청각계
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
청각계는 소리 파동을 감각 세포로 전달하는 외이, 중이, 내이(달팽이관)로 구성된 청각 기관의 구조와 기능을 총칭한다. 외이는 귓바퀴와 외이도로 소리 파동을 고막으로 전달하고, 중이는 고막의 진동을 이소골을 통해 내이로 증폭하여 전달한다. 내이는 달팽이관 내 코르티 기관에서 기계적 운동을 전기 신호로 변환하여 청각 정보를 처리한다. 청각 경로는 청각 정보를 뇌의 여러 영역으로 전달하며, 청각 피질에서 소리 지각이 이루어진다. 청각계의 손상은 청각 질환, 청각 처리 장애, 과청각증, 이명 등을 유발하며, 조기 진단 및 치료가 중요하다.
더 읽어볼만한 페이지
| 청각계 | |
|---|---|
| 개요 | |
| 계통 | 감각 기관 |
| 기능 | 듣기 |
| 구조 | |
| 외부 | 귓바퀴 외이도 |
| 중간 | 고막 이소골 유스타키오관 |
| 내부 | 달팽이관 전정기관 반고리관 |
| 신경 | |
| 관련 신경 | 내이신경 |
2. 청각 기관의 구조와 기능
음파는 바깥귀를 통해 고막으로 전달되고, 가운데귀의 이소골을 거쳐 속귀의 달팽이관으로 전달된다. 이 과정에서 음파는 증폭되고, 액체 진동으로 변환되어 코르티 기관의 청각 세포를 자극한다.[31]
'''청각 기관의 구조'''는 다음과 같다.
- 바깥귀: 귓바퀴와 외이도로 구성되어 소리를 모으고 고막으로 전달한다.
- 가운데귀: 고막, 망치뼈, 모루뼈, 등자뼈 (이소골)로 구성되어 소리 진동을 증폭하여 난원창을 통해 달팽이관으로 전달한다.
- 속귀: 달팽이관, 전정기관, 반고리관으로 구성된다. 달팽이관은 소리를 감지하고, 전정기관과 반고리관은 몸의 균형을 담당한다.
전정관과 고실관은 달팽이액으로, 달팽이관은 내림프액으로 채워져 있다.[1][2][3] 코르티 기관의 외부 세포는 달팽이액 진동에 의해 프레스틴을 방출하여 세포를 움직이게 하고, 이는 기저막 움직임에 영향을 주어 소리 진폭을 증폭시킨다.[4]
기저막은 유모 세포 위치에 따라 감지되는 주파수를 조절한다. 달팽이관 기저부는 좁고 강하며 고주파수를, 정점은 넓고 약하며 저주파수를 감지한다. 덮개막은 유모세포를 자극하여 달팽이관 증폭을 돕고, 기저막과 함께 주파수 구분에 기여한다.[5][6][7][8][9][10]
2. 1. 외이 (Outer ear)
바깥귀는 소리 진동을 고막으로 유입하여 중간 주파수 범위에서 음압을 증가시키는 역할을 한다. 바깥귀는 귓바퀴와 외이도로 구성된다.2. 1. 1. 귓바퀴 (Pinna)
귓바퀴를 둘러싼 연골의 주름을 이개라고 한다. 소리 파동이 귓바퀴에 부딪히면 반사되고 감쇠되며, 이러한 변화는 뇌가 소리의 방향을 결정하는 데 도움이 되는 추가 정보를 제공한다.[31]2. 1. 2. 외이도 (External auditory canal)
소리 파동은 속이 빈 튜브인 외이도로 들어간다. 외이도는 3~12 kHz 사이의 소리를 증폭시킨다.[31] 외이도 끝에 있는 고막은 중이의 시작을 나타낸다.2. 2. 중이 (Middle ear)
바깥귀는 소리 진동을 고막으로 유입하여 중간 주파수 범위에서 음압을 증가시킨다. 가운데귀의 이소골은 진동 압력을 약 20배 더 증폭시킨다. 등골의 바닥은 난원창을 통해 달팽이관으로 진동을 전달하며, 난원창은 달팽이액 액체를 진동시키고(전체 속귀에 존재) 정원창이 난원창이 팽창하면서 밖으로 튀어나오게 한다.[31]소리 파동은 귓길을 통해 고막에 부딪히고, 이 파동은 망치뼈, 모루뼈, 등자뼈를 통해 중이강을 가로지른다. 이 뼈들을 이소골이라 하며, 지레 작용을 통해 고막의 진동을 타원창에서 더 높은 압력으로 변환한다. 망치뼈는 고막과, 등자뼈는 타원창과 연결된다. 타원창 너머 내이는 액체를 포함하고 있어, 더 높은 압력이 필요하다. 중이 근육의 등골 반사는 등골근을 활성화하여 소리 전달을 줄여 내이를 보호한다. 중이는 파동 형태의 소리 정보를 달팽이관에서 신경 임펄스로 변환한다.
2. 2. 1. 고막 (Tympanic membrane)
소리 파동은 귓길을 통해 이동하여 고막에 부딪힌다. 이 파동 정보는 망치뼈(망치), 모루뼈(모루), 등골뼈(등자)와 같은 일련의 섬세한 뼈들을 통해 공기로 채워진 중이강을 가로지른다. 이 소골들은 지레 역할을 하여 저압의 고막 소리 진동을 타원창(또는 안뜰창)이라고 하는 또 다른 더 작은 막에서 더 높은 압력의 소리 진동으로 변환한다. 망치뼈의 자루는 고막과 연결되고, 등골뼈의 발판(바닥)은 타원창과 연결된다. 타원창 너머의 내이는 공기가 아닌 액체를 포함하고 있기 때문에 고막보다 타원창에서 더 높은 압력이 필요하다. 중이 근육의 등골 반사는 소리에 반응하여 등골근이 활성화될 때 소리 에너지 전달을 줄여 내이를 손상으로부터 보호하는 데 도움이 된다.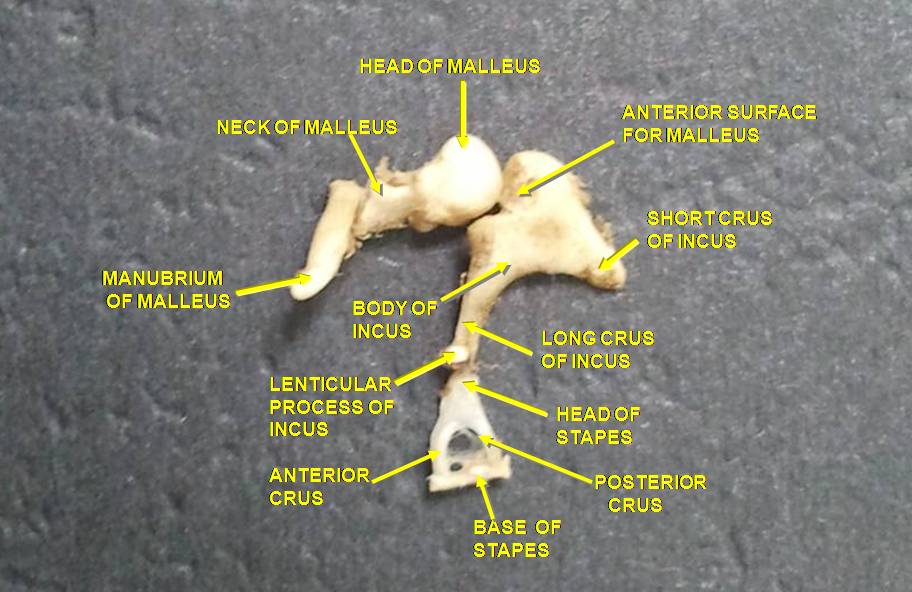
2. 2. 2. 이소골 (Ossicles)
소리 파동은 귓길을 통해 이동하여 고막에 부딪힌다. 이 파동 정보는 망치뼈, 모루뼈, 등자뼈와 같은 일련의 섬세한 뼈들을 통해 공기로 채워진 중이강을 가로질러 이동한다. 이 뼈들을 이소골(소골)이라고 하며, 지레 역할을 하여 저압 고막 소리 진동을 타원창(안뜰창)이라고 하는 또 다른 더 작은 막에서 더 높은 압력의 소리 진동으로 변환한다. 망치뼈의 자루는 고막과 연결되고, 등골뼈의 발판(바닥)은 타원창과 연결된다. 타원창 너머의 내이는 공기가 아닌 액체를 포함하고 있기 때문에 고막보다 타원창에서 더 높은 압력이 필요하다. 중이 근육의 등골 반사는 소리에 반응하여 등골근이 활성화될 때 소리 에너지 전달을 줄여 내이를 손상으로부터 보호하는 데 도움이 된다.[1] 중이는 여전히 파동 형태의 소리 정보를 포함하고 있으며, 이는 달팽이관에서 신경 임펄스로 변환된다.[1]2. 2. 3. 이관 (Eustachian tube)
이관은 중이와 관련된 내용이 아니므로, 주어진 `source`에서 이관에 대한 정보를 찾을 수 없습니다. 따라서 이 섹션에는 내용을 작성할 수 없습니다. (이 출력은 변경할 내용이 없습니다.)2. 3. 내이 (Inner ear)
내이는 달팽이관과 여러 비청각 구조로 구성된다. 바깥귀를 통해 들어온 공기 진동은 고막을 거쳐 중이의 이소골에 의해 증폭되어 내이에 기계적으로 전달된다.내이에서는 기계적 운동이 바깥 림프에 전해져 림프에 파동을 일으키고, 이것이 코르티 기관에 도달하면 청각 세포를 자극하여 소리를 감지한다. 공기 진동은 액체 진동으로 바뀐 다음 감각 세포에 도달한다.[31]
전정관과 고실관은 달팽이액으로 채워져 있으며, 그 사이의 달팽이관은 내림프액으로 채워져 있다.[1][2][3] 전정관의 달팽이액 진동은 코르티 기관 외부 세포를 구부려 프레스틴을 방출하게 한다. 이는 세포가 화학적으로 늘어나고 수축하도록 하고, 모발 다발이 이동하여 기저막의 움직임에 전기적으로 영향을 미친다. 이러한 운동은 전달 파동 진폭을 40배 이상 증폭시킨다.[4]
기저막은 스칼라 사이의 장벽이며, 그 가장자리를 따라 유모 세포가 위치한다. 기저막의 너비와 강성은 유모세포에 의해 감지되는 주파수를 제어한다. 달팽이관 기저부에서 기저막은 가장 좁고 강하며(고주파수), 달팽이관 정점에서 가장 넓고 약하다(저주파수). 덮개막은 유모세포를 자극하여 달팽이관 증폭을 돕는다. 덮개막의 너비와 강성은 기저막과 평행하며 주파수 구분에 도움이 된다.[5][6][7][8][9][10]
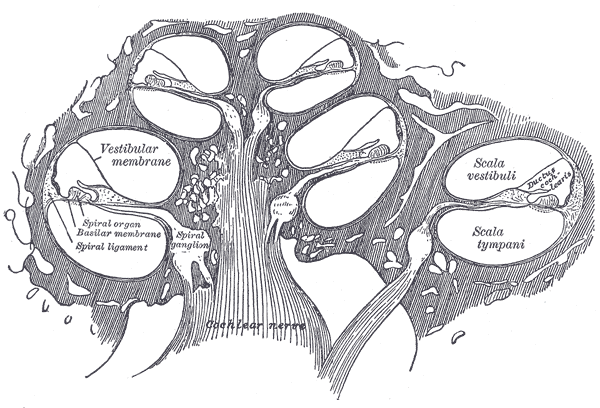
2. 3. 1. 달팽이관 (Cochlea)
내이는 달팽이관과 여러 비청각 구조로 구성된다. 달팽이관은 중간 계단, 고실 계단, 전정 계단의 세 유체 구획으로 채워져 있으며, 두 구획을 분리하는 바닥막을 가로지르는 압력에 의해 구동되는 유체 파동을 지지한다. 달팽이관 도관 또는 ''중간 계단''에는 내림프가 포함되어 있다. 코르티 기관은 바닥막의 이 도관에 위치하며, 기계적 파동을 뉴런의 전기 신호로 변환한다. ''고실 계단''과 ''전정 계단''은 뇌척수액과 유사한 성분인 주위림프로 채워진 골 미로 내에 위치한다. 내림프와 주위림프 유체 간의 화학적 차이는 칼륨 이온과 칼슘 이온 간의 전위차로 인해 내이의 기능에 중요하다.[26]
인간 달팽이관의 평면도는 (모든 포유류와 대부분의 척추동물의 전형적인) 특정 주파수가 그 길이를 따라 발생하는 위치를 보여준다. 주파수는 코르티 기관 내의 달팽이관 길이에 대한 대략적인 지수 함수이다. 박쥐와 돌고래와 같은 일부 종에서는 활동적인 음파 탐지 기능을 지원하기 위해 특정 영역에서 관계가 확장된다.
코르티 기관은 달팽이관 전체 ''달팽이관 중간계''를 따라 길게 뻗어 있는 감각 상피의 띠를 형성한다. 이 기관의 유모 세포는 액체 파동을 신경 신호로 변환한다. 수많은 신경의 여정은 이 첫 번째 단계에서 시작된다. 여기에서 추가적인 처리를 거쳐 다양한 청각 반응과 감각을 만들어낸다.
털세포는 기둥 모양의 세포로, 각 세포의 상단에는 100~200개의 특수화된 입체섬모로 이루어진 "털 묶음"이 있어 털세포라고 불린다. 청각 시스템에는 두 가지 유형의 털세포, 즉 ''내부'' 및 ''외부'' ''털'' ''세포''가 있다. 내털세포는 청각을 위한 기계 수용체로, 소리의 진동을 신경 섬유의 전기적 활동으로 변환하여 뇌로 전달한다. 외털세포는 운동 구조이다. 소리 에너지는 이러한 세포의 모양을 변화시켜 주파수별 방식으로 소리 진동을 증폭시킨다. 내털세포의 가장 긴 섬모 위에 가볍게 놓여 있는 것은 덮개막으로, 소리의 각 주기에 따라 앞뒤로 움직이며 섬모를 기울여 털세포의 전기적 반응을 유발한다.
내털세포는 눈의 광수용체와 마찬가지로 다른 뉴런의 활동 전위 전형적인 단계적 반응을 보인다. 이러한 단계적 전위는 활동 전위의 "전부 또는 전무" 속성에 얽매이지 않는다.
털 묶음의 흔들림이 막 전위의 차이를 유발하는 원리는 섬모가 한 섬모의 끝을 다른 섬모에 연결하는 구조인 "팁 링크"에 의해 서로 연결되어 있다는 것이다. 팁 링크는 늘어나고 압축되면서 이온 채널을 열어 털세포에서 수용체 전위를 생성할 수 있다. 최근에 cadherin-23 CDH23와 protocadherin-15 PCDH15가 이러한 팁 링크와 관련된 접착 분자임이 밝혀졌다.[26] 칼슘 구동 모터가 이러한 링크를 짧게 하여 장력을 재생성하는 것으로 생각된다. 이러한 장력의 재생성은 장기간의 청각 자극을 파악할 수 있게 해준다.[27]
2. 3. 2. 청신경 (Auditory nerve)
털세포는 기둥 모양의 세포로, 각 세포의 상단에는 100~200개의 특수화된 입체섬모로 이루어진 "털 묶음"이 있어 털세포라고 불린다. 청각 시스템에는 두 가지 유형의 털세포, 즉 ''내부'' 털세포 및 ''외부'' 털세포가 있다. 내털세포는 청각을 위한 기계 수용체로, 소리의 진동을 신경 섬유의 전기적 활동으로 변환하여 뇌로 전달한다. 외털세포는 운동 구조이다. 소리 에너지는 이러한 세포의 모양을 변화시켜 주파수별 방식으로 소리 진동을 증폭시킨다. 내털세포의 가장 긴 섬모 위에 가볍게 놓여 있는 것은 덮개막으로, 소리의 각 주기에 따라 앞뒤로 움직이며 섬모를 기울여 털세포의 전기적 반응을 유발한다.내털세포는 눈의 광수용체와 마찬가지로 다른 뉴런의 활동 전위가 아닌 전형적인 단계적 반응을 보인다. 이러한 단계적 전위는 활동 전위의 "전부 또는 전무" 속성에 얽매이지 않는다.
이 시점에서 이러한 털 묶음의 흔들림이 막 전위의 차이를 어떻게 유발하는지 질문할 수 있다. 현재 모델은 섬모가 한 섬모의 끝을 다른 섬모에 연결하는 구조인 "팁 링크"에 의해 서로 연결되어 있다는 것이다. 팁 링크는 늘어나고 압축되면서 이온 채널을 열어 털세포에서 수용체 전위를 생성할 수 있다. 최근에 cadherin-23 CDH23와 protocadherin-15 PCDH15가 이러한 팁 링크와 관련된 접착 분자임이 밝혀졌다.[26] 칼슘 구동 모터가 이러한 링크를 짧게 하여 장력을 재생성하는 것으로 생각된다. 이러한 장력의 재생성은 장기간의 청각 자극을 파악할 수 있게 해준다.[27]
구심성 뉴런은 달팽이관 내이모세포를 지배하며, 글루탐산 신경전달물질이 털 세포에서 일차 청각 뉴런의 가지돌기로 신호를 전달하는 시냅스에서 작용한다.
달팽이관에는 구심성 신경 섬유보다 내이모세포가 훨씬 적으며, 많은 청각 신경 섬유가 각 털 세포를 지배한다. 신경 가지돌기는 청각 신경의 뉴런에 속하며, 이 신경은 다시 전정 신경과 합쳐져 전정와우 신경, 즉 뇌신경 VIII번을 형성한다.[28] 특정 구심성 신경 섬유에 입력을 공급하는 기저막의 영역을 그 수용 영역으로 간주할 수 있다.
뇌에서 달팽이관으로의 원심성 투사 또한 소리 지각에 중요한 역할을 하지만, 아직 잘 알려져 있지 않다. 원심성 시냅스는 외이모세포와 내이모세포 아래의 구심성(뇌로 향하는) 가지돌기에서 발생한다.
3. 청각 정보 처리 과정
음파는 청각 기관을 통해 감각 세포로 전달된다. 청각기는 바깥귀, 가운데귀, 속귀(달팽이관)로 구성된다. 바깥귀로 들어온 공기의 진동은 고막을 진동시키고, 가운데귀의 이소골을 통해 속귀로 증폭되어 전달된다.[31]
속귀에서는 기계적 진동이 바깥 림프액에 파동을 일으켜 코르티 기관의 청각 세포를 자극한다. 공기 진동은 청각 세포에 직접 작용하지 않고, 중간 장치에서 증폭되어 액체 진동으로 바뀐 후 감각 세포에 도달한다. 바깥귀는 소리 진동을 고막으로 모아 음압을 높이고, 가운데귀의 이소골은 진동 압력을 약 20배 증폭시킨다. 등자뼈 바닥은 난원창을 통해 달팽이관으로 진동을 전달하고, 난원창은 달팽이액을 진동시켜 정원창을 밖으로 튀어나오게 한다.[31]
전정관과 고실관은 달팽이액으로 채워져 있고, 그 사이의 달팽이관은 내림프액으로 채워져 있다.[1][2][3] 전정관의 달팽이액 진동은 코르티 기관 외부 세포를 구부려 프레스틴을 방출하게 한다. 이는 세포가 늘어나고 수축하게 하고, 모발 다발을 움직여 기저막 움직임에 영향을 준다. 이러한 외부 유모 세포는 전달 파동 진폭을 40배 이상 증폭시킨다.[4] 외부 유모 세포는 나선 신경절에 의해 최소한으로 지배받는 반면, 내부 유모 세포는 하나의 털당 30개 이상의 신경 섬유와 연결되어 있다.
기저막은 스칼라 사이의 장벽으로, 그 가장자리를 따라 내부 유모 세포와 외부 유모 세포가 위치한다. 기저막의 너비와 강성은 내부 유모 세포가 감지하는 주파수를 제어한다. 달팽이관 기저부에서 기저막은 가장 좁고 강하며(고주파수), 달팽이관 정점에서는 가장 넓고 약하다(저주파수). 개막은 외부 유모 세포와 내부 유모 세포를 자극하여 달팽이관 증폭을 돕는다. 개막 너비와 강성은 기저막과 유사하게 주파수 구분에 기여한다.[5][6][7][8][9][10]
올리브 상핵은 뇌교에 위치하며, 좌우 달팽이관 펄스가 처음으로 합쳐지는 지점이다. 사다리꼴체는 대부분의 달팽이핵 섬유가 교차하는 곳으로, 소리 위치 파악에 도움을 준다.[15] 달팽이관 신경 섬유는 각각 가장 민감한 주파수를 가지며 광범위한 수준에서 반응한다.[16][17]
신경 섬유의 신호는 부시 세포에 의해 올리브 복합체의 이음향 영역으로 전달되고, 신호 피크와 밸리는 별 모양 세포에 의해 기록되며, 신호 타이밍은 문어 세포에 의해 추출된다. 가쪽 섬유띠의 복부 핵은 위둔덕이 위상 및 긴장성 반응을 제공하여 진폭 변조 소리를 해독하는 데 도움을 준다. 위둔덕은 시각, 뇌교, 척수, 시상 등 다양한 입력을 받는다.[18]
내측 무릎핵은 복부, 등쪽, 내측으로 나뉜다. 청각 피질은 소리를 인식하고 식별하며, 소리의 기원 위치를 파악한다. 청각 피질은 서로 다른 화음, 타이밍, 음조에 반응하는 묶음이 있는 지형 주파수 맵이다. 오른손 청각 피질은 음성에, 왼손 청각 피질은 소리의 미세한 순차적 차이에 더 민감하다.[19][20]
전전두피질의 전내측 및 복외측은 각각 음조 공간에서의 활성화와 단기 기억 저장에 관여한다.[21] 헤슐 고랑/가로 측두 고랑은 베르니케 영역 및 기능을 포함하며, 감정-소리, 감정-얼굴 표정, 소리-기억 과정에 관여한다. 내후각 피질은 시각 및 청각 기억을 돕고 저장한다.[22][23] 덧가쪽이랑은 언어 이해를 돕고 동정심 있는 반응을 담당하며, 소리를 모이랑으로 단어에 연결하고 단어 선택에 도움을 준다. 덧가쪽이랑은 촉각, 시각, 청각 정보를 통합한다.[24][25]
3. 1. 달팽이핵 (Cochlear nucleus)
구심성 뉴런은 달팽이관 내이모세포를 지배하며, 글루탐산 신경전달물질이 털 세포에서 일차 청각 뉴런의 가지돌기로 신호를 전달하는 시냅스에서 작용한다.달팽이관에는 구심성 신경 섬유보다 내이모세포가 훨씬 적으며, 많은 청각 신경 섬유가 각 털 세포를 지배한다. 신경 가지돌기는 청각 신경의 뉴런에 속하며, 이 신경은 다시 전정 신경과 합쳐져 전정와우 신경, 즉 뇌신경 VIII번을 형성한다.[28] 특정 구심성 신경 섬유에 입력을 공급하는 기저막의 영역을 그 수용 영역으로 간주할 수 있다.
뇌에서 달팽이관으로의 원심성 투사 또한 소리 지각에 중요한 역할을 하지만, 아직 잘 알려져 있지 않다. 원심성 시냅스는 외이모세포와 내이모세포 아래의 구심성(뇌로 향하는) 가지돌기에서 발생한다. 달팽이핵은 내이에서 새롭게 변환된 "디지털" 데이터를 신경적으로 처리하는 첫 번째 지점이다(양이 감각 융합 참조). 포유류에서 이 영역은 해부학적으로 그리고 생리학적으로 배쪽 달팽이핵 (DCN)과 복측 달팽이핵 (VCN)의 두 영역으로 나뉜다. VCN은 신경근에 의해 후복측 달팽이핵 (PVCN)과 전복측 달팽이핵 (AVCN)으로 더 나뉜다.[29]
3. 2. 상올리브 복합체 (Superior olivary complex)
상올리브 복합체(SOC)는 교뇌에 위치하며, 주로 복측 달팽이핵으로부터 입력을 받지만, 배측 달팽이핵도 복측 음향 줄기를 통해 입력을 전달한다. 상올리브 복합체는 왼쪽 및 오른쪽 달팽이관에서 오는 신호가 처음으로 합쳐지는 곳이다.[11] 상올리브 복합체는 14개의 핵으로 구성되어 있는데, 여기서는 그 약칭을 사용한다(전체 이름은 올리브 상핵 참조).[11][12][13][14]- MSO (내측 상올리브): 양쪽 귀에서 들어오는 정보의 시간 차이를 측정하여 소리가 나는 방향을 결정한다.
- LSO (외측 상올리브): 양쪽 귀 사이의 소리 크기 차이를 이용하여 소리의 방향을 파악하는 데 도움을 준다. LSO는 주로 내유모세포(IHC)의 신호를 받는다.
- VNTB (복측 사다리꼴체 핵): 외유모세포(OHC)의 신호를 주로 받는다.
- MNTB (내측 사다리꼴체 핵): 글리신을 통해 LSO를 억제한다.
- LNTB (외측 사다리꼴체 핵): 글리신에 영향을 받지 않으며, 빠른 신호 처리에 사용된다.
- DPO (배측 주변올리브 핵): 고주파 소리에 반응하며, 위치에 따라 반응이 달라진다.
- DLPO (배외측 주변올리브 핵): 저주파 소리에 반응하며, 위치에 따라 반응이 달라진다.
- VLPO (복외측 주변올리브 핵): DPO와 기능은 같지만, 다른 영역에서 작용한다.
- PVO, CPO, RPO, VMPO, ALPO, SPON (글리신에 의해 억제됨): 다양한 신호 처리 및 억제 기능을 담당한다.
상올리브 복합체 내부에는 외측 상올리브(LSO)와 내측 상올리브(MSO)가 있다. LSO는 양쪽 귀의 소리 크기 차이를 감지하는 데 중요하며, MSO는 양쪽 귀에 도달하는 시간 차이를 구별하는 데 중요하다.[13]
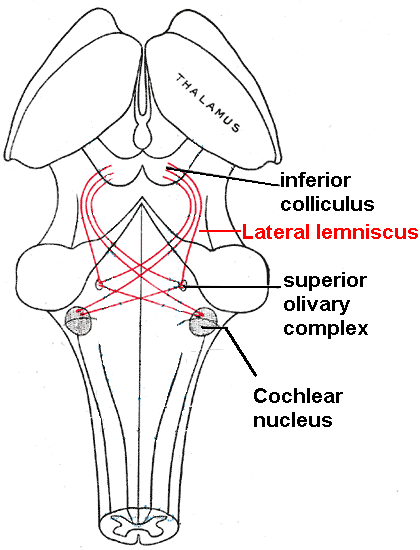
3. 3. 하구 (Inferior colliculus)
가쪽 섬유띠는 뇌줄기 내의 축삭 다발로, 달팽이핵에서 다양한 뇌줄기 핵을 거쳐 반대쪽 중간뇌의 아래둔덕(Inferior colliculus, 하구)으로 소리 정보를 전달한다.하구(IC)는 상구 바로 아래에 위치한다. 하구의 중심핵은 상행 청각 시스템에서 거의 필수적인 중계 역할을 하며, 상올리브 복합체[12]와 등쪽 와우 핵에서 소스 위치 정보와 관련된 정보를 통합한 후 시상 및 대뇌 피질로 보낸다.[31] 하구는 또한 청각 피질과 청각 시상 (내측 슬상핵)으로부터 하행 입력을 받는다.[32]
3. 4. 내측 슬상핵 (Medial geniculate nucleus)
내측 슬상핵은 시상의 일부로, 청각 정보를 중계하는 역할을 한다.3. 5. 청각 피질 (Auditory cortex)
제1 청각 피질은 제2 청각 피질로 둘러싸여 있으며, 서로 연결되어 있다. 이러한 이차 영역은 상측두이랑, 상측두구의 등쪽 가장자리, 전두엽의 추가 처리 영역과 상호 연결된다. 인간의 경우, 이러한 영역과 중측두이랑의 연결은 언어 지각에 중요할 것이다. 청각 지각의 근간이 되는 전두-측두엽 시스템은 소리를 언어, 음악 또는 소음으로 구별할 수 있게 해준다.3. 5. 1. 제1 청각 피질 (Primary auditory cortex)
제1 청각 피질은 청각 입력을 처음으로 받는 대뇌 피질의 영역이다.소리 지각은 왼쪽 뒤쪽 상측두이랑과 관련이 있다. 상측두이랑에는 브로드만 영역 41번과 42번을 포함하여 뇌의 여러 중요한 구조가 포함되어 있으며, 이는 음높이 및 리듬과 같은 소리의 기본적인 특성에 대한 감각을 담당하는 피질 영역인 제1 청각 피질의 위치를 나타낸다.[33][34][35][36][37][38][39] 비인간 영장류 연구를 통해 제1 청각 피질을 기능적으로 구별 가능한 하위 영역으로 더 나눌 수 있다는 것을 알고 있다.
제1 청각 피질의 뉴런은 다양한 청각 주파수를 포괄하는 수용 영역을 가지고 있으며, 조화로운 음높이에 선택적으로 반응한다고 여겨질 수 있다.[40] 두 귀에서 정보를 통합하는 뉴런은 청각 공간의 특정 영역을 포괄하는 수용 영역을 가지고 있다.
3. 5. 2. 청각 연합 피질 (Auditory association cortex)
제1 청각 피질은 청각 입력을 처음으로 받는 대뇌 피질의 영역이다.소리 지각은 왼쪽 뒤쪽 상측두이랑과 관련이 있다. 상측두이랑에는 브로드만 영역 41번과 42번을 포함하여 뇌의 여러 중요한 구조가 포함되어 있으며, 이는 음높이 및 리듬과 같은 소리의 기본적인 특성에 대한 감각을 담당하는 피질 영역인 제1 청각 피질의 위치를 나타낸다. 제1 청각 피질의 뉴런은 다양한 청각 주파수를 포괄하는 수용 영역을 가지고 있으며, 조화로운 음높이에 선택적으로 반응한다고 여겨질 수 있다.[40] 두 귀에서 정보를 통합하는 뉴런은 청각 공간의 특정 영역을 포괄하는 수용 영역을 가지고 있다.
제1 청각 피질은 제2 청각 피질로 둘러싸여 있으며, 서로 연결되어 있다. 이러한 이차 영역은 상측두이랑, 상측두구의 등쪽 가장자리, 전두엽의 추가 처리 영역과 상호 연결된다. 인간의 경우, 이러한 영역과 중측두이랑의 연결은 언어 지각에 중요할 것이다. 청각 지각의 근간이 되는 전두-측두엽 시스템은 소리를 언어, 음악 또는 소음으로 구별할 수 있게 해준다.
4. 청각 경로 (Auditory pathways)
바깥귀는 소리 진동을 고막으로 모아 음압을 증폭시키고, 가운데귀의 이소골은 진동 압력을 약 20배 더 증폭시킨다. 등골은 난원창을 통해 달팽이관으로 진동을 전달하며, 이는 달팽이액을 진동시키고 정원창을 팽창시킨다.[31]
전정관과 고실관은 달팽이액으로, 달팽이관은 내림프액으로 채워져 있다.[1][2][3] 달팽이액 진동은 코르티 기관 외부 세포(4열)를 구부려 프레스틴을 방출, 세포가 수축과 이완을 반복하게 한다(체성 운동). 이 운동은 모발 다발을 움직여 기저막 움직임에 전기적 영향을 주며, 전달 파동 진폭을 40배 이상 증폭시킨다.[4] 외부 유모 세포(OHC)는 나선 신경절에 의해 적게 지배받는 반면, 내부 유모 세포(IHC)는 아페렌트 신경 지배만 가지지만 연결성이 높다. OHC는 IHC보다 3~4배 더 많다.
기저막(BM)은 IHC와 OHC가 위치하는 곳으로, 너비와 강성에 따라 IHC가 감지하는 주파수가 달라진다. 달팽이관 기저부에서 BM은 좁고 강하며(고주파수), 정점에서는 넓고 약하다(저주파수). 개막은 OHC와 IHC를 자극하여 달팽이관 증폭을 돕는다. TM 너비와 강성은 BM과 유사하게 주파수 구분에 기여한다.[5][6][7][8][9][10]
올리브 상핵(SOC)은 교뇌에 위치하며, 좌우 달팽이관 펄스의 첫 수렴 지점이다. MSO는 좌우 정보의 시간차로 소리 각도를, LSO는 귀 사이 소리 수준을 정규화하여 소리 강도로 각도를 파악한다. LSO는 IHC를, VNTB는 OHC를 지배한다. MNTB는 글리신으로 LSO를 억제하고, LNTB는 글리신 면역성이며 빠른 신호 처리에 사용된다. DPO, DLPO, VLPO, PVO, CPO, RPO, VMPO, ALPO, SPON 등 다양한 핵이 신호 및 억제에 관여한다.[11][12][13][14]
사다리꼴체는 대부분의 달팽이관 핵 (CN) 섬유가 교차하는 곳으로, 소리 위치 파악에 도움을 준다.[15] CN은 복부(VCN)와 등쪽(DCN) 영역으로 나뉜다. VCN의 부시 세포는 타이밍 정보를, 별 모양(초퍼) 세포는 소리 스펙트럼을, 문어 세포는 청각 타이밍 코드를 해독한다. DCN의 방추형 세포는 정보를 통합하여 위치에 대한 스펙트럼 단서를 결정한다. 달팽이관 신경 섬유(30,000개 이상)는 각각 민감한 주파수를 가지며 광범위한 수준에서 반응한다.[16][17]
신경 섬유 신호는 부시 세포에 의해 올리브 복합체의 이음향 영역으로, 신호 피크와 밸리는 별 모양 세포, 신호 타이밍은 문어 세포에 의해 처리된다. 가쪽 섬유띠의 핵들은 진폭 변조 소리 해독에 기여한다. 위둔덕(IC)은 시각, 교뇌, 척수, 시상 등 다양한 입력을 받아 '놀람 반응'과 안구 반사에 관여하며, 다중 감각 통합, 음조 감지, 이음향 청취 시간차 결정 등을 수행한다.[18]
내측 무릎 핵은 복부, 등쪽, 내측으로 나뉘며, 각각 주파수, 강도, 이음향 정보, 체감각 정보, 소리 지속 시간 등을 처리한다. 청각 피질(AC)은 소리 인식/지각을 담당하며, 소리 식별 및 위치 파악을 수행한다. AC는 지형 주파수 맵을 가지며, 오른손 AC는 음성, 왼손 AC는 소리의 미세한 순차적 차이에 민감하다.[19][20]
전전두피질의 전내측 및 복외측은 음조 공간 활성화와 단기 기억 저장에 관여한다.[21] 헤슐의 고랑/가로 측두 고랑은 베르니케 영역 및 기능을 포함하며, 감정-소리, 감정-얼굴 표정, 소리-기억 프로세스에 관여한다. 내후각 피질은 시각 및 청각 기억을 돕고 저장한다.[22][23] 덧가쪽이랑(SMG)은 언어 이해와 동정심 반응을 담당하며, 소리를 각이랑으로 연결하고 단어 선택을 돕는다. SMG는 촉각, 시각, 청각 정보를 통합한다.[24][25]
4. 1. 배쪽 청각 경로 (What pathway)
일차 청각 피질에서 두 개의 별도 경로, 즉 청각 배쪽 스트림과 청각 등쪽 스트림이 발생한다.[41] 청각 배쪽 스트림에는 전 상측두이랑, 전 상측두구, 중간 측두이랑 및 측두극이 포함된다. 이 영역의 뉴런은 소리 인식과 문장에서 의미 추출을 담당한다. 청각 등쪽 스트림에는 후 상측두이랑 및 구, 하두정 소엽 및 두정 내구가 포함된다. 두 경로는 인간의 하전두이랑으로 투사된다. 영장류에서 청각 등쪽 스트림의 가장 확립된 역할은 소리 위치 파악이다. 인간의 경우, 좌반구의 청각 등쪽 스트림은 또한 언어 반복 및 발성, 단어 이름의 음운론적 장기 인코딩 및 언어 작업 기억을 담당한다.4. 2. 등쪽 청각 경로 (Where pathway)
일차 청각 피질에서는 청각 배쪽 스트림과 청각 등쪽 스트림, 두 개의 별도 경로가 발생한다.[41] 청각 등쪽 스트림에는 후 상측두이랑 및 구, 하두정 소엽, 두정 내구가 포함된다. 영장류에서 청각 등쪽 스트림의 가장 확립된 역할은 소리 위치 파악이다. 인간의 경우, 좌반구의 청각 등쪽 스트림은 언어 반복 및 발성, 단어 이름의 음운론적 장기 인코딩, 언어 작업 기억도 담당한다.5. 임상적 의의 (Clinical significance)
바깥귀는 소리 진동을 고막으로 보내 음압을 증폭시키고, 가운데귀의 이소골은 진동 압력을 약 20배 더 증폭시킨다. 등골 바닥은 난원창을 통해 달팽이관으로 진동을 전달하며, 이는 달팽이액을 진동시키고 정원창이 팽창하게 한다.[31]
전정관과 고실관은 달팽이액으로, 그 사이의 달팽이관은 내림프액으로 채워져 있다.[1][2][3] 전정관의 달팽이액 진동은 코르티 기관 외부 세포(4열)를 구부려 프레스틴을 방출하게 한다. 이는 세포가 화학적으로 늘어나고 수축하게 하고(체성 운동), 모발 다발이 이동하여 기저막 움직임에 전기적으로 영향을 미친다(모발 다발 운동). 이러한 운동기(외부 유모 세포)는 전달 파동 진폭을 40배 이상 증폭시킨다.[4] 외부 유모 세포(OHC)는 내부 유모 세포(IHC)보다 3~4배 더 많다.
기저막(BM)은 IHC와 OHC가 위치한 곳으로, 너비와 강성은 IHC에 의해 가장 잘 감지되는 주파수를 제어한다. 개막은 OHC와 IHC를 자극하여 달팽이관 증폭을 돕는다.[5][6][7][8][9][10]
올리브 상핵(SOC)은 교뇌에 위치하며, 왼쪽 및 오른쪽 달팽이관 펄스의 첫 번째 수렴 지점이다. MSO는 소리가 나온 각도를, LSO는 소리 강도를 사용하여 소리 각도를 결정하는 데 도움을 준다.[11][12][13][14]
사다리꼴체는 대부분의 달팽이관 핵(CN) 섬유가 교차하는 곳으로, 소리 위치 파악에 도움이 된다.[15] CN은 복부(VCN)와 등쪽(DCN) 영역으로 나뉜다. VCN의 부시 세포는 타이밍 정보를, 별 모양(초퍼) 세포는 소리 스펙트럼을, 문어 세포는 청각 타이밍 코드를 해독한다. DCN의 방추형 세포는 정보를 통합하여 위치에 대한 스펙트럼 단서를 결정한다. 달팽이관 신경 섬유(30,000개 이상)는 각각 가장 민감한 주파수를 가지며 광범위한 수준에서 반응한다.[16][17]
신경 섬유 신호는 부시 세포에 의해 올리브 복합체의 이음향 영역으로 운반되는 반면, 신호 피크와 밸리는 별 모양 세포에 의해 기록되고 신호 타이밍은 문어 세포에 의해 추출된다. 가쪽 섬유띠 복부 핵은 위쪽 둔덕(IC)이 위상 및 토닉 반응을 모두 제공하여 진폭 변조 소리를 해독하는 데 도움을 준다. IC는 시각, 교뇌, 척수, 시상 등 다중 감각 통합을 담당하며, '놀람 반응'과 안구 반사에 관여한다. IC는 특정 진폭 변조 주파수에 반응하여 음조 감지를 가능하게 하고, 이음향 청취 시간 차이를 결정한다.[18]
내측 무릎 핵은 복부, 등쪽, 내측으로 나뉜다. 청각 겉질(AC)은 소리를 인식/지각하고 식별하며, 소리의 기원 위치를 식별한다. AC는 지형 주파수 맵을 가지며, 오른손 AC는 음성에, 왼손 AC는 소리의 미세한 순차적 차이에 더 민감하다.[19][20]
전전두피질 전내측 및 복외측은 각각 음조 공간에서의 활성화와 단기 기억 저장에 관여한다.[21] 헤슐 고랑/가로 측두 고랑은 베르니케 영역 및 기능을 포함하며, 감정-소리, 감정-얼굴 표정 및 소리-기억 프로세스에 관여한다. 내후각 피질은 '해마 시스템'의 일부로 시각 및 청각 기억을 돕고 저장한다.[22][23] 덧가쪽이랑(SMG)은 언어 이해를 돕고 동정심 있는 반응을 담당하며, 소리를 각이랑으로 단어에 연결하고 단어 선택에 도움을 준다. SMG는 촉각, 시각 및 청각 정보를 통합한다.[24][25]
5. 1. 청각 질환
청각계의 기능이 제대로 이루어지면 주변의 소리를 듣고, 처리하고, 이해할 수 있다. 소리를 감지하고 처리하며 이해하는 데 어려움이 있으면 의사 소통, 학습, 그리고 일상적인 업무를 효과적으로 수행하는 능력에 부정적인 영향을 미칠 수 있다.[42]어린이의 경우, 청각계 기능 손상에 대한 조기 진단 및 치료는 사회적, 학업적, 언어 발달의 중요한 이정표를 달성하는 데 매우 중요하다.[43]
청각계 손상에는 다음이 포함될 수 있다.
- 청성뇌간반응 및 신생아 청력 검사를 위한 ABR 청력 검사
- 청각 처리 장애
- 과청각증
- 이중청
- 이명
- 내이 현상
참조
[1]
서적
Kaplan medical anatomy flashcards
Kaplan Publishing
2013
[2]
서적
Barron's anatomy flash cards
Barron's Educational Series
2016
[3]
웹사이트
How Does My Hearing Work?
https://www.audiolog[...]
NZ Audiological Society
2016-03-27
[4]
논문
Prestin is the motor protein of cochlear outer hair cells
2000-05
[5]
논문
Tectorial membrane stiffness gradients
2007-09
[6]
논문
The effect of tectorial membrane and basilar membrane longitudinal coupling in cochlear mechanics
2010-03
[7]
논문
Dynamic material properties of the tectorial membrane: a summary
2003-06
[8]
논문
A targeted deletion in alpha-tectorin reveals that the tectorial membrane is required for the gain and timing of cochlear feedback
2000-10
[9]
논문
The effect of acoustic trauma on the tectorial membrane, stereocilia, and hearing sensitivity: possible mechanisms underlying damage, recovery, and protection
1988
[10]
논문
Basilar membrane and tectorial membrane stiffness in the CBA/CaJ mouse
2014-10
[11]
논문
Afferent projections of the superior olivary complex
2000-11
[12]
논문
Ascending efferent projections of the superior olivary complex
2000-11
[13]
논문
Organization of the human superior olivary complex
2000-11
[14]
논문
The superior olivary nucleus and its influence on nucleus laminaris: a source of inhibitory feedback for coincidence detection in the avian auditory brainstem
1999-03
[15]
논문
Temporal processing from the auditory nerve to the medial nucleus of the trapezoid body in the rat
2001-09
[16]
논문
Topographic organization of the dorsal nucleus of the lateral lemniscus in the cat
1999-05
[17]
서적
Integrative Functions in the Mammalian Auditory Pathway
Springer
2002
[18]
서적
The Inferior Colliculus
Springer
2005
[19]
논문
The cortical topography of tonal structures underlying Western music
2002-12
[20]
논문
Human primary auditory cortex: cytoarchitectonic subdivisions and mapping into a spatial reference system
2001-04
[21]
논문
Dual streams of auditory afferents target multiple domains in the primate prefrontal cortex
1999-12
[22]
논문
Left ventrolateral prefrontal cortex and the cognitive control of memory
2007-10
[23]
논문
Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps
2005-12
[24]
논문
A PET study of the functional neuroanatomy of writing impairment in Alzheimer's disease. The role of the left supramarginal and left angular gyri
1995-06
[25]
논문
Supramarginal gyrus involvement in visual word recognition
2009-10
[26]
논문
Development and regeneration of sensory transduction in auditory hair cells requires functional interaction between cadherin-23 and protocadherin-15
2010-08
[27]
논문
Integrating the biophysical and molecular mechanisms of auditory hair cell mechanotransduction
2011-11
[28]
웹사이트
Meddean – CN VIII. Vestibulocochlear Nerve
http://www.meddean.l[...]
2007-10-27
[29]
서적
Encyclopedia of Neuroscience
Academic Press
[30]
서적
Encyclopedia of Clinical Neuropsychology
Springer
2011
[31]
논문
Anatomophysiology of the central auditory nervous system: basic concepts
[32]
서적
The Inferior Colliculus
https://link.springe[...]
Springer Science+ Business Media, Incorporated
2022-05-17
[33]
논문
Anatomy of the auditory cortex
[34]
논문
Subdivisions of auditory cortex and levels of processing in primates
[35]
논문
Auditory processing in primate cerebral cortex
1999-04
[36]
논문
Subdivisions of auditory cortex and processing streams in primates
2000-10
[37]
논문
Architectonic identification of the core region in auditory cortex of macaques, chimpanzees, and humans
2001-12
[38]
논문
The neuroanatomical and functional organization of speech perception
2003-02
[39]
논문
Functional specialization in rhesus monkey auditory cortex
2001-04
[40]
논문
The harmonic organization of auditory cortex
2013-12
[41]
논문
The cortical organization of speech processing
2007-05
[42]
웹사이트
Hearing Loss
http://hearingloss.o[...]
National Academy on an Aging Society
2018-01-28
[43]
논문
Moderate-Severe Hearing Loss in Children: A Diagnostic and Rehabilitative Challenge
2017-12
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com