961(실루리아기) - 120cm
- ''Pterygotus floridanus'' Kjellesvig-Waering, 1950(데본기) - 90cm
- ''Pterygotus gaspesiensis'' Russell, 1953(데본기) - 체장 불명
- ?''Pterygotus grandidentatus'' Kjellesvig-Waering, 1961(실루리아기) - 175cm(협각의 단편에 의한 추정)
- ?''Pterygotus impacatus'' Kjellesvig-Waering, 1964(실루리아기) - 165cm(협각의 단편에 의한 추정)
- ''Pterygotus kopaninensis'' Barrande, 1872(실루리아기) - 50cm(협각의 단편에 의한 추정)
- ''Pterygotus lanarkensis'' Kjellesvig-Waering, 1964(실루리아기)
- ''Pterygotus lightbodyi'' Kjellesvig-Waering, 1961(실루리아기) - 75cm
- ''Pterygotus ludensis'' Salter, 1859(실루리아기) - 체장 불명
- ''Pterygotus marylandicus'' Kjellesvig-Waering, 1964(실루리아기)
- ''Pterygotus monroensis'' Sarle, 1902(실루리아기)
과거에는 본 속에 분류되었으며, 2020년 현재 별개의 속의 종, 그리고 그 동물 동의어가 된 무효의 학명은 다음과 같다.[74]
- 아크티라무스아속 '''''Pterygotus'' (''Acutiramus'')''' Ruedemann 1935 → 아크티라무스속 '''''Acutiramus''''' Ruedemann 1935
- *Størmer (1974)에 의한 승격[75]。
- *'''''Pterygotus bohemicus''''' Barrande, 1872 → 아크티라무스속: '''''Acutiramus영어 bohemicus''''' (Barrande, 1872)(실루리아기)
- **Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus comes'' Barrande, 1872
- ** ''= Pterygotus mediocris'' Barrande, 1872
- ** ''= Pterygotus blahai'' Semper, 1898
- ** ''= Pterygotus fissus'' Seemann, 1906
- * '''''Pterygotus cummingsi''''' Grote & Pitt, 1875 → 아크티라무스속: '''''Acutiramus cummingsi''''' (Grote & Pitt, 1875)(실루리아기)
- ** Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus acuticaudatus'' Pohlman, 1882
- ** ''= Pterygotus buffaloensis'' Pohlman, 1881
- ** ''= Pterygotus quadraticaudatus'' Pohlman, 1882
- * '''''Pterygotus macrophthalmus''''' Hall, 1859 → 아크티라무스속: '''''Acutiramus macrophthalmus''''' (Hall, 1859)(실루리아기)
- ** Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus osborni'' Hall, 1859
- ** ''= Pterygotus cobbi'' var. ''juvenis'' Clarke & Ruedemann, 1912
- 에레토프테루스아속 '''''Pterygotus'' (''Erettopterus영어'')''' Salter 1859 → 에레토프테루스속 '''''Erettopterus''''' Salter 1859
- *Kjellesvig-Waering (1961)에 의한 승격[71]。
- *'''''Pterygotus bilobus''''' Salter, 1856 → 에레토프테루스속: '''''Erettopterus bilobus''''' (Salter, 1856)(실루리아기)
- ** Kjellesvig-Waering (1961)에 의한 재분류[71]。
- **= ''Eurypterus perornatus'' Salter, 1856
- ** = ''Pterygotus bilobus'' var. ''acidens'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''crassus'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''inornatus'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''perornatus'' Woodward, 1878
- ** = ''Pterygotus perornatus'' var. ''plicatissimus'' Salter in Huxley & Salter, 1859
- *'''''Pterygotus canadensis''''' Dawson, 1879 → 에레토프테루스속: '''''Erettopterus canadensis''''' (Dawson, 1879)(실루리아기)
- *'''''Pterygotus holmi''''' Størmer, 1934b → 에레토프테루스속: '''''Erettopterus holmi''''' (Størmer, 1934b)(실루리아기)
- '''''Pterygotus howelli''''' Salter, 1856 → 자켈로프테루스속: '''''Jaekelopterus영어 howelli''''' (Salter, 1856)(데본기)
- * Tetlie (2007)에 의한 재분류[76]。
- *= ''Pterygotus mcgrewi'' Kjellesvig-Waering & Richardson In Kjellesvig-Waering (1986)
- '''''Pterygotus rhenaniae''''' Jaekel, 1914 → 자켈로프테루스속: '''''Jaekelopterus rhenaniae''''' (Jaekel, 1914)(데본기)
- *Waterston (1964)에 의한 재분류[72]。
- '''''Pterygotus''? ''sarlei''''' Ciurca & Tetlie, 2007[77] → 치우르코프테루스속: '''''Ciurcopterus영어 sarlei''''' (Ciurca & Tetlie, 2007)(실루리아기)
- *Tetlie & Briggs (2009)에 의한 재분류[69]。
- '''''Pterygotus ventricosus''''' Kjellesvig-Waering, 1948a[78] → 치우르코프테루스속: '''''Ciurcopterus ventricosus''''' (Kjellesvig-Waering, 1948a)(실루리아기)
- *Tetlie & Briggs (2009)에 의한 재분류[69]。
5. 1. 하위 종
프테리고투스(Pterygotus)는 프테리고티오이데아(Pterygotioidea) 상과 프테리고투스과(Pterygotidae)에 속하며, 과와 상과 모두에 이름을 부여했다.[16] 가장 분화된 세 종의 프테리고투스과 유립테리드인 아쿠티라무스(Acutiramus), 자켈로프테루스(Jaekelopterus), 프테리고투스(Pterygotus)는 서로 매우 유사하며, 특히 ''자켈로프테루스''(Jaekelopterus)와는 생식 부속기와 텔슨(미부)의 특징에서만 사실상 구별된다.[33]생식 부속기의 유사성은 세 속이 과거에 분류되었던 것처럼 서로 동의어일 수 있음을 시사한다. 집게발의 형태는 생활 방식에 따라 달라지고 개체 발생 전반에 걸쳐 변화하는 것으로 관찰되었기 때문에 속을 구분하는 기준으로 의문이 제기되었다. 텔슨 형태는 때때로 구별 특징으로 사용되지만, 세 속의 텔슨은 모두 노 모양이다(''Jaekelopterus''의 텔슨은 삼각형이지만, 다른 속의 형태적 범위에 속할 수 있다).[34] 이러한 속들이 서로 동의어인지 여부를 확인하기 위해서는 ''Acutiramus'', ''Pterygotus'', ''Jaekelopterus''의 여러 종을 포함하는 포괄적인 계통 발생 분석이 필요하다.[34]
프테리고투스(Pterygotus)는 바다전갈과(Pterygotidae)[68](Pterygotidae) 중에서 처음으로 발견·기재된 모식 속이다. 시울코프테루스(Ciurcopterus)와 에레토프테루스(Erettopterus)보다 파생적이며, 제켈로프테루스(Jaekelopterus)와 아쿠티라무스(Acutiramus)보다 먼저 분기된 것으로 여겨진다.[63][69][60][70] 20세기 중반까지는 본 과의 모든 종이 본 속에만 분류되었으며, 그 중에서도 에레토프테루스속과 아쿠티라무스속은 각각 "''Pterygotus'' (''Erettopterus'')"와 "''Pterygotus'' (''Acutiramus'')"[55][72]로 본 속의 아속으로 취급되기도 했다.[77]
프테리고투스는 미절이 둥글고 후단이 뾰족하며, 협각은 끝이 매끄럽게 굽어 부동지가 현저하게 특화된 이빨을 결여하는 것으로 동과의 다른 속과 구분된다.[57][63] 그러나 이 분류 체계는 일부 문헌에서 의문시되고 있으며, 이 기준으로는 제켈로프테루스와 아쿠티라무스의 종류도 본 속에 포함될 수 있다.[73] 본 속 중에서 계통 분석이 이루어질 정도로 발견과 연구가 진행된 것은 모식 종인 ''P. anglicus'' 뿐이다.[63][69][70]
2020년 현재, 불확실한 종을 포함하여 18종이 알려져 있으며[74], 최대 체장은 Lamsdell & Braddy (2009)에 근거한다.[69]
- ''Pterygotus anglicus'' Agassiz, 1849(데본기) - 160cm
- 본 속의 모식 종. 협각의 가동지·부동지 모두 끝이 굽어 수직에서 비스듬히 기울어져 있으며, 집게를 닫은 상태에서 가동지의 끝은 부동지의 끝에 의해 둘러싸인다. 가동지의 이빨은 뒤쪽으로 기울어져 있고, 부동지의 이빨은 수직이거나 약간 뒤쪽으로 기울어져 있다. 이빨의 표면은 줄무늬가 있으며, 부동지의 최대 이빨은 가동지의 최대 이빨의 앞쪽 가장자리에 맞물리도록 기울어지는 경우가 있다[57][58] .
- = ''Pterygotus atlanticus'' Clarke & Ruedemann, 1912
- = ''Pterygotus minor'' Woodward, 1864
- ''Pterygotus arcuatus'' Salter, 1859(실루리아기) - 60cm
- ?''Pterygotus australis'' McCoy, 1899(실루리아기) - 체장 불명
- ''Pterygotus barrandei'' Semper, 1898(실루리아기) - 126cm(협각의 단편에 의한 추정)
- = ''Pterygotus beraunensis'' Semper, 1898
- ''Pterygotus bolivianus'' Kjellesvig-Waering, 1964(데본기) - 55cm
- ''Pterygotus carmani'' Kjellesvig-Waering, 1961(데본기) - 체장 불명
- ''Pterygotus cobbi'' Hall, 1859(실루리아기) - 140cm
- ''Pterygotus denticulatus'' Kjellesvig-Waering, 1961(실루리아기) - 120cm
- ''Pterygotus floridanus'' Kjellesvig-Waering, 1950(데본기) - 90cm
- ''Pterygotus gaspesiensis'' Russell, 1953(데본기) - 체장 불명
- ?''Pterygotus grandidentatus'' Kjellesvig-Waering, 1961(실루리아기) - 175cm(협각의 단편에 의한 추정)
- ?''Pterygotus impacatus'' Kjellesvig-Waering, 1964(실루리아기) - 165cm(협각의 단편에 의한 추정)
- ''Pterygotus kopaninensis'' Barrande, 1872(실루리아기) - 50cm(협각의 단편에 의한 추정)
- ''Pterygotus lanarkensis'' Kjellesvig-Waering, 1964(실루리아기)
- ''Pterygotus lightbodyi'' Kjellesvig-Waering, 1961(실루리아기) - 75cm
- ''Pterygotus ludensis'' Salter, 1859(실루리아기) - 체장 불명
- ''Pterygotus marylandicus'' Kjellesvig-Waering, 1964(실루리아기)
- ''Pterygotus monroensis'' Sarle, 1902(실루리아기)
과거에는 본 속에 분류되었으며, 2020년 현재 별개의 속의 종, 그리고 그 동물 동의어가 된 무효의 학명은 다음과 같다.[74]
- 아크티라무스아속 '''''Pterygotus'' (''Acutiramus'')''' Ruedemann 1935 → 아크티라무스속 '''''Acutiramus''''' Ruedemann 1935
- *Størmer (1974)에 의한 승격[75]。
- *'''''Pterygotus bohemicus''''' Barrande, 1872 → 아크티라무스속: '''''Acutiramus bohemicus''''' (Barrande, 1872)(실루리아기)
- **Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus comes'' Barrande, 1872
- ** ''= Pterygotus mediocris'' Barrande, 1872
- ** ''= Pterygotus blahai'' Semper, 1898
- ** ''= Pterygotus fissus'' Seemann, 1906
- * '''''Pterygotus cummingsi''''' Grote & Pitt, 1875 → 아크티라무스속: '''''Acutiramus cummingsi''''' (Grote & Pitt, 1875)(실루리아기)
- ** Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus acuticaudatus'' Pohlman, 1882
- ** ''= Pterygotus buffaloensis'' Pohlman, 1881
- ** ''= Pterygotus quadraticaudatus'' Pohlman, 1882
- * '''''Pterygotus macrophthalmus''''' Hall, 1859 → 아크티라무스속: '''''Acutiramus macrophthalmus''''' (Hall, 1859)(실루리아기)
- ** Størmer (1974)에 의한 재분류[75]。
- **''= Pterygotus osborni'' Hall, 1859
- ** ''= Pterygotus cobbi'' var. ''juvenis'' Clarke & Ruedemann, 1912
- 에레토프테루스아속 '''''Pterygotus'' (''Erettopterus'')''' Salter 1859 → 에레토프테루스속 '''''Erettopterus''''' Salter 1859
- *Kjellesvig-Waering (1961)에 의한 승격[71]。
- *'''''Pterygotus bilobus''''' Salter, 1856 → 에레토프테루스속: '''''Erettopterus bilobus''''' (Salter, 1856)(실루리아기)
- ** Kjellesvig-Waering (1961)에 의한 재분류[71]。
- **= ''Eurypterus perornatus'' Salter, 1856
- ** = ''Pterygotus bilobus'' var. ''acidens'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''crassus'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''inornatus'' Woodward, 1878
- ** = ''Pterygotus bilobus'' var. ''perornatus'' Woodward, 1878
- ** = ''Pterygotus perornatus'' var. ''plicatissimus'' Salter in Huxley & Salter, 1859
- *'''''Pterygotus canadensis''''' Dawson, 1879 → 에레토프테루스속: '''''Erettopterus canadensis''''' (Dawson, 1879)(실루리아기)
- *'''''Pterygotus holmi''''' Størmer, 1934b → 에레토프테루스속: '''''Erettopterus holmi''''' (Størmer, 1934b)(실루리아기)
- '''''Pterygotus howelli''''' Salter, 1856 → 야켈로프테루스속: '''''Jaekelopterus howelli''''' (Salter, 1856)(데본기)
- * Tetlie (2007)에 의한 재분류[76]。
- *= ''Pterygotus mcgrewi'' Kjellesvig-Waering & Richardson In Kjellesvig-Waering (1986)
- '''''Pterygotus rhenaniae''''' Jaekel, 1914 → 야켈로프테루스속: '''''Jaekelopterus rhenaniae''''' (Jaekel, 1914)(데본기)
- *Waterston (1964)에 의한 재분류[72]。
- '''''Pterygotus''? ''sarlei''''' Ciurca & Tetlie, 2007[77] → 치우르코프테루스속: '''''Ciurcopterus sarlei''''' (Ciurca & Tetlie, 2007)(실루리아기)
- *Tetlie & Briggs (2009)에 의한 재분류[69]。
- '''''Pterygotus ventricosus''''' Kjellesvig-Waering, 1948a[78] → 치우르코프테루스속: '''''Ciurcopterus ventricosus''''' (Kjellesvig-Waering, 1948a)(실루리아기)
- *Tetlie & Briggs (2009)에 의한 재분류[69]。
6. 연구사
''프테리고투스''의 첫 번째 화석은 로흐코비안-프라기안 (초기 데본기) 시대의 퇴적층에서 스코틀랜드와 잉글랜드 서부의 채석공에 의해 발견되었으며, 그들은 큰 화석 유해를 "세라핌"이라고 불렀다.[10] 루이 아가시는 스위스계 미국인 생물학자이자 지질학자로, 1839년에 화석을 묘사하고 이를 "날개 달린 것"을 의미하는 ''프테리고투스''(Pterygotus)로 명명했다. 아가시는 그것들이 큰 물고기의 화석 유해라고 믿었다.[10] 아가시가 잉글랜드에서 묘사한 표본은 그가 ''프테리고투스 프로블레마티쿠스''(Pterygotus problematicus)라고 명명한 종으로 분류되었다. 아가시는 5년 후인 1844년 스코틀랜드의 올드 레드 사암에서 더 완전한 화석을 검토한 후, 화석의 진정한 본질이 절지동물 유해임을 처음으로 인식했다.[11][14]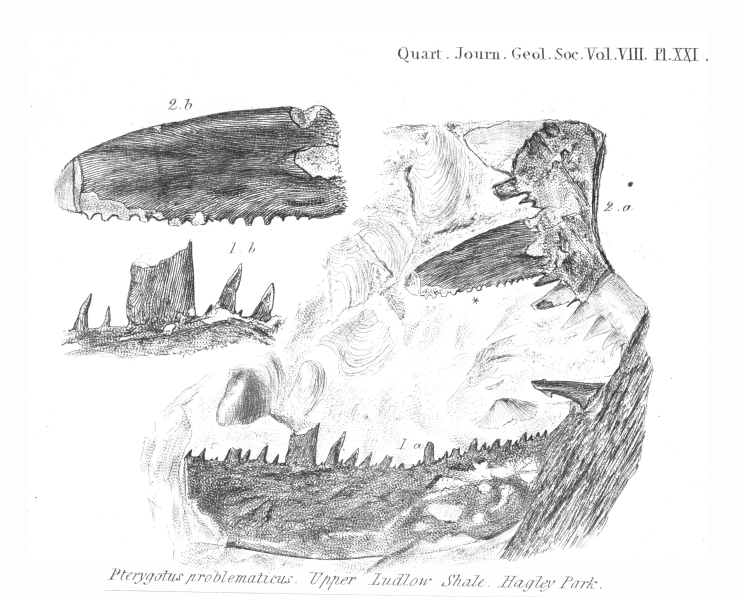
비록 ''프테리고투스''의 화석을 절지동물로 인식한 것이 현대의 계통 발생학적 위치에 더 가까웠지만, 아가시는 ''프테리고투스''가 Entomostraca 아강의 갑각류를 나타내는 것으로 간주했다. 비록 프레데릭 맥코이가 1849년에 ''프테리고투스''가 말발굽게과 이전에 발견된 유라프테리드 ''유리프테루스''와 유사하다는 점을 언급했지만, 그는 ''유리프테루스''와 ''프테리고투스''를 모두 갑각류로 분류했다.[12]
새로운 스코틀랜드 화석은 1849년에 ''P. 앵글리쿠스''(P. anglicus)라는 종으로 명명되었으며, 이는 ''프테리고투스''의 가장 널리 알려진 종으로 남아 있으며, 이후 발견된 종들과는 구부러진 말단 이빨과 1차 및 중간 이빨이 약간 뒤로 기울어져 있다는 점에서 구별된다.[13]
''P. 프로블레마티쿠스''는 또한 1852년 존 윌리엄 샐터가 잉글랜드 서부의 웨일스 변경 지역에서 발견한 불완전한 협각을 지칭하는 데 사용되었지만, 현대에는 종을 정의하는 것이 불가능하므로 ''무효명''( "근거 없는 이름")으로 간주된다. 그것이 묘사된 화석 자료는 진단할 수 없으며 의미 있는 특성을 확립하기에 불충분하며, 그 때문에 많은 단편적인 프테리고투스과 화석이 그 긴 역사 동안 그것으로 언급되었으며, 이는 효과적으로 프테리고투스과와 동의어가 되었다. 대부분의 경우, 이러한 조각들은 프테리고투스과 외골격의 조각으로 구성되어 있으며, 연구자들이 오랫동안 단지 ''프테리고투스'' 또는 ''P. 프로블레마티쿠스''의 특징이라고 잘못 믿었던 비늘 모양의 장식을 보존하고 있다. 이러한 장식은 모든 프테리고투스과 속에서 알려져 있으므로 단일 종의 진단적 특징으로 사용할 수 없다.[14] 비록 ''P. 프로블레마티쿠스''가 ''프테리고투스'' 종에 사용된 가장 초기의 이름이지만, 그 이름이 더 이상 사용되지 않으므로 기준 종으로 간주되지 않는다. 대신, 1844년에 아가시가 묘사한 것처럼 여러 진단적 특징을 기반으로 하고 적절하게 삽화된 ''P. 앵글리쿠스''가 ''프테리고투스''의 기준 종으로 간주된다.[14]
19세기 동안 잉글랜드에서 오늘날까지 속으로 지정된 두 개의 종이 더 묘사되었으며, 프리돌리 (후기 실루리아기) 시대의 ''P. 루덴시스''(P. ludensis)와 러들로 (후기 실루리아기) 시대의 ''P. 아르쿠아투스''(P. arcuatus), 그리고 미국의 프리돌리에서 발견된 북미 종인 ''P. 코비''(P. cobbi)가 있다. ''P. 아르쿠아투스''는 원래 존 윌리엄 샐터에 의해 5개의 개별 화석 표본을 포함하는 합성형 계열(특정 모식표본이 지정되지 않은 일련의 표본)의 화석 유해로 묘사되었다. 1961년, 묘사된 지 102년 후, 에릭 N. 켈레스비그-와에링은 이러한 표본 중 하나(런던 지질 조사 및 박물관의 89587번, 협각의 자유 가지)만이 실제로 이 독특한 종에 해당할 수 있으며, 다른 화석(등판, 콕사 및 결정할 수 없는 조각)은 진단적이지 않기 때문에 다른 종의 화석을 나타낼 수 있다고 언급했다.[14] 1859년 샐터가 묘사한 ''P. 루덴시스''는 텔손의 시작부터 꼬리 가시까지 이어지는 더 발달되고 연장된 용골(또는 능선)에 의해 다른 종과 구별될 수 있다.[15]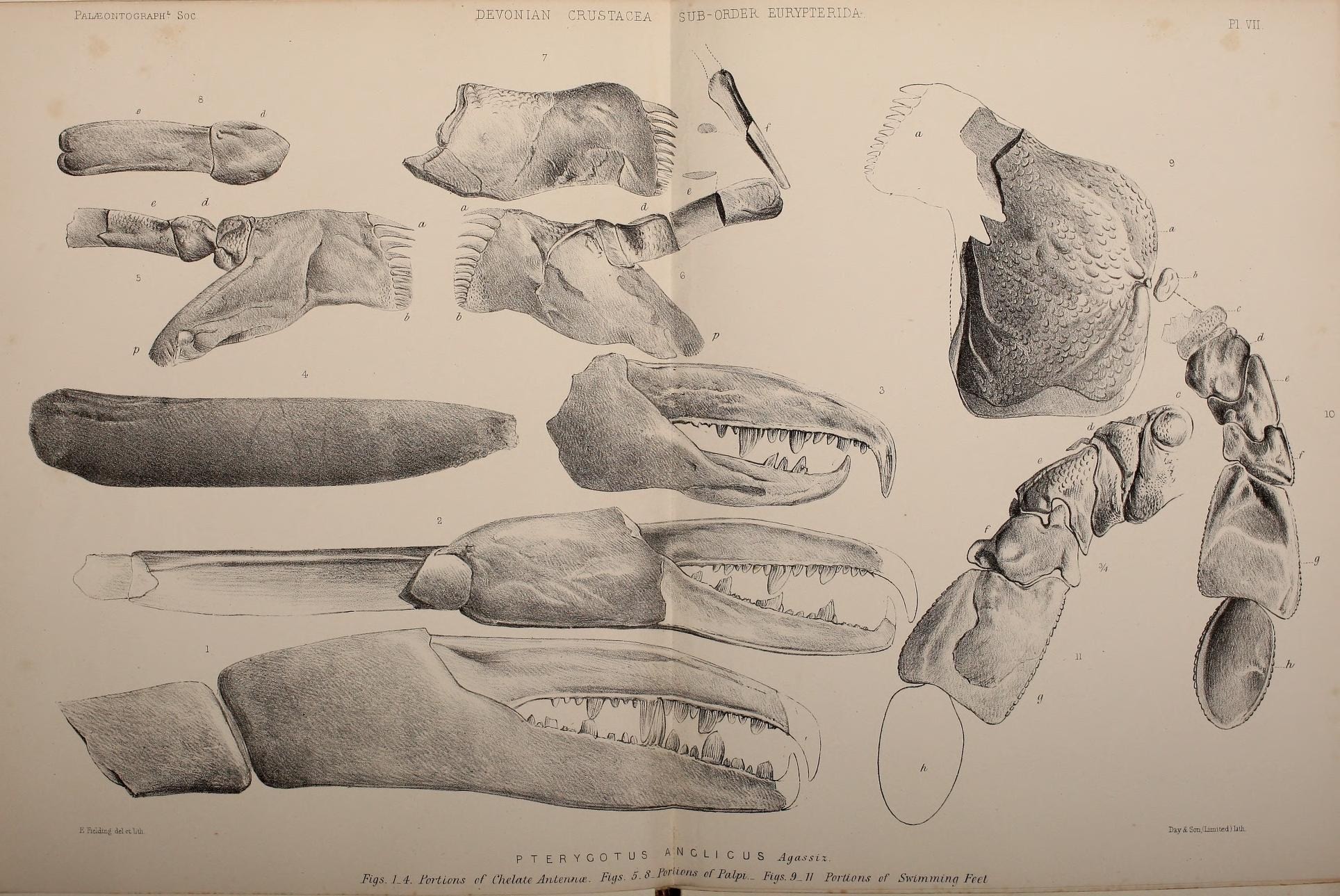
1859년 제임스 홀이 미국 뉴욕에서 발견된 화석을 기반으로 묘사한 희귀종 ''P. 코비''는 스코틀랜드 외부에서 묘사된 최초의 ''프테리고투스'' 종이었으며, 속의 알려진 범위를 상당히 확장시켰다. 홀은 이 새로운 종과 함께 두 개의 다른 북미 종을 묘사했다. 즉, ''P. 매크로프탈무스''(현재 ''아큐티라무스''로 분류됨)와 ''P. 오스보르니''(나중에 동의어가 ''P. 매크로프탈무스''와 됨). 자유 가지(움직이는 발톱 부분)의 원위 치아는 다른 종보다 덜 두드러졌으며, 이는 ''아큐티라무스 커밍시''의 자유 가지의 원위 치아와 유사하다는 점에 주목했다. 비록 ''P. 코비''가 열악한 화석 자료에 기반하고, 자유 가지에서만 알려져 있지만, 체코에서 발견된 특정 종(예: ''P. 바라데이'')과 다른 프테리고투스과 종(예: ''아큐티라무스 매크로프탈무스'')보다 유사하다는 점을 고려하여 여전히 별개의 종으로 인식된다.[6]
1859년까지 10종이 이 속에 할당되었고,[16] 존 윌리엄 살터는 텔슨의 형태에 따라 이 종들을 아속으로 나눌 수 있음을 인식했다. 살터는 이엽 텔슨을 가진 종을 위해 아속 ''프테리고투스'' (''에레토프테루스'')를 세웠다.[17] 더 많은 차이점이 종들 사이에서 발견되면서 추가적인 아속이 명명되었으며, 이러한 아속에는 협각의 소치(이빨) 특징을 기반으로 1935년에 명명된 ''프테리고투스'' (''Curviramus'')와 ''프테리고투스'' (''아쿠티라무스'')가 포함된다.[17] ''프테리고투스'' (''Curviramus'')는 나중에 레프 스퇴르머에 의해 같은 해에 ''프테리고투스'' (''프테리고투스'')와 동의어로 인정되었고,[11] ''에레토프테루스''와 ''아쿠티라무스''는 별개의 속이지만 밀접한 관련이 있는 속으로 인정되었다(1961년 에릭 N. 켈레스비그-웨링에 의한 ''에레토프테루스'',[11] 1974년 스퇴르머에 의한 ''아쿠티라무스'').[17]
1912년, 존 메이슨 클라크와 루돌프 루드먼은 유절류 속 ''프테리고투스'', ''슬리모니아'', ''휴밀레리아'' 및 ''하스티미마''를 포함하기 위해 1912년에 프테리고투스과(Pterygotidae)를 세웠다. 후자의 세 속은 1951년 에릭 N. 켈레스비그-웨링에 의해 휴밀레리아과(Hughmilleriidae)의 구성원으로 재분류되어, ''프테리고투스''와 이전 아속이 유일한 프테리고투스과 유절류로 남게 되었다.[9]
초기의 ''프테리고투스'' 발견이 잉글랜드와 북아메리카에 국한되었지만, 20세기 동안의 화석 발견은 ''프테리고투스''가 거의 전 세계적 분포를 이루었다는 것을 입증하는 데 도움이 되었다.[18] 호주에서 처음 발견된 유르테리드는 ''Pterygotus australis''였으며, 그 화석은 다길 형성의 루드로우기 멜버른 그룹에서 발견되었다. 이 화석은 1899년 멜버른 아래 배수 시설 공사 중 발굴 과정에서 발견된 네 개의 조각으로 구성되어 있으며, 세그먼트의 약 절반을 차지한다. 이 단편적인 화석은 당시 ''프테리고투스'' 종으로 분류되었던 ''Erettopterus bilobus''의 화석과 매우 유사하여, ''프테리고투스''로의 분류에 의문을 제기할 수 있다.[19] 2020년, 이 종은 pterygotid의 나머지 종과 ''P. australis''를 구분할 충분한 진단 자료가 부족하여 ''nomen dubium''(의심스러운 종)으로 표시되었다.[20]
Kjellesvig-Waering은 1964년 볼리비아의 엠시안-아이펠리안(초기~중기 데본기) 시대 퇴적물에서 회수된 화석을 바탕으로 ''P. bolivianus'' 종을 명명했다. 이 종은 남아메리카에서 처음 발견된 프테리고티드이며, 남반구의 퇴적물에서 회수된 최초의 데본기 프테리고티드이며, 또한 프테리고티드과의 마지막 생존 개체 중 하나를 나타낸다. 표본(PE6173, 시카고 자연사 박물관 소장)에는 잘 보존된 협각과 가지의 앞부분 절반이 포함되어 있다. 가지의 이빨은 짧고 넓으며 원뿔형이며, 모두 약간 뒤로 구부러져 있다. 말단 치아는 더 크지만, 그 바로 뒤에 있는 치아보다 약간 크며, 발톱의 안쪽 굽힘은 ''P. lightbodyi'', ''P. impacatus'' 및 ''Erettopterus brodiei''와 같은 다른 종에서도 나타나는 일종의 "이중 치아 조합"이 존재할 수 있음을 시사한다. 화석화된 가지의 총 길이는 2.35cm(1인치)이지만, 전체 가지의 약 절반만을 나타낼 가능성이 높다. 다른 종(그리고 일반적으로 프테리고투스과)과 마찬가지로, 이빨은 미세하게 갈비뼈 모양이다. 좁지만 짧고 균일한 크기의 치아와 말단 치아가 특별한 발달을 보이지 않는다는 점에서 다른 모든 ''프테리고투스'' 종과 구별할 수 있다.[9] 2019년, 콜롬비아의 쿠체 형성에서 새로운 단편적인 협각의 가지가 발견되었다. 홀로타입과의 유사성으로 인해 불확실하게 ''P. bolivianus''로 지정된 표본(SGC-MGJRG.2018.I.5)은 콜롬비아의 첫 번째 유르테리드이자 남아메리카의 네 번째 유르테리드를 나타낸다. 이 화석은 프라스니안(후기 데본기)으로 연대가 측정되어, ''프테리고투스''가 이전에 생각했던 것처럼 중기 데본기에 멸종되지 않았음을 보여준다.[21]
면밀한 검토와 새로운 화석 증거의 발견에 따라, 더 많은 속이 ''프테리고투스''에서 분리되었다. ''P. rhenaniae''는 1964년 Charles D. Waterston에 의해 자체 속인 ''Jaekelopterus''의 일부로 분류되었다. 그는 이 종이 세분된 생식 부속지(나중에 잘못된 것으로 밝혀진 특징)와 좁고 긴 협각, 그리고 주요 치아가 약간 앞쪽으로 기울어져 있다는 점에서 다른 ''프테리고투스'' 종과 충분히 구별된다고 간주했다.[22] 또 다른 종인 ''P. ventricosus''는 2007년 O. Erik Tetlie와 Derek E. G. Briggs에 의해 별도의 속인 ''Ciurcopterus''로 분류되었으며, 주로 ''Slimonia''의 부속지와 유사한 부속지를 갖는 것과 같이, 더 기저적인 pterygotioid 유르테리드와 몇 가지 특징을 공유하는 것으로 구별되었다.[23]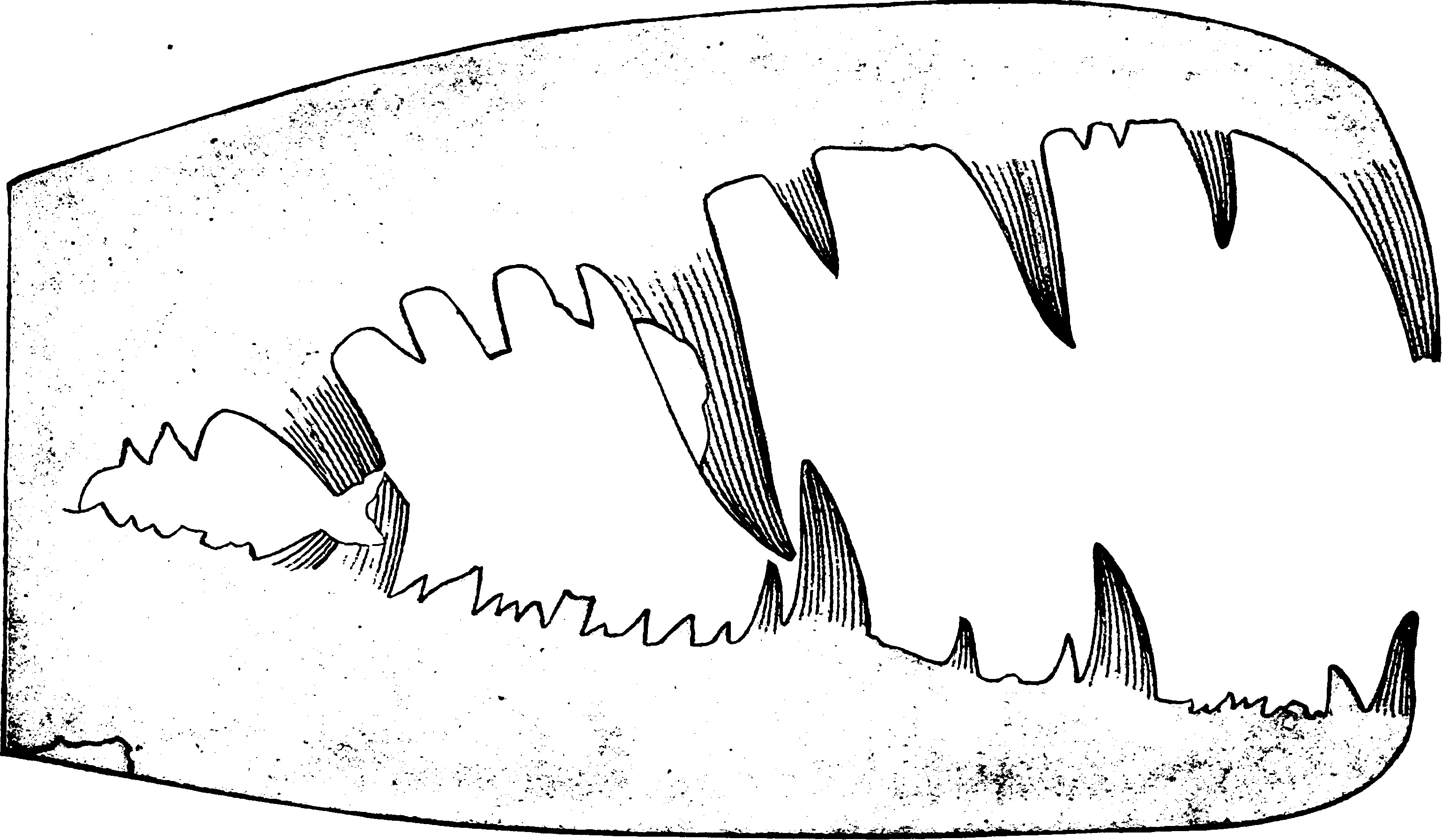
새로운 화석 발견은 이전에 알려지지 않았던 여러 유럽 국가에서 ''프테리고투스''(Pterygotus)의 존재를 밝혀냈고, 분류학적으로 매우 다양한 속임을 입증했다. ''P. barrandei''는 1898년에 명명되었으며, 체코의 프리돌리 시대 퇴적물에서 화석이 발견된다. ''P. barrandei''는 ''P. cobbi''와 매우 유사한 것으로 알려져 있으며, 두 종 사이의 밀접한 관계가 추정된다. 많은 유사점에도 불구하고, 두 종은 몇 가지 차이점이 있는데, 가장 두드러진 것은 자유 지체의 협각 치아이다. ''P. barrandei''의 자유 지체에서 가장 큰 치아는 ''P. cobbi''의 해당 치아보다 훨씬 길고, ''P. barrandei''의 자유 지체의 치아는 일반적으로 더 앞쪽으로 향해 있다. ''P. barrandei''의 화석은 희귀하며, 화석 발견은 보헤미아의 프리돌리 시대 지층 몇 곳에 국한되어 있다. 알려진 화석으로는 일부 불완전한 협각과 배판이 있다. 몇몇 추가 화석 잔해도 이 종에 할당되었는데, 이는 콕사와 생식 부속지를 포함하지만, 이 종에 대한 할당은 의심스럽다.[24]
1872년에 기술된 종인 ''P. nobilis''는 오늘날 체코에서 발견된 작고 조각난 협각을 기반으로 한다. 이 발톱에서 보이는 치아 배열은 대부분의 치아가 보존되지 않았지만, 연구자 막스 셈퍼는 1897년에 ''프테리고투스''의 모식종인 ''P. anglicus''에서 보이는 것과 거의 또는 전혀 유사하지 않으며, 실제로 ''P. buffaloensis''와 ''P. bohemicus''에서 보이는 것과 더 유사하다고 언급했다. 오늘날 ''P. buffaloensis''는 ''P. bohemicus''의 주니어 동의어로 간주되며, 이는 밀접하게 관련된 속인 ''Acutiramus''의 일부로 재분류되었다.[17] ''P. nobilis''의 조각난 잔해는 정확한 정체성에 대한 추가 연구를 어렵게 만들며, 셈퍼는 이것이 ''Acutiramus bohemicus''와 동의어일 수 있다고 제안했지만, "질문[그 정체성에 대한]은 나에게 제공된 자료에서 답변할 수 없다"고 언급했다.[25]
1872년에 명명된 또 다른 종인 ''P. kopaninensis''는 프라하에 위치한 자드니 코파니나 마을 주변의 코파니나 지층에서 발견된 단일하고 불완전한 고정된 협각 가지(표본 번호 L1396)에서 알려져 있다. 이 표본은 길이가 4.3cm(1.7인치)이며, 한때 "가지의 특이한 갈고리 모양 종단" 때문에 ''Erettopterus''에 할당되었는데, 이 특징은 화석에 자유 가지의 잔해가 존재하기 때문에 나타나는 것으로 알려져 있다. 발톱의 치아 패턴과 모양은 ''프테리고투스''에 할당하는 것이 더 타당함을 시사한다.[24]
1926년 엠브릭 스트랜드에 의해 기술된 ''P. siemiradzkii''는 서부 우크라이나의 화석 자료를 기반으로 하며, 진단적 가치가 거의 없는 매우 조각난 자료를 기반으로 한다. ''P. siemiradzkii''로 지정된 단일 표본인 텔슨은 알려진 ''프테리고투스'' 종, 또는 기타 프테리고티드 유르테리드의 속에서는 볼 수 없는 특이하고 두드러진 능선이 있어, ''프테리고투스''로의 할당에 의문이 제기된다.[26]
1964년, Kjellesvig-Waering에 의해 기술된 두 종은 ''프테리고투스''의 알려진 범위를 스코틀랜드(''P. lanarkensis'')와 에스토니아(''P. impacatus'')로 확장시켰으며, 둘 다 루드러우기에 해당한다.[3][9] ''P. lanarkensis''는 스코틀랜드 라나크셔의 레스마하고우에서 발견된 잘 알려진 유르테리드 동물군에서 두 번째로 발견된 프테리고티드였다. 프테리고티드는 여러 관련 속과 연관되어 나타나는 것이 일반적이므로, 레스마하고우에 단일 종인 ''Erettopterus bilobus''만 존재한다는 것은 이례적인 일로 간주되었다. ''P. lanarkensis''의 화석 잔해는 1868년부터 알려져 있었지만(1855–1860년에 로버트 슬리몬이 처음 수집), 1964년 Kjellesvig-Waering에 의해 처음으로 인식되었다. 원래 ''Erettopterus bilobus''로 의심스럽게 언급된 큰 협각(표본 번호는 영국 자연사 박물관의 53890)에 의해 대표되는 ''P. lanarkensis''는 ''Erettopterus'' 종보다 ''P. anglicus''와 더 밀접하게 유사하다. (이 표본에서 파손된) 말단 치아는 길이가 0.9cm이고, 중앙 치아는 이례적으로 짧다. 이례적으로 긴 말단 치아와 발달하지 않은 중앙 치아의 조합은 ''P. lanarkensis''를 다른 ''프테리고투스'' 종과 구별한다. ''P. lanarkensis''가 발견된 ''Erettopterus''와 구별되는 몇 가지 특징이 있으며, 여기에는 ''P. lanarkensis''의 더 견고한 협각이 포함된다.[9]
에스토니아 사레마의 키엘콘드에서 루드러우 시대의 퇴적물에서 발견된 ''P. impacatus''는 진단적이고 잘 보존된 세부 사항을 보존하는 협각의 조각난 자유 가지로 구성된 홀로타입 표본(표본 번호 7059/7, 비교 동물학 박물관에 소장)으로 대표된다. 이 표본과 함께, 이 종에 언급된 중요한 표본에는 파라타입(No. 7059/3, 조각난 협각)이 포함된다. 이 표본은 발톱의 말단 부분의 두껍고 긴 치아에 이어지는 수직 치아와 같이 ''P. impacatus''를 진단하는 여러 특징을 포함한다. 특히 진단적 가치가 있는 것은 말단 치아가 처음 시작되는 지점에 치아가 존재한다는 것이다. 자유 가지의 중앙 치아는 이 종에서 이례적으로 크다. 말단 치아는 길이가 2.3cm(1인치)이다. ''P. impacatus''의 가지는 크고 뾰족한 소포(피부의 융기)로 장식되어 있으며, 이 특징은 ''P. impacatus''의 표본을 그 잔해가 발견되는 화석 부지의 다른 프테리고티드와 구별하는 데 도움이 된다.[9] 2007년, O. 에릭 테틀리에 의해 ''P. impacatus''를 ''프테리고투스''에 할당하는 것에 의문이 제기되었는데, 눈과 등갑의 모양이 ''Erettopterus''에서 이들 신체 부위의 모양과 유사했기 때문이다.[17]

20세기에는 북아메리카에서도 실루리아기의 ''P. marylandicus''(1964년, 메릴랜드, 미국)와 ''P. monroensis''(1902년, 뉴욕, 미국), 초기 데본기의 ''P. carmani''(1961년, 미국)와 ''P. floridanus''(1950년, 플로리다, 미국), 중기 데본기의 ''P. gaspesiensis''(1953년, 퀘벡, 캐나다) 등 ''프테리고투스''의 추가적인 종들이 발견되어 기술되었다.[3][27]
프테리고투스 유해류의 화석 유물은 이 그룹에서 알려진 뚜렷한 비늘 모양 장식을 나타내며, 연구자 윌리엄 에드먼드 로건이 1846년 이전에 캐나다 동부에서 보고했는데, 그는 "머치슨의 ''Pterygotus problematicus''와 강한 유사성을 보이는" 동물의 발생을 개스페, 퀘벡의 실루리아기-데본기 퇴적층에서 보고했다. 결국 데본기 시대의 것으로 밝혀진 화석은 처음에는 ''P. atlanticus''(현재 ''P. anglicus''와 동의어)로 잠정적으로 언급되었는데, 이는 개스페 화석과 비교적 가까운 곳에서 발견되었으며, ''P. atlanticus'' 재료가 단편적이어서 동일한 종을 나타내는지 여부를 알 수 없었기 때문이다. 이들은 1953년 로리스 S. 러셀에 의해 새로운 종 ''P. gaspesiensis''로 기술되었다.[28]
존 윌리엄 도슨은 1861년 개스페에서 발견된 줄기와 가지의 잔해를 바탕으로 석송 식물의 새로운 종 ''Selaginites formosus''를 명명했다. 샐터는 도슨에게 ''S. formosus''의 화석이 실제로 유해류의 단편적인 잔해임을 설득했으며, 이는 추가적인 발견으로 확인되었다.[29][30] 도슨은 1871년 이를 유해류로 재분류했다. 1964년 셸레스비히-와에링은 이를 ''프테리고투스''의 의문스러운 종으로 지정했다.[11]
1921년, 루데만은 뉴욕주 피츠포드의 버논 지층에서 발견된 유해류 화석군을 기술했다. 그 중 ''P. vernonensis''는 두 개의 작고 짧은 두흉갑을 기반으로 세워졌다. 눈의 윤곽과 위치는 ''프테리고투스'' 속으로 분류될 가능성을 시사하며, 거의 직사각형 모양이고 직선의 횡단 전두부를 가진다는 점에서 ''P. monroensis''와 다르다. 그는 ''슬리모니아''와의 관계를 제시했지만, 후자의 특징을 나타내는 더 많은 재료가 부족하여 그렇게 지정하지 않았다.[31] 나중에 웨링오프테루스 속으로 분류되었지만, 사무엘 J. 치우르카 주니어와 O. 에릭 테틀리는 2007년에 홀로타입이 실제로 눈을 가지고 있지 않으며 불완전한 신체 분절에 불과하다고 결론지었다. 따라서 그들은 그 종을 ''노멘 더비움''으로 간주했다.[17]
플로리다주 로크코비안 시대의 퇴적층에서 발견된 ''P. floridanus''는 대륙에서 유해류의 알려진 범위를 800 km(500마일) 이상 남쪽으로 확장했다. 이 발견 이전에 북아메리카의 유해류는 대륙 북부 지역에서만 알려져 있었으며, 뉴욕주는 화석이 가장 풍부한 주로 여겨졌다. ''P. floridanus''의 잔해는 G. 아서 쿠퍼에 의해 플로리다주 스와니 카운티에서 처음 발견되었으며, 화석은 협각의 고정된 가지와 복부 판 및 등판의 파편으로 구성되어 있으며, 1950년 에릭 N. 셸레스비히-와에링에 의해 새로운 종의 ''프테리고투스''로 결론지어졌다. 이는 ''P. cobbi''와 ''P. barrandei''와 가장 유사하며, ''P. cobbi''와는 중심 치아가 더 발달하고 발톱의 작은 치아 배열이 다르며, ''P. barrandei''와는 ''P. floridanus''가 더 가늘다는 점에서 다르다.[32]
''P. carmani''은 오하이오주 루카스 카운티의 하부 데본기 퇴적층에서 가장 흔히 발견되는 유해류이다. 1961년 에릭 N. 셸레스비히-와에링에 의해 기술되었으며, ''P. carmani'' 잔해가 발견된 지역에서 유해류를 처음 발견한 오하이오 주립 대학교의 J. 어니스트 카만 박사를 기리기 위해 명명된 ''P. carmani''는 주로 불완전한 협각과 턱 기저부에서 알려져 있다. 원래 설명의 일부인 두 개의 턱 기저부와 세 개의 협각과 함께, 알려진 화석 유물에는 복대와 전형부도 포함된다. 이 모든 원래 화석 표본은 종의 원래 설명에 따라 셸레스비히-와에링에 의해 파라타입 표본으로 지정되었다. 지정된 유형 표본은 불완전한 협각인 PE5105로, 파라타입 표본과 함께 시카고 자연사 박물관에 보관되어 있다. 이 종은 협각 치아의 특징으로 다른 ''프테리고투스'' 종과 구별할 수 있으며, 이러한 치아가 ''P. carmani''에서 덜 발달하고 더 두꺼우며, 치아가 발톱에서 현저하게 다른 배열을 가지고 있다는 점에서 ''P. barrandei'' 및 ''P. cobbi''와 다르다.
루드포디안 시대(후기 실루리아기)의 퇴적층에서 발견된 ''P. marylandicus''는 매켄지 지층에서 발견된 단편적이고 작은 텔슨으로 알려져 있으며, 1964년 셸레스비히-와에링에 의해 처음 기술되었으며, 그는 이를 ''프테리고투스''의 텔슨으로 인식했다. 표본(미국 국립 박물관의 140901번)은 매우 넓고 0.75 cm이며, 앞쪽으로 수렴하는 가장자리가 있는 거의 직선의 기저부를 가지고 있다. 일부 종과 달리 텔슨에는 톱니가 없으며 가시는 무딘 형태이다. 이 종은 텔슨의 모양으로 다른 모든 실루리아기 ''프테리고투스'' 종과 구별할 수 있다는 점에서 매우 독특하다. 초기 데본기 시대의 ''Jaekelopterus'' 종인 ''J. howelli''는 넓고 절단된 텔슨 모양과 유사하지만, 톱니와 훨씬 더 큰 말단 가시를 가지고 있어 쉽게 구별된다.[9]
미국 뉴욕주에서 후기 웬록에서 루드로 시대의 퇴적층에서 알려진 종 ''P. monroensis''는 2007년 사무엘 J. 치우르카 주니어와 O. 에릭 테틀리에 의해 눈과 두흉갑의 유사한 모양을 기반으로 ''Erettopterus osiliensis''의 동의어를 나타낼 것으로 제안되었다. 이러한 재지정은 ''P. impacatus''가 잠재적으로 ''E. osiliensis''의 동의어를 나타낼 수 있다는 점에서 다른 ''프테리고투스'' 종에도 영향을 미칠 것이다.[17] 후속 연구와 유해류 종 목록은 ''P. monroensis''와 ''P. impacatus''를 ''프테리고투스''의 별개의 종으로 계속 취급해 왔다.[16]
7. 한국 고생물학에서의 의의
8. 대중문화 속 프테리고투스
참조
[1]
서적
Museumsführer
Nationaler Geotop, National Geographic Society
2012
[2]
논문
Giant claw reveals the largest ever arthropod
[3]
논문
Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
https://www.research[...]
[4]
논문
Distribution and dispersal history of Eurypterida (Chelicerata)
http://fossilinsects[...]
[5]
논문
Eurypterids of the Devonian Holland Quarry shale of Ohio
https://www.biodiver[...]
1961
[6]
논문
The Eurypterida of New York
https://en.wikisourc[...]
1912
[7]
논문
The pterygotid telson as a biological rudder
1988
[8]
서적
Part P Arthropoda 2, Chelicerata
[9]
논문
A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)
1964
[10]
서적
The Silurian System, Founded on Geological Researches in the Counties of Salop, Hereford, Radnor, Montgomery, Caermarthen, Brecon, Pembroke, Monmouth, Gloucester, Worcester, and Stafford: With Descriptions of the Coalfields and Overlying Formations
https://archive.org/[...]
Albemarle Street
[11]
논문
A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)
1964
[12]
논문
XLI.—On the classification of some British fossil Crustacea, with notices of new forms in the University Collection at Cambridge
https://zenodo.org/r[...]
[13]
논문
Pterygotus anglicus Agassiz (Chelicerata: Eurypterida) from Atholville, Lower Devonian Campbelltown Formation, New Brunswick, Canada
2007
[14]
논문
The Silurian Eurypterida of the Welsh Borderland
1961
[15]
논문
The Horse Foot Crab
1870
[16]
간행물
A summary list of fossil spiders and their relatives
http://www.wsc.nmbe.[...]
World Spider Catalog. Natural History Museum Bern
2018
[17]
논문
Pterygotids (Chelicerata; Eurypterida) from the Silurian Vernon Formation of New York
https://www.cambridg[...]
[18]
논문
Distribution and dispersal history of Eurypterida (Chelicerata)
2007
[19]
논문
I.—Note on a New Australian Pterygotus
https://zenodo.org/r[...]
1899
[20]
논문
Re-evaluating evidence of Australian eurypterids
https://www.scienced[...]
[21]
논문
New insights into Late Devonian vertebrates and associated fauna from the Cuche Formation (Floresta Massif, Colombia)
https://www.research[...]
[22]
논문
II.—Observations on Pterygotid Eurypterids
https://www.research[...]
1964
[23]
논문
The origin of pterygotid eurypterids (Chelicerata: Eurypterida)
2009
[24]
논문
Pterygotid eurypterids (Arthropoda, Chelicerata) in the Silurian and Devonian of Bohemia
http://www.jgeosci.o[...]
1994
[25]
논문
Die Gigantostraken des Älteren Böhmischen Palaeozoicum
http://www.zobodat.a[...]
1897
[26]
논문
Embrik Strand's eurypterids
https://www.research[...]
2006
[27]
논문
Coastal-fluvial palaeoenvironments and plant palaeoecology of the Lower Devonian (Emsian), Gaspé Bay, Québec, Canada
http://sp.lyellcolle[...]
2000
[28]
논문
Pterygotus anglicus Agassiz (Chelicerata: Eurypterida) from Atholville, Lower Devonian Campbellton Formation, New Brunswick, Canada
https://www.research[...]
2007
[29]
서적
The Fossil Plants of the Devonian and Upper Silurian Formations of Canada
Geological Survey of Canada
1871
[30]
논문
The Devonian System in Canada
https://zenodo.org/r[...]
1899
[31]
논문
A recurrent Pittsford (Salina) fauna
https://biodiversity[...]
[32]
논문
A New Silurian Eurypterid from Florida
1950
[33]
논문
Do giant claws mean giant bodies? An alternative view on exaggerated scaling relationships
2008
[34]
논문
An isolated pterygotid ramus (Chelicerata: Eurypterida) from the Devonian Beartooth Butte Formation, Wyoming
2010
[35]
논문
Chapter 17: Gigantism, Dwarfism, and Cope's Rule: "Nothing in Evolution Makes Sense without a Phylogeny"
2004
[36]
논문
All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids
2015
[37]
논문
Pterygotid eurypterids (Arthropoda, Chelicerata) in the Silurian and Devonian of Bohemia
https://petrol.natur[...]
2018-05-12
[38]
논문
Nineteenth century collections of Pterygotus anglicus Agassiz (Chelicerata; Eurypterida) from the Campbellton Formation, New Brunswick, Canada
https://journals.lib[...]
2007
[39]
웹사이트
The Old Red Sandstone of Great Britain (Geological Conservation Review Series No. 31) {{!}} JNCC Resource Hub
https://hub.jncc.gov[...]
2023-08-20
[40]
웹사이트
Fossilworks: Eurypterids of the Melbourne Group, ?Dargile Formation, at Melbourne (Silurian of Australia)
http://www.fossilwor[...]
2018-02-27
[41]
웹사이트
Dargile Formation, Winneke Reservoir Site (Silurian of Australia)
https://paleobiodb.o[...]
2018-07-27
[42]
서적
Habitat of Llandoverian-Lochkovian eurypterids
A. J. Boucot, J. D. Lawson (eds.), Paleocommunities – a case study from the Silurian and Lower Devonian
1999
[43]
웹사이트
Eurypterid-Associated Biota of the Rootsikula Horizon, Saaremaa, Estonia: Rootsikula, Estonia
https://paleobiodb.o[...]
2018-07-27
[44]
웹사이트
Eurypterid-Associated Biota of the Pittsford Shale, Pittsford, New York: Ludlow, New York
https://paleobiodb.o[...]
2018-07-27
[45]
웹사이트
Platy limestone-shale facies, Upper Pridolian, Barrandian S-D boundary section: Pridoli, Czech Republic
https://paleobiodb.o[...]
2018-07-27
[46]
웹사이트
Eurypterus remipes tetragonopthalmus Community, Ustje subsuite, Podolia (Silurian of Ukraine)
https://paleobiodb.o[...]
2018-07-27
[47]
웹사이트
Eurypterid-Associated Biota of the Chortkov Horizon in Podolia (Devonian of Ukraine)
https://paleobiodb.o[...]
2018-07-27
[48]
웹사이트
Eurypterids of the Keefer Sst. Mbr, Mifflintown Fmn., Hancock, Maryland: Wenlock, Maryland
https://paleobiodb.o[...]
2018-07-27
[49]
웹사이트
Eurypterid-Associated Biota of the Ledbury Formation, Ewyas Harold, England: Pridoli, United Kingdom
https://paleobiodb.o[...]
2018-07-27
[50]
웹사이트
Sun Oil Co. well, Core 44 (3552–3568 feet): Lochkovian, Florida
https://paleobiodb.o[...]
2018-07-27
[51]
웹사이트
Gaspe community, Gaspe Sandstone (Devonian of Canada)
https://paleobiodb.o[...]
2018-07-27
[52]
서적
カンブリアンモンスター図鑑 = An Illustrated Book of cambrian monsters: カンブリア爆発の不思議な生き物たち : 図說
https://www.worldcat[...]
秀和システム
2015
[53]
서적
とても巨大な絶滅せいぶつ図鑑
https://www.worldcat[...]
世界文化社
2019
[54]
서적
よみがえる恐竜・古生物 超ビジュアルCG版
https://www.worldcat[...]
フトバンククリエイティブ
2006-07
[55]
논문
A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)
https://www.jstor.or[...]
1964
[56]
논문
Segmentation and tagmosis in Chelicerata
https://www.academia[...]
[57]
논문
Pterygotus Anglicus Agassiz (chelicerata: Eurypterida) from Atholville, Lower Devonian Campbellton Formation, New Brunswick, Canada
https://onlinelibrar[...]
2007
[58]
논문
Nineteenth century collections of Pterygotus anglicus Agassiz (Chelicerata; Eurypterida) from the Campbellton Formation, New Brunswick, Canada
https://journals.lib[...]
2007
[59]
논문
Morphology, taxonomy, and classification of the order Eurypterida Burmeister, 1843
https://www.cambridg[...]
1989-09
[60]
논문
All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids
https://www.ncbi.nlm[...]
2015-08
[61]
논문
A new identity for the Silurian arthropod Necrogammarus
https://www.palass.o[...]
[62]
논문
Ichnology of the Devonian (Emsian) Campbellton Formation, New Brunswick, Canada
https://journals.lib[...]
2017-02-08
[63]
논문
Giant claw reveals the largest ever arthropod
https://www.ncbi.nlm[...]
2008-02-23
[64]
논문
Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates
https://www.research[...]
2009
[65]
논문
The pterygotid telson as a biological rudder
https://onlinelibrar[...]
1988
[66]
서적
カラー図解古生物たちのふしぎな世界 : 繁栄と絶滅の古生代3億年史
https://www.worldcat[...]
講談社
2017-06
[67]
웹사이트
Fossilworks: Pterygotus
http://fossilworks.o[...]
2020-12-24
[68]
서적
節足動物の多様性と系統
https://www.worldcat[...]
裳華房
2008
[69]
논문
The origin of pterygotid eurypterids (Chelicerata: Eurypterida)
https://onlinelibrar[...]
2009
[70]
논문
The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa
https://doi.org/10.1[...]
2015-09-01
[71]
논문
The Silurian Eurypterida of the Welsh Borderland
https://www.jstor.or[...]
1961
[72]
논문
II.—Observations on Pterygotid Eurypterids*
https://www.cambridg[...]
1964
[73]
논문
An isolated pterygotid ramus (Chelicerata: Eurypterida) from the Devonian Beartooth Butte Formation, Wyoming
https://www.research[...]
2010-11
[74]
간행물
A summary list of fossil spiders and their relatives
https://wsc.nmbe.ch/[...]
Natural History Museum Bern
2020
[75]
간행물
Arthropods from the Lower Devonian (Lower Emsian) of Alken an der Mosel, Germany, Pt. 4, Eurypterida, Drepanopteridae and other groups
1974
[76]
논문
Distribution and dispersal history of Eurypterida (Chelicerata)
http://www.sciencedi[...]
2007-09-03
[77]
논문
Pterygotids (Chelicerata; Eurypterida) from the Silurian Vernon Formation of New York
https://www.cambridg[...]
2007-07
[78]
논문
Two New Eurypterids from the Silurian of Indiana
https://www.jstor.or[...]
1948
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com