쿠스쌀쥐
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
쿠스쌀쥐(Oryzomys couesi)는 오리조미스속에 속하는 쥐의 일종으로, 멕시코 남부에서 코스타리카, 콜롬비아에 걸쳐 널리 분포한다. 다양한 아종으로 분류되며, 털은 황갈색에서 붉은색을 띠고 배는 흰색 또는 황갈색이다. 반수생 생활을 하며, 물 속에서 수영과 잠수가 가능하다. 식물 재료, 작은 물고기, 곤충 등을 먹고 살며, 둥지를 짓고 야행성으로 활동한다. IUCN은 이 종을 "관심 필요"종으로 분류하지만, 서식지 파괴와 도입종에 의한 위협을 받기도 한다. 텍사스에서는 멸종 위협을 받고 있으며, 코수멜 개체군은 서식지 교란으로 인해 감소했다.
더 읽어볼만한 페이지
| 쿠스쌀쥐 - [생물]에 관한 문서 | |
|---|---|
| 일반 정보 | |
 | |
| 학명 | Oryzomys couesi |
| 명명자 | (Edward Richard Alston, 1877) |
| 보존 상태 | 최소 관심(LC) |
| 학명 이명 | Hesperomys couesi Alston, 1877 Oryzomys couesi Thomas, 1893 Oryzomys palustris couesi Hall, 1960 |
| 무게 | 43~82g |
| 몸길이 | 약 1m |
| 분포 | 쿠스쌀쥐 분포(붉은색) |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 목 | 쥐목 |
| 아목 | 쥐아목 |
| 상과 | 쥐상과 |
| 과 | 비단털쥐과 |
| 아과 | 목화쥐아과 |
| 족 | 쌀쥐족 |
| 속 | 쌀쥐속 |
| 형태 및 습성 | |
| 서식지 | 반수생 설치류 |
| 먹이 | 잡식성 (씨앗, 과일, 곤충) |
| 활동 | 주로 야행성이지만, 주간 활동도 관찰됨 |
| 번식 | 번식기는 다양하며, 한 번에 여러 마리의 새끼를 낳음 |
| 특징 | 물갈퀴가 있는 발 꼬리는 몸길이와 거의 같거나 약간 짧음 털은 거칠고 방수성이 있음 다양한 환경에 잘 적응함 수영과 잠수에 능숙함 |
| 분포 및 서식지 | |
| 분포 지역 | 멕시코 남부 중앙 아메리카 남아메리카 북서부 |
| 서식지 | 습지, 강둑, 관목 지대, 농경지 등 다양한 환경 |
| 보전 상태 | |
| IUCN 적색 목록 | 최소 관심(LC) |
| 위협 요인 | 서식지 파괴, 농업 확장, 수질 오염 등 |
| 기타 | |
| 참고 | 쿠스쌀쥐는 생태계에서 중요한 역할을 수행함 씨앗 분산, 곤충 개체수 조절 등에 기여함 인간 활동으로 인해 서식지가 위협받고 있으며, 보전 노력이 필요함 |
2. 분류
오리조미스속(Oryzomys) 내에서 '''''쿠스쌀쥐'''''(''Oryzomys couesi'')와 주변에 분포하는 좁은 범위의 최소 6종은 함께 ''O. couesi'' 그룹을 형성한다. 이 속의 8번째 종인 습지쌀쥐(''O. palustris'')는 자체 그룹의 유일한 구성원이다[6] (서부 개체군이 별도의 종, 즉 ''O. texensis''로 분류되지 않는 한).[7] ''Oryzomys''는 이전에 다른 많은 종을 포함했는데, 2006년 마르셀로 웩슬러(Marcelo Weksler)와 동료들의 기여로 절정에 달한 다양한 연구에서 40종 이상이 이 속에서 제거되면서 재분류되었다.[8] 이들은 모두 100종이 넘는 다양한 집단인 오리조미니족(Oryzomyini) ("쌀쥐")에 속하며,[9] 상위 분류군으로는 주로 작은 설치류 수백 종과 함께 시그모돈아과(Sigmodontinae)에 속하며 비단털쥐과(Cricetidae)에 속한다.[10]
==== 역사 ====
에드워드 알스톤은 1877년 멕시코와 과테말라에서 채집한 세 개의 표본을 사용하여 ''Oryzomys couesi''(쿠스쌀쥐)를 처음으로 기술했다.[11] 그는 이 동물의 이름을 ''Hesperomys couesi''로 명명하고, 현재는 폐지된 속인 ''Hesperomys''에 속하게 했으며, 습지쌀쥐(당시에는 ''Hesperomys palustris''로 불림)와 현재 ''Tylomys''에 속하는 두 종과의 유사점을 언급했다.[12] 종명인 ''couesi''는 북미 설치류에 대해 많은 연구를 한 미국 박물학자 Elliott Coues를 기리기 위해 붙여졌다.[11]
1893년 Oldfield Thomas는 당시 ''Oryzomys couesi''로 속이 Oryzomys에 속해 있던 이 종이 세 표본(과테말라 코반에서 1개, 멕시코에서 2개)이 알스톤이 실제로 두세 종에 속해 있었기 때문에 그 정체성에 대해 많은 혼란을 야기했다고 썼다. 그는 ''couesi''라는 이름을 과테말라의 동물로 제한하고, 멕시코 동물 중 하나에 새로운 이름 ''Oryzomys fulgens''를 부여했다.[13] 1890년대 초부터 다른 몇몇 관련 종들이 묘사되었고,[18] 1901년 클린턴 하트 메리엄은 이들 중 다수를 습지쌀쥐를 포함하는 ''palustris-mexicanus'' 종 그룹으로 통합했다.[14]
에드워드 알폰소 골드먼은 1918년에 북미 ''Oryzomys''를 수정하여 많은 형태를 단일 종 ''Oryzomys couesi''로 통합했으며, 이는 남부 텍사스와 멕시코 서부에서 코스타리카에 이르는 10개의 아종으로 분포했다. 그는 그것을 습지쌀쥐와 더 제한적인 분포를 가진 몇몇 종을 포함하는 ''Oryzomys palustris'' 그룹에 배치했는데, 그는 이를 ''O. couesi''와 관련이 있지만 별도의 종으로 분류될 만큼 충분히 독특하다고 여겼다.[15]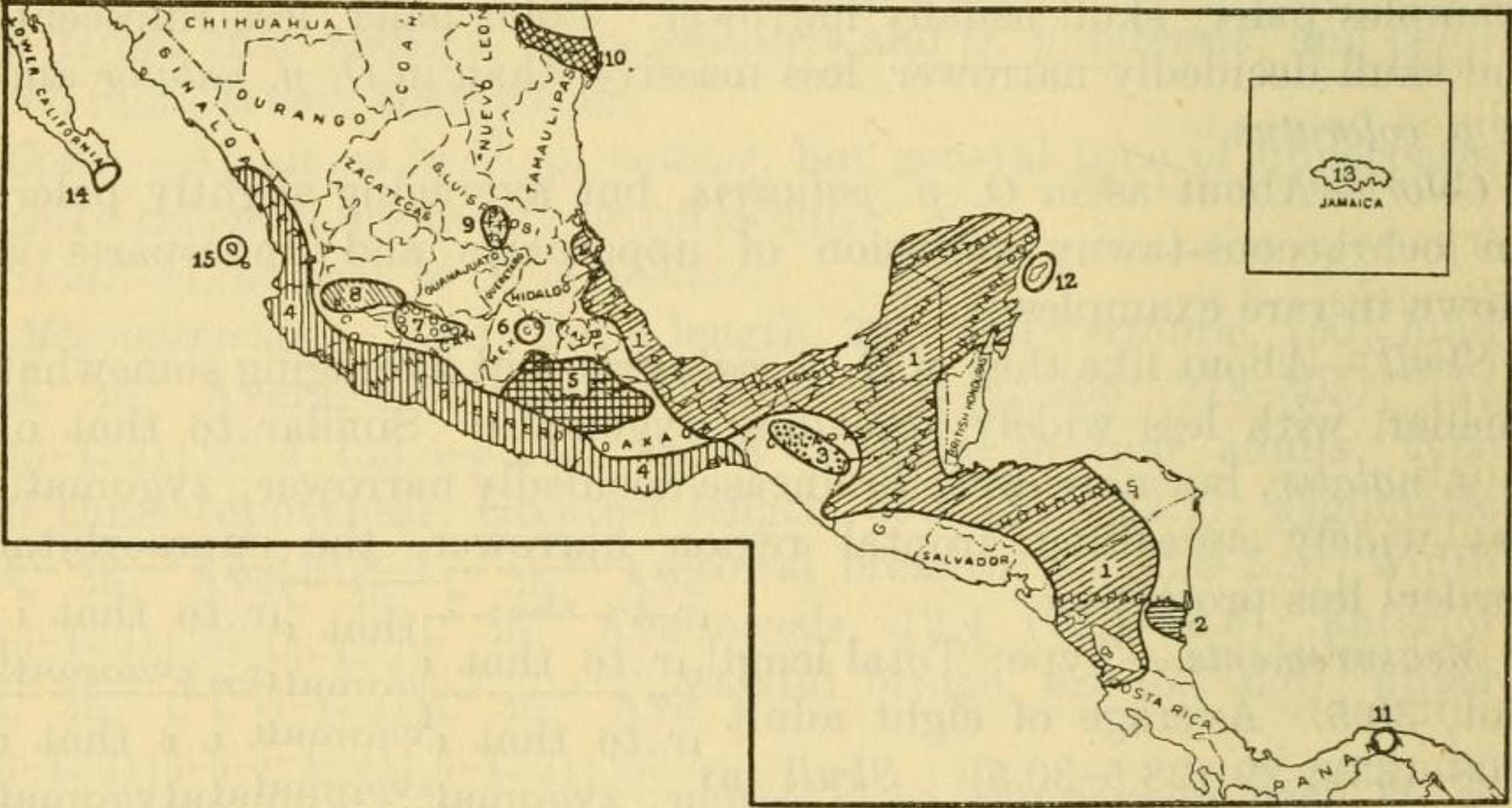
1930년대에 ''O. couesi''와 관련된 몇 가지 형태가 더 묘사되었다.[18]
1960년, 레이먼드 홀은 미국 종인 습지쌀쥐와 ''Oryzomys couesi''의 분포 범위가 텍사스 남부에서 만나는 지점에서 채집한 표본을 검토한 결과, 두 종을 분리할 근거를 찾지 못했다. 따라서 그는 ''O. couesi''를 습지쌀쥐의 아종으로 축소했다.[16] 다른 연구자들은 이러한 통합을 계속했고, 1971년까지 골드먼이 ''O. palustris'' 그룹에 배치한 다른 모든 종은 1937년에 종으로 묘사된 파나마의 ''Oryzomys azuerensis''와 함께 습지쌀쥐 아래에 분류되었다.[18]
1979년에 발표된 연구는 텍사스에서의 ''palustris–couesi'' 접촉 지점에 대한 추가 연구는 더 많은 표본과 특성을 사용하여 두 종이 실제로 거기에서 쉽게 구별될 수 있음을 나타냈다. 따라서 ''O. couesi''는 이후 습지쌀쥐와 구별되는 종으로 간주되었다.[18] 이후 ''O. couesi'' 또는 ''O. palustris'' 아래에 동의어로 묶인 다른 형태 중 일부는 서부 멕시코 마리아스 제도의 ''O. nelsoni'', 자메이카의 ''O. antillarum''과 같은 별도의 종으로 부활했다.[19]
2009년, 마이클 칼턴과 Joaquin Arroyo-Cabrales는 서부 멕시코 ''Oryzomys''를 검토하여 ''O. nelsoni''의 뚜렷함을 재확인하고, 바하칼리포르니아 반도의 끝 부분에서 ''O. peninsulae''와 멕시코 내륙의 ''O. albiventer''를 종으로 복원했다.[20] 그럼에도 불구하고, ''O. couesi''는 22개의 동의어를 포함했고,[21] 칼턴과 Arroyo-Cabrales는 ''O. couesi''와 관련 종에 대한 추가 연구가 확실히 추가적인 종의 인정을 초래할 것이라고 썼다.[22]
2010년 Delton Hanson과 동료들의 연구는 미토콘드리아 유전자 사이토크롬 ''b'' (''Cytb'')와 두 개의 핵 마커, 엑손 1의 간시세포 간 망막 결합 단백질 유전자 (''Rbp3'')와 인트론 2의 알코올 탈수소효소 유전자 1 (''Adh1-I2'')의 DNA 염기서열 데이터를 사용하여 습지쌀쥐와 ''O. couesi'' 개체군 간의 관계를 연구했다.[23] ''Cytb'' 데이터는 연구된 모든 ''O. couesi'' 표본을 습지쌀쥐와 자매 계통군에 배치했다. 두 그룹 간의 평균 유전적 거리는 11.30%였으며,[24] 이는 관련 속 ''Melanomys''와 ''Nectomys''의 자매 종 간의 거리(각각 7.48% 및 7.52%)보다 훨씬 컸다.[138] ''O. couesi'' 계통군 내에서, 파나마와 코스타리카에서 온 두 개체군은 다른 표본보다 연속적으로 기저 계통군이었는데, 이 표본은 두 개의 큰 아군(subclade)으로 나뉘었다. 하나는 멕시코 서부에서 엘살바도르에 이르는 태평양 해안에서 온 동물을 포함하고 다른 하나는 텍사스에서 니카라과에 이르는 동부 해안에서 온 쥐를 포함했다. 파나마와 코스타리카 개체군은 다른 것들과 6.53%에서 11.93% 차이를 보였고 서부와 동부 아군은 평균 4.41% 차이를 보였다.[24] 더 느리게 진화하는 핵 마커인 ''Rbp3''와 ''Adh1-I2''의 데이터도 ''Oryzomys''의 표본을 두 개의 주요 계통군에 배치했지만 ''O. couesi''의 서부 및 동부 그룹을 별도의 계통군으로 복구하지는 않았다. 또한, ''Adh1-I2''는 코스타리카 개체군을 습지쌀쥐 계통군 내에 배치하고 일부 서부 ''O. couesi'' 표본을 ''O. couesi'' 그룹보다 습지쌀쥐에 더 가깝게 배치했다.[25] 결합된 데이터 세트는 ''O. couesi'' 내의 서부 및 동부 계통군을 지원하고 코스타리카 개체군을 ''O. couesi''보다 습지쌀쥐에 약간 더 가깝게 배치했다.[26] 유전적 종 개념을 사용하여 저자들은 ''O. couesi'' 내에서 발견된 네 그룹을 별개의 종으로 인식해야 한다고 제안했다. 이 제안이 따르면, 동부 아군은 ''Oryzomys couesi''라는 이름을 유지하고, 서부 그룹은 ''Oryzomys mexicanus''로 명명되며, 파나마와 코스타리카 종에 적절한 이름은 아직 불분명하다.[27]
==== 하위 분류 ====
'''==== 멕시코 서부 ~ 엘살바도르 (''Oryzomys mexicanus'') ===='''
할리스코 (멕시코 서부)에서 엘살바도르 동부에 이르는 ''Oryzomys couesi'' 개체군은 단일 ''Cytb'' 계통을 형성하며, 한슨과 동료들은 이 계통을 종 ''Oryzomys mexicanus''로 인식할 것을 제안했다.[33] 이 동물들은 엄밀한 의미의 ''Oryzomys couesi''와 4.4% 차이를 보이며,[34] 북쪽과 동쪽에서 발견되고, 산맥으로 분리되어 있으며, 다른 종의 한타바이러스를 가지고 있으며, 메리엄(1901)에 따르면 더 튼튼한 두개골을 가지고 있으며 더 큰 어금니, 더 강한 관골궁, 그리고 두개골의 안와 사이 영역 (눈 사이)의 가장자리를 따라 더 잘 발달된 융기를 가지고 있다.[35] "''Oryzomys mexicanus''" 계통 내에서 ''Cytb'' 염기서열 차이는 평균 2.06%이며, 서부(할리스코 ~ 오아하카) 및 동부(치아파스 및 엘살바도르) 그룹은 별개의 하위 계통을 형성한다. 한슨과 동료들은 이들을 서부의 ''mexicanus''와 동부의 ''zygomaticus''로 다른 아종으로 인식했다.[36]
2009년 칼튼과 아로요-카브랄레스가 정의한 바에 따르면, 아종 ''Oryzomys couesi mexicanus''는 소노라 중부에서 오아하카 남동부에 이르는 태평양 연안을 따라, 그리고 내륙의 강을 따라 미초아칸 중부, 모렐로스 남부, 푸에블라 남부 및 오아하카 북서부까지 분포한다. 일반적으로 1000m 고도 이하에서 살지만, 할리스코에서는 1525m에서 발견되었다.[37] 이러한 분포 패턴은 ''Sigmodon mascotensis'', ''Hodomys alleni'', ''Peromyscus perfulvus'', 그리고 ''Osgoodomys banderanus''와 같은 다른 멕시코 서부 설치류와 유사하며, 일부 검토에서 별개의 생물지리 구역으로 인식되어 왔다.[41] ''O. c. mexicanus''는 다른 세 종의 ''Oryzomys''—''O. albiventer'', ''O. peninsulae'', 그리고 ''O. nelsoni''—과 가깝게 서식하는데, 이들은 더 크고 비율과 색상의 세부 사항이 다르다.[38]
조엘 아사프 앨런은 1897년 할리스코 표본을 바탕으로 ''Oryzomys mexicanus''를 완전한 종으로 처음 기술했다. 같은 출판물에서 그는 또한 인근 나야리트에서 ''Oryzomys bulleri''를 기술했지만, 이 두 종을 서로 비교하지는 않았다. 메리엄은 1901년에 나야리트에서 두 번째 종인 ''Oryzomys rufus''를 추가했는데, 이는 ''mexicanus''보다 작고 더 붉다는 점에 주목했다. 골드만은 1918년에 이 세 종을 ''O. couesi mexicanus''로 동의어로 처리했으며, 2009년 칼튼과 아로요-카브랄레스는 ''rufus''와 ''mexicanus''의 차이가 연령 관련성이 있으며 동물의 정상적인 변동 범위 내에 있다고 주장하며 이에 동의했다.[39] 또 다른 아종인 ''Oryzomys couesi lambi''는 1934년 버트에 의해 소노라 중부 해안에서 기술되었으며,[29] 당시 종의 범위를 400mi 확장했다. 이 형태는 짙은 회색-갈색으로 ''mexicanus''보다 훨씬 어둡고 꼬리가 짧으며 관골이 약하다.[40] 칼튼과 아로요-카브랄레스는 이것이 ''mexicanus''와 유사하지만, 아종으로 인정해야 하는지 여부를 결정하기 위해서는 추가 연구가 필요하다고 썼다. 시날로아 북부의 큰 ''O. couesi''도 이 형태에 속할 수 있다.[41] 골드만은 ''mexicanus''가 기준 ''couesi''와 매우 유사하지만, 일반적으로 털이 더 창백하다고 썼다. 상체는 ''couesi''보다 더 황갈색이며, 하체는 일반적으로 흰색이지만 황갈색일 수 있으며, 이는 ''couesi''의 일반적인 색상이다.[42]
''Oryzomys zygomaticus''는 메리엄이 1901년에 별도의 종으로 처음 기술했으며,[32] ''mexicanus''와 유사하지만 관골궁이 넓게 퍼져 아래로 굽어 있다.[73] 이를 ''couesi''의 아종으로 축소한 골드만은 과테말라 남서부와 인근 치아파스에서 이를 기록했으며, ''O. c. couesi''보다 약간 창백하지만 ''O. c. mexicanus''보다 어둡다고 기술했다.[43] 엘살바도르 중부의 세 표본은 ''zygomaticus''와 유사한 ''Cytb'' 염기서열을 가지고 있지만,[44] 『엘살바도르의 포유류』(1961)에서 버트와 스티어튼은 이 나라에서 아종 ''couesi''만을 기록했으며, 일부 지역의 표본이 다른 표본보다 약간 창백하다는 점에 주목했다.[45]
'''==== 멕시코 내륙 ===='''
골드만은 멕시코 중부 내륙 고원의 4종의 쿠스쌀쥐 아종, 즉 'albiventer', 'crinitus', 'aztecus', 'regillus'를 함께 묶었다.[49] 이 중 3종('albiventer'는 할리스코,[50] 'crinitus'는 연방구,[47] 'aztecus'는 모렐로스)[46]는 메리엄이 1901년에 기술했고, 골드만은 1915년에 미초아칸에서 'regillus'를 직접 기술했다.[48] 골드만에 따르면, 'aztecus'는 창백하고 이빨이 크고,[51] 'crinitus'는 크고 어둡고 이빨이 크며,[52] 'regillus'는 크고 어둡고,[49] 'albiventer'는 크고 비교적 창백하다.[53]
2009년 멕시코 서부의 ''Oryzomys''에 대한 검토에서, 칼턴과 아로요-카브랄레스는 명확한 형태 계측적 차이를 근거로 평지 ''mexicanus''와는 별개의 종으로 ''Oryzomys albiventer''를 분류했다.[54] 또한 'crinitus', 'regillus', 'aztecus'의 지위에 대한 몇 가지 의견을 제시했으며, 3가지 형태의 모식표본을 형태 계측 분석에 포함했다. 'regillus'와 'aztecus'의 모식표본은 멕시코 서부 저지대에서 발견된 대규모 ''mexicanus'' 표본의 변이 범위 상한에 속했고, 'crinitus'는 바하칼리포르니아 반도 끝자락에서 발견된 ''O. peninsulae'' 표본과 함께 묶였다. 그들은 'regillus'와 'aztecus'가 ''mexicanus''의 튼튼한 고지대 개체군에 지나지 않을 수 있지만, 다른 종을 나타낼 가능성을 배제할 수 없다고 제안했다. 멕시코 계곡의 2000 m 이상 고도에서 발견되는 'crinitus'가 바하칼리포르니아 반도 저지대의 'peninsulae'와 같은 종이라는 점은 받아들일 수 없었고, 'crinitus'의 관계를 결정하기 위한 추가 연구를 권장했다.[55] 미초아칸 내륙의 한 표본은 ''Cytb'' 데이터가 ''mexicanus''의 특징을 나타냈지만,[56] 한슨과 동료들은 다른 멕시코 내륙 ''Oryzomys''에 대한 데이터를 가지고 있지 않았다.[57]
토마스가 1893년에 기술한 종인 ''Oryzomys fulgens''의 모식표본은 "멕시코"라는 더 정확한 지역이 없지만, 멕시코 계곡이 그 기원으로 제안되었다.[58] 그것은 크고, 거친 털을 가진, 밝은 적갈색,[13] 넓게 퍼진 광대뼈 아치를 가진 넓은 두개골을 가진 긴 꼬리 종이다.[59] 골드만은 이것이 'crinitus'와 유사하지만 색상이 더 강렬하고 안와 간 부위의 형태가 다르다고 기록했으며, 더 많은 조사가 있을 때까지 별도의 종으로 유지했다.[60] 칼턴과 아로요-카브랄레스는 보관 연구를 통해 다른 멕시코 중부 ''Oryzomys''의 이전 이름으로 이를 확립할 수 있는 ''O. fulgens''의 정확한 기원이 밝혀질 수 있다고 언급했다.[61]
'''==== 텍사스 ~ 니카라과 (''Oryzomys couesi'') ===='''
텍사스에서 니카라과까지의 ''Oryzomys'' 개체군은 평균 염기서열 차이가 1.28%인 단일 ''Cytb'' 분류군을 형성하며,[74] Hanson과 동료들은 이 분류군에 ''Oryzomys couesi''라는 이름을 제한할 것을 제안했다.[27] 이러한 개체군은 Goldman이 인정한 두 아종(''O. c. aquaticus'' 및 ''O. c. couesi'')과 종으로 유지한 섬 형태(''O. cozumelae'')에 해당한다. Goldman이 인정한 다른 두 아종, ''O. c. richmondi''와 ''O. c. peragrus'' 및 Goldman의 논문 이후에 설명된 세 번째 아종, ''O. c. pinicola''는 동일한 지역에서 발생하지만 유전적으로 연구되지 않았다.[75]
''Oryzomys couesi''의 최북단 개체군, 즉 텍사스 남부와 인근 멕시코 타마울리파스주에 있는 개체군은 1891년에 별도의 종, ''Oryzomys aquaticus''로 기술된 아종 ''aquaticus''로 분류된다.[77] 여기에서 ''O. couesi''의 분포 범위는 습지쌀쥐의 분포 범위와 만난다.[78] 텍사스주 케네디 카운티, 윌라시 카운티 및 카메론 카운티의 일부와 타마울리파스주 북동부에서 이 두 종은 공존한다(같은 장소에서 발생). 접촉 구역에서 ''couesi''는 내륙에서 더 멀리 발생하고, 습지 쌀쥐는 해안을 따라 산다.[79] 실험 조건에서 두 종은 교배에 실패하고[80] 유전자 분석 결과 야생에서 유전자 흐름이나 잡종화의 증거가 없다.[81] 남쪽의 개체군과 비교하여 ''aquaticus''는 더 크고 창백하며 더 강건한 두개골을 가지고 있다.[82] 타마울리파스주에서 온 표본은 텍사스에서 온 표본보다 약간 더 어둡다.[83] ''aquaticus'' 표본의 ''Cytb'' 염기서열은 별도의 그룹을 형성하지만, 더 남쪽에서 온 ''O. c. couesi'' 표본 내에 무리를 이룬다.[84]
''peragrus'' 형태는 멕시코 남부의 산루이스포토시주 리오 베르데 분지, 이달고주, 그리고 베라크루스주 북부에서 알려져 있다.[85] 이 형태의 플라이스토세 후기 화석이 타마울리파스주 쿠에바 데 아브레에서 발견되었다.[86] Goldman에 따르면, 그것은 ''O. c. aquaticus''와 ''O. c. couesi'' 사이의 중간 색상을 가지지만 ''aquaticus''와 유사한 두개골을 가지고 있다.[87]
Goldman은 베라크루스 북부에서 멕시코 동부, 과테말라, 온두라스, 니카라과를 거쳐 코스타리카 북서부까지 이르는 개체군을 명명 아종인 ''Oryzomys couesi couesi''로 통합했다. 그는 베라크루스에서 온 Allen과 Chapman의 1897년 종인 ''Oryzomys jalapae'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys jalapae rufinus'', 타바스코주에서 온 Merriam의 1901년 종인 ''Oryzomys teapensis'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys goldmani'', 베라크루스에서 온 Eliot의 1904년 종인 ''Oryzomys jalapae apatelius'' 및 니카라과에서 온 Allen의 1910년 종인 ''Oryzomys richardsoni''를 이 형태의 완전한 동의어로 배치했다.[88] Goldman에 따르면, 아종 내의 개인적 변이는 광범위하여 많은 수의 출판된 동의어를 낳았지만, 분포 범위의 모든 부분에서 온 개체군은 본질적으로 유사하다.[89]
아종 ''Oryzomys couesi pinicola''는 1932년에 서부 영국령 온두라스(현재 벨리즈)의 소나무 능선에서 기술되었다. 이 종은 명명 아종인 ''couesi''보다 작고 어두우며, 벨리즈에서도 발생하며, 더 섬세한 두개골을 가지고 있다.[90] 1901년에 Merriam은 코수멜 섬의 ''Oryzomys''를 별도의 종인 ''Oryzomys cozumelae''로 기술했으며, Goldman은 큰 크기, 어두운 털, 긴 꼬리 때문에 이를 유지했다.[91] 그러나 1965년에 Knox Jones와 Timothy Lawlor는 ''cozumelae''와 본토 ''couesi'' 사이의 차이가 사소하다고 판단하고 ''cozumelae''가 본토 ''Oryzomys'' 개체군의 변동 범위 내에 있음을 발견했다. 따라서 그들은 이 섬 형태를 아종으로 강등시켰다.[92] 1989년에 Mark Engstrom과 동료들은 이 결론을 재확인했다.[93] 섬 형태의 경우, 이 개체군은 유전적으로 매우 다양하다.[94] ''Cytb'' 염기서열 데이터에서 명명 아종인 ''couesi''의 개체군에 속한다.[57] ''Oryzomys couesi''는 벨리즈 해안의 턴네페 환초와 온두라스 해안의 로아탄에서도 발견된다.[95]
니카라과 동부 저지대의 ''Oryzomys''는 1901년에 Merriam에 의해 별도의 종인 ''Oryzomys richmondi''로 기술되었으며, Goldman은 뚜렷하게 어두운 털을 근거로 ''O. couesi''의 아종으로 유지했다.[97] 1986년에 니카라과 ''Oryzomys''를 검토하면서 Jones와 Engstrom은 색상 차이가 아종을 인식하기에는 너무 작다고 생각하여 ''richmondi''를 별도로 유지하지 않았다.[98] 회색 아래 부분을 가진 작고 어두운 ''Oryzomys''인 ''Oryzomys dimidiatus''는 니카라과 남동부에서 ''O. couesi''와 함께 발생한다.[99] Jones와 Engstrom에 따르면, 니카라과 호의 오메테페 섬에서 온 쌀쥐는 큰 두개골과 작은 외부 측정값, 특히 짧은 꼬리, 위는 오렌지색-갈색이고 아래는 황갈색인 부드러운 털, 접형골 와 (sphenopalatine vacuities)(뼈 구개 끝 뒤의 간극인 중간 익상와의 열린 부분)의 부재로 특징적이다. 그들은 이 개체군이 별도의 아종을 나타낼 가능성이 높다고 생각했지만, 성체 표본이 하나뿐이었기 때문에 새로운 이름을 제안하는 것을 거부했다.[100] 니카라과에서 ''O. couesi''는 최대 1250 m의 고도에서 발생한다.[101]
'''==== 코스타리카, 파나마, 콜롬비아 ===='''
코스타리카의 ''Oryzomys''는 역사적으로 ''O. c. couesi''로 언급되어 왔지만,[104] 한슨과 동료들은 코스타리카 북동부의 레푸지오 나시오날 데 비다 실베스트레 믹스토 마퀜케에서 채집된 두 표본이 다른 ''O. couesi''와 11.93% ''Cytb'' 염기 서열 차이를 보이며, 이는 늪쌀쥐와 ''O. couesi''의 차이(11.30%)만큼 큰 차이라는 것을 발견했다. 그들은 이 동물들이 ''O. couesi''와는 다른 종을 나타낸다고 제안했지만, ''dimidiatus'' 또는 ''richmondi''의 표본을 검사할 수 없었기 때문에 이 종에 적합한 이름을 결정할 수 없었다.[105]
''Oryzomys''는 파나마에서 드물다.[106] 파나마의 ''Oryzomys''는 1912년 골드먼에 의해 처음으로 기술되었으며, 그는 운하 지대의 가툰에서 채집된 표본에 대해 ''Oryzomys gatunensis''라는 이름을 도입했다.[103] 1918년 골드먼은 이 동물을 별개의 종으로 유지하면서, 그 동물이 ''richmondi''와 유사하지만, 안와 부위 가장자리를 따라 잘 발달된 능선, 짧은 두정골 및 긴 비골에서 뚜렷하다고 언급했다.[107] 1937년 볼은 파나마의 또 다른 ''Oryzomys'' 종인 ''Oryzomys azuerensis''를 베라구아스 주의 파라코테에서 기술했다.[102] 이 종은 갈색 형태이며, 주변 개체군의 붉은 색조가 없고, 짧은 로스트럼(앞 부분)과 ''gatunensis''의 것과 같은 안와 부위의 능선이 있는 넓은 두개골을 가지고 있다.[108] 골드먼은 그에게 ''gatunensis''와 ''azuerensis''를 모두 ''couesi''의 아종으로 취급할 것을 권장했지만, 볼은 ''azuerensis''가 지리적으로 가장 가까운 형태인 ''gatunensis''와 ''couesi'' 사이
2. 1. 역사
에드워드 알스톤은 1877년 멕시코와 과테말라에서 채집한 세 개의 표본을 사용하여 ''Oryzomys couesi''(쿠스쌀쥐)를 처음으로 기술했다.[11] 그는 이 동물의 이름을 ''Hesperomys couesi''로 명명하고, 현재는 폐지된 속인 ''Hesperomys''에 속하게 했으며, 습지쌀쥐(당시에는 ''Hesperomys palustris''로 불림)와 현재 ''Tylomys''에 속하는 두 종과의 유사점을 언급했다.[12] 종명인 ''couesi''는 북미 설치류에 대해 많은 연구를 한 미국 박물학자 Elliott Coues를 기리기 위해 붙여졌다.[11]1893년 Oldfield Thomas는 당시 ''Oryzomys couesi''로 속이 Oryzomys에 속해 있던 이 종이 세 표본(과테말라 Cobán에서 1개, 멕시코에서 2개)이 알스톤이 실제로 두세 종에 속해 있었기 때문에 그 정체성에 대해 많은 혼란을 야기했다고 썼다. 그는 ''couesi''라는 이름을 과테말라의 동물로 제한하고, 멕시코 동물 중 하나에 새로운 이름 ''Oryzomys fulgens''를 부여했다.[13] 1890년대 초부터 다른 몇몇 관련 종들이 묘사되었고,[18] 1901년 Clinton Hart Merriam은 이들 중 다수를 습지쌀쥐를 포함하는 ''palustris-mexicanus'' 종 그룹으로 통합했다.[14]
Edward Alphonso Goldman은 1918년에 북미 ''Oryzomys''를 수정하여 많은 형태를 단일 종 ''Oryzomys couesi''로 통합했으며, 이는 남부 텍사스와 멕시코 서부에서 코스타리카에 이르는 10개의 아종으로 분포했다. 그는 그것을 습지쌀쥐와 더 제한적인 분포를 가진 몇몇 종을 포함하는 ''Oryzomys palustris'' 그룹에 배치했는데, 그는 이를 ''O. couesi''와 관련이 있지만 별도의 종으로 분류될 만큼 충분히 독특하다고 여겼다.[15]
1930년대에 ''O. couesi''와 관련된 몇 가지 형태가 더 묘사되었다.[18]
1960년, Raymond Hall은 미국 종인 습지쌀쥐와 ''Oryzomys couesi''의 분포 범위가 텍사스 남부에서 만나는 지점에서 채집한 표본을 검토한 결과, 두 종을 분리할 근거를 찾지 못했다. 따라서 그는 ''O. couesi''를 습지쌀쥐의 아종으로 축소했다.[16] 다른 연구자들은 이러한 통합을 계속했고, 1971년까지 골드먼이 ''O. palustris'' 그룹에 배치한 다른 모든 종은 1937년에 종으로 묘사된 파나마의 ''Oryzomys azuerensis''와 함께 습지쌀쥐 아래에 분류되었다.[18]
1979년에 발표된 연구는 텍사스에서의 ''palustris–couesi'' 접촉 지점에 대한 추가 연구는 더 많은 표본과 특성을 사용하여 두 종이 실제로 거기에서 쉽게 구별될 수 있음을 나타냈다. 따라서 ''O. couesi''는 이후 습지쌀쥐와 구별되는 종으로 간주되었다.[18] 이후 ''O. couesi'' 또는 ''O. palustris'' 아래에 동의어로 묶인 다른 형태 중 일부는 서부 멕시코 마리아스 제도의 ''Oryzomys nelsoni'', 자메이카의 ''Oryzomys antillarum''과 같은 별도의 종으로 부활했다.[19]
2009년, Michael Carleton과 Joaquin Arroyo-Cabrales는 서부 멕시코 ''Oryzomys''를 검토하여 ''O. nelsoni''의 뚜렷함을 재확인하고, 바하칼리포르니아 반도의 끝 부분에서 ''O. peninsulae''와 멕시코 내륙의 ''O. albiventer''를 종으로 복원했다.[20] 그럼에도 불구하고, ''O. couesi''는 22개의 동의어를 포함했고,[21] Carleton과 Arroyo-Cabrales는 ''O. couesi''와 관련 종에 대한 추가 연구가 확실히 추가적인 종의 인정을 초래할 것이라고 썼다.[22]
2010년 Delton Hanson과 동료들의 연구는 미토콘드리아 유전자 사이토크롬 ''b'' (''Cytb'')와 두 개의 핵 마커, 엑손 1의 간시세포 간 망막 결합 단백질 유전자 (''Rbp3'')와 인트론 2의 알코올 탈수소효소 유전자 1 (''Adh1-I2'')의 DNA 염기서열 데이터를 사용하여 습지쌀쥐와 ''O. couesi'' 개체군 간의 관계를 연구했다.[23] ''Cytb'' 데이터는 연구된 모든 ''O. couesi'' 표본을 습지쌀쥐와 자매 clade에 배치했다. 두 그룹 간의 평균 유전적 거리는 11.30%였으며,[24] 이는 관련 속 ''Melanomys''와 ''Nectomys''의 자매 종 간의 거리(각각 7.48% 및 7.52%)보다 훨씬 컸다.[138] ''O. couesi'' clade 내에서, 파나마와 코스타리카에서 온 두 개체군은 다른 표본보다 연속적으로 기저 clade였는데, 이 표본은 두 개의 큰 subclade로 나뉘었다. 하나는 멕시코 서부에서 엘살바도르에 이르는 태평양 해안에서 온 동물을 포함하고 다른 하나는 텍사스에서 니카라과에 이르는 동부 해안에서 온 쥐를 포함했다. 파나마와 코스타리카 개체군은 다른 것들과 6.53%에서 11.93% 차이를 보였고 서부와 동부 subclade는 평균 4.41% 차이를 보였다.[24] 더 느리게 진화하는 핵 마커인 ''Rbp3''와 ''Adh1-I2''의 데이터도 ''Oryzomys''의 표본을 두 개의 주요 clade에 배치했지만 ''O. couesi''의 서부 및 동부 그룹을 별도의 clade로 복구하지는 않았다. 또한, ''Adh1-I2''는 코스타리카 개체군을 습지쌀쥐 clade 내에 배치하고 일부 서부 ''O. couesi'' 표본을 ''O. couesi'' 그룹보다 습지쌀쥐에 더 가깝게 배치했다.[25] 결합된 데이터 세트는 ''O. couesi'' 내의 서부 및 동부 clade를 지원하고 코스타리카 개체군을 ''O. couesi''보다 습지쌀쥐에 약간 더 가깝게 배치했다.[26] 유전적 종 개념을 사용하여 저자들은 ''O. couesi'' 내에서 발견된 네 그룹을 별개의 종으로 인식해야 한다고 제안했다. 이 제안이 따르면, 동부 subclade는 ''Oryzomys couesi''라는 이름을 유지하고, 서부 그룹은 ''Oryzomys mexicanus''로 명명되며, 파나마와 코스타리카 종에 적절한 이름은 아직 불분명하다.[27]
2. 2. 하위 분류
'''==== 멕시코 서부 ~ 엘살바도르 (''Oryzomys mexicanus'') ===='''할리스코 (멕시코 서부)에서 엘살바도르 동부에 이르는 ''Oryzomys couesi'' 개체군은 단일 ''Cytb'' 계통을 형성하며, 한슨과 동료들은 이 계통을 종 ''Oryzomys mexicanus''로 인식할 것을 제안했다.[33] 이 동물들은 엄밀한 의미의 ''Oryzomys couesi''와 4.4% 차이를 보이며,[34] 북쪽과 동쪽에서 발견되고, 산맥으로 분리되어 있으며, 다른 종의 한타바이러스를 가지고 있으며, 메리엄(1901)에 따르면 더 튼튼한 두개골을 가지고 있으며 더 큰 어금니, 더 강한 관골궁, 그리고 두개골의 안와 사이 영역 (눈 사이)의 가장자리를 따라 더 잘 발달된 융기를 가지고 있다.[35] "''Oryzomys mexicanus''" 계통 내에서 ''Cytb'' 염기서열 차이는 평균 2.06%이며, 서부(할리스코 ~ 오아하카) 및 동부(치아파스 및 엘살바도르) 그룹은 별개의 하위 계통을 형성한다. 한슨과 동료들은 이들을 서부의 ''mexicanus''와 동부의 ''zygomaticus''로 다른 아종으로 인식했다.[36]
2009년 칼튼과 아로요-카브랄레스가 정의한 바에 따르면, 아종 ''Oryzomys couesi mexicanus''는 소노라 중부에서 오아하카 남동부에 이르는 태평양 연안을 따라, 그리고 내륙의 강을 따라 미초아칸 중부, 모렐로스 남부, 푸에블라 남부 및 오아하카 북서부까지 분포한다. 일반적으로 1000m 고도 이하에서 살지만, 할리스코에서는 1525m에서 발견되었다.[37] 이러한 분포 패턴은 ''Sigmodon mascotensis'', ''Hodomys alleni'', ''Peromyscus perfulvus'', 그리고 ''Osgoodomys banderanus''와 같은 다른 멕시코 서부 설치류와 유사하며, 일부 검토에서 별개의 생물지리 구역으로 인식되어 왔다.[41] ''O. c. mexicanus''는 다른 세 종의 ''Oryzomys''—''O. albiventer'', ''O. peninsulae'', 그리고 ''O. nelsoni''—과 가깝게 서식하는데, 이들은 더 크고 비율과 색상의 세부 사항이 다르다.[38]
조엘 아사프 앨런은 1897년 할리스코 표본을 바탕으로 ''Oryzomys mexicanus''를 완전한 종으로 처음 기술했다. 같은 출판물에서 그는 또한 인근 나야리트에서 ''Oryzomys bulleri''를 기술했지만, 이 두 종을 서로 비교하지는 않았다. 메리엄은 1901년에 나야리트에서 두 번째 종인 ''Oryzomys rufus''를 추가했는데, 이는 ''mexicanus''보다 작고 더 붉다는 점에 주목했다. 골드만은 1918년에 이 세 종을 ''O. couesi mexicanus''로 동의어로 처리했으며, 2009년 칼튼과 아로요-카브랄레스는 ''rufus''와 ''mexicanus''의 차이가 연령 관련성이 있으며 동물의 정상적인 변동 범위 내에 있다고 주장하며 이에 동의했다.[39] 또 다른 아종인 ''Oryzomys couesi lambi''는 1934년 버트에 의해 소노라 중부 해안에서 기술되었으며,[29] 당시 종의 범위를 400mi 확장했다. 이 형태는 짙은 회색-갈색으로 ''mexicanus''보다 훨씬 어둡고 꼬리가 짧으며 관골이 약하다.[40] 칼튼과 아로요-카브랄레스는 이것이 ''mexicanus''와 유사하지만, 아종으로 인정해야 하는지 여부를 결정하기 위해서는 추가 연구가 필요하다고 썼다. 시날로아 북부의 큰 ''O. couesi''도 이 형태에 속할 수 있다.[41] 골드만은 ''mexicanus''가 기준 ''couesi''와 매우 유사하지만, 일반적으로 털이 더 창백하다고 썼다. 상체는 ''couesi''보다 더 황갈색이며, 하체는 일반적으로 흰색이지만 황갈색일 수 있으며, 이는 ''couesi''의 일반적인 색상이다.[42]
''Oryzomys zygomaticus''는 메리엄이 1901년에 별도의 종으로 처음 기술했으며,[32] ''mexicanus''와 유사하지만 관골궁이 넓게 퍼져 아래로 굽어 있다.[73] 이를 ''couesi''의 아종으로 축소한 골드만은 과테말라 남서부와 인근 치아파스에서 이를 기록했으며, ''O. c. couesi''보다 약간 창백하지만 ''O. c. mexicanus''보다 어둡다고 기술했다.[43] 엘살바도르 중부의 세 표본은 ''zygomaticus''와 유사한 ''Cytb'' 염기서열을 가지고 있지만,[44] 『엘살바도르의 포유류』(1961)에서 버트와 스티어튼은 이 나라에서 아종 ''couesi''만을 기록했으며, 일부 지역의 표본이 다른 표본보다 약간 창백하다는 점에 주목했다.[45]
'''==== 멕시코 내륙 ===='''
골드만은 멕시코 중부 내륙 고원의 4종의 쿠스쌀쥐 아종, 즉 'albiventer', 'crinitus', 'aztecus', 'regillus'를 함께 묶었다.[49] 이 중 3종('albiventer'는 할리스코,[50] 'crinitus'는 연방구,[47] 'aztecus'는 모렐로스)[46]는 메리엄이 1901년에 기술했고, 골드만은 1915년에 미초아칸에서 'regillus'를 직접 기술했다.[48] 골드만에 따르면, 'aztecus'는 창백하고 이빨이 크고,[51] 'crinitus'는 크고 어둡고 이빨이 크며,[52] 'regillus'는 크고 어둡고,[49] 'albiventer'는 크고 비교적 창백하다.[53]
2009년 멕시코 서부의 ''Oryzomys''에 대한 검토에서, 칼턴과 아로요-카브랄레스는 명확한 형태 계측적 차이를 근거로 평지 ''mexicanus''와는 별개의 종으로 ''Oryzomys albiventer''를 분류했다.[54] 또한 'crinitus', 'regillus', 'aztecus'의 지위에 대한 몇 가지 의견을 제시했으며, 3가지 형태의 모식표본을 형태 계측 분석에 포함했다. 'regillus'와 'aztecus'의 모식표본은 멕시코 서부 저지대에서 발견된 대규모 ''mexicanus'' 표본의 변이 범위 상한에 속했고, 'crinitus'는 바하칼리포르니아 반도 끝자락에서 발견된 ''O. peninsulae'' 표본과 함께 묶였다. 그들은 'regillus'와 'aztecus'가 ''mexicanus''의 튼튼한 고지대 개체군에 지나지 않을 수 있지만, 다른 종을 나타낼 가능성을 배제할 수 없다고 제안했다. 멕시코 계곡의 2000 m 이상 고도에서 발견되는 'crinitus'가 바하칼리포르니아 반도 저지대의 'peninsulae'와 같은 종이라는 점은 받아들일 수 없었고, 'crinitus'의 관계를 결정하기 위한 추가 연구를 권장했다.[55] 미초아칸 내륙의 한 표본은 ''Cytb'' 데이터가 ''mexicanus''의 특징을 나타냈지만,[56] 한슨과 동료들은 다른 멕시코 내륙 ''Oryzomys''에 대한 데이터를 가지고 있지 않았다.[57]
토마스가 1893년에 기술한 종인 ''Oryzomys fulgens''의 모식표본은 "멕시코"라는 더 정확한 지역이 없지만, 멕시코 계곡이 그 기원으로 제안되었다.[58] 그것은 크고, 거친 털을 가진, 밝은 적갈색,[13] 넓게 퍼진 광대뼈 아치를 가진 넓은 두개골을 가진 긴 꼬리 종이다.[59] 골드만은 이것이 'crinitus'와 유사하지만 색상이 더 강렬하고 안와 간 부위의 형태가 다르다고 기록했으며, 더 많은 조사가 있을 때까지 별도의 종으로 유지했다.[60] 칼턴과 아로요-카브랄레스는 보관 연구를 통해 다른 멕시코 중부 ''Oryzomys''의 이전 이름으로 이를 확립할 수 있는 ''O. fulgens''의 정확한 기원이 밝혀질 수 있다고 언급했다.[61]
'''==== 텍사스 ~ 니카라과 (''Oryzomys couesi'') ===='''
텍사스에서 니카라과까지의 ''Oryzomys'' 개체군은 평균 염기서열 차이가 1.28%인 단일 ''Cytb'' 분류군을 형성하며,[74] Hanson과 동료들은 이 분류군에 ''Oryzomys couesi''라는 이름을 제한할 것을 제안했다.[27] 이러한 개체군은 Goldman이 인정한 두 아종(''O. c. aquaticus'' 및 ''O. c. couesi'')과 종으로 유지한 섬 형태(''O. cozumelae'')에 해당한다. Goldman이 인정한 다른 두 아종, ''O. c. richmondi''와 ''O. c. peragrus'' 및 Goldman의 논문 이후에 설명된 세 번째 아종, ''O. c. pinicola''는 동일한 지역에서 발생하지만 유전적으로 연구되지 않았다.[75]
''Oryzomys couesi''의 최북단 개체군, 즉 텍사스 남부와 인근 멕시코 타마울리파스주에 있는 개체군은 1891년에 별도의 종, ''Oryzomys aquaticus''로 기술된 아종 ''aquaticus''로 분류된다.[77] 여기에서 ''O. couesi''의 분포 범위는 습지쌀쥐의 분포 범위와 만난다.[78] 텍사스주 케네디 카운티, 윌라시 카운티 및 카메론 카운티의 일부와 타마울리파스주 북동부에서 이 두 종은 공존한다(같은 장소에서 발생). 접촉 구역에서 ''couesi''는 내륙에서 더 멀리 발생하고, 습지 쌀쥐는 해안을 따라 산다.[79] 실험 조건에서 두 종은 교배에 실패하고[80] 유전자 분석 결과 야생에서 유전자 흐름이나 잡종화의 증거가 없다.[81] 남쪽의 개체군과 비교하여 ''aquaticus''는 더 크고 창백하며 더 강건한 두개골을 가지고 있다.[82] 타마울리파스주에서 온 표본은 텍사스에서 온 표본보다 약간 더 어둡다.[83] ''aquaticus'' 표본의 ''Cytb'' 염기서열은 별도의 그룹을 형성하지만, 더 남쪽에서 온 ''O. c. couesi'' 표본 내에 무리를 이룬다.[84]
''peragrus'' 형태는 멕시코 남부의 산루이스포토시주 리오 베르데 분지, 이달고주, 그리고 베라크루스주 북부에서 알려져 있다.[85] 이 형태의 플라이스토세 후기 화석이 타마울리파스주 쿠에바 데 아브레에서 발견되었다.[86] Goldman에 따르면, 그것은 ''O. c. aquaticus''와 ''O. c. couesi'' 사이의 중간 색상을 가지지만 ''aquaticus''와 유사한 두개골을 가지고 있다.[87]
Goldman은 베라크루스 북부에서 멕시코 동부, 과테말라, 온두라스, 니카라과를 거쳐 코스타리카 북서부까지 이르는 개체군을 명명 아종인 ''Oryzomys couesi couesi''로 통합했다. 그는 베라크루스에서 온 Allen과 Chapman의 1897년 종인 ''Oryzomys jalapae'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys jalapae rufinus'', 타바스코주에서 온 Merriam의 1901년 종인 ''Oryzomys teapensis'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys goldmani'', 베라크루스에서 온 Eliot의 1904년 종인 ''Oryzomys jalapae apatelius'' 및 니카라과에서 온 Allen의 1910년 종인 ''Oryzomys richardsoni''를 이 형태의 완전한 동의어로 배치했다.[88] Goldman에 따르면, 아종 내의 개인적 변이는 광범위하여 많은 수의 출판된 동의어를 낳았지만, 분포 범위의 모든 부분에서 온 개체군은 본질적으로 유사하다.[89]
아종 ''Oryzomys couesi pinicola''는 1932년에 서부 영국령 온두라스(현재 벨리즈)의 소나무 능선에서 기술되었다. 이 종은 명명 아종인 ''couesi''보다 작고 어두우며, 벨리즈에서도 발생하며, 더 섬세한 두개골을 가지고 있다.[90] 1901년에 Merriam은 코수멜 섬의 ''Oryzomys''를 별도의 종인 ''Oryzomys cozumelae''로 기술했으며, Goldman은 큰 크기, 어두운 털, 긴 꼬리 때문에 이를 유지했다.[91] 그러나 1965년에 Knox Jones와 Timothy Lawlor는 ''cozumelae''와 본토 ''couesi'' 사이의 차이가 사소하다고 판단하고 ''cozumelae''가 본토 ''Oryzomys'' 개체군의 변동 범위 내에 있음을 발견했다. 따라서 그들은 이 섬 형태를 아종으로 강등시켰다.[92] 1989년에 Mark Engstrom과 동료들은 이 결론을 재확인했다.[93] 섬 형태의 경우, 이 개체군은 유전적으로 매우 다양하다.[94] ''Cytb'' 염기서열 데이터에서 명명 아종인 ''couesi''의 개체군에 속한다.[57] ''Oryzomys couesi''는 벨리즈 해안의 턴네페 환초와 온두라스 해안의 로아탄에서도 발견된다.[95]
니카라과 동부 저지대의 ''Oryzomys''는 1901년에 Merriam에 의해 별도의 종인 ''Oryzomys richmondi''로 기술되었으며, Goldman은 뚜렷하게 어두운 털을 근거로 ''O. couesi''의 아종으로 유지했다.[97] 1986년에 니카라과 ''Oryzomys''를 검토하면서 Jones와 Engstrom은 색상 차이가 아종을 인식하기에는 너무 작다고 생각하여 ''richmondi''를 별도로 유지하지 않았다.[98] 회색 아래 부분을 가진 작고 어두운 ''Oryzomys''인 ''Oryzomys dimidiatus''는 니카라과 남동부에서 ''O. couesi''와 함께 발생한다.[99] Jones와 Engstrom에 따르면, 니카라과 호의 오메테페 섬에서 온 쌀쥐는 큰 두개골과 작은 외부 측정값, 특히 짧은 꼬리, 위는 오렌지색-갈색이고 아래는 황갈색인 부드러운 털, 접형골 와 (sphenopalatine vacuities)(뼈 구개 끝 뒤의 간극인 중간 익상와의 열린 부분)의 부재로 특징적이다. 그들은 이 개체군이 별도의 아종을 나타낼 가능성이 높다고 생각했지만, 성체 표본이 하나뿐이었기 때문에 새로운 이름을 제안하는 것을 거부했다.[100] 니카라과에서 ''O. couesi''는 최대 1250 m의 고도에서 발생한다.[101]
'''==== 코스타리카, 파나마, 콜롬비아 ===='''
코스타리카의 ''Oryzomys''는 역사적으로 ''O. c. couesi''로 언급되어 왔지만,[104] 한슨과 동료들은 코스타리카 북동부의 레푸지오 나시오날 데 비다 실베스트레 믹스토 마퀜케에서 채집된 두 표본이 다른 ''O. couesi''와 11.93% ''Cytb'' 염기 서열 차이를 보이며, 이는 늪쌀쥐와 ''O. couesi''의 차이(11.30%)만큼 큰 차이라는 것을 발견했다. 그들은 이 동물들이 ''O. couesi''와는 다른 종을 나타낸다고 제안했지만, ''dimidiatus'' 또는 ''richmondi''의 표본을 검사할 수 없었기 때문에 이 종에 적합한 이름을 결정할 수 없었다.[105]
''Oryzomys''는 파나마에서 드물다.[106] 파나마의 ''Oryzomys''는 1912년 골드먼에 의해 처음으로 기술되었으며, 그는 운하 지대의 가툰에서 채집된 표본에 대해 ''Oryzomys gatunensis''라는 이름을 도입했다.[103] 1918년 골드먼은 이 동물을 별개의 종으로 유지하면서, 그 동물이 ''richmondi''와 유사하지만, 안와 부위 가장자리를 따라 잘 발달된 능선, 짧은 두정골 및 긴 비골에서 뚜렷하다고 언급했다.[107] 1937년 볼은 파나마의 또 다른 ''Oryzomys'' 종인 ''Oryzomys azuerensis''를 베라구아스 주의 파라코테에서 기술했다.[102] 이 종은 갈색 형태이며, 주변 개체군의 붉은 색조가 없고, 짧은 로스트럼(앞 부분)과 ''gatunensis''의 것과 같은 안와 부위의 능선이 있는 넓은 두개골을 가지고 있다.[108] 골드먼은 그에게 ''gatunensis''와 ''azuerensis''를 모두 ''couesi''의 아종으로 취급할 것을 권장했지만, 볼은 ''azuerensis''가 지리적으로 가장 가까운 형태인 ''gatunensis''와 ''couesi'' 사이에서 중간 형태를 보이지 않고, 코스타리카 북서부와 니카라과 남동부에서 알려진 가장 가까운 ''O. couesi'' 개체군과 큰 간격으로 떨어져 있기 때문에 ''azuerensis''를 종으로 기술했다.[109] 1966년 파나마 포유류에 대한 검토에서, 찰스 핸들리는 ''gatunensis''와 ''azuerensis''를 모두 늪쌀쥐의 아종으로 축소했고(당시 ''O. couesi''가 포함됨),[110] ''O. couesi''가 별도의 종으로 복원되었을 때 이 형태들도 함께 했다.[18] ''azuerensis''의 기준 산지 근처의 표본은 다른 ''O. couesi''와 ''Cytb'' 염기 서열에서 약 7%의 차이를 보이며, 이는 그들이 별도의 종을 나타낼 수 있음을 시사한다. 그러나 한슨과 동료들은 ''gatunensis''의 표본을 검사할 수 없었기 때문에 ''azuerensis''를 종으로 복원하지 않았다.[35]
''Oryzomys couesi''는 1987년 필립 허시코비츠가 콜롬비아 북서부 코르도바 주의 몬테리아에서의 발생에 대해 보고했을 때 콜롬비아에서 처음 보고되었다.[111] 콜롬비아 표본은 전체적으로 황토색이며 허시코비츠에 따르면 과테말라의 표본과 거의 동일하지만, 윗입술이 흰색이라는 점에서 뚜렷하다. 그는 ''O. couesi''가 콜롬비아 서부의 엘 초코 태평양 저지대에서도 발견될 수 있다고 제안했다.[112]
2. 2. 1. 멕시코 서부 ~ 엘살바도르 (''Oryzomys mexicanus'')
할리스코 (멕시코 서부)에서 엘살바도르 동부에 이르는 ''Oryzomys couesi'' 개체군은 단일 ''Cytb'' 계통을 형성하며, 한슨과 동료들은 이 계통을 종 ''Oryzomys mexicanus''로 인식할 것을 제안했다.[33] 이 동물들은 엄밀한 의미의 ''Oryzomys couesi''와 4.4% 차이를 보이며,[34] 북쪽과 동쪽에서 발견되고, 산맥으로 분리되어 있으며, 다른 종의 한타바이러스를 가지고 있으며, 메리엄(1901)에 따르면 더 튼튼한 두개골을 가지고 있으며 더 큰 어금니, 더 강한 관골궁, 그리고 두개골의 안와 사이 영역 (눈 사이)의 가장자리를 따라 더 잘 발달된 융기를 가지고 있다.[35] "''Oryzomys mexicanus''" 계통 내에서 ''Cytb'' 염기서열 차이는 평균 2.06%이며, 서부(할리스코 ~ 오아하카) 및 동부(치아파스 및 엘살바도르) 그룹은 별개의 하위 계통을 형성한다. 한슨과 동료들은 이들을 서부의 ''mexicanus''와 동부의 ''zygomaticus''로 다른 아종으로 인식했다.[36]2009년 칼튼과 아로요-카브랄레스가 정의한 바에 따르면, 아종 ''Oryzomys couesi mexicanus''는 소노라 중부에서 오아하카 남동부에 이르는 태평양 연안을 따라, 그리고 내륙의 강을 따라 미초아칸 중부, 모렐로스 남부, 푸에블라 남부 및 오아하카 북서부까지 분포한다. 일반적으로 1000m 고도 이하에서 살지만, 할리스코에서는 1525m에서 발견되었다.[37] 이러한 분포 패턴은 ''Sigmodon mascotensis'', ''Hodomys alleni'', ''Peromyscus perfulvus'', 그리고 ''Osgoodomys banderanus''와 같은 다른 멕시코 서부 설치류와 유사하며, 일부 검토에서 별개의 생물지리 구역으로 인식되어 왔다.[41] ''O. c. mexicanus''는 다른 세 종의 ''Oryzomys''—''O. albiventer'', ''O. peninsulae'', 그리고 ''O. nelsoni''—과 가깝게 서식하는데, 이들은 더 크고 비율과 색상의 세부 사항이 다르다.[38]
조엘 아사프 앨런은 1897년 할리스코 표본을 바탕으로 ''Oryzomys mexicanus''를 완전한 종으로 처음 기술했다. 같은 출판물에서 그는 또한 인근 나야리트에서 ''Oryzomys bulleri''를 기술했지만, 이 두 종을 서로 비교하지는 않았다. 메리엄은 1901년에 나야리트에서 두 번째 종인 ''Oryzomys rufus''를 추가했는데, 이는 ''mexicanus''보다 작고 더 붉다는 점에 주목했다. 골드만은 1918년에 이 세 종을 ''O. couesi mexicanus''로 동의어로 처리했으며, 2009년 칼튼과 아로요-카브랄레스는 ''rufus''와 ''mexicanus''의 차이가 연령 관련성이 있으며 동물의 정상적인 변동 범위 내에 있다고 주장하며 이에 동의했다.[39] 또 다른 아종인 ''Oryzomys couesi lambi''는 1934년 버트에 의해 소노라 중부 해안에서 기술되었으며,[29] 당시 종의 범위를 400mi 확장했다. 이 형태는 짙은 회색-갈색으로 ''mexicanus''보다 훨씬 어둡고 꼬리가 짧으며 관골이 약하다.[40] 칼튼과 아로요-카브랄레스는 이것이 ''mexicanus''와 유사하지만, 아종으로 인정해야 하는지 여부를 결정하기 위해서는 추가 연구가 필요하다고 썼다. 시날로아 북부의 큰 ''O. couesi''도 이 형태에 속할 수 있다.[41] 골드만은 ''mexicanus''가 기준 ''couesi''와 매우 유사하지만, 일반적으로 털이 더 창백하다고 썼다. 상체는 ''couesi''보다 더 황갈색이며, 하체는 일반적으로 흰색이지만 황갈색일 수 있으며, 이는 ''couesi''의 일반적인 색상이다.[42]
''Oryzomys zygomaticus''는 메리엄이 1901년에 별도의 종으로 처음 기술했으며,[32] ''mexicanus''와 유사하지만 관골궁이 넓게 퍼져 아래로 굽어 있다.[73] 이를 ''couesi''의 아종으로 축소한 골드만은 과테말라 남서부와 인근 치아파스에서 이를 기록했으며, ''O. c. couesi''보다 약간 창백하지만 ''O. c. mexicanus''보다 어둡다고 기술했다.[43] 엘살바도르 중부의 세 표본은 ''zygomaticus''와 유사한 ''Cytb'' 염기서열을 가지고 있지만,[44] 『엘살바도르의 포유류』(1961)에서 버트와 스티어튼은 이 나라에서 아종 ''couesi''만을 기록했으며, 일부 지역의 표본이 다른 표본보다 약간 창백하다는 점에 주목했다.[45]
2. 2. 2. 멕시코 내륙
골드만은 멕시코 중부 내륙 고원의 4종의 쿠스쌀쥐 아종, 즉 'albiventer', 'crinitus', 'aztecus', 'regillus'를 함께 묶었다.[49] 이 중 3종('albiventer'는 할리스코,[50] 'crinitus'는 연방구,[47] 'aztecus'는 모렐로스)[46]는 메리엄이 1901년에 기술했고, 골드만은 1915년에 미초아칸에서 'regillus'를 직접 기술했다.[48] 골드만에 따르면, 'aztecus'는 창백하고 이빨이 크고,[51] 'crinitus'는 크고 어둡고 이빨이 크며,[52] 'regillus'는 크고 어둡고,[49] 'albiventer'는 크고 비교적 창백하다.[53]2009년 멕시코 서부의 ''Oryzomys''에 대한 검토에서, 칼턴과 아로요-카브랄레스는 명확한 형태 계측적 차이를 근거로 평지 ''mexicanus''와는 별개의 종으로 ''Oryzomys albiventer''를 분류했다.[54] 또한 'crinitus', 'regillus', 'aztecus'의 지위에 대한 몇 가지 의견을 제시했으며, 3가지 형태의 모식표본을 형태 계측 분석에 포함했다. 'regillus'와 'aztecus'의 모식표본은 멕시코 서부 저지대에서 발견된 대규모 ''mexicanus'' 표본의 변이 범위 상한에 속했고, 'crinitus'는 바하칼리포르니아 반도 끝자락에서 발견된 ''O. peninsulae'' 표본과 함께 묶였다. 그들은 'regillus'와 'aztecus'가 ''mexicanus''의 튼튼한 고지대 개체군에 지나지 않을 수 있지만, 다른 종을 나타낼 가능성을 배제할 수 없다고 제안했다. 멕시코 계곡의 2000 m 이상 고도에서 발견되는 'crinitus'가 바하칼리포르니아 반도 저지대의 'peninsulae'와 같은 종이라는 점은 받아들일 수 없었고, 'crinitus'의 관계를 결정하기 위한 추가 연구를 권장했다.[55] 미초아칸 내륙의 한 표본은 ''Cytb'' 데이터가 ''mexicanus''의 특징을 나타냈지만,[56] 한슨과 동료들은 다른 멕시코 내륙 ''Oryzomys''에 대한 데이터를 가지고 있지 않았다.[57]
토마스가 1893년에 기술한 종인 ''Oryzomys fulgens''의 모식표본은 "멕시코"라는 더 정확한 지역이 없지만, 멕시코 계곡이 그 기원으로 제안되었다.[58] 그것은 크고, 거친 털을 가진, 밝은 적갈색,[13] 넓게 퍼진 광대뼈 아치를 가진 넓은 두개골을 가진 긴 꼬리 종이다.[59] 골드만은 이것이 'crinitus'와 유사하지만 색상이 더 강렬하고 안와 간 부위의 형태가 다르다고 기록했으며, 더 많은 조사가 있을 때까지 별도의 종으로 유지했다.[60] 칼턴과 아로요-카브랄레스는 보관 연구를 통해 다른 멕시코 중부 ''Oryzomys''의 이전 이름으로 이를 확립할 수 있는 ''O. fulgens''의 정확한 기원이 밝혀질 수 있다고 언급했다.[61]
2. 2. 3. 텍사스 ~ 니카라과 (''Oryzomys couesi'')
텍사스에서 니카라과까지의 ''Oryzomys'' 개체군은 평균 염기서열 차이가 1.28%인 단일 ''Cytb'' 분류군을 형성하며,[74] Hanson과 동료들은 이 분류군에 ''Oryzomys couesi''라는 이름을 제한할 것을 제안했다.[27] 이러한 개체군은 Goldman이 인정한 두 아종(''O. c. aquaticus'' 및 ''O. c. couesi'')과 종으로 유지한 섬 형태(''O. cozumelae'')에 해당한다. Goldman이 인정한 다른 두 아종, ''O. c. richmondi''와 ''O. c. peragrus'' 및 Goldman의 논문 이후에 설명된 세 번째 아종, ''O. c. pinicola''는 동일한 지역에서 발생하지만 유전적으로 연구되지 않았다.[75]''Oryzomys couesi''의 최북단 개체군, 즉 텍사스 남부와 인근 멕시코 타마울리파스주에 있는 개체군은 1891년에 별도의 종, ''Oryzomys aquaticus''로 기술된 아종 ''aquaticus''로 분류된다.[77] 여기에서 ''O. couesi''의 분포 범위는 습지쌀쥐의 분포 범위와 만난다.[78] 텍사스주 케네디 카운티, 윌라시 카운티 및 카메론 카운티의 일부와 타마울리파스주 북동부에서 이 두 종은 공존한다(같은 장소에서 발생). 접촉 구역에서 ''couesi''는 내륙에서 더 멀리 발생하고, 습지 쌀쥐는 해안을 따라 산다.[79] 실험 조건에서 두 종은 교배에 실패하고[80] 유전자 분석 결과 야생에서 유전자 흐름이나 잡종화의 증거가 없다.[81] 남쪽의 개체군과 비교하여 ''aquaticus''는 더 크고 창백하며 더 강건한 두개골을 가지고 있다.[82] 타마울리파스주에서 온 표본은 텍사스에서 온 표본보다 약간 더 어둡다.[83] ''aquaticus'' 표본의 ''Cytb'' 염기서열은 별도의 그룹을 형성하지만, 더 남쪽에서 온 ''O. c. couesi'' 표본 내에 무리를 이룬다.[84]
''peragrus'' 형태는 멕시코 남부의 산루이스포토시주 리오 베르데 분지, 이달고주, 그리고 베라크루스주 북부에서 알려져 있다.[85] 이 형태의 플라이스토세 후기 화석이 타마울리파스주 쿠에바 데 아브레에서 발견되었다.[86] Goldman에 따르면, 그것은 ''O. c. aquaticus''와 ''O. c. couesi'' 사이의 중간 색상을 가지지만 ''aquaticus''와 유사한 두개골을 가지고 있다.[87]
Goldman은 베라크루스 북부에서 멕시코 동부, 과테말라, 온두라스, 니카라과를 거쳐 코스타리카 북서부까지 이르는 개체군을 명명 아종인 ''Oryzomys couesi couesi''로 통합했다. 그는 베라크루스에서 온 Allen과 Chapman의 1897년 종인 ''Oryzomys jalapae'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys jalapae rufinus'', 타바스코주에서 온 Merriam의 1901년 종인 ''Oryzomys teapensis'', 베라크루스에서 온 Merriam의 1901년 종인 ''Oryzomys goldmani'', 베라크루스에서 온 Eliot의 1904년 종인 ''Oryzomys jalapae apatelius'' 및 니카라과에서 온 Allen의 1910년 종인 ''Oryzomys richardsoni''를 이 형태의 완전한 동의어로 배치했다.[88] Goldman에 따르면, 아종 내의 개인적 변이는 광범위하여 많은 수의 출판된 동의어를 낳았지만, 분포 범위의 모든 부분에서 온 개체군은 본질적으로 유사하다.[89]
아종 ''Oryzomys couesi pinicola''는 1932년에 서부 영국령 온두라스(현재 벨리즈)의 소나무 능선에서 기술되었다. 이 종은 명명 아종인 ''couesi''보다 작고 어두우며, 벨리즈에서도 발생하며, 더 섬세한 두개골을 가지고 있다.[90] 1901년에 Merriam은 코수멜 섬의 ''Oryzomys''를 별도의 종인 ''Oryzomys cozumelae''로 기술했으며, Goldman은 큰 크기, 어두운 털, 긴 꼬리 때문에 이를 유지했다.[91] 그러나 1965년에 Knox Jones와 Timothy Lawlor는 ''cozumelae''와 본토 ''couesi'' 사이의 차이가 사소하다고 판단하고 ''cozumelae''가 본토 ''Oryzomys'' 개체군의 변동 범위 내에 있음을 발견했다. 따라서 그들은 이 섬 형태를 아종으로 강등시켰다.[92] 1989년에 Mark Engstrom과 동료들은 이 결론을 재확인했다.[93] 섬 형태의 경우, 이 개체군은 유전적으로 매우 다양하다.[94] ''Cytb'' 염기서열 데이터에서 명명 아종인 ''couesi''의 개체군에 속한다.[57] ''Oryzomys couesi''는 벨리즈 해안의 턴네페 환초와 온두라스 해안의 로아탄에서도 발견된다.[95]
니카라과 동부 저지대의 ''Oryzomys''는 1901년에 Merriam에 의해 별도의 종인 ''Oryzomys richmondi''로 기술되었으며, Goldman은 뚜렷하게 어두운 털을 근거로 ''O. couesi''의 아종으로 유지했다.[97] 1986년에 니카라과 ''Oryzomys''를 검토하면서 Jones와 Engstrom은 색상 차이가 아종을 인식하기에는 너무 작다고 생각하여 ''richmondi''를 별도로 유지하지 않았다.[98] 회색 아래 부분을 가진 작고 어두운 ''Oryzomys''인 ''Oryzomys dimidiatus''는 니카라과 남동부에서 ''O. couesi''와 함께 발생한다.[99] Jones와 Engstrom에 따르면, 니카라과 호의 오메테페 섬에서 온 쌀쥐는 큰 두개골과 작은 외부 측정값, 특히 짧은 꼬리, 위는 오렌지색-갈색이고 아래는 황갈색인 부드러운 털, 접형골 와 (sphenopalatine vacuities)(뼈 구개 끝 뒤의 간극인 중간 익상와의 열린 부분)의 부재로 특징적이다. 그들은 이 개체군이 별도의 아종을 나타낼 가능성이 높다고 생각했지만, 성체 표본이 하나뿐이었기 때문에 새로운 이름을 제안하는 것을 거부했다.[100] 니카라과에서 ''O. couesi''는 최대 1250 m의 고도에서 발생한다.[101]
2. 2. 4. 코스타리카, 파나마, 콜롬비아
쿠스쌀쥐(''Oryzomys couesi'')는 중간 크기에서 큰 쥐[192]로, 등쪽은 황갈색에서 붉은색을 띠고 옆구리와 뺨으로 갈수록 옅어지며 엉덩이와 얼굴은 더 어두운 거친 털을 가지고 있다. 배쪽은 흰색에서 황갈색이다. 털은 늪쌀쥐보다 짧고, 더 밝으며, 색상이 더 강렬하다. 주둥이는 무디게 끝나고 적당히 큰 눈은 붉은색의 눈부심을 보인다. 작은 귀는 바깥쪽은 검은색이고 안쪽은 짧고 회색에서 황갈색 또는 붉은색 털로 덮여 있다. 긴 꼬리는 위쪽은 짙은 갈색이고 아래쪽은 흰색에서 밝은 갈색이다.[128] 발은 길고 튼튼하다.[129] 앞발에는 발톱 털이 있다.[130] 뒷발의 많은 발바닥은 줄어들었고, 발톱 털도 줄어들었으며,[131] 작발가락 사이 물갈퀴가 적어도 일부 표본에 존재할 수 있다.[6] 이러한 특성 중 일부는 쌀쥐족에서 물속 생활에 대한 일반적인 적응이다.[132] 다른 대부분의 쌀쥐족처럼, 암컷은 8개의 젖꼭지를 가지고 있다.[133] 머리와 몸통의 길이는 98~142mm, 꼬리 길이는 107~152mm, 뒷발 길이는 27~33mm, 귀 길이는 13~18mm, 체중은 43~82g이다.[129] 텍사스와 엘살바도르에서 수행된 연구에 따르면 수컷이 암컷보다 약간 더 크다.[134]
코스타리카의 ''Oryzomys''는 역사적으로 ''O. c. couesi''로 언급되어 왔지만,[104] 한슨과 동료들은 코스타리카 북동부의 레푸지오 나시오날 데 비다 실베스트레 믹스토 마퀜케에서 채집된 두 표본이 다른 ''O. couesi''와 11.93% ''Cytb'' 염기 서열 차이를 보이며, 이는 늪쌀쥐와 ''O. couesi''의 차이(11.30%)만큼 큰 차이라는 것을 발견했다. 그들은 이 동물들이 ''O. couesi''와는 다른 종을 나타낸다고 제안했지만, ''dimidiatus'' 또는 ''richmondi''의 표본을 검사할 수 없었기 때문에 이 종에 적합한 이름을 결정할 수 없었다.[105]
''Oryzomys''는 파나마에서 드물다.[106] 파나마의 ''Oryzomys''는 1912년 골드먼에 의해 처음으로 기술되었으며, 그는 운하 지대의 가툰에서 채집된 표본에 대해 ''Oryzomys gatunensis''라는 이름을 도입했다.[103] 1918년 골드먼은 이 동물을 별개의 종으로 유지하면서, 그 동물이 ''richmondi''와 유사하지만, 안와 부위 가장자리를 따라 잘 발달된 능선, 짧은 두정골 및 긴 비골에서 뚜렷하다고 언급했다.[107] 1937년 볼은 파나마의 또 다른 ''Oryzomys'' 종인 ''Oryzomys azuerensis''를 베라구아스 주의 파라코테에서 기술했다.[102] 이 종은 갈색 형태이며, 주변 개체군의 붉은 색조가 없고, 짧은 로스트럼(앞 부분)과 ''gatunensis''의 것과 같은 안와 부위의 능선이 있는 넓은 두개골을 가지고 있다.[108] 골드먼은 그에게 ''gatunensis''와 ''azuerensis''를 모두 ''couesi''의 아종으로 취급할 것을 권장했지만, 볼은 ''azuerensis''가 지리적으로 가장 가까운 형태인 ''gatunensis''와 ''couesi'' 사이에서 중간 형태를 보이지 않고, 코스타리카 북서부와 니카라과 남동부에서 알려진 가장 가까운 ''O. couesi'' 개체군과 큰 간격으로 떨어져 있기 때문에 ''azuerensis''를 종으로 기술했다.[109] 1966년 파나마 포유류에 대한 검토에서, 찰스 핸들리는 ''gatunensis''와 ''azuerensis''를 모두 늪쌀쥐의 아종으로 축소했고(당시 ''O. couesi''가 포함됨),[110] ''O. couesi''가 별도의 종으로 복원되었을 때 이 형태들도 함께 했다.[18] ''azuerensis''의 기준 산지 근처의 표본은 다른 ''O. couesi''와 ''Cytb'' 염기 서열에서 약 7%의 차이를 보이며, 이는 그들이 별도의 종을 나타낼 수 있음을 시사한다. 그러나 한슨과 동료들은 ''gatunensis''의 표본을 검사할 수 없었기 때문에 ''azuerensis''를 종으로 복원하지 않았다.[35]
''Oryzomys couesi''는 1987년 필립 허시코비츠가 콜롬비아 북서부 코르도바 주의 몬테리아에서의 발생에 대해 보고했을 때 콜롬비아에서 처음 보고되었다.[111] 콜롬비아 표본은 전체적으로 황토색이며 허시코비츠에 따르면 과테말라의 표본과 거의 동일하지만, 윗입술이 흰색이라는 점에서 뚜렷하다. 그는 ''O. couesi''가 콜롬비아 서부의 엘 초코 태평양 저지대에서도 발견될 수 있다고 제안했다.[112]
3. 형태
위는 시그모돈타아과의 특징적인 패턴(1실-반선상): 각 절개에 의해 두 개의 방으로 나뉘지 않고 앞부분(전정)은 선상 상피로 덮여 있다.[135] 담낭은 쌀쥐족의 공유파생형질 (공유된 파생 형질)으로 존재하지 않는다.[136] 핵형은 56개의 염색체와 56개의 상염색체 팔(2n = 56, FNa = 56)의 기본 수를 포함한다. 상염색체는 긴 팔과 매우 짧은 팔을 가진 26쌍의 착사중심 염색체와 한 팔이 다른 팔보다 짧은 중간 크기의 부착중심 염색체 쌍을 포함한다.[137] X 염색체는 긴 팔과 짧은 팔을 가진 착사중심 염색체이거나, 길고 흔적이 있는 팔을 가진 말단착사중심 염색체이다.[138] 성 염색체의 형태는 늪쌀쥐와 ''Oryzomys couesi''를 구별하는 데 사용되었지만, 둘 사이에는 일관된 차이점이 없다.[139]
시그모돈타아과의 특징과 같이 ''Oryzomys couesi''는 복잡한 음경을 가지고 있으며, 음경골은 끝에 세 개의 연골 돌기로 끝난다.[140] 음경의 바깥 표면은 대부분 음경 가시로 덮여 있지만, 비가시 조직의 넓은 띠가 있다.[141] 음경의 등쪽(위쪽)에 있는 유두(젖꼭지 모양의 돌출부)는 작은 가시로 덮여 있으며, 이 특징은 조사된 쌀쥐족 중에서 ''Oryzomys couesi''가 작은쌀쥐속과 늪쌀쥐와만 공유한다.[142] 음경 끝의 구덩이에 위치한 요도 돌기에서[143] 육질의 돌기(아첨단 소엽)가 존재하며, 늪쌀쥐와 ''Holochilus brasiliensis''를 제외하고는 연구된 음경을 가진 다른 모든 쌀쥐족에서 존재하지 않는다.[144]
=== 두개골 ===
비골과 전상악골은 누골, 전두골, 상악골이 만나는 지점 뒤로 확장되지 않는다.[76] 관골판은 관골궁의 납작한 앞부분으로 넓으며 앞쪽 끝에 노치가 발달한다.[145] 판의 뒷쪽 가장자리는 첫 번째 위 어금니 앞에 위치한다.[146] 관골궁의 일부인 협골은 쌀쥐족에서 흔히 볼 수 있듯이 줄어들었다.[147] 어금니 위 두개골 측면에 있는 구멍(공간)인 접형구개공은 작다. 이는 늪쌀쥐에서 훨씬 더 크다.[148] 안와 사이 영역의 가장 좁은 부분은 앞쪽을 향하고 가장자리는 눈에 띄는 선반으로 둘러싸여 있다.[149] 두정골은 뇌두의 측면까지 확장된다.[150] 간두정골은 좁고 쐐기 모양이어서 두정골과 측두골이 광범위하게 만난다.[151]
절치공은 구개 앞부분의 구멍으로 어금니 사이 뒤쪽까지 뻗어 있다.[152] 구개는 길고 세 번째 어금니 뒤로 상당히 확장되어 쌀쥐족의 일반적인 상태이다.[147] 세 번째 어금니 근처의 뒷부분에는 일반적으로 와(함몰) 안으로 들어간 눈에 띄는 후외측 구개와가 천공되어 있다.[153] 접형구개 공포는 일반적으로 없지만 일부 개체군에서 보고되었다.[154] 일부 쌀쥐족에서 두개골의 두 개의 공극을 분리하는 접형골의 확장인 날개접형골 지주가 없다.[155] 머리 동맥의 상태는 매우 공유 파생형이다.[156] 측두하창은 측두골의 모양에 따라 결정되는 두개골 뒷부분의 구멍이 존재한다.[157] 측두골에는 고실개 즉 고실강의 지붕에 접촉하는 지지 과정이 없으며, 이는 쌀쥐족의 정의적인 특징이다.[158] 유양돌기에 일부 구멍이 있다.[159]
하악골 (아래턱)에서 첫 번째 어금니 바로 앞에 있는 구멍인 이공은 다른 몇몇 쌀쥐족에서와 같이 위쪽이 아닌 옆쪽으로 열린다.[160] 일부 씹는 근육을 고정하는 위 및 아래 교근 융기는 첫 번째 어금니 아래 지점에서 합쳐지며 그 지점을 넘어 앞쪽으로 확장되지 않는다.[161] 피막돌기는 하악골 뒤쪽의 뼈가 위로 솟아 앞니의 뒤쪽 끝을 수용하며 크다.[162]
=== 치아 ===
쿠스쌀쥐의 치식은 1.0.0.3/1.0.0.3 (위턱 앞니 1개, 아래턱 앞니 1개, 턱 양쪽에 위 어금니 3개, 아래 어금니 3개)이며,[192] 이는 쥐상과 설치류에서 일반적인 형태이다.[164] 위 앞니는 후굴치로, 씹는 가장자리가 치아의 수직 평면 뒤에 위치한다.[165] 어금니는 구치로, 융선보다 융기가 높으며, 다른 대부분의 쌀쥐족과 마찬가지로 단관치, 즉 치관이 낮다.[166] 위 어금니의 중엽과 아래 어금니의 중엽을 포함한 많은 보조 융선이 존재하며, 이는 ''O. couesi''가 다른 쌀쥐족 대부분과 공유하는 또 다른 특징이다.[167] 어금니의 순측(바깥쪽) 측면의 굴곡 및 굴곡(융기 및 융선 사이의 계곡)은 치대 (능선)로 닫혀 있다.[168]
첫 번째 및 두 번째 위 어금니에서 굴곡은 어금니의 중앙선까지 확장되지 않는다.[168] 위 첫 번째 어금니의 앞쪽 융기인 전원추는 앞쪽의 함몰부(전중선 굴곡)에 의해 두 부분으로 나뉘지 않는다. 전엽이라는 융선이 순측 소엽 뒤에 존재한다.[169] 대부분의 쌀쥐족과 마찬가지로 위 어금니는 모두 내측(설측) 측면에 뿌리가 하나 있고 외측(순측) 측면에 뿌리가 두 개 있으며, 또한 첫 번째 위 어금니는 일반적으로 또 다른 작은 순측 뿌리를 갖는다.[165]
첫 번째 아래 어금니에서 전원소립의 순측 및 설측 원소립은 전중측 와에 의해 분리된다.[170] 두 번째 아래 어금니는 다른 쌀쥐족에서 어금니의 앞쪽 가장자리를 형성하는 두 개의 융기인 원형원 및 후원소립 앞에 전엽이라는 융기를 갖는다.[171] 원형원 앞에 어금니의 바깥 앞쪽(전순측) 가장자리에 뚜렷한 융선(전순측 치대)이 있다.[170] 세 번째 아래 어금니도 전엽과 전순측 치대를 갖는다.[172] 첫 번째 아래 어금니는 치아 앞쪽과 뒤쪽에 큰 뿌리가 있고, 순측 및 설측 측면에 그 사이에 두 개의 작은 뿌리가 있다. 두 번째 및 세 번째 아래 어금니는 앞쪽에 하나, 뒤쪽에 하나, 두 개의 큰 뿌리가 있다.[165]
=== 골격 ===
쌀쥐족에서 흔히 볼 수 있듯이, 12개의 갈비뼈가 있다. 첫 번째 갈비뼈는 마지막 경추와 첫 번째 흉추에 모두 관절하며, 이는 시그모돈티나아과의 공유 파생 형질이다.[173] 척추 뒤쪽의 돌기인 부신경돌기는 다섯 번째 요추에 존재하지 않는다.[174] 두 번째와 세 번째 미추 사이에는 가시 모양의 뒷면을 가진 혈관궁(작은 뼈)이 존재한다.[175] 내상과공은 시그모돈티나아과의 모든 구성원에서와 마찬가지로 존재하지 않는다. 다른 설치류에서처럼 존재한다면, 이 구멍은 상완골 (위팔뼈)의 원위부(먼) 끝을 관통한다.[176]
3. 1. 두개골
비골과 전상악골은 누골, 전두골, 상악골이 만나는 지점 뒤로 확장되지 않는다.[76] 관골판은 관골궁의 납작한 앞부분으로 넓으며 앞쪽 끝에 노치가 발달한다.[145] 판의 뒷쪽 가장자리는 첫 번째 위 어금니 앞에 위치한다.[146] 관골궁의 일부인 협골은 오리조미니에서 흔히 볼 수 있듯이 줄어들었다.[147] 어금니 위 두개골 측면에 있는 공간(구멍)인 접형구개공은 작다. 이는 늪쌀쥐에서 훨씬 더 크다.[148] 안와 사이 영역의 가장 좁은 부분은 앞쪽을 향하고 가장자리는 눈에 띄는 선반으로 둘러싸여 있다.[149] 두정골은 뇌두의 측면까지 확장된다.[150] 간두정골은 좁고 쐐기 모양이어서 두정골과 측두골이 광범위하게 만난다.[151]
절치공은 구개 앞부분의 구멍으로 어금니 사이 뒤쪽까지 뻗어 있다.[152] 구개는 길고 세 번째 어금니 뒤로 상당히 확장되어 오리조미니의 일반적인 상태이다.[147] 세 번째 어금니 근처의 뒷부분에는 일반적으로 와(함몰) 안으로 들어간 눈에 띄는 후외측 구개와가 천공되어 있다.[153] 접형구개 공포는 일반적으로 없지만 일부 개체군에서 보고되었다.[154] 일부 오리조미니에서 두개골의 두 개의 공극을 분리하는 접형골의 확장인 날개접형골 지주가 없다.[155] 머리 동맥의 상태는 매우 공유 파생형이다.[156] 측두하창은 측두골의 모양에 따라 결정되는 두개골 뒷부분의 구멍이 존재한다.[157] 측두골에는 고실개 즉 고실강의 지붕에 접촉하는 지지 과정이 없으며, 이는 오리조미니의 정의적인 특징이다.[158] 유양돌기에 일부 구멍이 있다.[159]
하악골 (아래턱)에서 첫 번째 어금니 바로 앞에 있는 구멍인 이공은 다른 몇몇 오리조미니에서와 같이 위쪽이 아닌 옆쪽으로 열린다.[160] 일부 씹는 근육을 고정하는 위 및 아래 교근 융기는 첫 번째 어금니 아래 지점에서 합쳐지며 그 지점을 넘어 앞쪽으로 확장되지 않는다.[161] 피막돌기는 하악골 뒤쪽의 뼈가 위로 솟아 앞니의 뒤쪽 끝을 수용하며 크다.[162]
3. 2. 치아
쿠스쌀쥐의 치식은 (위턱 앞니 1개, 아래턱 앞니 1개, 턱 양쪽에 위 어금니 3개, 아래 어금니 3개)이며,[192] 이는 쥐상과 설치류에서 일반적인 형태이다.[164] 위 앞니는 후굴치로, 씹는 가장자리가 치아의 수직 평면 뒤에 위치한다.[165] 어금니는 구치로, 융선보다 융기가 높으며, 다른 대부분의 오리자미아족과 마찬가지로 단관치, 즉 치관이 낮다.[166] 위 어금니의 중엽과 아래 어금니의 중엽을 포함한 많은 보조 융선이 존재하며, 이는 ''O. couesi''가 다른 오리자미아족 대부분과 공유하는 또 다른 특징이다.[167] 어금니의 순측(바깥쪽) 측면의 굴곡 및 굴곡(융기 및 융선 사이의 계곡)은 치대 (능선)로 닫혀 있다.[168]
첫 번째 및 두 번째 위 어금니에서 굴곡은 어금니의 중앙선까지 확장되지 않는다.[168] 위 첫 번째 어금니의 앞쪽 융기인 전원추는 앞쪽의 함몰부(전중선 굴곡)에 의해 두 부분으로 나뉘지 않는다. 전엽이라는 융선이 순측 소엽 뒤에 존재한다.[169] 대부분의 오리자미아족과 마찬가지로 위 어금니는 모두 내측(설측) 측면에 뿌리가 하나 있고 외측(순측) 측면에 뿌리가 두 개 있으며, 또한 첫 번째 위 어금니는 일반적으로 또 다른 작은 순측 뿌리를 갖는다.[165]
첫 번째 아래 어금니에서 전원소립의 순측 및 설측 원소립은 전중측 와에 의해 분리된다.[170] 두 번째 아래 어금니는 다른 오리자미아족에서 어금니의 앞쪽 가장자리를 형성하는 두 개의 융기인 원형원 및 후원소립 앞에 전엽이라는 융기를 갖는다.[171] 원형원 앞에 어금니의 바깥 앞쪽(전순측) 가장자리에 뚜렷한 융선(전순측 치대)이 있다.[170] 세 번째 아래 어금니도 전엽과 전순측 치대를 갖는다.[172] 첫 번째 아래 어금니는 치아 앞쪽과 뒤쪽에 큰 뿌리가 있고, 순측 및 설측 측면에 그 사이에 두 개의 작은 뿌리가 있다. 두 번째 및 세 번째 아래 어금니는 앞쪽에 하나, 뒤쪽에 하나, 두 개의 큰 뿌리가 있다.[165]
3. 3. 골격
오리조미스족에서 흔히 볼 수 있듯이, 12개의 갈비뼈가 있다. 첫 번째 갈비뼈는 마지막 목뼈와 첫 번째 등뼈에 모두 관절하며, 이는 시그모돈티나아과의 공유 파생 형질이다.[173] 척추 뒤쪽의 돌기인 부신경돌기는 다섯 번째 허리뼈에 존재하지 않는다.[174] 두 번째와 세 번째 꼬리뼈 사이에는 가시 모양의 뒷면을 가진 혈관궁(작은 뼈)이 존재한다.[175] 내상과공은 시그모돈티나아과의 모든 구성원에서와 마찬가지로 존재하지 않는다. 다른 설치류에서처럼 존재한다면, 이 구멍은 상완골 (위팔뼈)의 원위부(먼) 끝을 관통한다.[176]
4. 생태
쿠스쌀쥐(''Oryzomys couesi'')는 땅에서 생활하며 반수생으로, 물에서 많은 시간을 보낸다.[106] 나무를 잘 타기도 한다.[183] 코스타리카에서의 연구에 따르면 쿠스쌀쥐는 수영과 잠수를 잘하며, 꼬리를 사용하여 추진력을 얻는다. 잠수 능력은 생태학적으로 유사한 면 쥐 ''Sigmodon hirsutus''와의 지위 분화에 기여한다.[184] 방해를 받으면 물에 들어가 헤엄쳐 도망간다.[106] 주로 야행성이다.[185]
갈대 사이의 물이나 땅 위 약 1m 높이에 엮인 식물로 둥근 둥지를 짓는다.[186] 텍사스에서는 더 큰 개체가 더 큰 둥지를 짓는다.[187] 일반적으로 식생에 자체 통로를 만들지 않지만 면 쥐와 같은 다른 설치류의 통로를 사용할 수 있다.[193]
개체군 밀도는 ha당 5~30마리이다.[1] 코수멜에서는 ha당 약 14.5~16.5마리이지만 계절에 따라 큰 변동을 보인다.[188] 멕시코 서부의 한 연구에 따르면 구름 숲에서는 ha당 3마리이고, 교란된 지역에서는 ha당 1마리였다.[189] 24시간 동안 텍사스 수컷 쿠스쌀쥐는 최대 153m까지 이동하고 암컷은 최대 126m까지 이동한다.[190]
식단에는 씨앗과 녹색 부분과 같은 식물 재료와 작은 물고기, 갑각류, 달팽이, 개미와 딱정벌레와 같은 곤충, 기타 무척추동물이 포함된다.[191] 일년 내내 번식할 가능성이 있으며, 임신 기간은 21~28일이며,[192] 암컷은 2~7마리의 새끼를 낳으며, Reid의 ''중앙 아메리카 및 남동 멕시코의 포유류''에 따르면 평균 3.8마리이다.[106] 니카라과에서 온 28마리의 임신한 암컷의 경우, 새끼 수는 1~8마리로 다양했으며 평균 4.4마리였다.[126] 새끼는 7주가 되면 생식 능력을 갖게 되며 생물학적 생애 주기는 짧다.[192]
도입종 뱀 ''Boa constrictor''는 코수멜에서 쿠스쌀쥐를 포식한다.[181] 베라크루스에서 쿠스쌀쥐에서 기록된 기생충으로는 확인되지 않은 진드기, 진드기, 벼룩 및 파리 유충이 있다.[193] 벼룩 ''Polygenis odiosus''는 코수멜에서 온 쿠스쌀쥐에서 발견되었다.[194] 산루이스포토시의 10마리의 쿠스쌀쥐 중 5마리씩은 선충류 벌레 ''Hassalstrongylus musculi''와 ''H. bocqueti''에 감염되었으며, 쥐당 약 25마리의 벌레가 있었고, 2마리는 ''Raillietina'' 속의 1~2개의 촌충에 감염되었다.[195] 진드기 ''Eubrachylaelaps circularis''와 ''Gigantolaelaps boneti''는 오악사카에서 쿠스쌀쥐에서 발견되었으며,[196] 빨판이 ''Hoplopleura oryzomydis''는 니카라과에서,[197] 진드기 ''Laelaps oryzomydis'', ''Echinonyssus microchelae'', ''Ornithonyssus bacoti'', ''Prolistrophorus frontalis'', ''Prolistrophorus bakeri''는 콜리마에서, 그리고 아피콤플렉사인 ''Eimeria couesii''는 멕시코에서 발견되었다.[198] 이 종은 두 개의 한타바이러스—온두라스의 Catacamas virus와 멕시코 서부의 Playa de Oro virus—에 감염되는데, 이는 미국에서 한타바이러스 감염의 흔한 원인인 늪 쌀쥐를 감염시키는 Bayou virus와 관련이 있다. 그러나 인간의 한타바이러스 감염이 쿠스쌀쥐 한타바이러스와 연관된 적은 없다.[200] 치아파스 쿠스쌀쥐는 베네수엘라 말 뇌염 바이러스를 포함한 여러 아르보바이러스로 실험적으로 감염되어도 쉽게 살아남아, 이 종이 해당 바이러스의 자연 숙주 역할을 할 수 있음을 시사한다.[201]
4. 1. 행동
쿠스쌀쥐(''Oryzomys couesi'')는 땅에서 생활하며 반수생으로, 물에서 많은 시간을 보낸다.[106] 나무를 잘 타기도 한다.[183] 코스타리카에서의 연구에 따르면 쿠스쌀쥐는 수영과 잠수를 잘하며, 꼬리를 사용하여 추진력을 얻는다. 잠수 능력은 생태학적으로 유사한 면 쥐 ''Sigmodon hirsutus''와의 지위 분화에 기여한다.[184] 방해를 받으면 물에 들어가 헤엄쳐 도망간다.[106] 주로 야행성이다.[185]갈대 사이의 물이나 땅 위 약 1m 높이에 엮인 식물로 둥근 둥지를 짓는다.[186] 텍사스에서는 더 큰 개체가 더 큰 둥지를 짓는다.[187] 일반적으로 식생에 자체 통로를 만들지 않지만 면 쥐와 같은 다른 설치류의 통로를 사용할 수 있다.[193]
개체군 밀도는 ha당 5~30마리 (에이커당 2~12마리)이다.[1] 코수멜에서는 ha당 약 14.5~16.5마리 (에이커당 5.9~6.7마리)이지만 계절에 따라 큰 변동을 보인다.[188] 멕시코 서부의 한 연구에 따르면 구름 숲에서는 ha당 3마리 (에이커당 1.2마리)이고, 교란된 지역에서는 ha당 1마리 (에이커당 0.4마리)였다.[189] 24시간 동안 텍사스 수컷 쿠스쌀쥐는 최대 153m까지 이동하고 암컷은 최대 126m까지 이동한다.[190]
식단에는 씨앗과 녹색 부분과 같은 식물 재료와 작은 물고기, 갑각류, 달팽이, 개미와 딱정벌레와 같은 곤충, 기타 무척추동물이 포함된다.[191] 일년 내내 번식할 가능성이 있으며, 임신 기간은 21~28일이며,[192] 암컷은 2~7마리의 새끼를 낳으며, Reid의 ''중앙 아메리카 및 남동 멕시코의 포유류''에 따르면 평균 3.8마리이다.[106] 니카라과에서 온 28마리의 임신한 암컷의 경우, 새끼 수는 1~8마리로 다양했으며 평균 4.4마리였다.[126] 새끼는 7주가 되면 생식 능력을 갖게 되며 생물학적 생애 주기는 짧다.[192]
도입종 뱀 ''Boa constrictor''는 코수멜에서 쿠스쌀쥐를 포식한다.[181] 베라크루스에서 쿠스쌀쥐에서 기록된 기생충으로는 확인되지 않은 진드기, 진드기, 벼룩 및 파리 유충이 있다.[193] 벼룩 ''Polygenis odiosus''는 코수멜에서 온 쿠스쌀쥐에서 발견되었다.[194] 산루이스포토시의 10마리의 쿠스쌀쥐 중 5마리씩은 선충류 벌레 ''Hassalstrongylus musculi''와 ''H. bocqueti''에 감염되었으며, 쥐당 약 25마리의 벌레가 있었고, 2마리는 ''Raillietina'' 속의 1~2개의 촌충에 감염되었다.[195] 진드기 ''Eubrachylaelaps circularis''와 ''Gigantolaelaps boneti''는 오악사카에서 쿠스쌀쥐에서 발견되었으며,[196] 빨판이 ''Hoplopleura oryzomydis''는 니카라과에서,[197] 진드기 ''Laelaps oryzomydis'', ''Echinonyssus microchelae'', ''Ornithonyssus bacoti'', ''Prolistrophorus frontalis'', ''Prolistrophorus bakeri''는 콜리마에서, 그리고 아피콤플렉사인 ''Eimeria couesii''는 멕시코에서 발견되었다.[198] 이 종은 두 개의 한타바이러스—온두라스의 Catacamas virus와 멕시코 서부의 Playa de Oro virus—에 감염되는데, 이는 미국에서 한타바이러스 감염의 흔한 원인인 늪 쌀쥐를 감염시키는 Bayou virus와 관련이 있다. 그러나 인간의 한타바이러스 감염이 쿠스쌀쥐 한타바이러스와 연관된 적은 없다.[200] 치아파스 쿠스쌀쥐는 베네수엘라 말 뇌염 바이러스를 포함한 여러 아르보바이러스로 실험적으로 감염되어도 쉽게 살아남아, 이 종이 해당 바이러스의 자연 숙주 역할을 할 수 있음을 시사한다.[201]
4. 2. 번식
쿠스쌀쥐는 일년 내내 번식할 가능성이 있으며, 임신 기간은 21~28일이다.[192] 암컷은 2~7마리의 새끼를 낳으며, Reid의 ''중앙 아메리카 및 남동 멕시코의 포유류''에 따르면 평균 3.8마리이다.[106] 니카라과에서 온 28마리의 임신한 암컷의 경우, 새끼 수는 1~8마리로 다양했으며 평균 4.4마리였다.[126] 새끼는 7주가 되면 생식 능력을 갖게 되며 생물학적 생애 주기는 짧다.[192]5. 인간과의 관계
5. 1. 질병 매개
5. 2. 보존 상태
IUCN은 ''쿠스쌀쥐''(Oryzomys couesi)가 널리 분포하고 흔하며 다양한 서식지에 적응하며 많은 보호 구역에 서식하기 때문에 "관심 필요"종으로 분류한다.[1] 습지 파괴와 같은 서식지 파괴는 일부 개체군을 위협할 수 있다.[1] 많은 지역에서 너무 흔해서 해충으로 간주되기도 한다.[192] 2006년에 게재된 사진에서 알 수 있듯이, 멕시코 계곡에서도 개체군이 존재한다.[202] 그러나 이 종은 서식지 손실로 인해 분포가 매우 제한적인 텍사스에서는 위협받는 종으로 지정되었다.[203] 1979년 벤슨과 게일바흐는 텍사스 개체군 규모를 약 15,000마리로 추정했다.[204] 2001년 연구에서는 기후 변화로 인해 적합한 서식지가 더 이상 존재하지 않아 텍사스 개체군이 멸종될 것으로 예측했다.[205] 코수멜 개체군은 1980년대 중반 이후 서식지 교란과 도입된 종의 포식으로 인해 상당히 감소했다.[188]5. 3. 한국과의 관계
참조
[1]
논문
Linzey et al.
2016
[2]
논문
Hanson et al.
2010
[3]
논문
Dickinson
2005
[4]
논문
Dickinson
2005
[5]
논문
Hall
1960
[6]
논문
Carleton and Arroyo-Cabrales
2009
[7]
논문
Hanson et al.
2010
[8]
논문
Weksler et al.
2006
[9]
논문
Weksler
2006
[10]
논문
Musser and Carleton
2005
[11]
논문
Alston
1877
[12]
논문
Alston
1877
[13]
논문
Thomas
1893
[14]
논문
Merriam
1901
[15]
논문
Goldman
1918
[16]
논문
Hall
1960
[17]
논문
Goldman
1918
[18]
논문
Musser and Carleton
2005
[19]
논문
Musser and Carleton
2005
[20]
논문
Carleton and Arroyo-Cabrales
2009
[21]
논문
Carleton and Arroyo-Cabrales
2009
[22]
논문
Carleton and Arroyo-Cabrales
2009
[23]
논문
Hanson et al.
2010
[24]
논문
Hanson et al.
2010
[25]
논문
Hanson et al.
2010
[26]
논문
Hanson et al.
2010
[27]
논문
Hanson et al.
2010
[28]
논문
Allen
1897
[29]
논문
Burt
1934
[30]
논문
Allen
1897
[31]
논문
Merriam
1901
[32]
논문
Merriam
1901
[33]
논문
Hanson et al.
2010
[34]
논문
Hanson et al.
2010
[35]
논문
Hanson et al.
2010
[36]
논문
Hanson et al., 2010, fig. 2, table 1, appendix I
[37]
논문
Carleton and Arroyo-Cabrales, 2009, p. 119
[38]
논문
Carleton and Arroyo-Cabrales, 2009, pp. 118, 121–122
[39]
논문
Carleton and Arroyo-Cabrales, 2009, pp. 118–120; Goldman, 1918, p. 34
[40]
논문
Burt, 1934, p. 108
[41]
논문
Carleton and Arroyo-Cabrales, 2009, p. 120
[42]
논문
Goldman, 1918, p. 34
[43]
논문
Goldman, 1918, p. 33
[44]
논문
Hanson et al., 2010, figs. 1–2, appendix I
[45]
논문
Burt and Stirton, 1961, p. 61
[46]
논문
Merriam, 1901b, p. 282
[47]
논문
Merriam, 1901b, p. 281
[48]
논문
Goldman, 1915, p. 129
[49]
논문
Goldman, 1918, p. 37
[50]
논문
Merriam, 1901b, p. 279
[51]
논문
Goldman, 1918, p. 35
[52]
논문
Goldman, 1918, p. 36
[53]
논문
Goldman, 1918, p. 38
[54]
논문
Carleton and Arroyo-Cabrales, 2009, p. 118
[55]
논문
Carleton and Arroyo-Cabrales, 2009, p. 113
[56]
논문
Hanson et al., 2010, fig. 1, appendix I
[57]
논문
Hanson et al., 2010, fig. 1
[58]
논문
Carleton and Arroyo-Cabrales, 2009, pp. 113–114
[59]
논문
Thomas, 1893, p. 404
[60]
논문
Goldman, 1918, p. 282
[61]
논문
Carleton and Arroyo-Cabrales, 2009, p. 114
[62]
논문
Eliot, 1904, p. 266
[63]
논문
Allen, 1891, p. 289
[64]
논문
Merriam, 1901a, p. 103
[65]
논문
Merriam, 1901a, p. 103; Merriam, 1901b, p. 280
[66]
논문
Poole and Schantz, 1942, p. 306
[67]
논문
Merriam, 1901b, p. 288
[68]
논문
Allen and Chapman, 1897, p. 206
[69]
논문
Merriam, 1901b, p. 283
[70]
논문
Murie, 1932, p. 1
[71]
논문
Allen, 1910, p. 99
[72]
논문
Merriam, 1901b, p. 284
[73]
논문
Merriam, 1901b, p. 286
[74]
논문
Hanson et al., 2010, fig. 1, table 1
[75]
논문
Goldman, 1918, fig. 3; Murie, 1932, p. 1; Handon et al., 2010, fig. 1
[76]
논문
Goldman, 1918, plate I
[77]
논문
Goldman, 1918, pp. 39–40; Hanson et al., 2010, appendix A
[78]
논문
Schmidt and Engstrom, 1994, p. 914
[79]
논문
Schmidt and Engstrom, 1994, pp. 916–917
[80]
논문
Schmidt and Engstrom, 1994, pp. 915–916
[81]
논문
Schmidt and Engstrom, 1994, p. 920
[82]
논문
Goldman, 1918, p. 40
[83]
논문
Baker, 1951, p. 215
[84]
논문
Hanson et al., 2010, fig. 2
[85]
논문
Goldman, 1918, p. 39; Jones et al., 1983, p. 379; Hall and Dalquest, 1963, p. 289
[86]
논문
Dalquest and Roth, 1970, pp. 220, 226
[87]
논문
Goldman, 1918, p. 39
[88]
논문
Goldman, 1918, pp. 29–30
[89]
논문
Goldman, 1918, p. 31
[90]
논문
Murie, 1932, pp. 1–2; Murie, 1935, p. 26
[91]
논문
Goldman, 1918, p. 43
[92]
논문
Jones and Lawlor, 1965, p. 413
[93]
논문
Schmidt and Engstrom, 1989, p. 414
[94]
논문
Vega et al., 2004, p. 210
[95]
논문
Platt et al., 2000, p. 167
[96]
논문
Koopman, 1959, p. 237
[97]
논문
Goldman, 1918, p. 32
[98]
논문
Jones and Engstrom, 1986, p. 10
[99]
논문
Genoways and Jones, 1971, p. 834
[100]
논문
Jones and Engstrom, 1986, pp. 10, 12
[101]
논문
Jones and Engstrom, 1986, p. 7
[102]
논문
Bole, 1937, p. 165
[103]
논문
Goldman, 1912, p. 7
[104]
논문
Goldman, 1918, p. 31; Harris, 1943, p. 12
[105]
논문
Hanson et al., 2010, p. 343, appendix A
[106]
문헌
2009
[107]
문헌
1918
[108]
문헌
1937
[109]
문헌
1937
[110]
문헌
1966
[111]
문헌
1987
[112]
문헌
1987
[113]
문헌
1905
[114]
문헌
1918
[115]
문헌
1905, 1918, 2008, 2004
[116]
문서
Number of specimens measured.
[117]
문서
Only averages available.
[118]
문헌
2009
[119]
문헌
1961
[120]
문헌
1918
[121]
문서
With claws.
[122]
문헌
1918
[123]
문헌
1918
[124]
문헌
1932
[125]
문서
Head and body length instead of total length.
[126]
문헌
1986
[127]
문헌
1918
[128]
문헌
1918, 2009
[129]
문헌
2009
[130]
문헌
2006
[131]
문헌
1989, 2006
[132]
문헌
2006
[133]
문헌
2006
[134]
문헌
1979, 1961
[135]
문헌
2006
[136]
문헌
2006
[137]
문헌
1979
[138]
문헌
2010
[139]
문헌
1994, 2010
[140]
문헌
2006
[141]
서적
Weksler, 2006, pp. 56–57
[142]
서적
Hooper and Musser, 1964, p. 13; Weksler, 2006, p. 57
[143]
서적
Hooper and Musser, 1964, p. 7
[144]
서적
Weksler, 2006, p. 57
[145]
서적
Weksler, 2006, p. 27, table 5
[146]
서적
Weksler, 2006, pp. 31–32
[147]
서적
Weksler, 2006, p. 32
[148]
서적
Weksler, 2006, p. 32, table 5
[149]
서적
Schmidt and Engstrom, 1994, p. 917
[150]
서적
Weksler, 2006, p. 28, table 5
[151]
서적
Weksler, 2006, p. 30
[152]
서적
Weksler, 2006, p. 31
[153]
서적
Weksler, 2006, pp. 34–35
[154]
서적
Weksler, 2006, p. 35
[155]
서적
Weksler, 2006, p. 37; Carleton and Arroyo-Cabrales, 2009, p. 117; Sánchez et al., 2001, p. 210
[156]
서적
Weksler, 2006, p. 38, table 5
[157]
서적
Weksler, 2006, p. 37
[158]
서적
Weksler, 2006, pp. 38–39
[159]
서적
Weksler, 2006, p. 40
[160]
서적
Weksler, 2006, pp. 40–41
[161]
서적
Weksler, 2006, p. 41, table 5
[162]
서적
Weksler, 2006, p. 42
[163]
서적
Weksler, 2006, pp. 41–42
[164]
서적
Carleton and Musser, 1984, p. 292
[165]
서적
Weksler, 2006, p. 43
[166]
서적
Weksler, 2006, pp. 43–44
[167]
서적
Weksler, 2006, pp. 44–49
[168]
서적
Weksler, 2006, p. 44
[169]
서적
Weksler, 2006, pp. 44–45
[170]
서적
Weksler, 2006, p. 49
[171]
서적
Weksler, 2006, p. 52
[172]
서적
Weksler, 2006, p. 53
[173]
서적
Weksler, 2006, p. 52, table 5
[174]
서적
Weksler, 2006, pp. 52–53
[175]
서적
Weksler, 2006, p. 53; fig. 28
[176]
논문
[177]
논문
[178]
논문
[179]
논문
[180]
논문
[181]
논문
[182]
논문
[183]
논문
[184]
논문
[185]
논문
[186]
논문
[187]
논문
[188]
논문
[189]
논문
[190]
논문
[191]
논문
[192]
논문
[193]
논문
[194]
논문
[195]
논문
[196]
논문
[197]
논문
[198]
논문
[199]
논문
[200]
논문
[201]
논문
[202]
논문
[203]
논문
[204]
논문
[205]
논문
[206]
논문
[207]
저널
"''Oryzomys couesi''"
http://www.iucnredli[...]
IUCN
2017-06-25
[208]
논문
[209]
서적
Mammal Species of the World: a taxonomic and geographic reference. 3rd ed
http://www.bucknell.[...]
The Johns Hopkins University Press
[210]
논문
[211]
서적
2005
[212]
서적
1877
[213]
서적
1893
[214]
서적
1960
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com