위둔덕
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
위둔덕은 중간뇌의 일부로, 시각 정보 처리와 눈 운동을 제어하는 데 중요한 역할을 하는 뇌 구조이다. 중간뇌 덮개에 위치하며, 아래둔덕과 함께 사두체를 형성한다. 위둔덕은 층상 구조를 가지며, 시각 정보 입력과 운동 관련 뇌 영역으로의 투사를 담당하는 표층과 심층으로 구분된다. 시각 자극에 대한 빠른 반응, 눈 운동 조절, 주의 산만과 관련된 기능을 수행하며, 영장류, 고양이, 박쥐, 설치류 등 다양한 종에서 발견된다. 포유류가 아닌 척추동물에서는 시각 덮개라고 불리며, 뇌간의 운동 조절에 기여한다.
더 읽어볼만한 페이지
- 사구체 - 아래둔덕
아래둔덕은 중뇌에 위치하여 청각 정보를 처리하고 음원 위치 결정 및 반사 작용에 중요한 역할을 하며, 다양한 뇌 영역과 연결되어 소리 분석 등의 기능을 수행한다. - 사구체 - 사구체 (신경해부학)
사구체는 신경해부학에서 뇌와 척수 밖에서 발견되는 신경 세포체의 집합으로, 감각 정보 처리, 자율신경계 조절 등 다양한 기능을 수행하며, 신경 세포체, 신경 섬유, 지지 세포로 구성된다. - 중간뇌 덮개 - 아래둔덕
아래둔덕은 중뇌에 위치하여 청각 정보를 처리하고 음원 위치 결정 및 반사 작용에 중요한 역할을 하며, 다양한 뇌 영역과 연결되어 소리 분석 등의 기능을 수행한다. - 중간뇌 덮개 - 사구체 (신경해부학)
사구체는 신경해부학에서 뇌와 척수 밖에서 발견되는 신경 세포체의 집합으로, 감각 정보 처리, 자율신경계 조절 등 다양한 기능을 수행하며, 신경 세포체, 신경 섬유, 지지 세포로 구성된다. - 중간뇌 - 흑색질
흑색질은 기저핵의 일부로 중뇌에서 가장 큰 핵이며, 안구 운동, 운동 계획, 보상 회로 등 다양한 뇌 기능에 관여하고, 특히 치밀부의 도파민성 뉴런은 운동 조절에 중요한 역할을 수행한다. - 중간뇌 - 배쪽 피개부
배쪽 피개부는 뇌의 주요 도파민 시스템으로서 보상, 동기 부여, 인지 기능, 약물 의존과 관련된 다양한 뇌 영역에 신경 섬유를 투사하며, 편도체로부터 감정적 정보를 처리하고 회피 및 공포 조건화에 관여하여 약물 중독, 조현병, ADHD와 같은 질환과 관련되는 중요한 역할을 한다.
| 위둔덕 | |
|---|---|
| 개요 | |
| 라틴어 명칭 | colliculus superior |
| 설명 | '위쪽 언덕'을 의미 |
| 구조 | |
| 위치 | 중뇌의 덮개 |
| 구성 요소 | (제공된 정보 없음) |
| 혈액 공급 | |
| 동맥 | (제공된 정보 없음) |
| 정맥 | (제공된 정보 없음) |
| 기능 | |
| 주요 역할 | 시각 반사 공간적 주의 조절 안구 운동 조절 머리와 몸의 방향 조절 청각 정보 처리 (일부) |
| 추가 정보 | |
| 관련 신경 | 눈돌림신경 (oculomotor nerve) |
| 외부 링크 | NeuroNames ID: 473 MeSH: Superior+Colliculus |
| 그레이 해부학 | '188, 806쪽' |
| 이미지 | |
 | |
| |
2. 구조
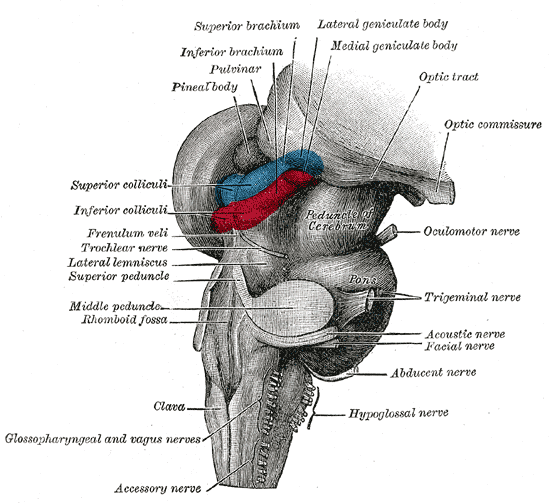
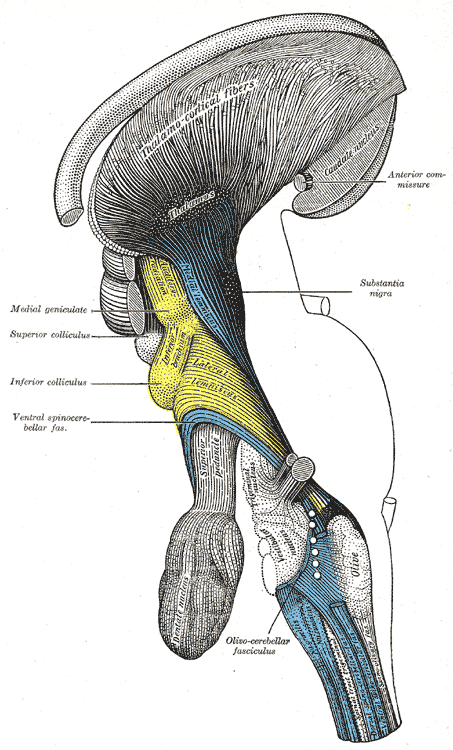
위둔덕은 등쪽 중간뇌에 쌍으로 존재하는 구조이며, 중간뇌 덮개의 일부이다. 두 개의 위둔덕은 솔방울샘과 뇌량 팽대의 아래쪽에 위치한다. 이는 시상의 시상 침윤핵에 의해 덮여 있으며, 시상의 내측 슬상핵은 각 위둔덕의 가쪽에 자리한다.[8] 두 개의 아래둔덕은 위둔덕 바로 아래에 위치하며, 아래둔덕과 위둔덕을 통틀어 사두체(四丘體, 라틴어: corpora quadrigemina)라고 부른다. 위둔덕은 아래둔덕보다 크지만, 아래둔덕이 더 두드러져 보인다.[9]
'''위둔덕 팔'''(영어: superior brachium)은 위둔덕에서 가쪽으로 뻗어 나와 시상 침윤핵과 내측 슬상핵 사이를 지나 시상으로 이어지는 신경 섬유 다발이다. 이 중 일부는 외측 슬상핵이라는 융기로 이어지고, 일부는 시신경로로 연결된다.
위둔덕은 인접한 구조인 '''파라비제미날 핵'''(영어: parabigeminal nucleus)과 관련이 있으며, 이 핵은 종종 위둔덕의 위성(satellite)으로 불린다. 시각 덮개에서는 이 인접 구조를 협부핵(영어: isthmic nucleus)이라고 한다.[10]
2. 1. 층판 구조
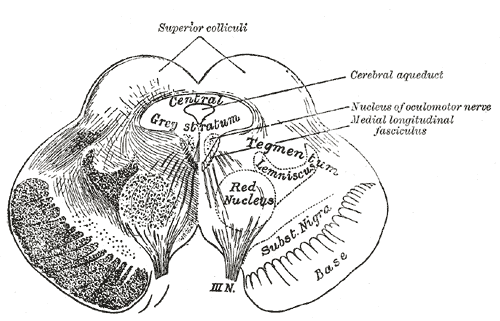
위둔덕은 시냅스 층상 구조를 가지고 있다.[51] 위둔덕과 시각 덮개의 미세 구조는 종에 따라 다르다. 일반적으로 위둔덕은 두 부분으로 나뉘는데, 하나는 주로 시각계로부터 입력을 받아 시각 반응을 보이는 표층(superficial layers)이고, 다른 하나는 다양한 유형의 입력을 받아 수많은 운동 관련 뇌 영역으로 신호를 보내는 심층(deep layers)이다. 이 두 영역의 구분은 매우 명확하고 일관적이어서, 일부 해부학자들은 이를 별개의 뇌 구조로 보아야 한다고 제안하기도 했다.포유류에서는 다음과 같이 7개의 층이 확인된다.[11]
'''표층 (Superficial Layers)'''
상위 3개 층을 표층이라고 부른다.
- '''층 I''' 또는 '''SZ''' (stratum zonale|스트라툼 조날레la, 띠층): 작은 수초화된 축삭과 가장자리 세포 및 수평 세포로 구성된 얇은 층이다.
- '''층 II''' 또는 '''SGS''' (stratum griseum superficiale|스트라툼 그리세움 수페르피시알레la, 표층 회백질층): 다양한 모양과 크기의 많은 뉴런이 포함되어 있다.
- '''층 III''' 또는 '''SO''' (stratum opticum|스트라툼 옵티쿰la, 시각층): 주로 시신경로에서 오는 축삭으로 구성된다.
'''중간층 (Intermediate Layers)'''
다음 두 층은 중간층이다.
- '''층 IV''' 또는 '''SGI''' (stratum griseum intermedium|스트라툼 그리세움 인테르메디움la, 중간 회백질층): 가장 두꺼운 층으로, 다양한 크기의 많은 뉴런으로 채워져 있다. 이 층은 종종 다른 모든 층을 합친 것만큼 두꺼우며, 때때로 "상부"와 "하부" 부분으로 나뉜다.
- '''층 V''' 또는 '''SAI''' (stratum album intermedium|스트라툼 알붐 인테르메디움la, 중간 백질층): 주로 다양한 출처에서 온 신경 섬유로 구성된다.
'''심층 (Deep Layers)'''
마지막 두 층은 심층이다.
- '''층 VI''' 또는 '''SGP''' (stratum griseum profundum|스트라툼 그리세움 프로푼둠la, 심층 회백질층): 느슨하게 배열된 뉴런과 수초화된 섬유로 구성된다.
- '''층 VII''' 또는 '''SAP''' (stratum album profundum|스트라툼 알붐 프로푼둠la, 심층 백질층): 중뇌수도 회백질 바로 위에 위치하며, 전적으로 신경 섬유로 구성된다.
2. 2. 신경 회로
얕은 층(표층)은 주로 망막, 시각 관련 대뇌 피질 영역(일차 시각 피질, 이차 시각 피질(브로드만 영역 18, 브로드만 영역 19), 전두엽 안구 운동 영역), 전개막, 그리고 이중반구 핵으로부터 신경 신호를 받는다. 망막에서의 입력은 얕은 층 전체에 걸쳐 양쪽에서 오지만, 반대쪽에서 오는 신호가 더 광범위하다.얕은 층이 주로 시각 정보를 받는 것과 달리, 중간층과 깊은 층은 매우 다양한 감각 및 운동 구조로부터 신호를 받는다. 대부분의 대뇌 피질 영역이 이 층들로 신호를 보내지만, 특정 감각이나 운동을 직접 담당하는 영역보다는 여러 정보를 통합하는 "연합" 영역에서의 입력이 더 강한 경향이 있다.[12] 그러나 어떤 피질 영역이 연결되고 그 신호의 강도는 동물 종에 따라 다르다.[13] 또 다른 중요한 입력은 기저핵의 일부인 흑질 망상부에서 오는데, 이는 억제성 신경전달물질인 GABA를 사용하며 위둔덕의 활동을 조절하는 "게이팅(gating, 문지기)" 역할을 하는 것으로 여겨진다. 중간층과 깊은 층은 또한 얼굴의 체성감각 정보를 전달하는 척수 삼차 신경 핵, 시상 하부, 불확실대, 시상, 그리고 하구로부터도 입력을 받는다.
입력 경로뿐만 아니라 출력 경로도 층별로 다르다. 얕은 층의 가장 중요한 출력 중 하나는 시상의 베개와 외측 중간 영역으로 가며, 이 신호는 다시 안구 운동을 제어하는 대뇌 피질 영역으로 전달된다. 얕은 층은 또한 전개막 핵, 시상의 외측 슬상체, 그리고 이중반구 핵으로도 신호를 보낸다. 깊은 층에서의 출력은 더 광범위하다. 뇌간과 척수로 내려가는 두 개의 큰 하행 경로와 안구 운동 생성에 관여하는 여러 경로를 포함하여, 다양한 감각 및 운동 중심부로 향하는 수많은 상행 신호 전달 경로가 있다.
위둔덕과 하구 모두 중앙 교뇌 망상체와 척수로 하행 신호를 보내며, 이를 통해 대뇌 피질의 복잡한 처리 과정을 거치지 않고도 자극에 빠르게 반응할 수 있다.
사람의 위둔덕은 사카데(단속성 안구 운동)와 눈과 머리의 움직임을 조화시키는 데 중요한 역할을 한다. 위둔덕으로 들어오는 구심성 신경 섬유는 대뇌 피질, 하구, 망막, 기저핵, 척수 등에서 시작된다. 위둔덕에서 나가는 원심성 신경 섬유는 방정중교뇌망상체(PPRF), 척수 및 다른 부위로 향한다. 인간을 포함한 많은 척추동물에서 중뇌로 전달된 감각 정보는 시상을 거쳐 대뇌 피질에서 처리되지만, 위둔덕은 대뇌 피질의 개입 없이도 안구 운동을 조절할 수 있다.
위둔덕 내에서도 시각 정보를 받는 것은 주로 얕은 층이며, 깊은 층은 청각 및 체성감각 입력을 받아 뇌의 여러 감각 운동 영역과 연결된다. 전체적으로 위둔덕은 머리와 눈을 시각적 또는 청각적 자극이 있는 방향으로 향하게 하는 역할을 한다.[75][76][77][78]
2. 3. 출력
위둔덕의 얕은 영역과 깊은 영역은 각각 독특한 출력 경로를 가진다.얕은 층의 가장 중요한 출력 중 하나는 시상의 베개와 외측 중간 영역으로 향하며, 이는 다시 안구 운동 제어에 관여하는 대뇌 피질 영역으로 투사된다. 또한 얕은 층은 전개막 핵, 시상의 외측 슬상체, 그리고 이중반구 핵으로도 투사한다.
깊은 층에서의 투사는 더 광범위하다. 다양한 감각 및 운동 중심부로 향하는 수많은 상행 투사가 있으며, 뇌간과 척수로 이동하는 두 개의 큰 하행 경로와 안구 운동 생성에 관여하는 여러 경로를 포함한다.
위둔덕의 얕은 층과 깊은 층 모두 중앙 교뇌 망상체와 척수로 하행 투사를 보내며, 이를 통해 대뇌 피질의 처리를 거치기 전에 자극에 더 빠르게 반응할 수 있다.
사람의 상구(위둔덕)는 특히 사카데안구 운동과 눈과 머리의 협응에 중요한 역할을 한다. 상구에서 나가는 원심성 섬유는 방정중교뇌망상체(PPRF), 척수, 그리고 다른 부위들에 도달한다. 중뇌로 전달된 감각 정보는 보통 시상을 거쳐 대뇌 피질에서 처리되지만, 상구는 피질의 개입 없이도 안구 운동을 조절할 수 있다.
상구의 얕은 층은 주로 시각 정보를 처리하는 반면, 깊은 층은 청각 및 체성 감각 입력을 받아 뇌의 여러 감각 운동 영역과 연결된다. 전체적으로 상구는 머리와 눈을 시각 또는 청각 자극이 있는 방향으로 향하게 하는 기능을 수행한다.[75][76][77][78]
2. 4. 모자이크 구조
윗둔덕의 층들은 매끄러운 단일 층이 아니라, 개별적인 기둥들이 벌집처럼 배열된 구조를 이룬다.[14] 이러한 기둥 구조는 부곁두덩핵에서 기원하는 콜린성 입력 신호가 덮개의 위에서 아래까지 일정한 간격으로 분포하며 종말 묶음을 형성하는 모습에서 가장 뚜렷하게 나타난다.[15] 또한 칼레티닌, 파르부민, GAP-43, NMDA 수용체와 같은 여러 신경화학적 지표들과 뇌간 및 간뇌의 여러 다른 뇌 구조와의 연결에서도 이러한 비균질성이 확인된다.[16] 전체 기둥의 수는 약 100개 정도로 추정된다.[14] 이 기둥 구조의 구체적인 기능은 아직 명확히 밝혀지지 않았지만, 최근 연구에 따르면 콜린성 입력이 덮개 내에서 특정 신호만 선택하는 '승자 독식' 방식의 신경 회로와 관련될 수 있다는 증거가 제시되고 있다.이러한 구획화는 포유류와 비포유류를 포함하여 조사된 모든 종에서 관찰되지만, 배열의 세부적인 모습에는 약간의 차이가 있다.[15] 예를 들어, 토끼나 사슴과 같이 눈이 옆에 있고 줄무늬형 망막을 가진 종에서는 구획이 위둔덕 전체에 걸쳐 나타난다. 반면, 중심와가 있는 종에서는 위둔덕의 앞쪽(머리쪽) 부분에서 이러한 구획화가 뚜렷하지 않게 나타난다. 이 앞쪽 부분에는 눈이 한 곳을 응시하며 고정되어 있을 때 지속적으로 활동하는 '고정' 뉴런들이 많이 분포한다.
3. 기능
위둔덕 기능에 대한 연구는 시대에 따라 초점이 변화해 왔다. 초기에는 어류, 개구리 등 비포유류의 주요 시각 중심으로 여겨졌으나,[17] 1970년대 이후 포유류 연구에서는 안구 운동 제어 역할이 강조되었다. 1990년대 후반부터는 머리와 눈의 통합된 움직임인 '시선 이동'과 다감각 통합 등 더 넓은 기능에 대한 관심이 높아졌다.[18]
행동 연구에 따르면, 위둔덕은 물체를 인식하는 데는 필수적이지 않지만, 특정 물체를 향해 행동하도록 지시하는 능력에 중요한 역할을 한다. 이러한 능력은 대뇌 피질이 손상된 경우에도 유지될 수 있다.[18] 예를 들어, 시각 피질이 손상된 고양이는 물체를 알아보지는 못하지만 움직이는 자극을 따라갈 수 있다. 반면, 위둔덕의 일부가 손상되면 해당 방향의 물체에 반응하지 못하게 된다.
전반적으로 위둔덕은 시각, 청각, 체성 감각 정보를 통합하여[75][76][77][78] 해당 자극이 있는 방향으로 머리와 눈을 돌리는 반사적인 움직임을 유도하는 데 핵심적인 역할을 한다. 이러한 기능은 대뇌 피질의 개입 없이도 이루어질 수 있다.
3. 1. 안구 운동 (Eye Movements)
영장류에서 눈 운동은 여러 유형으로 나눌 수 있다. 눈이 움직이지 않는 물체를 향하고 머리의 움직임을 보상하기 위해 눈 운동만 하는 주시, 눈이 움직이는 물체를 따라가기 위해 눈이 꾸준히 움직이는 추종 운동, 눈이 한 위치에서 다른 위치로 매우 빠르게 움직이는 빠른 눈 운동(사카데), 그리고 단일 양안 시력을 얻거나 유지하기 위해 눈이 동시에 반대 방향으로 움직이는 폭주가 있다. 위둔덕(상구)은 이 모든 종류의 눈 운동에 관여하지만, 특히 빠른 눈 운동에서의 역할이 가장 집중적으로 연구되었다.[19][20][21]뇌 양쪽에 하나씩 있는 두 개의 위둔덕 각각은 시야의 절반을 나타내는 2차원 지도를 포함한다. 시력에서 가장 민감한 영역인 중심와는 이 지도의 앞쪽 가장자리에 해당하고, 시야의 주변부는 뒤쪽 가장자리에 해당한다. 눈 운동은 위둔덕의 깊은 층에 있는 신경 세포(뉴런)의 활동에 의해 유발된다. 주시 중에는 앞쪽 가장자리, 즉 중심와 영역 근처의 뉴런이 지속적으로 활성화된다. 추종 운동 중에는 앞쪽 가장자리에서 약간 떨어진 뉴런이 활성화되어 작은 눈 움직임을 일으킨다. 빠른 눈 운동의 경우, 뉴런은 눈이 빠르게 움직여 향할 지점을 나타내는 영역에서 활성화된다. 빠른 눈 운동 직전에는 목표 위치에 해당하는 위둔덕 영역의 활동이 빠르게 증가하고 다른 부분의 활동은 감소한다. 이 활동은 비교적 넓게 퍼져 있어서, 특정 빠른 눈 운동에 대한 활동 모습은 위둔덕 지도 위에 상당한 부분을 차지하는 '언덕' 형태를 이룬다. 이 '언덕'의 정점이 가리키는 위치가 빠른 눈 운동의 목표 지점을 나타낸다.[22]
위둔덕은 시선 이동의 목표 지점을 부호화(encoding)하지만, 그 목표 지점에 도달하기 위해 필요한 정확한 움직임 자체를 지정하지는 않는 것으로 보인다.[23] 시선 이동을 머리 움직임과 눈 움직임으로 나누고, 빠른 눈 운동 동안 눈의 정확한 궤적을 결정하는 것은 위둔덕에서 오는 신호와 다른 경로의 신호를 하위 운동 영역에서 통합하는 과정에 달려 있으며, 이 과정은 아직 완전히 밝혀지지 않았다. 움직임이 어떻게 유발되든 위둔덕은 이를 '망막 중심 좌표계'로 부호화한다. 즉, 위둔덕 활동 '언덕'의 위치는 망막 상의 고정된 위치에 해당한다. 이는 위둔덕의 한 지점을 자극했을 때 초기 눈의 방향에 따라 다른 방향으로 시선이 이동할 수 있다는 관찰 결과와 모순되는 것처럼 보일 수 있다. 그러나 이는 자극된 망막 위치가 목표 위치, 눈의 방향, 그리고 눈의 구형 기하학적 구조 사이의 비선형적인 함수 관계 때문인 것으로 밝혀졌다.[24]
과거에는 위둔덕이 단순히 눈 운동 명령을 내리고 실행은 다른 구조에 맡기는지, 아니면 빠른 눈 운동의 실행 과정에 적극적으로 참여하는지에 대한 논란이 있었다. 1991년 무뇨스(Munoz) 등은 빠른 눈 운동 동안 위둔덕의 활동 '언덕'이 점진적으로 이동하며, 이는 눈이 목표 위치에서 벗어난 정도를 실시간으로 반영한다고 주장했다(이동 언덕 가설).[25] 현재 지배적인 견해는 활동 '언덕'이 빠른 눈 운동 동안 약간 이동하기는 하지만, '이동 언덕' 가설에서 예측하는 것처럼 꾸준하고 비례적인 방식으로 이동하지는 않는다는 것이다.[26] 그러나 이동하는 활동 언덕은 다른 역할을 할 수 있다. 최근 연구에서는 눈이 천천히 움직이는 동안에도 별도의 빠른 눈 운동 목표 지점이 유지될 때, 시각적 기억과 관련된 활동 언덕이 지속적으로 움직이는 것이 관찰되었다.[27]
위둔덕의 운동 관련 영역에서 나가는 신호는 중뇌와 뇌간에 있는 일련의 핵으로 전달된다. 이 핵들은 위둔덕이 사용하는 '위치' 부호를 눈 운동 신경 세포가 사용하는 '속도' 부호로 변환하는 역할을 한다. 눈 운동은 세 쌍의 외안근에 의해 조절되므로, 최종적으로 눈 운동은 데카르트 좌표계와 유사한 방식으로 부호화된다.
위둔덕은 망막으로부터 직접적이고 강한 입력을 받지만, 영장류에서는 주로 눈 운동을 결정하는 데 관여하는 대뇌 피질의 여러 영역으로부터 제어를 받는다.[28] 운동 피질의 일부인 전두안야는 의도적인 빠른 눈 운동을 유발하는 데 관여하고, 인접한 보조 안야는 여러 빠른 눈 운동을 순서대로 구성하는 데 관여한다. 뇌 뒤쪽에 위치한 두정 안야는 주로 시야의 변화에 반응하여 일어나는 반사적인 빠른 눈 운동에 관여한다. 최근 연구[29][30]는 일차 시각 피질(V1)이 외부 시각 자극에 대해 V1 현저성 가설에 따라 반사적인 눈 운동을 유도할 수 있음을 시사한다.[31]
위둔덕의 얕은 층은 주로 시각 정보를 받지만, 깊은 층은 청각 및 체성감각 입력도 받으며 뇌의 많은 감각 운동 영역과 연결되어 있다. 위둔덕 전체는 보고 들은 자극 쪽으로 머리와 눈을 향하게 하는 데 중요한 역할을 하는 것으로 생각된다.[32][33][34][35][75][76][77][78] 위둔덕은 아래둔덕(하구)으로부터 청각 정보를 받으며, 이 정보는 기존의 시각 정보와 통합되어 복화술 효과를 만들어낸다.
사람의 위둔덕은 빠른 눈 운동(사카데)과 눈과 머리의 협응 운동에 관여한다. 위둔덕으로 들어오는 신경 섬유(구심성 섬유)는 대뇌 피질, 아래둔덕, 망막, 기저핵, 척수 등에서 시작된다. 위둔덕에서 나가는 신경 섬유(원심성 섬유)는 방정중교뇌망상체(PPRF), 척수 및 다른 뇌 영역으로 이어진다. 인간을 포함한 많은 척추동물에서 중뇌로 전달된 감각 정보는 시상을 거쳐 대뇌 피질에서 처리되지만, 위둔덕은 대뇌 피질의 관여 없이도 눈 운동을 조절할 수 있다.
3. 2. 주의 산만 (Distractibility)
상구는 눈 운동과 관련된 기능 외에도 주의 산만 회로망에서 중요한 역할을 하는 것으로 보인다. 주의 산만이 증가하는 것은 정상적인 노화 과정에서 나타나기도 하며,[36] ADHD를 포함한 여러 의학적 상태의 핵심 특징이기도 하다.[37]연구에 따르면 여러 동물 종에서 상구에 병변이 생기면 주의 산만이 증가할 수 있다.[38][39] 인간의 경우에도 전전두피질에서 상구에 대한 억제 조절을 제거하여 상구의 활동을 증가시키면 주의 산만이 증가하는 것으로 나타났다.[40]
ADHD의 동물 모델인 자발 고혈압 쥐를 이용한 연구에서도 상구 의존적인 행동 변화[41][42] 및 생리학적 변화가 관찰되었다.[42] 또한, ADHD의 주요 치료제로 사용되는 암페타민은 건강한 동물에서 상구의 활동을 억제하는 효과를 보인다.[43]
4. 다양한 종 (Other Animals)
(내용 없음 - 주어진 정보와 제약 조건 하에 해당 섹션에 작성할 내용이 없습니다.)
4. 1. 영장류 (Primates)
일반적으로 영장류의 상구(superior colliculus)는 다른 포유류와 달리 반대쪽 눈이 보는 시야에 대한 완전한 지도를 포함하지 않는다는 특징이 있다.[44][79] 대신, 시각 피질이나 외측 슬상체처럼 각 상구는 시야의 반대쪽 절반만을 중간선까지 나타내며, 같은 쪽(동측) 시야의 절반에 대한 정보는 포함하지 않는다.[44][79] 이러한 기능적 특징은 영장류의 경우, 망막 신경절 세포 중 시야의 귀쪽(이측 또는 측두엽 쪽) 절반을 담당하는 세포와 반대쪽 상구 사이에 해부학적 연결이 없기 때문이다.[44][79] 반면, 다른 포유류에서는 반대쪽 눈의 망막 전체에서 오는 신경절 세포가 반대쪽 상구로 투사된다.영장류와 다른 포유류 사이의 이러한 상구 연결 방식의 차이는 1986년 오스트레일리아의 신경과학자 잭 페티그루(Jack Pettigrew)가 제안한 비행 영장류 가설(flying primates theory)을 뒷받침하는 주요 증거 중 하나로 여겨진다.[45][80] 페티그루는 박쥐의 일종인 과일박쥐가 망막과 상구 사이의 해부학적 연결 패턴에서 영장류와 유사하다는 점을 발견하고 이 가설을 제시했다.[45][80]
4. 2. 고양이 (Cats)
고양이의 경우 상구는 그물체 형성을 통해 투사되며 뇌간의 운동 신경원과 상호 작용한다.[46]4. 3. 박쥐 (Bats)
박쥐는 일반적으로 알려진 것과 달리 앞을 보지 못하는 것은 아니지만, 길을 찾거나 먹이를 잡을 때 시각보다는 반향 정위 능력에 훨씬 더 크게 의존한다. 박쥐는 음파(짹짹거리는 소리)를 내보내고 그 소리가 주변 사물에 부딪혀 돌아오는 메아리를 분석하여 주변 환경에 대한 정보를 얻는다. 박쥐의 뇌는 이러한 반향 정위 과정을 처리하기 위해 매우 특화되어 있으며, 이러한 전문화의 일부는 위둔덕(상구)에서 나타난다.[47]박쥐의 경우, 시각 정보가 전달되는 망막 투영은 위둔덕 표면 바로 아래의 얇은 영역만을 차지한다. 대신, 청각 영역으로부터 광범위한 정보를 입력받으며, 이 정보는 귀, 머리, 또는 몸 전체를 특정 방향으로 향하게 하는 운동 영역으로 전달된다. 서로 다른 방향에서 돌아오는 메아리는 위둔덕의 각기 다른 위치에 있는 뉴런들을 활성화시키고,[48] 이렇게 활성화된 위둔덕 뉴런은 박쥐가 다음에 내보낼 소리의 특성에 영향을 미친다. 따라서 박쥐의 위둔덕은 다른 동물들이 시각 정보를 이용해 행동을 조절하는 것과 유사하게, 청각 정보를 바탕으로 박쥐의 행동을 유도하는 중요한 기능을 수행한다고 볼 수 있다.
박쥐는 크게 두 그룹으로 나뉜다. 전 세계적으로 가장 흔하게 발견되는 소형익수아목 박쥐와 아시아, 아프리카, 오스트랄라시아 등지에서 발견되는 과일 박쥐인 대형익수아목이 있다. 한 종을 제외하고, 대형박쥐는 반향 정위를 사용하지 않으며, 대신 잘 발달된 시각 능력에 의존하여 이동한다. 이러한 대형박쥐의 위둔덕에 있는 뉴런들은 고양이나 영장류의 위둔덕처럼 시각 수용장이 망막의 정보를 정확하게 반영하는 지도를 형성한다.
4. 4. 설치류 (Rodents)
설치류의 상구는 감각 정보를 바탕으로 접근하거나 회피하는 행동을 조절하는 것으로 여겨진다.[49][50] 특히 쥐의 상구에 대한 연구는 회로 분석 도구를 사용하여 중요한 기능들을 밝혀냈다.[51] 여러 연구를 통해, 쥐가 먹이를 잡거나 포식자를 피하는 행동을 시작하게 하는 상구 내의 'Ying-Yang 회로 모듈'이라는 특정 신경 회로 세트가 확인되었다.[52][53][54][55] 더 나아가, 단일 세포 RNA 시퀀싱 기법을 이용하여 상구 뉴런의 유전자 발현 패턴을 분석하고, 이 Ying-Yang 회로 모듈을 구성하는 세포들의 고유한 유전자 표지자를 찾아냈다.[56]4. 5. 기타 척추동물 (Other Vertebrates)

포유류가 아닌 척추동물의 중뇌에서 위둔덕에 해당하는 영역은 시각 덮개(optic tectum|옵틱 텍텀eng)라고 불린다. 이 구조는 뇌의 날개판에서 발달하며, 포유류가 아닌 동물의 주요 시각 중심 역할을 한다. 시각 덮개는 다양한 크기의 물체를 인식하고 이에 반응하는 데 중요한 역할을 하며, 이러한 과정은 글루탐산과 같은 흥분성 신경전달물질에 의해 촉진된다.[58]
시각 덮개는 다양한 척추동물 뇌의 기본적인 구성 요소 중 하나이다.[64] 구조적으로는 여러 층으로 이루어져 있는데, 표면층에는 시신경 입력이 밀집되어 있고 깊은 층에는 다른 감각 정보 입력이 강하게 들어온다는 공통점이 있다. 하지만 전체 층의 수나 세포 종류의 수는 종에 따라 매우 다양하다.[65]
시각 덮개는 좁은 등쪽 핵(isthmic nucleus|이스믹 뉴클리어스eng)이라는 인접 구조와 밀접하게 연관되어 있으며, 이 핵은 시각 덮개의 기능에 매우 중요한 기여를 하는 것으로 알려져 많은 연구가 이루어졌다.[66] (포유류의 상구에서는 부방차핵이 이와 유사한 구조이다.) 좁은 등쪽 핵은 '큰 세포가 있는 부분'인 '''좁은 등쪽 핵 대세포 부위'''(pars magnocellularis|파르스 마그노셀룰라리스lat, Imc)와 '작은 세포가 있는 부분'인 '''좁은 등쪽 핵 소세포 부위'''(pars parvocellularis|파르스 파르보셀룰라리스lat, Ipc)로 나뉜다. 시각 덮개, Ipc, Imc 세 영역 간의 연결은 지형적으로 이루어진다. 즉, 시각 덮개 표면층의 특정 지점 뉴런은 Ipc와 Imc의 해당 지점으로 신호를 보낸다. Ipc로 가는 신호는 매우 집중되는 반면, Imc로 가는 신호는 다소 분산되는 경향이 있다. Ipc는 Imc와 시각 덮개 양쪽으로 매우 집중된 콜린성 신호를 보내며, 시각 덮개에서는 이 신호가 위아래로 길게 뻗어 전체 기둥을 연결한다. 반대로 Imc는 Ipc와 시각 덮개로 GABA성 신호를 보내는데, 이 신호는 옆으로 넓게 퍼져 대부분의 시각 영역에 영향을 미친다. 이러한 덮개-Ipc-Imc 회로는 시각 덮개의 특정 영역이 활성화될 때, 주변의 작은 영역은 강하게 흥분시키고 멀리 떨어진 넓은 영역은 억제하는 순환 피드백을 만들어낸다.
시각 덮개는 물고기의 수영, 새의 비행, 개구리의 먹이 공격 등 다양한 생존 행동과 반응에 관여한다.
박쥐의 경우, 상구(시각 덮개에 해당)는 반향 정위 시스템을 구성하는 초음파 발성과 귀의 움직임에 영향을 미치는 것으로 알려져 있다.[81]
4. 5. 1. 어류 (Fish)
포유류가 아닌 척추동물에서 위둔덕에 해당하는 중뇌 영역은 '''시개'''(optic tectumeng)라고 불린다. 양서류, 파충류, 어류에서는 시개가 가장 중요한 시각 정보 처리 영역이다. 하지만 그 기능에 대해서는 아직 완전히 밝혀지지 않았다. 시개는 포식자와 먹이를 구별하는 데 필요하며, 위협을 피하거나 먹이를 사냥하는 행동과 관련이 있는 것으로 알려져 있다.
칠성장어는 초기 척추동물의 뇌 구조를 많이 반영하는 비교적 단순한 뇌를 가지고 있어 뇌 연구에 널리 활용된다. 1960년대 칼 로바이넨의 선구적인 연구[68]를 시작으로, 1970년대부터는 스톡홀름 카롤린스카 연구소의 스테인 그리너와 그의 동료들이 척추동물의 운동 제어 원리를 밝히기 위해 칠성장어를 모델 시스템으로 사용해왔다.[69] 다른 시스템과 마찬가지로 척수 내의 신경 회로는 수영의 기초적인 리듬 운동 패턴을 생성할 수 있는 것으로 보이며, 이러한 회로는 기저핵과 시개를 포함한 상위 뇌 구조의 영향을 받는 뇌간 및 중뇌의 특정 운동 영역에 의해 조절된다.[70]
2007년에 발표된 칠성장어 시개 연구에서는[71] 시개에 전기 자극을 주면 안구 운동, 몸통을 옆으로 구부리는 운동, 또는 수영 활동을 유발할 수 있다는 사실이 밝혀졌다. 이때 나타나는 운동의 종류, 크기, 방향은 시개 내에서 자극이 가해진 위치에 따라 달라졌다. 이는 칠성장어도 다른 동물들처럼 목표 지향적인 움직임을 만들어낸다는 가설을 뒷받침하는 결과로 해석되었다.
4. 5. 2. 조류 (Birds)
새의 시각 덮개는 비행과 관련이 있으며, 뇌의 가장 큰 구성 요소 중 하나이다. 조류의 시각 처리에 대한 연구는 인간을 포함한 포유류의 시각 처리 이해를 높이는 데 기여했다.[72]
4. 5. 3. 파충류 (Reptiles)
포유류가 아닌 척추동물, 예를 들어 양서류, 파충류, 어류에서 중뇌의 시각 정보 처리 영역은 '''시개'''(optic tectumeng)라고 불린다. 특히 파충류에서는 시개가 가장 중요한 시각 정보 처리 영역으로 기능한다. 시개는 포식자와 먹이를 구별하고, 도망치거나 사냥하는 행동과 관련된 역할을 하는 것으로 알려져 있지만, 그 기능 전반은 아직 완전히 밝혀지지 않았다.비단뱀이나 살무사처럼 적외선을 감지하여 '보는' 능력을 가진 뱀의 경우, 시각 정보 처리가 독특하게 이루어진다. 이 뱀들은 눈의 시신경 대신 삼차 신경을 통해 초기 적외선 감각 정보를 받아들인다. 이후의 정보 처리 과정은 일반적인 시각 경로와 유사하며, 여기서도 시개가 중요한 역할을 담당한다.
5. 추가 이미지 (Additional Images)
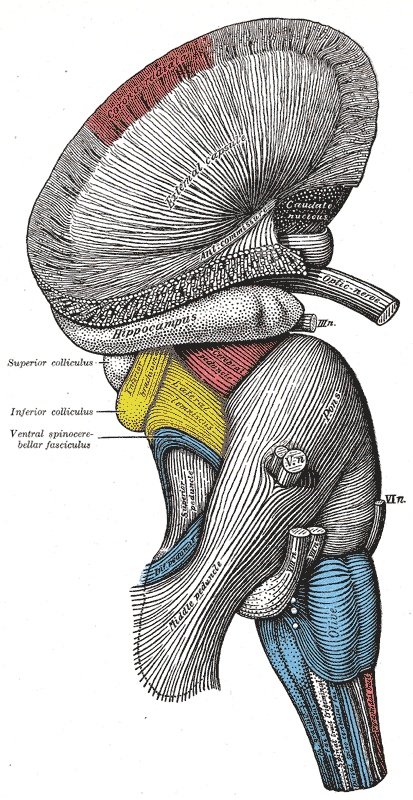
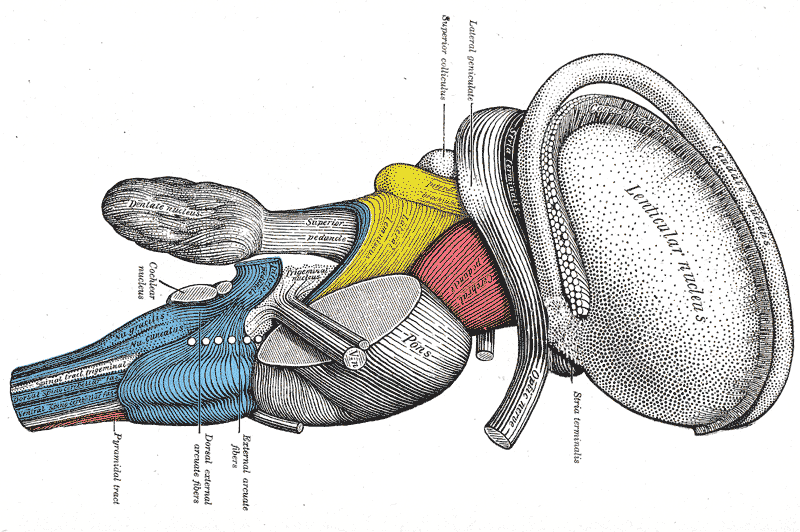
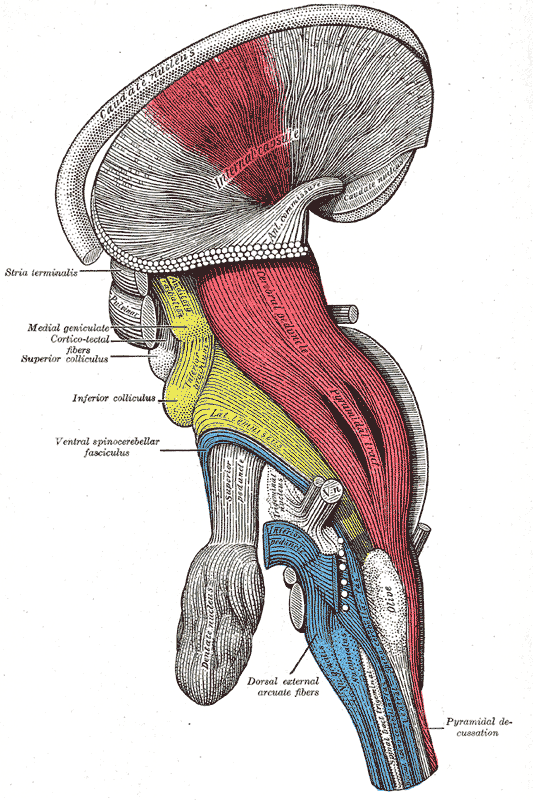
참조
[1]
서적
Fundamental neuroscience
Academic Press
2013
[2]
논문
Control from below: the role of a midbrain network in spatial attention.
2011-06
[3]
논문
Mosaic Organization of Body Pattern Control in the Optic Lobe of Squids
2017-01-25
[4]
논문
Circuits for Action and Cognition: A View from the Superior Colliculus.
2017-09-15
[5]
문서
Wallace et al., 1998
[6]
문서
Gandhi et al., 2011
[7]
문서
Lunenburger et al., 2001
[8]
서적
Last's Anatomy
Elsevier Australia
[9]
웹사이트
IX. Neurology. 4b. The Mid-brain or Mesencephalon. Gray, Henry. 1918. Anatomy of the Human Body.
https://www.bartleby[...]
2019-10-10
[10]
논문
The nucleus isthmi and dual modulation of the receptive field of tectal neurons in non-mammals
2003
[11]
문서
Huerta & Harting, 1984
[12]
논문
Cortical efferents of the perirhinal, postrhinal, and entorhinal cortices of the rat
2009-12
[13]
논문
Topographic organization of somatosensory corticotectal influences in cat
[14]
문서
Chavalier & Mana, 2000
[15]
문서
Illing, 1996
[16]
문서
Mana & Chevalier, 2001
[17]
논문
Coding Schemes in the Archerfish Optic Tectum
2018
[18]
문서
Sprague, 1996
[19]
논문
Circuits for Action and Cognition: A View from the Superior Colliculus
2017
[20]
논문
Activity in the human superior colliculus relating to endogenous saccade preparation and execution
2015
[21]
논문
The superior colliculus and eye movement control
1996
[22]
논문
Evidence against a moving hill in the superior colliculus during saccadic eye movements in the monkey
2002
[23]
문서
Sparks & Gandhi, 2003
[24]
문서
Klier et al., 2001
[25]
문서
Munoz et al., 1991
[26]
문서
Soetedjo et al., 2002
[27]
문서
Dash et al., 2015
[28]
문서
Pierrot-Deseilligny et al., 2003
[29]
논문
Bottom-up saliency and top-down learning in the primary visual cortex of monkeys
2018-10-09
[30]
논문
Neural Activities in V1 Create a Bottom-Up Saliency Map
2012-01-12
[31]
논문
A saliency map in primary visual cortex
https://www.scienced[...]
2002-01-01
[32]
문서
Kustov & Robinson, 1996
[33]
문서
Klier et al., 2003
[34]
문서
Krauzlis et al., 2004
[35]
문서
Sparks, 1999
[36]
논문
Age-related differences in distractibility and response to methylphenidate in monkeys.
1998-03-01
[37]
서적
Developmental neuropsychiatry
Guildford Press
[38]
논문
Visual orientation in the rat: A dissociation of deficits following cortical and collicular lesions
1978-03-01
[39]
논문
Go-left go-right discrimination performance and distractibility following lesions of prefrontal cortex or superior colliculus in stumptail macaques
1978-01-01
[40]
논문
A direct prefrontotectal tract against distractibility in the human brain
2003-04-01
[41]
논문
Abnormal air righting behaviour in the spontaneously hypertensive rat model of ADHD
2011-11-01
[42]
논문
Altered visual processing in a rodent model of Attention-Deficit Hyperactivity Disorder
https://kclpure.kcl.[...]
[43]
논문
Enhanced visual responses in the superior colliculus in an animal model of attention-deficit hyperactivity disorder and their suppression by d-amphetamine
[44]
문서
Lane et al., 1973
[45]
문서
Pettigrew, 1986
[46]
논문
Tectal influences on cat ocular motoneurons
1974
[47]
문서
Ulanovsky & Moss, 2008
[48]
문서
Valentine & Moss, 1997
[49]
논문
Output pathways from the rat superior colliculus mediating approach and avoidance have different sensory properties
1990-08
[50]
논문
Neural Correlates of Active Avoidance Behavior in Superior Colliculus
2010-06-23
[51]
논문
The Mouse Superior Colliculus: An Emerging Model for Studying Circuit Formation and Function.
2018
[52]
논문
A parvalbumin-positive excitatory visual pathway to trigger fear responses in mice
https://www.science.[...]
2015-06-26
[53]
논문
Divergent midbrain circuits orchestrate escape and freezing responses to looming stimuli in mice
2018-12
[54]
논문
A subcortical excitatory circuit for sensory-triggered predatory hunting in mice
http://www.nature.co[...]
2019-06
[55]
논문
The tectonigral pathway regulates appetitive locomotion in predatory hunting in mice
2021-12
[56]
논문
Transcriptomic encoding of sensorimotor transformation in the midbrain
2021-07-28
[57]
논문
NOGO-A induction and localization during chick brain development indicate a role disparate from neurite outgrowth inhibition
[58]
논문
An evaluation of L-glutamate as the transmitter released from optic nerve terminals of the pigeon.
1976
[59]
논문
Spontaneous Activity in the Zebrafish Tectum Reorganizes over Development and Is Influenced by Visual Experience
[60]
논문
Neural Circuits Underlying Visually Evoked Escapes in Larval Zebrafish
[61]
논문
Hypothalamic Projections to the Optic Tectum in Larval Zebrafish
[62]
논문
Visuomotor Behaviors in Larval Zebrafish after GFP-Guided Laser Ablation of the Optic Tectum
2003
[63]
논문
Sensorimotor Decision Making in the Zebrafish Tectum
2015
[64]
문서
Maximino, 2008
[65]
문서
Northcutt, 2002
[66]
논문
Nucleus Isthmi Is Required to Sustain Target Pursuit during Visually Guided Prey-Catching
2019-06-03
[67]
문서
Hartline et al., 1978
[68]
간행물
Neurobiology of lampreys. Physiological reviews 59:1007-1077.
1979
[69]
문서
Grillner, 2003
[70]
간행물
Thomas Graham Brown (1882--1965), Anders Lundberg (1920-), and the neural control of stepping. Brain Res Rev 59:74-95.
2008
[71]
문서
Saitoh et al., 2007
"#refSaitoh"
[72]
논문
The optic tectum of birds: mapping our way to understanding visual processing
2009-12
[73]
웹사이트
Primate Superior Colliculus
http://keck.ucsf.edu[...]
University of California
[74]
간행물
Multisensory Intergration in the Superior Colliculus of the Alert Cat
2005
[75]
간행물
Three-Dimensional Eye-Head Coordination Is Implemented Downstream From the Superior Colliculus
2003
[76]
간행물
Target selection and the superior colliculus: goals, choices and hypotheses
2004
[77]
간행물
Shared neural control of attentional shifts and eye movements
1996
[78]
간행물
Conceptual issues related to the role of the superior colliculus in the control of gaze
1999
[79]
간행물
The visuotopic organization of the superior colliculus of the owl monkey (Aotus trivirgatus) and the bush baby (Galago senegalensis)
1973
[80]
간행물
Flying primates? Megabats have the advanced pathway from eye to midbrain
1986
[81]
간행물
Spatially selective auditory responses in the superior colliculus of the echolocating bat
1997
[82]
서적
Fundamental neuroscience
https://archive.org/[...]
Academic Press
2013
[83]
저널
Control from below: the role of a midbrain network in spatial attention.
2011-06
[84]
저널
Mosaic Organization of Body Pattern Control in the Optic Lobe of Squids
2017-01-25
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com