세포막
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
세포막은 모든 세포를 둘러싸는 얇은 막으로, 세포 내외부 환경을 분리하고 세포의 형태를 유지하며 물질 수송, 신호 전달, 세포 부착, 효소 활성 등 다양한 기능을 수행한다. 세포막은 지질(인지질, 당지질, 스테롤), 단백질, 탄수화물로 구성되며, 유동 모자이크 모델로 설명된다. 세포막은 수동 수송, 능동 수송, 세포내이입, 세포외유출 등 다양한 기전을 통해 물질을 수송하며, 세포 신호 전달, 세포 부착, 효소 활성, 막 전위 유지 등에도 관여한다. 원핵세포의 세포막은 그람 양성균과 그람 음성균에 따라 구조가 다르며, 지방산 조성에 따라 유동성이 조절된다.
더 읽어볼만한 페이지
- 생화학 막에 관한 - 인지질
인지질은 글리세롤에 두 개의 지방산과 하나의 인산기가 결합된 양친매성 지질로, 세포막의 주요 구성 성분이며 글리세로인지질과 스핑고지질로 나뉘어 세포막 구조와 기능, 신호 전달 등에 중요한 역할을 하고 약물 전달 시스템 및 식품 산업에도 활용된다. - 세포해부학 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포해부학 - 세포벽
세포벽은 세포막 외부에 위치하여 세포 형태 유지, 기계적 스트레스 방어, 삼투압 안정화에 기여하는 외피로, 식물의 셀룰로오스, 균류의 키틴, 세균의 펩티도글리칸 등 생물 종에 따라 구성 성분과 구조가 다양하며, 고세균은 슈도펩티도글리칸 등 다른 유형의 세포벽을 가진다. - 세포소기관 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포소기관 - 호르몬
호르몬은 특정 기관의 기능을 조절하는 정보 전달 물질로, 다양한 화학적 구조와 신호 전달 방식을 가지며, 척추동물의 성장, 발달, 생식, 신진대사 등 생리 과정에 관여하고, 의약품으로도 사용되지만 부작용의 위험이 있다.
| 세포막 | |
|---|---|
| 일반 정보 | |
| 구조 | 지질 이중층 막 단백질 막 지질 |
| 기능 | |
| 주요 역할 | 세포 내부와 외부 환경을 분리하는 생물학적 막 |
| 추가 역할 | 세포 안팎으로 물질 이동 조절 세포 신호 전달 |
| 구성 요소 | |
| 지질 | 인지질 스핑고지질 스테롤 |
| 단백질 | 막 단백질 내재성 단백질 외재성 단백질 |
| 탄수화물 | 당지질 당단백질 |
| 구조적 특징 | |
| 형태 | 유동 모자이크 모델 |
| 유동성 | 세포막 내 지질과 단백질 이동 가능 |
| 투과성 | |
| 반투과성 | 특정 물질만 통과 가능 |
| 수동 수송 | 확산 삼투 |
| 능동 수송 | 에너지 소모를 통해 물질 이동 |
| 생체 막 형성 | |
| 형성 방법 | 생체 모방 결합 반응을 통해 조립 가능 |
| 기타 | |
| 인공 세포막 | 화학자가 인공적으로 합성 |
| 내부 막 점도 | 세포막 형태에 영향 |
2. 역사
세포막에 대한 연구는 로버트 훅(Robert Hooke)이 1665년에 세포를 발견하고 세포설이 제창되면서 시작되었지만, 초기에는 세포막의 중요성이 간과되었다. 당시에는 식물 세포만 관찰되었기 때문에 모든 세포에 단단한 세포벽이 있다고 오해[9]했다. 이후 현미경 기술이 발전하면서 19세기 초에는 식물 세포가 분리될 수 있다는 사실이 밝혀졌고, 세포는 개별적인 독립체이며 서로 연결되지 않고 각각의 세포벽으로 둘러싸여 있다는 인식이 확산되었다.
19세기 후반에도 현미경 기술은 세포막과 세포벽을 명확히 구분할 수준은 아니었다. 하지만 일부 학자들은 동물 세포 내 물질의 이동 특성을 근거로 세포막의 존재를 추론하고, 이것이 식물 세포벽과는 다른 구조임을 인지했다. 그러나 세포막이 모든 세포에 필수적인 것은 아니라는 주장도 있었고, 19세기 말까지 세포막의 존재 자체를 부정하는 시각도 많았다. 1890년에는 세포막이 존재하지만 부차적인 구조에 불과하다는 수정된 세포 이론이 제기되기도 했다.
삼투압과 투과성에 대한 연구가 진행되면서 세포막의 중요성이 부각되기 시작했고,[9] 1895년 어니스트 오버턴(Ernest Overton)은 세포막이 지질로 구성되어 있다고 제안했다.[10] 1925년 고르터(Gorter)와 그렌델(Grendel)은 결정학적 연구와 비누방울 관찰을 바탕으로 세포막이 지질 이중층 구조라는 가설[11]을 제시했다. 이들은 적혈구의 지질을 추출하여 표면적을 측정하고, 이를 통해 세포막이 지질 이중층으로 구성되어 있다는 결론[9][12]을 내렸다.
같은 해, 프릭(Fricke)은 적혈구와 효모 세포막의 두께가 3.3~4 nm로 지질 단층과 일치함을 확인했다. 또한, 레프토스코프(leptoscope)라는 새로운 장비를 통해 막의 두께를 더 정밀하게 측정할 수 있게 되었고, 이를 통해 지질 이중층 가설이 더욱 뒷받침되었다. 1930년대 후반에는 휴 데이브슨(Hugh Davson)과 제임스 대니얼리(James Danielli)가 파우시몰레큘러 모델(paucimolecular model)[13][9]을 제시하며, 지질 이중층이 두 개의 단백질 층 사이에 있다는 모델을 제안했다. 이 모델은 세이무어 조너선 싱어(Seymour Jonathan Singer)와 가스 니콜슨(Garth L. Nicolson)의 유동 모자이크 모델(1972)이 등장하기 전까지 30년간 지배적인 모델로 자리 잡았다.[13][9]
유동 모자이크 모델은 1970년대에 처음 제안된 이후 현재까지 세포막의 주요 모델로 인정받고 있다.[9] 이 모델은 세포막이 친수성 머리 부분과 소수성 꼬리 부분을 가진 지질 이중층으로 구성되어 있으며, 단백질이 이중층에 박혀 있거나 표면에 결합된 형태로 존재한다고 설명한다. 유동 모자이크 모델은 세포막의 유동성과 단백질의 다양한 위치 및 기능을 설명하며, 세포막 연구에 큰 영향을 미쳤다.
이처럼 세포막에 대한 인식은 오랜 시간에 걸쳐 발전해 왔다. 초기에는 세포막의 존재와 중요성이 간과되었지만, 현미경 기술의 발전과 다양한 연구를 통해 세포막의 구조와 기능이 점차 밝혀졌다.
2. 1. 세포막의 발견과 초기 연구
로버트 훅(Robert Hooke)이 1665년에 세포를 발견한 후 세포 이론이 제기되었다. 당시에는 식물 세포만 관찰할 수 있었기 때문에 처음에는 모든 세포에 단단한 세포벽이 있다고 생각되었다.[9] 19세기 초, 식물 세포가 분리될 수 있다는 사실이 밝혀지면서 세포는 개별적인 독립체이며, 서로 연결되지 않고 각각의 세포벽으로 둘러싸여 있다는 것이 인식되었다. 이 이론은 동물 세포까지 확장되어 세포 보호 및 발달에 대한 보편적인 메커니즘을 제시하였다.19세기 후반에도 현미경 기술은 세포막과 세포벽을 구별하기에는 충분하지 않았다. 그러나 일부 현미경 학자들은 당시에 보이지 않더라도 세포 내부 구성 요소는 내부적으로는 이동하지만 외부로는 이동하지 않는다는 점으로 미루어 동물 세포에는 세포막이 존재한다고 정확하게 파악했고, 세포막이 식물 세포벽과 동일하지 않다는 것을 인지했다. 또한 세포막이 모든 세포에 필수적인 구성 요소는 아니라는 점도 추론되었다. 19세기 말까지도 많은 사람들이 세포막의 존재를 부정했다. 1890년, 세포 이론의 수정을 통해 세포막이 존재하지만 단순히 이차적인 구조일 뿐이라는 주장이 제기되었다. 삼투압 및 투과성에 대한 후속 연구를 통해서야 세포막이 더욱 인정받게 되었다.[9] 1895년, 어니스트 오버턴(Ernest Overton)은 세포막이 지질로 구성되어 있다고 제안했다.[10]
1925년, 고르터(Gorter)와 그렌델(Grendel)이 제안한 지질 이중층 가설[11]은 결정학적 연구와 비누방울 관찰을 바탕으로 세포막 이중층 구조에 대한 추측을 만들어냈다. 이 가설을 받아들이거나 기각하기 위해 연구자들은 세포막의 두께를 측정했다. 연구자들은 인간 적혈구에서 지질을 추출하여 물 표면에 퍼졌을 때 지질이 덮는 표면적을 측정했다. 성숙한 포유류 적혈구는 핵과 세포질 소기관이 모두 없기 때문에 세포막은 세포 내 유일한 지질 함유 구조이다. 따라서 세포에서 추출한 모든 지질은 세포의 세포막에 있었다고 가정할 수 있다. 추출된 지질이 덮는 물의 표면적과 지질이 추출된 적혈구의 계산된 표면적의 비율은 약 2:1이었고, 그들은 세포막이 지질 이중층으로 구성되어 있다고 결론지었다.[9][12]
1925년, 프릭(Fricke)은 적혈구와 효모 세포막의 두께가 3.3~4 nm 범위에 있다는 것을 밝혀냈는데, 이는 지질 단층과 호환되는 두께였다. 이러한 연구에 사용된 비유전율의 선택에 의문이 제기되었지만, 이후 실험에서도 초기 실험 결과를 반박할 수 없었다. 독립적으로, 레프토스코프(leptoscope)가 발명되어 표본에서 반사되는 빛의 강도를 알려진 두께의 막 표준의 강도와 비교하여 매우 얇은 막을 측정할 수 있게 되었다. 이 장치는 pH 측정 및 8.6~23.2 nm 범위의 막 단백질 존재 여부에 따라 달라지는 두께를 측정할 수 있었는데, 더 낮은 측정값은 지질 이중층 가설을 뒷받침했다. 1930년대 후반, 휴 데이브슨(Hugh Davson)과 제임스 대니얼리(James Danielli)(1935)의 파우시몰레큘러 모델(paucimolecular model)이 세포막 구조 모델로 일반적으로 받아들여지게 되었다.[13][9] 이 모델은 기름과 극피동물 알 사이의 표면 장력 연구를 기반으로 했다. 표면 장력 값이 기름-물 계면에서 예상되는 값보다 훨씬 낮게 나타났기 때문에, 세포 표면의 계면 장력을 낮추는 물질이 있다고 가정되었다. 지질 이중층이 두 개의 얇은 단백질 층 사이에 있다는 것이 제안되었다. 파우시몰레큘러 모델은 즉시 인기를 얻었고, 세이무어 조너선 싱어(Seymour Jonathan Singer)와 가스 니콜슨(Garth L. Nicolson)(1972)의 유동 모자이크 모델에 의해 경쟁이 될 때까지 30년 동안 세포막 연구를 지배했다.[13][9]
유동 모자이크 모델 이전에 제안된 수많은 세포막 모델에도 불구하고, 유동 모자이크 모델은 1970년대에 처음 제안된 이후로 오랫동안 세포막의 주요 원형으로 남아 있다.[9] 유동 모자이크 모델은 현대의 발견들을 상세히 설명하도록 수정되었지만, 기본 원리는 일관되게 유지되었다. 즉, 세포막은 친수성 외부 머리와 소수성 내부로 구성된 지질 이중층이며, 단백질은 극성 상호 작용을 통해 친수성 머리와 상호 작용할 수 있지만, 이중층을 완전히 또는 부분적으로 가로지르는 단백질은 소수성 아미노산을 가지고 있어 비극성 지질 내부와 상호 작용한다. 유동 모자이크 모델은 세포막 역학에 대한 정확한 표현을 제공했을 뿐만 아니라, 나중에 생물학적 거대 분자를 설명하는 필수적인 기술적 한계로 발전할 소수성 힘의 연구를 향상시켰다.[9]
수세기 동안 인용된 과학자들은 세포막으로 보이는 구조의 중요성에 대해 의견이 분분했다. 거의 2세기 동안 세포막은 관찰되었지만 세포 기능에서 중요한 구조로 간주되지 않았다. 세포막의 중요성이 인정받은 것은 20세기에 이르러서였다. 마침내 고르터(Gorter)와 그렌델(Grendel)(1925)은 세포막이 "지질 기반"이라는 것을 발견했다. 이를 바탕으로 그들은 이 구조가 층을 모방한 형태를 취해야 한다는 생각을 더욱 발전시켰다. 더 자세히 연구한 결과, 세포 표면의 합과 지질의 표면을 비교하여 2:1의 비율을 추정하여 오늘날 알려진 이중층 구조의 첫 번째 기초를 제공했다. 이 발견은 다양한 과학 분야에서 전 세계적으로 새로운 연구를 시작하여 세포막의 구조와 기능이 널리 받아들여지도록 했다.[9]
2. 2. 세포막 구조 모델의 발전
로버트 훅(Robert Hooke)이 1665년에 세포를 발견한 후 세포 이론이 제기되었다. 당시에는 식물 세포만 관찰할 수 있었기 때문에 처음에는 모든 세포에 단단한 세포벽이 있다고 생각되었다.[9] 19세기 초, 식물 세포가 분리될 수 있다는 사실이 밝혀지면서 세포는 개별적인 독립체이며, 서로 연결되지 않고 각각의 세포벽으로 둘러싸여 있다는 것이 인식되었다. 이 이론은 동물 세포까지 확장되어 세포 보호 및 발달에 대한 보편적인 메커니즘을 제시하였다.19세기 후반에도 현미경 기술은 세포막과 세포벽을 구별하기에는 충분하지 않았다. 그러나 일부 현미경 학자들은 당시에 보이지 않더라도 동물 세포에는 세포막이 존재한다고 파악했고, 세포막이 식물 세포벽과 동일하지 않다는 것을 인지했다. 또한 세포막이 모든 세포에 필수적인 구성 요소는 아니라는 점도 추론되었다. 19세기 말까지도 많은 사람들이 세포막의 존재를 부정했다. 1890년, 세포 이론의 수정을 통해 세포막이 존재하지만 단순히 이차적인 구조일 뿐이라는 주장이 제기되었다. 삼투 및 투과성에 대한 후속 연구를 통해서야 세포막이 더욱 인정받게 되었다.[9] 1895년, 어니스트 오버턴(Ernest Overton)은 세포막이 지질로 구성되어 있다고 제안했다.[10]
1925년, 고르터(Gorter)와 그렌델(Grendel)이 제안한 지질 이중층 가설[11]은 결정학적 연구와 비누방울 관찰을 바탕으로 세포막 이중층 구조에 대한 추측을 만들어냈다. 연구자들은 인간 적혈구에서 지질을 추출하여 물 표면에 퍼뜨렸을 때 지질이 덮는 표면적을 측정했다. 성숙한 포유류 적혈구는 핵과 세포질 소기관이 모두 없기 때문에 세포막은 세포 내 유일한 지질 함유 구조이다. 따라서 세포에서 추출한 모든 지질은 세포의 세포막에 있었다고 가정할 수 있다. 추출된 지질이 덮는 물의 표면적과 지질이 추출된 적혈구의 계산된 표면적의 비율은 약 2:1이었고, 그들은 세포막이 지질 이중층으로 구성되어 있다고 결론지었다.[9][12]
1925년, 프릭(Fricke)은 적혈구와 효모 세포막의 두께가 3.3~4 nm 범위에 있다는 것을 밝혀냈는데, 이는 지질 단층과 호환되는 두께였다. 독립적으로, 레프토스코프(leptoscope)가 발명되어 매우 얇은 막을 측정할 수 있게 되었다. 이 장치는 pH 측정 및 8.6~23.2 nm 범위의 막 단백질 존재 여부에 따라 달라지는 두께를 측정할 수 있었는데, 더 낮은 측정값은 지질 이중층 가설을 뒷받침했다. 1930년대 후반, 휴 데이브슨(Hugh Davson)과 제임스 대니얼리(James Danielli)(1935)의 파우시몰레큘러 모델(paucimolecular model)이 세포막 구조 모델로 일반적으로 받아들여지게 되었다.[13][9] 이 모델은 기름과 극피동물 알 사이의 표면 장력 연구를 기반으로 했다. 지질 이중층이 두 개의 얇은 단백질 층 사이에 있다는 것이 제안되었다. 파우시몰레큘러 모델은 즉시 인기를 얻었고, 세이무어 조너선 싱어(Seymour Jonathan Singer)와 가스 니콜슨(Garth L. Nicolson)(1972)의 유동 모자이크 모델이 나타날 때까지 30년 동안 세포막 연구를 지배했다.[13][9]
유동 모자이크 모델은 1970년대에 처음 제안된 이후로 오랫동안 세포막의 주요 원형으로 남아 있다.[9] 유동 모자이크 모델은 현대의 발견들을 상세히 설명하도록 수정되었지만, 기본 원리는 일관되게 유지되었다. 즉, 세포막은 친수성 외부 머리와 소수성 내부로 구성된 지질 이중층이며, 단백질은 극성 상호 작용을 통해 친수성 머리와 상호 작용할 수 있지만, 이중층을 완전히 또는 부분적으로 가로지르는 단백질은 소수성 아미노산을 가지고 있어 비극성 지질 내부와 상호 작용한다. 유동 모자이크 모델은 세포막 역학에 대한 정확한 표현을 제공했을 뿐만 아니라, 나중에 생물학적 거대 분자를 설명하는 필수적인 기술적 한계로 발전할 소수성 힘의 연구를 향상시켰다.[9]
수세기 동안 인용된 과학자들은 세포막으로 보이는 구조의 중요성에 대해 의견이 분분했다. 거의 2세기 동안 세포막은 관찰되었지만 세포 기능에서 중요한 구조로 간주되지 않았다. 세포막의 중요성이 인정받은 것은 20세기에 이르러서였다. 마침내 고르터(Gorter)와 그렌델(Grendel)(1925)은 세포막이 "지질 기반"이라는 것을 발견했다. 이를 바탕으로 그들은 이 구조가 층을 모방한 형태를 취해야 한다는 생각을 더욱 발전시켰다. 더 자세히 연구한 결과, 세포 표면의 합과 지질의 표면을 비교하여 2:1의 비율을 추정하여 오늘날 알려진 이중층 구조의 첫 번째 기초를 제공했다. 이 발견은 다양한 과학 분야에서 전 세계적으로 새로운 연구를 시작하여 세포막의 구조와 기능이 널리 받아들여지도록 했다.[9]
3. 구성
세포막은 다양한 생체 분자, 특히 지질과 단백질을 포함하고 있다. 세포막의 구성은 고정되어 있지 않고, 유동성과 환경 변화에 따라 끊임없이 변화하며, 세포 발달 단계에 따라서도 변동한다. 특히, 사람의 일차 신경세포 세포막의 콜레스테롤 양은 변화하며, 이러한 구성 변화는 발달 단계 전반에 걸쳐 유동성에 영향을 미친다.[22]
물질은 다양한 메커니즘을 통해 세포막에 통합되거나 제거된다.
- 세포외유출 과정에서 세포 내 소낭이 세포막과 융합되어 소낭의 내용물을 배출하고, 소낭막의 구성 요소를 세포막에 통합한다.
- 세포내이입 과정에서 세포막은 세포외 물질 주위에 세포 돌기를 형성하여 소낭으로 분리될 수 있다.
- 세포막이 막 물질로 만들어진 관상 구조와 연속적인 경우, 관에서 물질이 세포막으로 지속적으로 유입될 수 있다.
- 수용액 상에서 막 구성 요소의 농도는 낮지만, 지질과 수용액 상 사이에서 분자 교환이 일어난다.
세포막에는 당단백질과 일부 당지질(세레브로시드와 갱글리오시드)을 포함한 탄수화물도 포함되어 있다. 탄수화물은 진핵생물에서 세포-세포 인식에 중요한 역할을 한다. 탄수화물은 세포 표면에 위치하여 숙주 세포를 인식하고 정보를 공유한다. 이러한 수용체를 이용하여 세포에 결합하는 바이러스는 감염을 일으킨다.[26]
3. 1. 지질
세포막은 지질과 단백질을 포함한 다양한 생체 분자로 구성되어 있다. 세포막의 구성은 고정되어 있지 않고 유동성과 환경 변화에 따라 끊임없이 변화하며, 세포 발달 단계에 따라서도 변동한다. 특히, 사람의 일차 신경세포 세포막의 콜레스테롤 양은 변화하며, 이러한 구성 변화는 발달 단계 전반에 걸쳐 유동성에 영향을 미친다.[22]물질은 다양한 메커니즘을 통해 세포막에 통합되거나 제거된다.
- 세포외유출 과정에서 세포 내 소낭이 세포막과 융합되어 소낭의 내용물을 배출하고, 소낭막의 구성 요소를 세포막에 통합한다.
- 세포내이입 과정에서 세포막은 세포외 물질 주위에 세포 돌기를 형성하여 소낭으로 분리될 수 있다.
- 세포막이 막 물질로 만들어진 관상 구조와 연속적인 경우, 관에서 물질이 세포막으로 지속적으로 유입될 수 있다.
- 수용액 상에서 막 구성 요소의 농도는 낮지만, 지질과 수용액 상 사이에서 분자 교환이 일어난다.

세포막은 인지질, 당지질, 스테롤의 세 가지 양친매성 지질로 구성된다. 각 지질의 양은 세포의 종류에 따라 다르지만, 대부분 인지질이 가장 풍부하며, 종종 세포막 전체 지질의 50% 이상을 차지한다.[23][24] 당지질은 약 2% 정도의 소량만을 차지하고, 스테롤이 나머지를 구성한다. 적혈구 연구에서는 세포막의 30%가 지질로 이루어져 있으며, 대부분의 진핵세포에서 세포막은 무게 기준으로 지질과 단백질이 각각 절반씩을 차지한다.
인지질과 당지질의 지방산 사슬은 일반적으로 짝수 개의 탄소 원자를 포함하며, 16개에서 20개 사이이다. 16개와 18개의 탄소 원자를 가진 지방산이 가장 흔하다. 지방산은 포화 또는 불포화될 수 있으며, 이중 결합의 배열은 거의 항상 "cis"형이다. 지방산 사슬의 길이와 불포화 정도는 막 유동성에 큰 영향을 미치는데, 불포화 지질은 굽은 모양을 만들어 지방산이 서로 밀접하게 붙는 것을 방지하여 막의 녹는 온도를 낮추고(유동성을 증가시키고) 결과적으로 막 유동성에 영향을 준다.[23][24] 지질 조성을 변화시킴으로써 세포막의 유동성을 조절하는 일부 생물의 능력을 항온적응이라고 한다.
전체 막은 소수성 꼬리의 비공유 결합 상호 작용을 통해 유지되지만, 구조는 매우 유동적이며 단단히 고정되어 있지 않다. 생리적 조건하에서 세포막의 인지질 분자는 액정 상태에 있다. 즉, 지질 분자는 자유롭게 확산되고 존재하는 층을 따라 빠른 측면 확산을 나타낸다.[23] 그러나 이중층의 세포 내 및 세포 외 소엽 사이에서 인지질 분자의 교환은 매우 느린 과정이다. 지질 뗏목과 카베올라는 세포막에서 콜레스테롤이 풍부한 미세 영역의 예이다.[24] 또한, 적분 막 단백질과 직접 접촉하는 지질의 일부는 단백질 표면에 단단히 결합되어 있으며, 환상 지질 껍질이라고 불리며 단백질 복합체의 일부로 작용한다.
콜레스테롤은 일반적으로 세포막 전체에 다양한 정도로 분산되어 있으며, 막 지질의 소수성 꼬리 사이의 불규칙적인 공간에 존재하여 막에 강성과 강화 효과를 부여한다.[4] 생물학적 막에서 콜레스테롤의 양은 생물, 세포 유형, 심지어 개별 세포에 따라 다르다. 콜레스테롤은 전체 막의 유동성을 조절하며, 농도에 따라 다양한 세포막 구성 요소의 이동량을 제어한다.[4] 고온에서는 콜레스테롤이 인지질 지방산 사슬의 이동을 억제하여 작은 분자의 투과성과 막 유동성을 감소시킨다. 저온에서는 콜레스테롤의 역할은 그 반대이다. 콜레스테롤 생산은 저온에 대한 반응으로 증가한다. 저온에서는 콜레스테롤이 지방산 사슬 상호 작용을 방해한다. 콜레스테롤은 부동액 역할을 하여 막의 유동성을 유지한다. 콜레스테롤은 따뜻한 지역 동물보다 추운 지역 동물에서 더 풍부하다. 콜레스테롤이 없는 식물에서는 피토스테롤이라고 하는 관련 화합물이 콜레스테롤과 같은 기능을 수행한다.[4]
3. 1. 1. 지질 소포 (리포솜)
지질 소낭(리포솜)은 지질 이중층으로 둘러싸인 거의 구형의 주머니 모양 구조이다.[25] 이러한 구조는 세포에 직접적으로 화학 물질을 전달하여 세포에 미치는 영향을 조사하거나, 세포막의 투과성에 대한 보다 자세한 지식을 얻는 등의 목적으로 실험실에서 사용된다. 지질 소낭과 리포솜은 먼저 지질을 수용액에 현탁하고, 그 혼합물을 초음파 처리로 교반하여 얻을 수 있다.[69] 소낭 내부에서 주변 용액으로의 유출 속도를 측정함으로써 막 투과성에 대한 이해를 높일 수 있게 되었다. 내부에 분자나 이온을 포함하는 소낭은 목적 물질을 포함하는 용액에서 소낭을 형성함으로써 얻을 수 있다. 또한 단백질도 목적 단백질을 계면활성제 존재 하에 가용화시켜 리포솜을 형성하는 인지질과 결합시킴으로써 막에 삽입할 수 있다. 이러한 기술을 통해 연구자들은 다양한 막 단백질의 기능을 조사할 수 있게 되었다.3. 2. 단백질
세포막은 다량의 단백질을 함유하고 있으며, 일반적으로 막 부피의 약 50%를 차지한다.[27] 이러한 단백질은 다양한 생물학적 활동을 담당하기 때문에 세포에 중요하다. 효모의 유전자 약 3분의 1이 특히 단백질을 암호화하며, 다세포 생물에서는 이 수치가 더 높다.[25]막 단백질은 막관통 단백질, 주변 단백질, 지질 연결 단백질의 세 가지 주요 유형으로 구성된다.[4]
| 유형 | 설명 | 예시 |
|---|---|---|
| 막관통 단백질 | 세포막을 관통하며, 내부 분자와 상호 작용하는 친수성 세포질 도메인, 세포막 내에 고정하는 소수성 막 관통 도메인, 그리고 외부 분자와 상호 작용하는 친수성 세포외 도메인을 갖는다. 소수성 도메인은 하나, 여러 개 또는 α-나선과 β-병풍 단백질 모티프의 조합으로 구성된다. | 이온 채널, 양성자 펌프, G 단백질 연결 수용체 |
| 지질 연결 단백질 | 하나 또는 여러 개의 지질 분자에 공유 결합되어 있으며, 소수성으로 세포막에 삽입되어 단백질을 고정한다. 단백질 자체는 막과 접촉하지 않는다. | G 단백질 |
| 주변 단백질 | 막관통 단백질에 부착되거나 지질 이중층의 주변 영역과 관련이 있다. 이러한 단백질은 생물학적 막과 일시적인 상호 작용을 하는 경향이 있으며, 반응이 일어나면 세포질에서 작용을 계속하기 위해 분리된다. | 일부 효소, 일부 호르몬 |
막관통 단백질의 예로는 이온 채널, 양성자 펌프, G 단백질 연결 수용체가 있다. 이온 채널은 나트륨, 칼륨, 칼슘, 염소와 같은 무기 이온이 막을 가로지르는 친수성 기공을 통해 전기화학적 기울기에 따라 지질 이중층을 확산하도록 허용한다. 신경 세포와 같은 세포의 전기적 거동은 이온 채널에 의해 조절된다.[4] 양성자 펌프는 지질 이중층에 내장된 단백질 펌프로, 양성자가 한 아미노산 측쇄에서 다른 아미노산 측쇄로 이동하여 막을 통과하도록 한다. 전자 전달 및 ATP 생성과 같은 과정은 양성자 펌프를 사용한다.[4] G 단백질 연결 수용체는 지질 이중층을 일곱 번 가로지르는 단일 폴리펩타이드 사슬로, 신호 분자 (예: 호르몬 및 신경 전달 물질)에 반응한다. G 단백질 연결 수용체는 세포 간 신호 전달, cAMP 생성 조절, 이온 채널 조절과 같은 과정에 사용된다.[4]
외부 환경에 노출된 세포막은 세포 간 통신의 중요한 부위이다. 따라서 항원과 같은 다양한 단백질 수용체와 식별 단백질이 세포막 표면에 존재한다. 막 단백질의 기능에는 세포 접촉, 표면 인식, 세포 골격 접촉, 신호 전달, 효소 활성 또는 막을 가로지르는 물질 수송이 포함될 수 있다.
대부분의 막 단백질은 어떤 방식으로든 막에 삽입되어야 한다.[28] 이를 위해 아미노산의 N 말단 "신호 서열"이 단백질을 소포체로 안내하여 지질 이중층에 단백질을 삽입한다. 삽입되면 단백질은 소포로 최종 목적지로 운반되고, 소포는 표적 막과 융합된다.
3. 3. 탄수화물
세포막에는 탄수화물도 포함되어 있는데, 주로 당단백질이지만 일부 당지질(세레브로시드와 갱글리오시드)도 포함되어 있다. 탄수화물은 진핵생물에서 세포-세포 인식에 중요한 역할을 한다. 탄수화물은 세포 표면에 위치하여 숙주 세포를 인식하고 정보를 공유한다. 이러한 수용체를 이용하여 세포에 결합하는 바이러스는 감염을 일으킨다.[26] 대부분의 경우 세포 내 막에서는 당쇄 변형이 일어나지 않고, 일반적으로는 세포막의 세포외 표면에서 당쇄 변형이 일어난다. 글리코칼릭스는 모든 세포, 특히 미세융모가 있는 상피세포에서 중요한 특징이다. 최근 자료에 따르면 글리코칼릭스는 세포 부착 등 여러 가지 과정에 참여하는 것으로 나타났다.[26] 골지체에서 당쇄 변형이 이루어지기 때문에, 끝에서 두 번째 당은 갈락토스, 끝의 당은 시알산이다.[71] 시알산은 음전하를 띠고 있으며, 전하를 띤 입자에 대한 외부 장벽이 된다.4. 구조
세포막은 인지질 이중층이라는 두 겹의 인지질로 구성되어 있으며, '유동 모자이크' 구조로 설명된다. 유동 모자이크란 지질이 자유롭게 떠다니는 2차원적인 흐름을 의미하며, 내부에는 세포막을 가로지르는 통로 혹은 수용체로 작용하는 단백질을 가지고 있다.
세포는 세포막의 유동성을 유지하기 위해서 지질의 종류 및 양을 다양하게 변화시킨다. 이중층 내부에 있는 콜레스테롤 분자(진핵세포의 경우)나 호파노이드(원핵세포의 경우)는 유동성을 유지시켜주는 데 도움을 준다.
세포골격은 세포막을 지지하면서 세포막 내부 단백질을 고정시키고, 특정한 면으로 향하게 해주며, 또한 단백질이 세포막 내부에서 움직일 수 있는 범위를 제한해준다. 세포막 표면은 항상 형태 없이 움직이는 유체가 아니라, 어느 정도의 구조를 보여준다. 시냅스는 보다 구조적인 세포막의 예이다.
세포막은 많은 단백질을 포함하고 있으며, 일반적으로 막 부피의 50%를 차지한다.[72] 이러한 단백질은 다양한 생물학적 활성을 담당하며 중요하다. 막 단백질은 내재성, 외재성, 지질 앵커 단백질이라는 세 가지 주요 유형으로 구성된다.[54]
| 유형 | 설명 | 예시 |
|---|---|---|
| 내재성 막 단백질 (막관통 단백질) | 막을 관통하여 존재하며, 친수성 세포질 도메인을 가지고 세포 내 분자와 상호 작용한다. 소수성 막관통 도메인은 세포막 내부에 존재하며, 친수성 세포외 도메인은 세포 외부의 분자와 상호 작용한다. 소수성 도메인은 α-나선 또는 β-시트로 구성된 단백질 모티프 하나 또는 여러 개, 또는 이들의 조합으로 구성된다. | 이온 채널, 양성자 펌프, G 단백질 연결 수용체 |
| 지질 앵커형 단백질 | 하나 또는 여러 개의 지질 분자에 공유 결합되어 있으며, 이들을 세포막에 소수성으로 삽입함으로써 단백질을 고정시킨다. 단백질 자체는 막과 반드시 상호 작용하는 것은 아니다. | G 단백질 |
| 외재성 막 단백질 | 내재성 막 단백질이나 지질 이중층의 주변 영역과 상호 작용한다. 이러한 단백질은 생체막과 일시적인 상호 작용만 하는 경향이 있으며, 일단 반응하면 분자는 해리되어 세포질로 이동하여 기능을 수행한다. | 일부 효소, 호르몬 |
세포막은 외부 환경에 노출되어 있으며, 세포 간 의사소통에 중요한 부위이다. 따라서 다양한 종류의 단백질 수용체와 항원 제시를 수행하는 단백질 등이 세포막 표면에 존재한다. 막 단백질의 기능에는 세포 간 연결, 표면 인식, 세포 골격 연결, 신호 전달, 효소 활성, 막을 가로지른 물질 수송 등이 포함된다.
4. 1. 유동 모자이크 모델
세포막은 인지질 이중층이라는 두 겹의 인지질로 구성되어 있으며, '유동 모자이크' 구조로 설명된다. 유동 모자이크란 지질이 자유롭게 떠다니는 2차원적인 흐름을 의미하며, 내부에는 세포막을 가로지르는 통로 혹은 수용체로 작용하는 단백질을 가지고 있다. 이러한 세포막 모형은 1971년 S.J. 싱어(S.J. Singer)가 지질 단백질 모형을 제안하고, 1972년 G.L. 니콜슨(G.L. Nicolson)과 함께 유동 특성을 제시하며 탄수화물이 붙어 있다고 하였다.[38] 세포는 세포막의 유동성을 유지하기 위해서 지질의 종류 및 양을 다양하게 변화시킨다. 이중층 내부에 있는 콜레스테롤 분자(진핵세포의 경우)나 호파노이드(원핵세포의 경우)는 유동성을 유지시켜주는 데 도움을 준다.세포골격은 세포막을 지지하면서 세포막 내부 단백질을 고정시키고, 특정한 면으로 향하게 해주며, 또한 단백질이 세포막 내부에서 움직일 수 있는 범위를 제한해준다. 세포막 표면은 항상 형태 없이 움직이는 유체가 아니라, 어느 정도의 구조를 보여준다. 시냅스는 보다 구조적인 세포막의 예이다.
싱어와 니콜슨이 1972년에 제시한 유동 모자이크 모델에 따르면, 생물학적 막은 지질 및 단백질 분자가 다소 자유롭게 확산되는 2차원 액체로 간주될 수 있다.[38] 막의 기초를 이루는 지질 이중층은 그 자체로 2차원 액체를 형성하지만, 세포막은 많은 양의 단백질을 포함하고 있으며, 이는 더 많은 구조를 제공한다. 이러한 구조의 예로는 단백질-단백질 복합체, 액틴 기반 세포골격에 의해 형성된 울타리, 그리고 잠재적으로 지질 뗏목이 있다.
4. 2. 지질 이중층
세포막은 인지질 이중층이라는 두 겹의 인지질로 구성되어 있으며, ''유동 모자이크'' 구조로 설명된다. 유동 모자이크란 지질이 자유롭게 떠다니는 2차원적인 흐름을 의미하며, 내부에는 세포막을 가로지르는 통로 혹은 수용체로 작용하는 단백질을 가지고 있다. 이러한 세포막 모형은 1971년 S.J. Singer가 지질 단백질 모형을 제안하고, 1972년 G.L. Nicolson과 함께 유동 특성을 제시하며 탄수화물이 붙어 있다고 하였다. 세포는 세포막의 유동성을 유지하기 위해서 지질의 종류 및 양을 다양하게 변화시킨다. 이중층 내부에 있는 콜레스테롤 분자(진핵세포의 경우)나 호파노이드(원핵세포의 경우)는 유동성을 유지시켜주는 데 도움을 준다.인지질 이중층은 자가 조립 과정을 통해 형성된다. 세포막은 주로 얇은 양친매성 인지질 층으로 구성되어 있으며, 이는 자발적으로 배열되어 소수성 "꼬리" 부분이 주변 물로부터 분리되고 친수성 "머리" 부분은 생성된 이중층의 세포 내(세포질) 및 세포 외 표면과 상호 작용하도록 한다. 이것은 연속적인 구형 인지질 이중층을 형성한다. 소수성 효과는 인지질 이중층 형성의 주요 원동력이다. 소수성 분자 사이의 상호 작용이 증가하면(소수성 영역의 밀집을 유발) 물 분자가 서로 더 자유롭게 결합하여 시스템의 엔트로피가 증가한다. 이러한 복잡한 상호 작용에는 반데르발스 힘, 정전기적 상호 작용 및 수소 결합과 같은 비공유 결합이 포함될 수 있다.
인지질 이중층은 일반적으로 이온과 극성 분자에 대해 불투과성이다. 인지질 이중층의 친수성 머리와 소수성 꼬리의 배열은 극성 용질(예: 아미노산, 핵산, 탄수화물, 단백질 및 이온)이 막을 가로질러 확산되는 것을 방지하지만, 일반적으로 소수성 분자의 수동 확산을 허용한다. 이는 세포가 기공, 채널 및 게이트와 같은 막관통 단백질 복합체를 통해 이러한 물질의 이동을 제어할 수 있도록 한다. 플립파제와 스크램블라제는 음전하를 띠는 포스파티딜세린을 내막에 집중시킨다. NANA와 함께 이것은 막을 통과하는 하전된 기의 이동에 대한 추가적인 장벽을 만든다.
일부 고세균의 세포막은 인지질 이중층이 아닌 예외적인 경우인데, 예를 들어 술폴로부스속(''Sulfolobus'') 등에서는 마주 보는 인지질의 소수성 사슬이 연결되어 인지질 단일층이 된다.
4. 3. 막 단백질
세포막은 많은 단백질을 포함하고 있으며, 일반적으로 막 부피의 50%를 차지한다.[72] 이러한 단백질은 다양한 생물학적 활성을 담당하며 중요하다. 효모에서는 약 3분의 1의 유전자가 막 단백질을 암호화하고 있으며, 이 비율은 다세포 생물에서 더 높아진다.[68] 막 단백질은 내재성, 외재성, 지질 앵커 단백질이라는 세 가지 주요 유형으로 구성된다.[54]| 유형 | 설명 | 예시 |
|---|---|---|
| 내재성 막 단백질 (막관통 단백질) | 막을 관통하여 존재하며, 친수성 세포질 도메인을 가지고 세포 내 분자와 상호 작용한다. 소수성 막관통 도메인은 세포막 내부에 존재하며, 친수성 세포외 도메인은 세포 외부의 분자와 상호 작용한다. 소수성 도메인은 α-나선 또는 β-시트로 구성된 단백질 모티프 하나 또는 여러 개, 또는 이들의 조합으로 구성된다. | 이온 채널, 양성자 펌프, G 단백질 연결 수용체 |
| 지질 앵커형 단백질 | 하나 또는 여러 개의 지질 분자에 공유 결합되어 있으며, 이들을 세포막에 소수성으로 삽입함으로써 단백질을 고정시킨다. 단백질 자체는 막과 반드시 상호 작용하는 것은 아니다. | G 단백질 |
| 외재성 막 단백질 | 내재성 막 단백질이나 지질 이중층의 주변 영역과 상호 작용한다. 이러한 단백질은 생체막과 일시적인 상호 작용만 하는 경향이 있으며, 일단 반응하면 분자는 해리되어 세포질로 이동하여 기능을 수행한다. | 일부 효소, 호르몬 |
내재성 막 단백질은 양친매성 막관통 단백질이다. 여기에는 이온 채널, 양성자 펌프, G 단백질 연결 수용체 등이 포함된다. 이온 채널을 통해 나트륨, 칼륨, 칼슘, 염소 등의 무기 이온이 전기화학적 기울기에 따른 확산이 일어난다. 이온 채널의 친수성 구멍을 통해 이온이 세포막을 통과한다. 세포의 전기적 거동(신경 세포 등)은 이온 채널에 의해 조절된다.[54] 양성자 펌프는 지질 이중층에 내장된 단백질 펌프이며, 양성자는 아미노산의 측쇄를 차례로 이동하면서 막을 가로질러 이동한다. 이러한 전자 전달 및 ATP 생산에는 양성자 펌프가 사용된다.[54] G 단백질 연결 수용체는 지질 이중층을 7번 관통하는 하나의 폴리펩타이드 사슬로, 신호 전달 물질(호르몬이나 신경전달물질)에 반응한다. G 단백질 연결 수용체는 세포 간 신호 전달, cAMP 생산 조절, 이온 채널 조절 등에 사용된다.[54]
세포막은 외부 환경에 노출되어 있으며, 세포 간 의사소통에 중요한 부위이다. 따라서 다양한 종류의 단백질 수용체와 항원 제시를 수행하는 단백질 등이 세포막 표면에 존재한다. 막 단백질의 기능에는 세포 간 연결, 표면 인식, 세포 골격 연결, 신호 전달, 효소 활성, 막을 가로지른 물질 수송 등이 포함된다.
막 단백질은 여러 가지 방법으로 막에 삽입된다. 한 가지 방법으로, N 말단 아미노산의 "신호 서열"이 단백질을 소포체로 향하게 하며, 거기서 지질 이중층에 삽입된다. 삽입된 단백질은 소포체 내에서 최종 목적지까지 수송되며, 거기서 소포체는 표적 막과 융합한다.[73]
4. 4. 세포골격과의 연결
세포골격은 세포질 내 세포막 아래에 존재하며, 세포막 단백질이 고정될 수 있는 지지체 역할을 하고 세포에서 돌출되는 세포소기관을 형성한다.[40] 사실, 세포골격 요소는 세포막과 광범위하고 밀접하게 상호 작용한다. 고정 단백질은 단백질을 특정 세포 표면(예: 척추동물 장관을 따라 늘어선 상피세포의 첨단부)에 제한하고, 이중층 내에서 확산되는 정도를 제한한다. 세포골격은 섬모와 같이 세포막으로 덮인 미세소관 기반 돌출부와, 액틴 기반 돌출부인 필로포디아와 같은 부속기관 모양의 세포소기관을 형성할 수 있다. 이러한 돌출부는 세포막으로 싸여 있으며, 세포 표면에서 돌출하여 외부 환경을 감지하거나 기질 또는 다른 세포와 접촉하기 위해 존재한다. 상피세포의 첨단부에는 미세융모라고 알려진 액틴 기반의 손가락 모양 돌출부가 밀집되어 있으며, 이는 세포 표면적을 증가시켜 영양소 흡수율을 높인다. 세포골격과 세포막의 국소적 분리는 블레브(bleb)를 형성한다.4. 5. 세포 내 막 구조
세포막 내부의 세포 내용물은 세포의 전체 기능에 기여하는 수많은 세포 소기관으로 구성되어 있다. 각 소기관의 기원, 구조 및 기능은 각 소기관의 고유한 특성으로 인해 세포 구성에 큰 차이를 보인다.- 미토콘드리아와 엽록체는 세포내 공생설로 알려진 박테리아에서 진화한 것으로 여겨진다. 이 이론은 ''파라코쿠스''와 ''로도슈도모나스''와 같은 박테리아가 미토콘드리아와 유사한 기능을 공유하고, 남세균(시아노박테리아)이 엽록체와 유사한 기능을 공유한다는 생각에서 비롯되었다. 세포내 공생설은 진화 과정을 통해 진핵생물 세포가 이 두 종류의 박테리아를 흡수하여 진핵세포 내부에 미토콘드리아와 엽록체가 형성되었다는 것을 제안한다. 이러한 흡수는 이러한 세포 소기관의 이중막 시스템으로 이어졌는데, 여기서 외막은 숙주 세포의 세포막에서 유래하고 내막은 내공생체의 세포막이었다. 미토콘드리아와 엽록체 모두 자신의 DNA를 포함하고 있다는 점은 이 두 소기관 모두 진핵세포 내부에서 번성하는 흡수된 박테리아에서 진화했다는 것을 더욱 뒷받침한다.[41]
- 진핵세포에서 핵막은 핵의 내용물을 세포질과 분리한다.[42] 핵막은 내막과 외막으로 구성되어 핵 안팎으로 물질의 엄격한 조절을 제공한다. 물질은 핵막의 핵공을 통해 세포질과 핵 사이를 이동한다. 세포의 핵이 전사에서 더 활동적인 경우 핵막에 더 많은 핵공이 있다. 많은 단백질이 확산을 통해 핵공을 통과할 수 없기 때문에 핵의 단백질 구성은 세포질과 크게 다를 수 있다. 핵막 내에서 내막과 외막의 단백질 구성은 다르며, 외막만 소포체(ER) 막과 연속적이다. ER과 마찬가지로 외막에는 두 막 사이의 공간으로 단백질을 생산하고 수송하는 역할을 하는 리보솜도 있다. 핵막은 유사분열 초기 단계에서 분해되고 유사분열 후기 단계에서 재조립된다.[43]
- 세포의 전체 막 함량의 상당 부분을 차지하는 세포내막계의 일부인 소포체(ER)은 관과 주머니로 구성된 폐쇄된 네트워크이며, 주요 기능은 단백질 합성과 지질 대사이다. ER에는 매끄러운 ER과 거친 ER의 두 가지 유형이 있다. 거친 ER에는 단백질 합성에 사용되는 리보솜이 부착되어 있고, 매끄러운 ER은 세포 내 독소 처리와 칼슘 조절에 더 많이 사용된다.[44]
- 골지체는 두 개의 상호 연결된 둥근 골지 시스터나를 가지고 있다. 골지체의 구획은 조직, 스택 연결 및 화물 수송을 담당하는 여러 관상-망상 네트워크를 형성하며, 50~60nm에 이르는 포도알처럼 연속적인 일련의 소포를 나타낸다. 골지체는 편평한 원반 모양의 시스터나와 관상-망상 네트워크 및 소포의 세 가지 주요 구획으로 구성된다.[45]
5. 기능
세포막은 세포질을 둘러싸고 세포 내 구성 요소를 세포 외 환경과 분리하는 중요한 역할을 한다. 균류, 세균, 대부분의 고세균, 식물은 세포벽을 추가로 가지는데, 이는 세포에 기계적 지지력을 제공하고 큰 분자의 통과를 막는다.
세포막은 선택적 투과성을 가지며, 세포 안팎으로의 물질 출입을 조절하여 생존에 필수적인 물질의 막 수송을 돕는다. 막을 통한 물질 이동은 수동 수송과 능동 수송으로 나뉜다. 또한 세포막은 막전위를 유지하여 특정 물질만 선택적으로 통과시키는 필터 역할을 한다. 세포는 다음과 같은 다양한 수송 메커니즘을 이용한다.
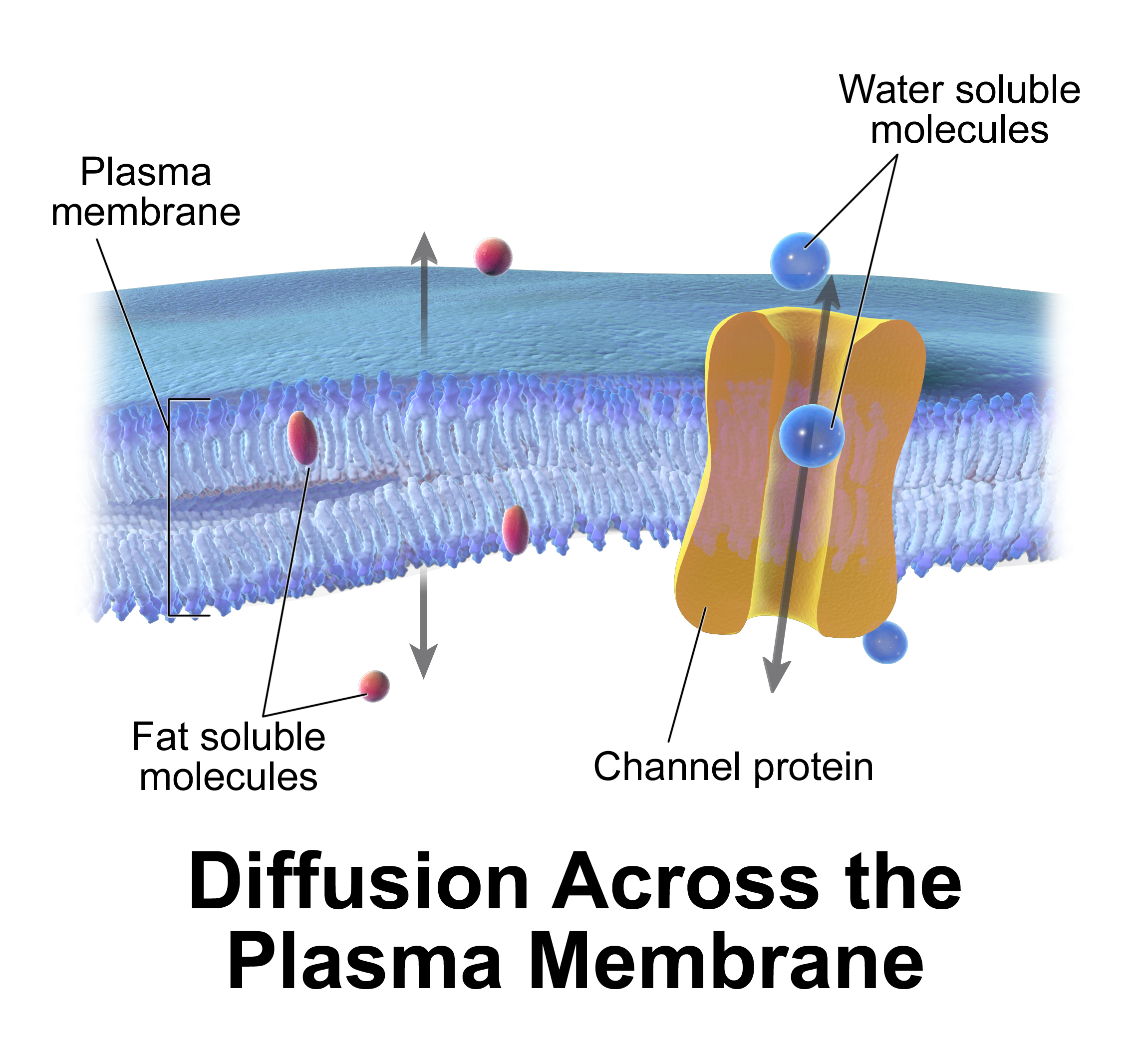
- 수동 삼투 및 확산: 이산화탄소(CO2), 산소(O2)와 같은 일부 물질은 확산에 의해 세포막을 통과한다. 막은 특정 물질이나 이온에 대한 장벽 역할을 하여 막 양쪽에 농도 차이를 만든다. 확산은 작은 분자나 이온이 고농도에서 저농도로 이동하는 현상으로, 에너지 소모 없이 농도 기울기에 의해 일어난다.[29] 반투과성 막을 경계로 한 농도 기울기는 물의 삼투 현상을 일으킨다. 생체 내 삼투는 용매가 반투과성 막을 통과하지만, 농도 기울기에 따라 이동하고 에너지를 소모하지 않으므로 수동적이다. 세포 내 용매는 주로 물이지만, 다른 액체나 초임계유체, 기체일 수도 있다.[30]
- 막관통 채널 및 수송체: 막관통 단백질은 막 양쪽에서 기능하며 분자의 막 통과를 돕는다.[31] 당이나 아미노산과 같은 영양소는 세포로 들어가야 하고, 특정 대사산물은 세포 밖으로 배출되어야 한다. 이들은 아쿠아포린과 같은 단백질 채널을 통해 촉진 확산되거나, 막관통 수송체에 의해 흡수된다. 투과 효소라고도 불리는 단백질 채널은 대개 매우 특이적이어서 제한된 종류의 화학 물질만 인식하고 수송한다. 막관통 단백질의 다른 예로는 세포 표면 수용체가 있으며, 신호 전달 물질을 이용해 세포 간 소통을 한다.[31]
- '''세포내이입''': 세포가 분자를 감싸 흡수하는 과정이다. 세포막은 함입(invagination)이라는 작은 변형을 안쪽으로 만들어 수송될 물질을 포획한다. 함입은 세포막 바깥쪽 단백질에 의해 일어나며, 이들은 수용체처럼 기능하여 함몰부에 모여 세포막 안쪽으로 단백질이나 지질 축적을 촉진한다.[32] 이후 변형 부분은 세포 안쪽으로 잘록해져 막에서 분리되고, 포획된 물질을 포함하는 소포를 생성한다. 식작용, 음세포작용 등의 경로가 있으며 에너지를 필요로 하는 능동 수송의 한 형태이다.
- '''세포외유출''': 소포 막이 세포막과 융합하여 내용물을 외부로 배출하는 과정이다. 엑소사이토시스는 엔도사이토시스로 흡수된 물질의 미소화 잔여물을 제거하거나, 호르몬, 효소 등을 분비하며, 물질을 세포 장벽 너머로 수송한다. 엑소사이토시스 과정에서 골지체에서 생성된 분비 소포 등은 세포골격에 의해 세포 표면으로 이동, 소포 막과 세포막이 접촉한다. 두 지질 이중층의 지질 분자는 서로 재배열되고, 두 막은 융합, 통로가 형성되고, 소포는 내용물을 세포 외부로 배출한다.

세포막은 세포골격을 지지하여 세포 형태를 유지하고, 세포외기질 및 다른 세포와 부착하여 조직을 형성한다. 세포막은 다양한 "초막" 구조를 형성하고 세포 연접을 통해 다른 세포와 연결되어, 세포 부착, 세포 간 통신, 세포내이입, 세포외유출을 담당한다.
세포막에는 다양한 종류의 막 단백질이 존재하며, 이들은 세포 간 연결, 표면 인식, 세포골격 연결, 신호전달, 효소 활성, 막을 가로지른 물질 수송 등 다양한 기능을 수행한다.
5. 1. 물질 수송
세포막은 반투과성을 지니는데, 이는 일부 분자만이 세포 내외로 수송될 수 있음을 의미한다. 통과 가능한 분자는 크기가 작거나 소수성이어야 한다. 그렇지 않은 대부분의 분자는 통과를 매개하는 각각의 막단백질이 세포막에 있을 경우에만 통과할 수 있다.분자에 따라, 수송은 다른 방식으로 일어나며, 크게 아데노신 삼인산(ATP) 형태로 존재하는 에너지를 소모하는 능동수송과 그렇지 않은 수동수송의 두 가지로 나눌 수 있다.
수동수송은 물질이 세포막을 통과할 때 아데노신 삼인산 등의 화학 에너지를 소모하지 않는 수송이다. 소수성(비극성) 혹은 작은 극성 분자, 그리고 가스는 막단백질의 매개가 필요없는 확산을 통해 운반되며, 극성을 띠거나 전하를 띤 작은 분자들 그리고 이온들은 각각에 대한 막단백질이 매개하는 촉진확산의 방식으로 운반된다. 물분자는 확산의 형태로도 세포막을 통과하지만, 대부분 아쿠아포린이라는 막단백질을 통해 삼투의 형태로 운반된다. 농도 기울기를 따라서 이동의 방향이 결정된다 (엄밀하게 말하면 양쪽 방향으로 이동이 일어나지만 총체적으로 농도가 높은 쪽에서 낮은 쪽으로 알짜 이동이 일어난다). 이온이나 전하를 띤 분자의 이동 방향은 단순한 농도 기울기뿐만 아니라 세포막을 중심으로 형성된 세포안과 밖의 전위차에 의해서도 영향을 받는다.
능동수송 과정에서 분자는 일반적으로 농도 기울기에 반해서 수송되며, 이는 엔트로피 법칙에 어긋나는 것이다. 이러한 농도 경사에 역행해서 이동이 일어나려면 아데노신 삼인산(ATP)의 가수분해에 의한 에너지가 필요하다. 두 가지의 능동수송이 가능한데, 하나는 운반체가 직접 아데노신 삼인산을 가수분해하며 이때 발생하는 에너지를 이용하여 수송하는 것으로서 1차 능동수송이라고 한다. 두 번째 방법은 다른 분자(주로 소듐 펌프)가 아데노신 삼인산을 가수분해할 때 나온 에너지를 이용하는 방법으로서 2차 능동수송이라고 한다. 소듐펌프는 아데노신 삼인산의 분해 에너지를 이용하여 소듐은 세포밖으로 퍼내고 포타슘은 세포 안으로 이동시킨다. 그러면 2차 능동 수송 막단백질들은 세포막을 경계로 형성된 소듐의 농도 기울기라는 화학적에너지를 이용하여 각자의 분자나 이온들을 농도 기울기에 역행해서 이동시킨다. 이 두 분자의 이동 방향은 같은 방향일수도 다른 방향일수도 있다.
세포막은 세포질을 둘러싸고 있으며, 세포 내 구성 요소를 세포 외 환경으로부터 물리적으로 분리한다. 또한, 세포막은 세포골격의 지지대가 되어 세포의 형태를 유지하고, 세포외기질 및 다른 세포와 부착하여 조직을 형성한다.
세포막은 선택적 투과성을 가지고 있으며, 세포의 출입을 조절하고, 생존에 필요한 물질의 수송을 촉진한다. 막을 통한 물질 이동은 세포의 에너지 투입 없이 일어나는 수동수송과 수송을 위해 세포가 에너지를 소비해야 하는 능동수송 중 하나에 의해 수행된다. 또한, 막은 막전위를 유지한다. 세포막은 선택적 필터 역할을 하여 특정 물질만 세포 안팎으로 출입할 수 있도록 한다. 세포는 생체막이 관여하는 다음과 같은 수송 메커니즘을 이용한다.
# 수동적인 삼투와 확산: 이산화탄소(CO2)나 산소(O2)와 같은 몇몇 물질은 확산에 의해 세포막을 통과할 수 있으며, 이는 수동적인 수송 과정이다. 막은 특정 물질이나 이온에 대한 장벽 역할을 하기 때문에, 막의 양쪽에는 다른 농도가 형성된다. 확산은 저분자나 이온이 고농도 쪽에서 저농도 쪽으로 자유롭게 이동할 때 발생한다. 이 과정은 에너지를 필요로 하지 않고, 막의 양쪽에 형성된 농도 기울기에 의해 추진되므로 수동 수송으로 간주된다.[29] 반투과성 막을 사이에 둔 농도 기울기는 물의 삼투류를 유발한다. 생체 내 삼투는 용매의 반투과성 막 통과를 수반하지만, 용매는 농도 기울기에 따라 이동하고 에너지를 필요로 하지 않으므로 마찬가지로 수동적이다. 세포 내 용매는 물이 가장 일반적이지만, 다른 액체나 초임계유체, 기체일 수도 있다.[30]
# 막관통 채널과 수송체: 막관통 단백질은 분자의 막 통과를 위해 막의 양쪽에서 기능한다.[31] 당이나 아미노산과 같은 영양소는 세포로 들어갈 필요가 있고, 특정 대사산물은 세포 밖으로 나갈 필요가 있다. 이러한 분자들은 아쿠아포린과 같은 단백질 채널을 수동적으로 통과하여 촉진 확산되거나, 막관통 수송체에 의해 막을 넘어 흡수된다. 단백질 채널은 투과 효소(permease)라고도 불리며, 일반적으로 매우 특이성이 높아 제한된 종류의 화학 물질만 (한 종류의 물질로 제한되는 경우도 많음) 인식하여 수송한다. 막관통 단백질의 다른 예로는 세포 표면의 수용체가 있으며, 신호 전달 물질을 이용하여 세포 간 의사소통을 한다.[31]
# 엔도사이토시스: 엔도사이토시스는 세포가 분자를 감싸서 흡수하는 과정이다. 세포막은 함입(invagination)이라고 불리는 안쪽으로의 작은 변형을 만들어내고, 수송될 물질이 그곳에 포착된다. 이 함입은 세포막 바깥쪽에 위치한 단백질에 의해 일어나며, 그들은 수용체처럼 기능하여 함몰부에 모여 세포막 안쪽으로의 단백질이나 지질의 축적을 촉진한다.[32] 그 후, 변형 부분은 세포 안쪽으로 잘록해져 막으로부터 분리되고, 포착된 물질을 안쪽에 포함하는 소포가 생성된다. 엔도사이토시스는 고체 입자를 흡수하거나(식작용) 저분자나 이온(음작용), 고분자를 흡수하는 경로이다. 엔도사이토시스는 에너지를 필요로 하므로 능동 수송의 한 형태이다.
# 엑소사이토시스: 함입이나 소포 형성에 의해 세포에 물질이 흡수되는 것과 마찬가지로, 소포의 막은 세포막과 융합하여 내용물을 외부로 배출할 수 있다. 이것이 엑소사이토시스 과정이다. 엑소사이토시스는 엔도사이토시스에 의해 흡수된 물질의 미소화 잔여물을 제거하거나, 호르몬이나 효소와 같은 물질을 분비하거나, 물질을 세포의 장벽을 넘어 수송하는 등의 목적으로 다양한 세포에서 수행된다. 엑소사이토시스 과정에서 미소화 잔여물을 포함하는 식포나 골지체에서 출아된 분비소포는 먼저 세포골격에 의해 세포 내부에서 표면으로 이동하여 소포의 막과 세포막이 접촉하게 된다. 두 지질 이중층의 지질 분자는 서로 재배열되고, 두 막은 융합한다. 막의 융합에 의해 통로가 형성되고, 소포는 그 내용물을 세포 외부로 배출한다.
5. 2. 세포 신호 전달
세포막은 세포질을 둘러싸고 있으며, 세포 내 구성 요소를 세포 외 환경으로부터 물리적으로 분리한다. 또한, 세포막은 세포골격의 지지대가 되어 세포의 형태를 유지하고, 세포외기질 및 다른 세포와 부착하여 조직을 형성한다. 균류, 세균, 대부분의 고세균, 그리고 식물은 세포벽도 가지고 있으며, 세포의 기계적인 지지 역할을 하고, 거대 분자의 통과를 막는다.세포막은 선택적 투과성을 가지고 있으며, 세포의 출입을 조절하고, 생존에 필요한 물질의 수송을 촉진한다. 막을 통한 물질 이동은 세포의 에너지 투입 없이 일어나는 수동수송과 수송을 위해 세포가 에너지를 소비해야 하는 능동수송 중 하나에 의해 수행된다. 또한, 막은 막전위를 유지한다. 세포막은 선택적 필터 역할을 하여 특정 물질만 세포 안팎으로 출입할 수 있도록 한다. 세포는 생체막이 관여하는 다음과 같은 수송 메커니즘을 이용한다.
1. 수동적인 삼투와 확산: 이산화탄소(CO2)나 산소(O2)와 같은 몇몇 물질은 확산에 의해 세포막을 통과할 수 있으며, 이는 수동적인 수송 과정이다. 막은 특정 물질이나 이온에 대한 장벽 역할을 하기 때문에, 막의 양쪽에는 다른 농도가 형성된다. 확산은 저분자나 이온이 고농도 쪽에서 저농도 쪽으로 자유롭게 이동할 때 발생한다. 이 과정은 에너지를 필요로 하지 않고, 막의 양쪽에 형성된 농도 기울기에 의해 추진되므로 수동 수송으로 간주된다.[29] 반투과성 막을 사이에 둔 농도 기울기는 물의 삼투류를 유발한다. 생체 내 삼투는 용매의 반투과성 막 통과를 수반하지만, 용매는 농도 기울기에 따라 이동하고 에너지를 필요로 하지 않으므로 마찬가지로 수동적이다. 세포 내 용매는 물이 가장 일반적이지만, 다른 액체나 초임계유체, 기체일 수도 있다.[30]
2. 막관통 채널과 수송체: 막관통 단백질은 분자의 막 통과를 위해 막의 양쪽에서 기능한다.[31] 당이나 아미노산과 같은 영양소는 세포로 들어갈 필요가 있고, 특정 대사산물은 세포 밖으로 나갈 필요가 있다. 이러한 분자들은 아쿠아포린과 같은 단백질 채널을 수동적으로 통과하여 촉진 확산되거나, 막관통 수송체에 의해 막을 넘어 흡수된다. 단백질 채널은 투과 효소(permease)라고도 불리며, 일반적으로 매우 특이성이 높아 제한된 종류의 화학 물질만 (한 종류의 물질로 제한되는 경우도 많음) 인식하여 수송한다. 막관통 단백질의 다른 예로는 세포 표면의 수용체가 있으며, 신호 전달 물질을 이용하여 세포 간 의사소통을 한다.[31]
3. 엔도사이토시스: 엔도사이토시스는 세포가 분자를 감싸서 흡수하는 과정이다. 세포막은 함입(invagination)이라고 불리는 안쪽으로의 작은 변형을 만들어내고, 수송될 물질이 그곳에 포착된다. 이 함입은 세포막 바깥쪽에 위치한 단백질에 의해 일어나며, 그들은 수용체처럼 기능하여 함몰부에 모여 세포막 안쪽으로의 단백질이나 지질의 축적을 촉진한다.[32] 그 후, 변형 부분은 세포 안쪽으로 잘록해져 막으로부터 분리되고, 포착된 물질을 안쪽에 포함하는 소포가 생성된다. 엔도사이토시스는 고체 입자를 흡수하거나(식작용) 저분자나 이온(음작용), 고분자를 흡수하는 경로이다. 엔도사이토시스는 에너지를 필요로 하므로 능동 수송의 한 형태이다.
4. 엑소사이토시스: 함입이나 소포 형성에 의해 세포에 물질이 흡수되는 것과 마찬가지로, 소포의 막은 세포막과 융합하여 내용물을 외부로 배출할 수 있다. 이것이 엑소사이토시스 과정이다. 엑소사이토시스는 엔도사이토시스에 의해 흡수된 물질의 미소화 잔여물을 제거하거나, 호르몬이나 효소와 같은 물질을 분비하거나, 물질을 세포의 장벽을 넘어 수송하는 등의 목적으로 다양한 세포에서 수행된다. 엑소사이토시스 과정에서 미소화 잔여물을 포함하는 식포나 골지체에서 출아된 분비소포는 먼저 세포골격에 의해 세포 내부에서 표면으로 이동하여 소포의 막과 세포막이 접촉하게 된다. 두 지질 이중층의 지질 분자는 서로 재배열되고, 두 막은 융합한다. 막의 융합에 의해 통로가 형성되고, 소포는 그 내용물을 세포 외부로 배출한다.
5. 3. 세포 부착
세포막은 소낭, 시냅스후 밀도, 포도좀, 침입족, 초점 부착과 같은 다양한 "초막" 구조를 형성하며, 세포 연접을 통해 다른 세포와 연결된다. 이러한 구조는 세포 부착, 세포 간 통신, 세포내이입, 세포외유출을 담당하며, 전자 현미경이나 형광 현미경으로 관찰할 수 있다. 이들은 인테그린, 카데린과 같은 특정 단백질로 구성된다.5. 4. 효소 활성
세포막은 많은 단백질을 포함하고 있으며, 일반적으로 막 부피의 50%를 차지한다.[72] 이러한 단백질은 다양한 생물학적 활성을 담당하며 중요하다. 막 단백질의 기능에는 세포 간 연결, 표면 인식, 세포 골격 연결, 신호전달, 효소 활성, 막을 가로지른 물질 수송 등이 포함된다. 효모에서는 약 3분의 1의 유전자가 막 단백질을 암호화하고 있으며, 이 비율은 다세포 생물에서 더 높아진다.[68]5. 5. 막 전위 형성 및 유지
세포막은 선택적 투과성을 가지며, 세포 안팎으로 물질 출입을 조절하여 생존에 필요한 물질의 수송을 촉진한다. 막을 통한 물질 이동은 세포가 에너지를 소모하지 않는 수동 수송과 에너지를 소모하는 능동 수송으로 나뉜다. 또한, 막은 막전위를 유지하며, 특정 물질만 세포 안팎으로 출입할 수 있게 하는 선택적 필터 역할을 한다. 세포는 생체막을 이용한 다양한 수송 메커니즘을 사용한다.# 수동적인 삼투와 확산: 이산화탄소(CO2)나 산소(O2)와 같은 일부 물질은 확산(수동 수송)을 통해 세포막을 통과한다.[74] 막은 특정 물질이나 이온에 대한 장벽 역할을 하므로, 막 양쪽에 서로 다른 농도가 형성된다. 확산은 작은 분자나 이온이 고농도에서 저농도로 자유롭게 이동하는 현상이며, 에너지 소모 없이 막 양쪽의 농도 기울기에 의해 추진된다. 반투과성 막을 경계로 한 농도 기울기는 물의 삼투류를 유발한다. 생체 내 삼투는 용매가 반투과성 막을 통과하지만, 농도 기울기에 따라 이동하고 에너지를 소모하지 않으므로 수동적이다. 세포 내 용매는 주로 물이지만, 다른 액체나 초임계유체, 기체일 수도 있다.[75]
# 막관통 채널과 수송체: 막관통 단백질은 막 양쪽에서 기능하며 분자의 막 통과를 돕는다.[76] 당이나 아미노산과 같은 영양소는 세포로 들어가야 하고, 특정 대사산물은 세포 밖으로 배출되어야 한다. 이러한 분자들은 아쿠아포린과 같은 단백질 채널을 통해 수동적으로 촉진 확산되거나, 막관통 수송체에 의해 막을 넘어 흡수된다. 투과 효소(permease)라고도 불리는 단백질 채널은 대개 매우 특이적이어서 제한된 종류의 화학 물질(때로는 한 종류)만 인식하고 수송한다. 막관통 단백질의 다른 예로는 세포 표면 수용체가 있으며, 신호 전달 물질을 이용해 세포 간 소통을 한다.[76]
# 엔도사이토시스: 세포가 분자를 감싸 흡수하는 과정이다. 세포막은 함입(invagination)이라는 작은 변형을 안쪽으로 만들어 수송될 물질을 포획한다. 함입은 세포막 바깥쪽 단백질에 의해 일어나며, 이들은 수용체처럼 기능하여 함몰부에 모여 세포막 안쪽으로 단백질이나 지질 축적을 촉진한다.[77] 이후 변형 부분은 세포 안쪽으로 잘록해져 막에서 분리되고, 포획된 물질을 포함하는 소포를 생성한다. 엔도사이토시스는 고체 입자(식작용)나 저분자, 이온(음세포작용), 고분자를 흡수하는 경로이다. 에너지를 필요로 하므로 능동 수송의 한 형태이다.
# 엑소사이토시스: 함입이나 소포 형성을 통해 세포에 물질이 흡수되는 것처럼, 소포 막은 세포막과 융합하여 내용물을 외부로 배출할 수 있다. 이것이 엑소사이토시스 과정이다. 엑소사이토시스는 엔도사이토시스로 흡수된 물질의 미소화 잔여물을 제거하거나, 호르몬이나 효소와 같은 물질을 분비하거나, 물질을 세포 장벽 너머로 수송하기 위해 다양한 세포에서 일어난다. 엑소사이토시스 과정에서 미소화 잔여물을 포함하는 식포나 골지체에서 생성된 분비소포는 먼저 세포골격에 의해 세포 내부에서 표면으로 이동하여 소포 막과 세포막이 접촉한다. 두 지질 이중층의 지질 분자는 서로 재배열되고, 두 막은 융합한다. 막 융합으로 통로가 형성되고, 소포는 내용물을 세포 외부로 배출한다.
6. 원핵세포의 막
원핵생물은 고세균과 세균의 두 가지 그룹으로 나뉘며, 세균은 다시 그람 양성균과 그람 음성균으로 더 세분화된다. 그람 음성균은 주변질에 의해 분리된 세포막과 외막을 모두 가지고 있지만, 다른 원핵생물은 세포막만 가지고 있다. 이 두 막은 여러 면에서 다르다. 그람 음성균의 외막은 이중층의 외부를 형성하는 인지질과 내부를 형성하는 리포단백질과 인지질 때문에 다른 원핵생물과 다르다.[33] 외막은 일반적으로 기공 형성 단백질인 그람 음성 포린과 같은 막 단백질의 존재로 인해 다공성을 띠게 된다. 내부 세포막은 일반적으로 대칭적이지만, 앞서 언급한 단백질들 때문에 외막은 비대칭적이다.
또한, 원핵생물 막의 유동성에는 여러 가지 요인이 영향을 미친다. 유동성에 영향을 미치는 주요 요인 중 하나는 지방산 조성이다. 예를 들어, 세균 ''Staphylococcus aureus''(황색포도상구균)를 37℃에서 24시간 배양했을 때, 막은 젤과 같은 상태 대신 더 유동적인 상태를 나타냈다. 이는 고온에서 막이 저온보다 더 유동적이라는 개념을 뒷받침한다. 막이 더 유동적이 되고 더 안정화될 필요가 있을 때, 막을 안정화하는 데 도움이 되도록 더 긴 지방산 사슬이나 포화 지방산 사슬을 만든다.[34]
세균은 또한 펩티도글리칸(아미노산과 당)으로 구성된 세포벽으로 둘러싸여 있다. 일부 진핵세포도 세포벽을 가지고 있지만, 펩티도글리칸으로 만들어진 세포벽은 없다. 그람 음성균의 외막은 세포의 자연 면역을 자극하는 다당류 또는 올리고당류와 탄수화물 지질 영역이 결합된 지질다당류가 풍부하다.[35] 외막은 스트레스 조건하에서 또는 숙주 표적 세포를 만났을 때 병원성 요구사항에 따라 돌출부로 팽출될 수 있으며, 따라서 이러한 돌출부는 병독성 소기관으로 작용할 수 있다.[36] 세균 세포는 원핵세포막이 유기체의 생태적 지위에 맞는 구조로 적응하는 다양한 방식의 수많은 예를 제공한다. 예를 들어, 특정 세균 세포 표면의 단백질은 활주 운동을 돕는다.[37] 많은 그람 음성균은 ATP 구동 단백질 수송 시스템을 포함하는 세포막을 가지고 있다.[37]
참조
[1]
웹사이트
Kimball's Biology pages, Cell Membranes
http://users.rcn.com[...]
[2]
서적
Bacteria in Biology, Biotechnology and Medicine
Wiley
[3]
논문
Physiology, Membrane
https://www.ncbi.nlm[...]
2019-03-02
[4]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
Garland Science
[5]
논문
Membrane assembly driven by a biomimetic coupling reaction
2012-01-01
[6]
웹사이트
Chemists Synthesize Artificial Cell Membrane
https://www.scienced[...]
ScienceDaily
2012-01-25
[7]
웹사이트
Chemists create artificial cell membrane
http://www.kurzweila[...]
kurzweilai.net
2012-01-26
[8]
논문
The effects of intra-membrane viscosity on lipid membrane morphology: complete analytical solution
2018-01-01
[9]
논문
Once upon a time the cell membranes: 175 years of cell boundary research
2014-12-01
[10]
웹사이트
Chronological history of lipid center
http://www.cyberlipi[...]
2017-11-11
[11]
논문
On Bimolecular Layers of Lipoids on the Chromocytes of the Blood
1925-03-01
[12]
서적
Cell and Molecular Biology
John Wiley & Sons, Inc.
[13]
논문
The fluid mosaic model of the structure of cell membranes
[14]
논문
Plasmolytische Studien über die Wand der Vakuolen
[15]
서적
Osmotische Untersuchungen: Studien zur Zell Mechanik
Engelmann, Leipzig
[16]
서적
The Physiology of Plants
https://www.biodiver[...]
[17]
서적
Introduction To Cytology
https://archive.org/[...]
McGraw Hill
[18]
서적
Charles Ernest Overton's concept of a cell membrane
https://books.google[...]
Academic Press, San Diego
[19]
논문
Structure and locomotion in Amoeba proteus
[20]
논문
Membranes in the plant cell. I. Morphological membranes at protoplasmic surfaces
[21]
서적
Plant Cell Biology: From Astronomy to Zoology
https://books.google[...]
Elsevier/Academic Press
2009-01-01
[22]
논문
Assessment of Membrane Fluidity Fluctuations during Cellular Development Reveals Time and Cell Type Specificity
2016-06-30
[23]
서적
Molecular Cell Biology
https://www.ncbi.nlm[...]
Scientific American Books
[24]
논문
Structure of the Plasma Membrane
https://www.ncbi.nlm[...]
2000-01-01
[25]
논문
Biomembranes: Structural Organization and Basic Functions
https://www.ncbi.nlm[...]
2000-01-01
[26]
논문
Cell-surface carbohydrates in cell recognition and response
1986-07-01
[27]
웹사이트
Membrane Structure
http://www.bio.david[...]
Davidson College
[28]
논문
Post-Translational Modifications and Quality Control in the Rough ER
https://www.ncbi.nlm[...]
2000-01-01
[29]
논문
Transport of Small Molecules
https://www.ncbi.nlm[...]
2000-01-01
[30]
논문
Osmosis is not driven by water dilution
2013-04-01
[31]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
2002-01-01
[32]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
Garland Science
2002-01-01
[33]
서적
Medical Microbiology
https://www.ncbi.nlm[...]
University of Texas Medical Branch at Galveston
1996-01-01
[34]
논문
Carotenoid-related alteration of cell membrane fluidity impacts Staphylococcus aureus susceptibility to host defense peptides
2011-02-01
[35]
논문
Bacterial lipopolysaccharides and innate immunity
2001-01-01
[36]
논문
A structural model for virulence organellae of gram negative organisms with reference to Salmonella pathogenicity in chicken ileum
https://www.research[...]
[37]
논문
Microcompartments and protein machines in prokaryotes
2013
[38]
논문
The fluid mosaic model of the structure of cell membranes
1972-02-00
[39]
웹사이트
Basolateral cell membrane
https://www.uniprot.[...]
2023-06-15
[40]
논문
Mediation, modulation, and consequences of membrane-cytoskeleton interactions
[41]
논문
From extracellular to intracellular: the establishment of mitochondria and chloroplasts
1979-04-00
[42]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
Garland Science
2002
[43]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
Garland Science
2002
[44]
논문
The Endoplasmic Reticulum
https://www.ncbi.nlm[...]
2000
[45]
논문
The asymmetrical structure of Golgi apparatus membranes revealed by in situ atomic force microscope
2013-04-16
[46]
논문
Skeletal muscle basement membrane-sarcolemma-cytoskeleton interaction minireview series
2003-04-00
[47]
논문
Modulation of the bilayer thickness of exocytic pathway membranes by membrane proteins rather than cholesterol
2004-03-00
[48]
논문
Structure and function of the sarcolemma of skeletal muscle
1966-07-00
[49]
논문
Cell surface changes in the egg at fertilization
2009-10-00
[50]
논문
Characteristics of the Neuron
https://www.ncbi.nlm[...]
1999
[51]
논문
The role of the axolemma in the initiation of traumatically induced axonal injury
1998-03-00
[52]
웹사이트
Kimball's Biology pages, Cell Membranes
http://users.rcn.com[...]
[53]
서적
Bacteria in Biology, Biotechnology and Medicine
Wiley
[54]
서적
Molecular Biology of the Cell
https://www.ncbi.nlm[...]
Garland Science
[55]
논문
Membrane Assembly Driven by a Biomimetic Coupling Reaction
2011-12-29
[56]
뉴스
Chemists Synthesize Artificial Cell Membrane
https://www.scienced[...]
ScienceDaily
2012-01-25
[57]
뉴스
Chemists create artificial cell membrane
http://www.kurzweila[...]
kurzweilai.net
2012-01-26
[58]
논문
Once upon a time the cell membranes: 175 years of cell boundary research
2014-12-19
[59]
웹사이트
Chronological history of lipid center
http://www.cyberlipi[...]
[60]
논문
On bimolecular layers of lipoids on the chromocytes of the blood
[61]
논문
The fluid mosaic model of the structure of cell membranes
[62]
논문
Assessment of Membrane Fluidity Fluctuations during Cellular Development Reveals Time and Cell Type Specificity
2016-06-30
[63]
서적
Molecular Cell Biology
https://www.ncbi.nlm[...]
Scientific American Books
[64]
논문
Structure of the Plasma Membrane
https://www.ncbi.nlm[...]
2000
[65]
논문
Fatty acid composition of membrane bilayers: importance of diet polyunsaturated fat balance
https://www.ncbi.nlm[...]
2012-05-00
[66]
논문
Homeoviscous adaptation--a homeostatic process that regulates the viscosity of membrane lipids in Escherichia coli
https://www.ncbi.nlm[...]
1974-02-00
[67]
서적
An introduction to biological membranes : composition, structure and function
https://www.worldcat[...]
[68]
논문
Biomembranes: Structural Organization and Basic Functions
https://www.ncbi.nlm[...]
2000
[69]
논문
Sonication-Based Basic Protocol for Liposome Synthesis
https://www.ncbi.nlm[...]
2017
[70]
논문
Cell-surface carbohydrates in cell recognition and response
1986-07-00
[71]
서적
Molecular biology of the cell
https://www.ncbi.nlm[...]
Garland Science
2002
[72]
웹사이트
Membrane Structure
http://www.bio.david[...]
Davidson College
2007-01-11
[73]
논문
Post-Translational Modifications and Quality Control in the Rough ER
https://www.ncbi.nlm[...]
2000
[74]
논문
Transport of Small Molecules
https://www.ncbi.nlm[...]
2000
[75]
논문
Osmosis is not driven by water dilution
http://linkinghub.el[...]
[76]
논문
Membrane Proteins
https://www.ncbi.nlm[...]
2002
[77]
논문
Transport into the Cell from the Plasma Membrane: Endocytosis
https://www.ncbi.nlm[...]
2002
[78]
서적
Medical Microbiology
https://www.ncbi.nlm[...]
University of Texas Medical Branch at Galveston
1996
[79]
논문
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com