엔도솜
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
엔도솜은 세포 내에서 물질을 분류하고 분해하는 데 중요한 역할을 하는 세포 소기관이다. 초기 엔도솜, 후기 엔도솜, 재활용 엔도솜의 세 가지 주요 유형으로 나뉘며, 각기 다른 마커와 기능을 갖는다. 엔도솜은 골지체, 세포막, 리소좀 등과 연결된 다양한 경로를 통해 물질을 수송하며, LDL, EGF, 트랜스페린 등 다양한 분자의 수송과 처리에 관여한다.
더 읽어볼만한 페이지
- 소포 (세포) - 액포
액포는 세포 내 막으로 둘러싸인 소기관으로, 액체와 물질을 저장하며 삼투압 조절, 노폐물 저장, 자가포식 등의 기능을 수행하고, 식물 세포에서 큰 비중을 차지하는 중심 액포를 포함하여 수축포, 식포, 가스포 등으로 나뉜다. - 소포 (세포) - 리소좀
리소좀은 세포 내 소기관으로서 가수분해 효소를 이용해 세포 내 물질을 분해하며, 자가 소화 작용, 외부 물질 분해, 칼슘 저장 등 다양한 기능에 관여하고, 기능 이상은 질병과 관련되며 노화에도 영향을 미친다. - 세포소기관 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포소기관 - 세포막
세포막은 세포질을 둘러싸고 세포 내외부 환경을 구분하는 선택적 투과성 막으로, 인지질 이중층과 단백질, 탄수화물로 구성되어 물질 수송, 세포 형태 유지, 세포 간 신호 전달, 세포 접합 등의 기능을 수행하며, 유동 모자이크 모델로 설명된다.
| 엔도솜 | |
|---|---|
| 기본 정보 | |
 | |
| 위치 | 세포질 내 |
| 기능 | 세포내이입된 물질의 수송 및 분류 |
| 세부 유형 | |
| 주요 유형 | 초기 엔도솜 후기 엔도솜 재활용 엔도솜 |
| 기타 유형 | 운반 엔도솜 다포체 |
| 식별 | |
| 유전자 | 다수 (예: Rab5, Rab7) |
| 추가 정보 | |
| 관련 질병 | 알츠하이머병, 니만-피크병 C형 |
2. 엔도솜의 기능
엔도솜은 리소좀에 도달하기 전 물질을 분류하는 환경을 제공한다.[2] 예를 들어 저밀도 지단백질(LDL)은 LDL 수용체에 결합하여 세포 내로 유입된 후 초기 엔도솜에서 분리되고, 수용체는 세포 표면으로 재활용되지만, LDL은 리소좀으로 전달되어 처리된다. 상피 성장 인자(EGF)와 EGF 수용체는 리소좀으로 전달될 때까지 pH 저항성 결합을 유지하며, 만노스 6-인산 수용체는 골지체에서 나온 리간드를 리소좀으로 운반한다.
2. 1. 분자 메커니즘
엔도솜은 분해성 리소좀에 도달하기 전에 물질을 분류하는 환경을 제공한다.[2] 예를 들어, 저밀도 지단백질(LDL)은 세포 표면에서 LDL 수용체에 결합하여 세포 내로 유입된다. 초기 엔도솜에 도달하면 LDL은 수용체로부터 분리되고, 수용체는 세포 표면으로 재활용될 수 있다. LDL은 엔도솜에 남아 리소좀으로 전달되어 처리된다. LDL은 V-ATPase라는 액포 막 양성자 펌프에 의해 생성된 초기 엔도솜의 약간 산성화된 환경 때문에 분리된다. 반면에, 상피 성장 인자(EGF)와 EGF 수용체는 분해를 위해 리소좀으로 전달될 때까지 지속되는 pH 저항성 결합을 가지고 있다. 만노스 6-인산 수용체는 유사한 메커니즘을 통해 리소좀으로 이동할 골지체에서 나온 리간드를 운반한다.3. 엔도솜의 종류
엔도솜은 세포 내부로 들어온 물질을 처리하는 세포 내 소기관이다. 엔도솜은 크게 초기 엔도솜, 후기 엔도솜, 재활용 엔도솜의 세 가지 유형으로 나뉜다.[2] 이들은 엔도사이토시스된 물질이 도달하는 데 걸리는 시간과 Rabs와 같은 특정 표지 단백질(마커)로 구분할 수 있으며, 서로 다른 형태를 띤다.[3]
내세포작용을 통해 세포 안으로 들어온 소포는 막을 둘러싸고 있던 단백질 코팅(클라트린)을 벗은 후 초기 엔도솜과 융합한다. 초기 엔도솜은 이후 리소좀과 융합되기 전에 후기 엔도솜으로 성숙한다.[4][5] 이 과정에서 V-ATPase의 작용으로 내부가 점점 산성화된다.[6] 재활용되는 분자들은 초기 엔도솜의 관 모양 영역에 모여 있다가 떨어져 나가고, 이로 인해 후기 엔도솜에는 관 모양 구조가 거의 남지 않게 된다. 또한 초기 엔도솜은 더 큰 소포들과 융합하여 크기가 커진다.[7]
일부 물질은 초기 엔도솜에서 직접 세포막으로 재활용되기도 하지만,[9] 대부분은 재활용 엔도솜을 거쳐 세포막으로 돌아간다.
극성화된 세포나 대식세포와 같은 특수한 세포에는 추가적인 하위 유형의 엔도솜이 존재한다. 식세포작용으로 형성된 파고솜(phagosome), 다량음세포작용으로 형성된 마크로피노솜(macropinosome), 자가포식 작용으로 형성된 오토파고솜(autophagosome) 등도 엔도솜과 유사한 방식으로 성숙하며, 정상적인 엔도솜과의 융합이 필요할 수 있다.[14] 일부 세포 내 병원체는 RAB7 획득을 방해하는 등 이러한 성숙 과정을 방해하기도 한다.[15]
후기 엔도솜/MVB는 때때로 '내세포 수송 소포'라고 불리기도 하지만, 이 용어는 원래 초기 엔도솜에서 떨어져 나와 후기 엔도솜과 융합하는 소포를 가리키는 말이었다. 그러나 최근 연구에 따르면, 이러한 두 엔도솜 사이의 물질 수송은 소포 수송보다는 성숙 과정을 통해 일어날 가능성이 더 높다.
각 엔도솜은 막을 구성하는 지질 성분에 차이가 있다. 예를 들어, 세포막에는 PI(4,5)P2가, 초기 엔도솜에는 PI(3)P가, 후기 엔도솜에는 PI(3,5)P2가, 트랜스 골지 네트워크에는 PI(4)P가 주로 존재한다.[16]
3. 1. 초기 엔도솜 (Early Endosome)
초기 엔도솜은 동적인 관형-소포 네트워크로 구성되어 있으며, 직경이 최대 1 μm인 소포와 약 50 nm 직경의 연결된 소관으로 이루어져 있다.[13] RAB5A, RAB4, Transferrin 및 그 수용체, EEA1과 같은 마커를 통해 확인할 수 있다.[13]내세포작용된 소포가 코팅을 벗으면 초기 엔도솜과 융합된다.[4] 초기 엔도솜은 V-ATPase의 활동을 통해 점점 더 산성화되며, 재활용되는 많은 분자는 초기 엔도솜의 관형 영역에 농축되어 제거된다.[6] 이 과정에서 소관이 재활용 경로로 손실되면서 후기 엔도솜에는 소관이 거의 남지 않게 된다. 또한, 초기 엔도솜은 더 큰 소포로 동형 융합되어 크기가 증가한다.[7]
분자는 또한 엔도솜 내강으로 perimeter 막에서 새싹이 생기는 더 작은 소포로 정렬되어 내강 소포 (ILV)를 형성한다. 이로 인해 후기 엔도솜은 다포성 외관을 가지게 되며, 다포성 엔도솜 또는 다포성 소체 (MVB)라고도 불린다. transferrin receptor 및 mannose 6-인산 수용체와 같은 재활용 분자의 제거는 이 기간 동안 계속되며, 소포가 엔도솜 밖으로 새싹이 생겨 나오는 방식으로 이루어지는 것으로 추정된다.[4]
최종적으로, 엔도솜은 RAB5A를 잃고 RAB7A를 획득하여 리소좀과의 융합이 가능하게 된다.[7] 일부 물질은 초기 엔도솜에서 직접 세포막으로 재활용되기도 한다.[9]
3. 2. 후기 엔도솜 (Late Endosome)
엔도솜에는 초기 엔도솜, 후기 엔도솜, 재활용 엔도솜의 세 가지 유형이 있다.[2] 후기 엔도솜은 RAB7, RAB9, 만노스 6-인산 수용체[10]와 같은 마커로 구별된다. 또한, 후기 엔도솜 막에는 다른 어떤 소기관 막에서도 발견되지 않는 특이하고 고유한 지질인 BMP 또는 LBPA가 포함되어 있다.[11][12]초기 엔도솜은 여러 단계를 거쳐 후기 엔도솜으로 성숙한다.
- V-ATPase의 활동을 통해 점점 더 산성화된다.[6]
- 재활용되는 분자들이 초기 엔도솜의 관형 영역에 농축되어 제거되면서 후기 엔도솜에는 소관이 거의 없게 된다.
- 초기 엔도솜이 더 큰 소포로 동형 융합되어 크기가 증가한다.[7]
- 분자들이 엔도솜 내강으로 향하는 작은 소포로 정렬되어 내강 소포(ILV)를 형성한다. 이로 인해 후기 엔도솜은 다포성 외관을 가지게 되며, 다포성 엔도솜 또는 다포성 소체(MVB)라고도 불린다.
- transferrin receptor 및 만노스 6-인산 수용체와 같은 재활용 분자의 제거가 계속된다.[4]
- 엔도솜은 RAB5A를 잃고 RAB7A를 획득하여 리소좀과의 융합이 가능하게 된다.[7]
후기 엔도솜과 리소좀의 융합은 두 소포 구획의 중간 특성을 가진 '하이브리드' 구획을 형성한다.[8] 리소좀은 정상적인 더 높은 밀도로 재응축되어 재형성되기 전에 더 많은 후기 엔도솜과 융합될 수 있다.
후기 엔도솜/MVB는 때때로 '내세포 수송 소포'라고 불리기도 하지만, 이 용어는 초기 엔도솜에서 새싹이 생겨 후기 엔도솜과 융합되는 소포를 설명하는 데 사용되었다. 그러나 현재는 소포 수송보다는 성숙 과정을 통해 이러한 두 구획 간의 수송이 발생할 가능성이 더 높다는 것이 밝혀졌다.
엔도솜 막의 지질 구성은 종류에 따라 다르다. PI(4,5)P2는 세포막에, PI(3)P는 초기 엔도솜에, PI(3,5)P2는 후기 엔도솜에, PI(4)P는 trans Golgi network에 존재한다.[16]
3. 3. 재활용 엔도솜 (Recycling Endosome)
일부 물질은 초기 엔도솜에서 직접 세포막으로 재활용되지만,[9] 대부분은 재활용 엔도솜을 통해 이동한다.재활용 엔도솜은 미세소관 조직 중심에 집중되어 있으며 주로 관형 네트워크로 구성되어 있다. RAB11을 마커로 사용한다.[13]
4. 엔도솜 관련 경로
엔도솜은 세포 내에서 다양한 경로를 통해 다른 소기관들과 물질을 주고받는다. 주요 경로는 다음과 같다.
- 골지체-엔도솜 경로: 소포체는 골지체와 엔도솜 사이를 양방향으로 이동하며, 특정 단백질들을 운반한다.
- 세포막-초기 엔도솜-재활용 엔도솜 경로: 세포막에서 세포내이입된 물질들은 초기 엔도솜을 거쳐 재활용되거나 리소좀으로 이동한다. 저밀도 지단백질(LDL), 상피 성장 인자(EGF), 트랜스페린 등이 이 경로를 따른다.
- 후기 엔도솜-리소좀 경로: 후기 엔도솜은 리소좀과 융합하여 내강의 물질들을 분해한다. 이 경로는 한 방향으로만 진행된다.
이러한 경로들은 서로 겹치기도 하고, 특정 세포에서는 더욱 복잡한 양상을 보인다. 예를 들어 상피 세포에서는 트랜스사이토시스라는 과정을 통해 물질이 세포의 한쪽에서 다른 쪽으로 이동하기도 한다. 또한, 후기 엔도솜이 리소좀 대신 세포막과 융합하여 엑소좀을 방출하는 경우도 있다.
4. 1. 골지체-엔도솜 경로
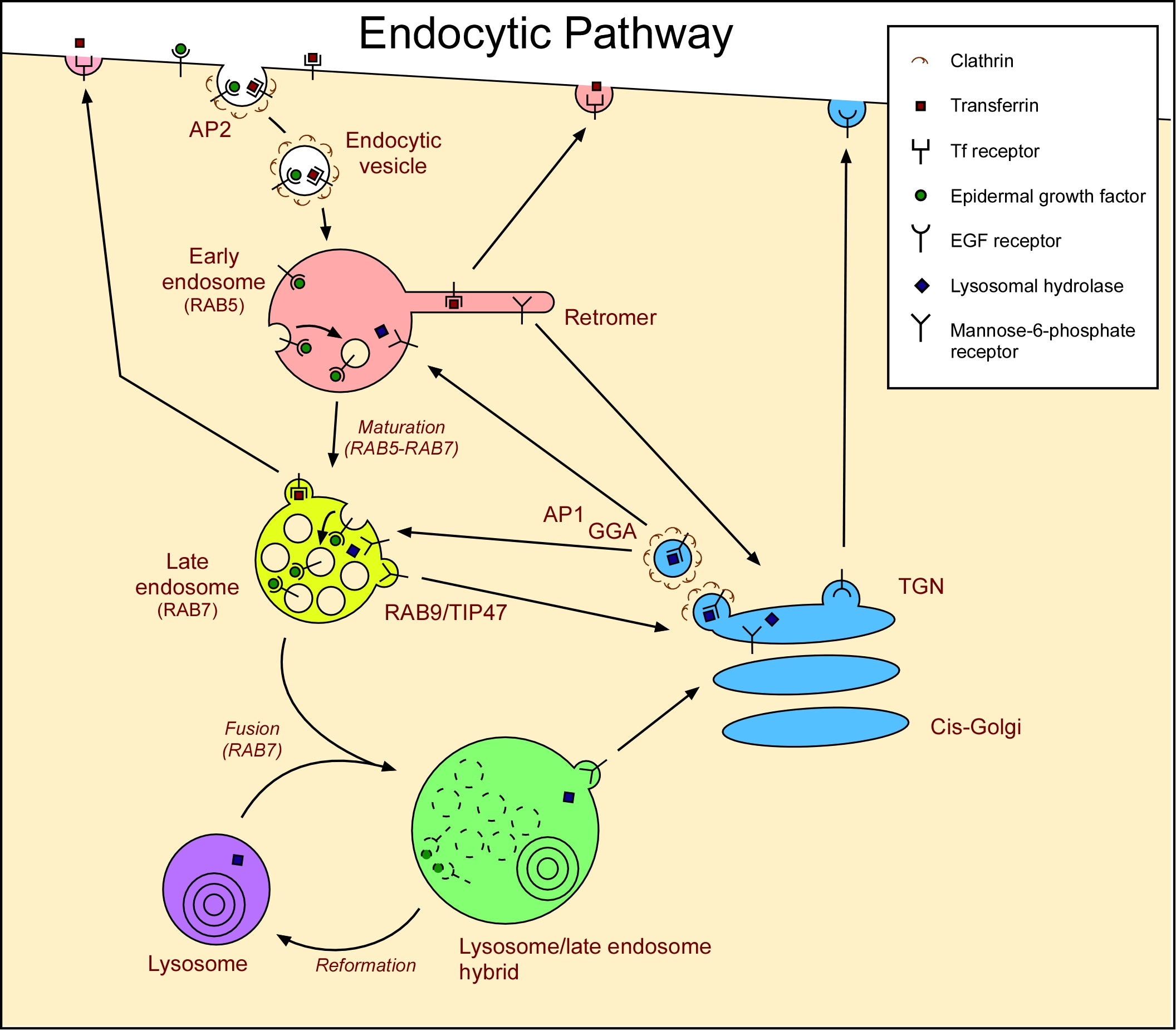
엔도솜과 연결되는 경로는 세 가지 주요 구획이 있다. 멜라닌 세포 및 분극 세포와 같은 특수 세포에는 더 많은 경로가 존재한다. 예를 들어, 상피 세포에서 트랜스사이토시스라는 특수한 과정을 통해 일부 물질이 세포의 한쪽으로 들어가 반대쪽에서 나올 수 있다. 또한, 어떤 상황에서는 후기 엔도솜/MVB가 리소좀 대신 세포막과 융합되어 루멘 소포( 엑소좀)를 세포외 매질로 방출한다.
이러한 경로의 정확한 특성에 대한 합의는 없으며, 주어진 상황에서 주어진 화물이 거치는 순차적인 경로는 논쟁의 대상이 되곤 한다.
소포체는 골지체와 엔도솜 사이를 양방향으로 이동한다. GGA와 AP-1 클라트린 코팅 소포 어댑터는 분자를 엔도솜으로 운반하는 소포체를 골지체에서 생성한다.[18] 반대 방향으로, 레트로머는 초기 엔도솜에서 소포체를 생성하여 분자를 골지체로 되돌려 보낸다. 일부 연구에서는 Rab9 및 TIP47에 의해 매개되는 후진 수송 경로가 후기 엔도솜에서 골지체로 이동한다고 설명하지만, 다른 연구에서는 이러한 발견에 이의를 제기한다. 이러한 경로를 따르는 분자에는 리소좀 가수분해효소를 내포성 경로로 운반하는 mannose-6-phosphate 수용체가 포함된다. 가수분해효소는 엔도솜의 산성 환경에서 방출되고, 수용체는 레트로머와 Rab9에 의해 골지체로 회수된다.
4. 2. 세포막-초기 엔도솜-재활용 엔도솜 경로
저밀도 지단백질(LDL)은 세포 표면에서 LDL 수용체에 결합하여 세포 내로 유입된다. 초기 엔도솜에 도달하면 LDL은 수용체로부터 분리되고, 수용체는 세포 표면으로 재활용될 수 있다. LDL은 엔도솜에 남아 리소좀으로 전달되어 처리된다. LDL은 V-ATPase라는 액포 막 양성자 펌프에 의해 생성된 초기 엔도솜의 약간 산성화된 환경 때문에 분리된다.[2]분자는 세포내이입 소포를 통해 세포막에서 초기 엔도솜으로 전달된다. 클라트린-코팅 소포에서 수용체 매개 세포내이입을 통해 내부화될 수 있으며, 카베올린을 사용하는 소포를 포함하여, 이 경로를 위해 다른 유형의 소포도 세포막에서 형성된다. 소포는 분자를 세포막으로 직접 다시 수송하지만, 많은 분자는 먼저 재활용 엔도솜과 융합되는 소포에서 수송된다.[19] 이 재활용 경로를 따르는 분자는 초기 엔도솜의 소관에 집중된다.
이러한 경로를 따르는 분자에는 수용체인 LDL, 상피 성장 인자(EGF), 철 수송 단백질 트랜스페린이 포함된다. 이러한 수용체의 세포막에서 내부화는 수용체 매개 세포내이입에 의해 발생한다. LDL은 낮은 pH로 인해 엔도솜에서 방출되고, 수용체는 세포 표면으로 재활용된다. 콜레스테롤은 주로 (LDL)에 의해 혈액으로 운반되며, LDL 수용체에 의한 수송은 콜레스테롤이 세포에 흡수되는 주요 메커니즘이다.
EGF 수용체(EGFR)은 EGF가 결합하면 활성화된다. 활성화된 수용체는 리소좀에서 자체 내부화 및 분해를 자극한다. EGF는 엔도솜으로 세포내이입되면 EGFR에 결합된 상태로 유지된다. 활성화된 EGFR은 자체 유비퀴틴화를 자극하며, 이는 루멘 소포로 향하게 하여 세포막으로 재활용되지 않도록 한다. 이는 시토졸에서 단백질의 신호 전달 부분을 제거하여 성장의 지속적인 자극을 방지한다.[20] EGF로 자극받지 않은 세포에서 EGFR은 EGF가 결합되지 않으므로 엔도솜에 도달하면 재활용된다.[21]
트랜스페린도 수용체와 관련되어 있지만, 산성 엔도솜에서 철은 트랜스페린에서 방출된 다음, 철이 없는 트랜스페린(여전히 트랜스페린 수용체에 결합)은 초기 엔도솜에서 세포 표면으로 직접 그리고 재활용 엔도솜을 통해 돌아온다.[22]
4. 3. 후기 엔도솜-리소좀 경로
후기 엔도솜에서 리소좀으로의 수송은 기본적으로 한 방향으로만 이루어진다. 후기 엔도솜은 리소좀과 융합되는 과정에서 "소모"되기 때문이다(때로는 엔도리소좀이라고도 함).[23][24] 따라서 엔도솜 내강에 있는 가용성 분자는 특별히 회수되지 않는 한 리소좀으로 이동한다. 막단백질은 리소좀의 주변 막 또는 내강으로 전달될 수 있다. 리소좀 내강으로 향하는 막단백질은 주변 막에서 엔도솜으로 싹트는 소포로 분류되는데, 이 과정은 초기 엔도솜에서 시작된다. 엔도솜 내에서 소포를 생성하는 과정은 후기 엔도솜, 엔도리소좀 또는 리소좀에서만 발견되는 특이한 지질인 BMP 또는 LBPA에 의해 향상되는 것으로 생각된다.[12]
엔도솜이 후기 엔도솜/MVB로 성숙하여 리소좀과 융합되면 내강 내 소포가 리소좀 내강으로 전달된다. 단백질은 유비퀴틴을 추가하여 이 경로로 표시된다.[25] 수송에 필요한 엔도좀 분류 복합체 (ESCRT)는 이 유비퀴틴을 인식하여 단백질을 형성되는 내강 소포로 분류한다.[26] 이러한 경로를 따르는 분자에는 LDL과 만노스-6-인산 수용체에 의해 전달되는 리소좀 가수분해효소, EGF에 결합된 막관통 EGFR 등이 있다. 이러한 가용성 분자는 엔도솜에 남아 있으므로 리소좀으로 전달된다.
참조
[1]
저널
Late endosomes derive from early endosomes by maturation
1991-05-01
[2]
저널
Endocytosis and molecular sorting
[3]
저널
Rab GTPases as coordinators of vesicle traffic
2009-08-01
[4]
저널
Multivesicular endosomes containing internalized EGF-EGF receptor complexes mature and then fuse directly with lysosomes
1996-03-01
[5]
저널
Lysosome-endosome fusion and lysosome biogenesis
2000-05-01
[6]
저널
Regulation of the V-ATPase along the endocytic pathway occurs through reversible subunit association and membrane localization
2008-07-01
[7]
저널
Rab conversion as a mechanism of progression from early to late endosomes
2005-09-01
[8]
저널
Fusion of lysosomes with late endosomes produces a hybrid organelle of intermediate density and is NSF dependent
1998-02-01
[9]
저널
Internalization and processing of transferrin and the transferrin receptor in human carcinoma A431 cells
1983-08-01
[10]
저널
Molecular mechanisms of late endosome morphology, identity and sorting
2006-08-01
[11]
저널
Separation and characterization of late endosomal membrane domains
2002-08-30
[12]
저널
Bis(monoacylglycero)phosphate, a peculiar phospholipid to control the fate of cholesterol: Implications in pathology
https://pubmed.ncbi.[...]
2009-11-01
[13]
저널
Rab11 regulates recycling through the pericentriolar recycling endosome
1996-11-01
[14]
저널
Autophagy and multivesicular bodies: two closely related partners
2009-01-01
[15]
저널
Biogenesis of Leishmania major-harboring vacuoles in murine dendritic cells
2006-02-01
[16]
저널
Membrane lipids: where they are and how they behave
2008-02-01
[17]
저널
Phosphoinositides in cell regulation and membrane dynamics
2006-10-01
[18]
저널
The GGA proteins: key players in protein sorting at the trans-Golgi network
2004-07-01
[19]
저널
Pathways and mechanisms of endocytic recycling
2009-09-01
[20]
저널
Human VPS34 is required for internal vesicle formation within multivesicular endosomes
2001-12-01
[21]
저널
Kinase activity controls the sorting of the epidermal growth factor receptor within the multivesicular body
1990-05-01
[22]
저널
Receptor-mediated endocytosis: the intracellular journey of transferrin and its receptor
1986-03-01
[23]
저널
Mechanisms of SARS-CoV-2 entry into cells
2022-01-01
[24]
저널
Role of Endolysosomes in Severe Acute Respiratory Syndrome Coronavirus-2 Infection and Coronavirus Disease 2019 Pathogenesis: Implications for Potential Treatments
2020-01-01
[25]
저널
Regulation of membrane protein transport by ubiquitin and ubiquitin-binding proteins
[26]
저널
ESCRT complexes and the biogenesis of multivesicular bodies
2008-02-01
[27]
저널
2011-02-20
[28]
저널
Rab9 GTPase Regulates Late Endosome Size and Requires Effector Interaction for Its Stability
http://www.pubmedcen[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com