나균
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
나균(Mycobacterium leprae)은 세포 내 기생 세균으로, 나병을 유발한다. 호기성 간균으로, 미콜산 왁스 코팅을 가지며, 그람 양성으로 염색되지만, Ziehl-Neelsen 염색에서 카볼 푸크신으로 염색된다. 발견 이후 인공 배양에 성공한 사례는 없으며, 누드 마우스의 발바닥이나 아르마딜로에서 증식시킬 수 있다. 숙주 범위는 좁아 인간 외 아홉띠아르마딜로와 붉은 다람쥐가 자연 숙주이며, 유전체는 환원 진화를 겪어 기능 유전자가 적고, 유사유전자가 많다. 대사 능력 저하로 숙주 의존성이 높으며, 11일의 세대 시간을 가진다. 증상은 피부 궤양, 신경 손상, 감각 능력 저하 등을 포함하며, 치료는 다제요법(MDT)을 사용한다. 1873년 게르하르트 아르마우어 한센에 의해 발견되었다.
더 읽어볼만한 페이지
- 게놈 해독이 완료된 세균 - 탄저균
탄저균은 바실러스속의 그람 양성 포자 형성 간균으로, 탄저병을 일으키는 병원체이며, 생물학적 무기로 사용될 수 있을 만큼 강력한 포자를 형성하고 캡슐과 외독소를 통해 숙주를 공격하며, 백신과 항생제로 예방 및 치료가 가능하다. - 게놈 해독이 완료된 세균 - 바킬루스 세레우스
바실루스 세레우스는 1887년 푸르쉬에 의해 분리된, 식품 오염과 관련이 깊은 막대 모양의 그람 양성균으로, 포자 형성, 생물막 형성, 운동성, 다양한 대사 작용, 항생제 내성 유전자 보유 등의 특징을 가지며, 식중독 및 다양한 질병을 유발할 수 있고, 위생 관리를 통해 예방이 가능하다. - 미코박테륨 - 소결핵균
소결핵균은 세포벽의 특성으로 여러 환경에 저항하며, 주로 감염된 소의 생 우유를 통해 인간에게 전파되고, 인수공통 결핵을 일으키며, 이소니아지드와 리팜피신으로 치료하고 BCG 백신의 조상이기도 하다. - 미코박테륨 - 결핵균
결핵균은 결핵을 유발하는 세균으로, 공기 감염을 통해 사람에게 전파되며, 항생제 내성으로 치료가 어려울 수 있고, 가래 검사 등으로 진단하며, 여러 항생제를 병용하는 화학 요법으로 치료한다.
| 나균 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
![피부 병변에서 채취한 [[Mycobacterium leprae]]의 현미경 사진. 작은 벽돌색 막대 모양의 세포가 클러스터로 나타남. 출처: CDC](https://cdn.onul.works/wiki/noimage.png) | |
| 학명 | Mycobacterium leprae |
| 명명자 | 게르하르 아르마우에르 한센, 1874 |
| 기타 명칭 | Hansen's coccus spirilly |
| 생물학적 분류 | |
| 도메인 | 세균 |
| 문 | 방선균문 |
| 강 | 방선균강 |
| 목 | 코리네박테리움목 |
| 과 | 마이코박테리움과 |
| 속 | 마이코박테리움속 |
| 특징 | |
| 관련 질병 | 나병 |
2. 미생물학

''나균''(la)은 세포 내, 다형성, 비포자 형성, 비운동성, 항산성, 병원성 세균이다.[5] 호기성 간균 (막대 모양 세균)으로, 평행한 면과 둥근 끝을 가지며, 미콜산의 특징적인 왁스 코팅으로 둘러싸여 있으며 이는 마이코박테리아에 고유하다. 그람 양성으로 그람 염색되지만, ''나균''은 전통적으로 Ziehl–Neelsen 염색에서 카볼 푸크신으로 염색되었다. 간균이 ''결핵균''(MTB)보다 항산성이 덜하기 때문에, 산 농도가 낮은 Fite-Faraco 염색 방법이 현재 사용된다.[6][7] 크기와 모양은 MTB와 매우 유사하다. 박테리아는 육아종 병변에서 발견되며 특히 결절에서 수가 많다. 이 박테리아는 나병의 나병성 병변 내에서 자주 다량으로 발생하며 일반적으로 울타리 형태로 함께 뭉쳐 있다.[8]
숙주 세포의 광학 현미경 검사를 통해, ''나균''은 단독으로 또는 "글로비"라고 하는 덩어리로 발견될 수 있으며, 간균은 곧거나 약간 구부러져 있고, 길이는 1–8 μm이며 직경은 0.3 μm이다.[9] 이 박테리아는 27~30 °C에서 가장 잘 성장하며, 이는 피부, 비강 점막 및 말초 신경을 ''나균'' 감염의 주요 표적으로 만든다.[10]
가늘고 긴 막대 모양의 세균이다.
발견 이후 현재까지 인공 배양에 성공한 사례는 없다.
누드 마우스의 발바닥 및 아르마딜로에서 증식시킴으로써 얻을 수 있다. 따라서 한센병 치료제 개발에 어려움을 겪었다. 세포 내에서만 증식하는 절대 세포 내 기생체이다. 특히 말초 신경세포에 친화성이 높다.
결핵균의 경우, 기능하고 있는 유전자의 비율이 90.8%인 데 반해, 나균은 49.5%로 극단적으로 적다. 따라서 생존과 증식을 전적으로 숙주 세포에 의존하고 있다.
세대 시간은 약 11일이다. 영장류에 감염된다. 증식하면 잎담배 모양의 배열을 보인다. 또한, 구형의 균괴(나구)를 형성한다.
항산성 염색으로 염색된다. 조직 검사에서는 Ziehl-Neelsen 염색과 Fite 염색이 진단에 활용되고 있다.
2. 1. 배양
나균(''Mycobacterium leprae'')은 배가 시간이 매우 길고(12~14일) 실험실에서 배양할 수 없는 특징을 가지고 있다.[13][14][15] 이는 나균이 절대 세포 내 기생체이기 때문에 독립 생존에 필요한 많은 유전자가 부족하기 때문이다.[16] 결핵균 속의 구성원을 파괴하기 어렵게 만드는 복잡하고 독특한 세포벽은 또한 매우 느린 증식 속도의 원인이기도 하다. 나균은 서늘한 온도, 약간 산성인 미호기성 환경을 선호하며, 당보다 지질을 에너지원으로 사용하는 것을 선호한다. 나균에 필요한 성장 조건은 알려져 있지만, 나균의 성장을 지원하는 정확한 무균 배지는 아직 발견되지 않았다.[16]일반적으로 생체 외 배양이 불가능하므로, 대신 생쥐 발 패드[12] 및 낮은 체온을 가진 아르마딜로에서 배양되었다.[17][16] 발견 이후 현재까지 인공 배양에 성공한 사례는 없다. 누드 마우스의 발바닥 및 아르마딜로에서 증식시킴으로써 얻을 수 있다. 따라서 한센병 치료제 개발에 어려움을 겪었다.
3. 숙주 범위
나균은 좁은 숙주 범위를 가지며, 인간 외에는 아홉띠아르마딜로와 붉은다람쥐만이 자연 숙주이다.[3] 아르마딜로는 인간에게 동물원성 나병의 원인으로 지목되어 왔다.[11] 실험실에서는 쥐를 감염시킬 수 있으며, 이는 유용한 동물 모델이다.[12]
4. 대사
나균 유전체가 겪은 환원 진화는 다른 마이코박테리움에 비해, 특히 이화 작용 경로에서 대사 능력을 저하시켰다.[18]
==== 이화 작용 ====
나균은 무균 배지에서 배양할 수 없으며, 이는 숙주로부터의 영양분과 중간체에 의존한다는 것을 나타낸다.[19] 다른 나균속에 존재하는 많은 이화 경로가 손상되었는데, 이는 영양분 분해에 중요한 역할을 하는 효소가 없기 때문이다.[19] 나균은 중심 에너지 대사 경로에서 아세테이트 및 갈락토스와 같은 일반적인 탄소원을 사용할 수 있는 능력을 잃었다.[26] 또한 지질 분해가 손상되어 주요 리파아제 효소 및 지방 분해에 관여하는 다른 단백질에 결함이 있다.[20] 해당 종에는 해당 경로, 오탄당 인산 경로, TCA 회로와 같은 기능적 탄소 이화 경로가 계속 존재한다.[19] 이러한 결핍은 미생물의 성장을 숙주 유래 중간체와 같은 제한된 수의 탄소원으로 광범위하게 제한한다.[26]
==== 동화 작용 ====
나균( ''Mycobacterium leprae'')의 동화 대사 경로는 환원 진화에 의해 크게 영향을 받지 않았다.[18] 이 종은 퓨린, 피리미딘, 뉴클레오티드, 뉴클레오사이드와 같은 유전 물질과 메티오닌, 라이신을 제외한 모든 아미노산의 합성에 대한 능력을 유지한다.[19]
4. 1. 이화 작용
나균은 무균 배지에서 배양할 수 없으며, 이는 숙주로부터의 영양분과 중간체에 의존한다는 것을 나타낸다.[19] 다른 나균속에 존재하는 많은 이화 경로가 손상되었는데, 이는 영양분 분해에 중요한 역할을 하는 효소가 없기 때문이다.[19] 나균은 중심 에너지 대사 경로에서 아세테이트 및 갈락토스와 같은 일반적인 탄소원을 사용할 수 있는 능력을 잃었다.[26] 또한 지질 분해가 손상되어 주요 리파아제 효소 및 지방 분해에 관여하는 다른 단백질에 결함이 있다.[20] 해당 종에는 해당 경로, 오탄당 인산 경로, TCA 회로와 같은 기능적 탄소 이화 경로가 계속 존재한다.[19] 이러한 결핍은 미생물의 성장을 숙주 유래 중간체와 같은 제한된 수의 탄소원으로 광범위하게 제한한다.[26]4. 2. 동화 작용
나균(''Mycobacterium leprae'')의 동화 대사 경로는 환원 진화에 의해 크게 영향을 받지 않았다.[18] 이 종은 퓨린, 피리미딘, 뉴클레오티드, 뉴클레오사이드와 같은 유전 물질과 메티오닌, 라이신을 제외한 모든 아미노산의 합성에 대한 능력을 유지한다.[19]5. 유전체
2001년에 나균의 첫 번째 유전체 염기 서열이 완료되었으며, 1604개의 단백질 코딩 유전자와 1,116개의 유사유전자가 밝혀졌다.[24] 2013년에는 인도 타밀나두에서 처음 분리되어 ''TN''으로 지정된 균주의 유전체 염기 서열이 완료되었다. 이 유전체 염기 서열은 3,268,203 염기쌍(bp)을 포함하며 평균 GC-함량은 57.8%인데, 이는 4,441,529 bp와 65.6%의 G+C를 갖는 ''결핵균''보다 훨씬 적다.[21]
결핵균(''Mycobacterium tuberculosis'')의 염기 서열과 비교하면 환원 진화의 극단적인 사례가 나타난다. 유전체의 절반도 안 되는 부분이 기능적인 유전자를 포함하고 있다. 결핵균속(''Mycobacterium leprae'') 유전체에서 약 2000개의 유전자가 소실된 것으로 추정된다.[24] 유전자 결실과 쇠퇴로 인해 사철소 생산, 산화 호흡의 일부, 대부분의 미호기성 및 혐기성 호흡 호흡 사슬, 수많은 이화 작용 시스템 및 조절 회로를 포함한 많은 중요한 대사 활동이 제거된 것으로 보인다.[22] 이러한 환원 진화는 이 유기체가 절대적 세포 내 세균으로 발달한 것과 크게 관련이 있다.[23]
5. 1. 유사유전자
나균(''Mycobacterium leprae'')은 숙주 유기체에 의존하기 때문에 많은 DNA 복구 기능이 손실되어 삭제 돌연변이 발생이 증가했다.[23] ''나균''은 많은 유전자를 잃으면서 게놈 크기가 극적으로 축소되었는데,[26] 병원균 게놈의 절반 이상이 환원 진화 과정을 거쳐 유사유전자로 구성되어 있다.[26] 공개된 게놈 중에서 ''나균''은 가장 많은 유사유전자(>1000개)를 가지고 있다.[28]일부 ''나균'' 유사유전자 발현 수준은 대식세포 감염 시 변경될 수 있으며, 이는 감염, 세포 내 복제 및 복제 기능도 할 수 있음을 시사한다.[28] 이러한 게놈 축소는 완료되지 않았다.[29] ''결핵균(M. tuberculosis)''과 비교하면 4.42 Mbp 게놈에서 3.27 Mbp 게놈으로 축소되면서 약 1200개의 단백질 암호화 서열이 손실된다.
나균과 결핵균의 공통 조상의 게놈에 존재했던 많은 유전자들이 나균 게놈에서 손실되었다.[24][29]
5. 2. 필수 효소
나균에 필수적인 효소는 8가지가 있으며, 그 중 하나는 알라닌 라세마아제(alr)이다.[30] 이 효소는 D-알라닌-D-알라닌 연결효소와 알라닌/아스파르트산 대사에 관여하기 때문에 중요하다. 다른 필수 효소로는 추정 dTDP4데히드로람노스 3, 5 에피머라아제 (rm1C)가 있으며, 이는 뉴클레오티드 당 대사와 폴리케타이드 당 단위 생합성 모두에서 중요한 역할을 한다. 펩티도글리칸 생합성에 필요한 murG, murF, MurE, murY, murC, murD는 나균에 필수적인 나머지 6가지 효소이다.[30]6. 분포
나균은 전 세계적으로 분포하지만, 사하라 이남 아프리카, 아시아, 남아메리카에서 유병률이 가장 높다.[31] Mycobacterium leprae영어의 지리적 발생 지역으로는 앙골라, 브라질, 중앙아프리카 공화국, 콩고 민주 공화국, 미크로네시아 연방, 인도, 키리바시, 마다가스카르, 네팔, 마셜 제도, 탄자니아 등이 있다.[32]
1980년대에 다제요법(MDT)이 도입된 이후 나병 환자 발생률은 95% 감소했다.[33] 이러한 감소로 인해 세계보건기구(WHO)는 나병을 공중 보건 문제에서 퇴치되었다고 선언했는데, 이는 인구 10,000명당 나병 환자가 1명 미만인 경우를 정의한다.[34] 감염된 인간으로부터의 Mycobacterium leprae영어 전파 외에도 환경적 요인 또한 중요한 저장소가 될 수 있다. Mycobacterium leprae영어 DNA는 방글라데시의 나병 환자 집, 수리남의 아르마딜로 구멍, 그리고 영국 제도 내 나종성 붉은 다람쥐의 서식지 토양에서 검출되었다.[35] 미국에서 아르마딜로와 접촉한 이력이 있는 나병 사례가 다수 보고되었으며,[33] 아르마딜로 노출을 통한 인수 공통 전파 경로가 제안되었다. 이전 연구에서 미국 남동부의 환자들은 아르마딜로 관련 Mycobacterium leprae영어 유전자형과 동일한 균에 감염된 것으로 나타났다.[36] 브라질 파라 주에서는 아르마딜로에서 높은 비율의 Mycobacterium leprae영어 감염이 관찰되었으며, 아르마딜로 고기를 자주 섭취하는 사람들은 섭취하지 않거나 덜 섭취하는 사람들에 비해 M. leprae 특이 항원인 페놀성 글리콜리피드 I(PGL-I)의 역가가 유의하게 더 높았다.[37][33]
7. 진화
''나균''(Mycobacterium leprae)의 가장 가까운 친척은 ''나균증균''(Mycobacterium lepromatosis)이다.[38] 현존하는 ''나균'' 균주의 최근 공통 조상은 3,607년 전 (95% 최고 사후 밀도 2204–5525년 전)에 살았던 것으로 계산되었다.[38] 나병은 동아프리카에서 기원하여 처음에는 유럽과 중동으로 퍼진 후 지난 500년 동안 서아프리카와 아메리카로 확산되었다.[39]
DNA 포획 기술과 고처리량 시퀀싱을 사용하여 유럽의 지리적 기원이 다른 나병 의심 골병변이 있는 중세 시대 골격에서 거의 완전한 ''나균'' 서열을 얻었다. 고대 서열을 다양한 유전자형과 지리적 기원을 나타내는 나병 환자의 생검에서 얻은 현대 균주와 비교하였다.[40] Verena J. Schuenemann 외 연구진은 지난 1000년 동안 놀라운 게놈 보존과 현대 균주와 고대 균주 사이의 근접한 유사성을 입증했으며, 이는 유럽에서 나병의 갑작스러운 감소가 독성 손실 때문이 아니라 다른 전염병, 숙주 면역의 변화 또는 개선된 사회적 조건과 같은 외부 요인 때문임을 시사한다.[40]
8. 병인
나균(''Mycobacterium leprae'')의 잠복기는 9개월에서 20년까지 다양하다.[41] 이 균은 조직구와 신경 세포 내에서 세포 내적으로 복제되며 결핵성 및 나종성 두 가지 형태를 갖는다.[42] 결핵성 나병은 세포 매개 반응을 유도하여 피부에 융기되고 붉게 솟아오른 가장자리가 있는 크고 평평한 반점을 유발하며, 이는 건조하고 창백하며 털이 없는 중심부와 피부 감각 상실을 동반한다.[44] 나종성 나병은 미생물이 침입 부위의 대식세포 내에서 증식하며, 얼굴과 귓불의 상피 조직 내에서도 증식한다.[42] 억제 T 세포는 많지만, 상피양 세포와 거대 세포는 드물거나 없다. 세포 매개 면역이 손상되면 많은 수의 나균이 대식세포에 나타나고 감염된 환자는 피부가 접힌 채로 침입 부위에 구진이 생기며, 피부 신경의 점진적인 파괴는 "전형적인 사자 얼굴" 현상으로 이어진다.[44]
9. 증상
나병의 증상은 창백한 색의 피부 궤양, 수 주 또는 수 개월이 지나도 사라지지 않는 혹 또는 덩어리, 신경 손상으로 인해 팔다리의 감각 능력에 합병증을 유발하고 근육 쇠약으로 이어질 수 있다.[45] 증상은 보통 노출 후 3~5년 후에 나타나지만, 일부는 20년이 지나서야 증상을 보이기도 한다.[45] 이러한 긴 잠복기는 나병 감염 시점을 정확하게 진단하기 어렵게 만든다.[45]
아르마딜로에서 ''나균''은 조직과 신경에 유사한 구조적 및 병리학적 변화를 동반한 파종성 감염을 유발한다.[46] 다람쥐의 경우, 주둥이, 입술, 눈꺼풀, 귀, 생식기, 때로는 발과 다리 아래 부위에 심한 부종과 털이 빠지는 증상이 나타나며, 벼룩, 진드기 같은 외부 기생충 감염이 심해진다.[47]
10. 치료
1984년 세계보건기구(WHO)는 다제요법(MDT)을 권장했으며, 이는 나병 치료의 표준이 되었다.[49] MDT는 1995년부터 풍토병 발생 국가에 WHO에 의해 무상으로 공급되어 왔다.[49] 단일 약물(단일 요법)로 나병을 치료하면 약물 내성이 생길 수 있기 때문에 MDT가 사용된다.[49] MDT에 사용되는 약물 조합은 질병의 분류에 따라 달라진다.[49] WHO는 다균형 나병 환자에게 리팜피신, 클로파지민 및 답손의 조합을 12개월 동안 사용할 것을 권장한다.[49] WHO는 핍균성 나병 환자에게 리팜피신과 답손의 조합을 6개월 동안 사용할 것을 권장한다.[49] ''나균''은 약물 내성이 생길 수 있으므로 치료가 완료될 때까지 항생제를 정기적으로 복용해야 한다.[50] 치료의 효과는 환자에게 여전히 존재하는 균의 수를 추정하기 위해 피부 도말 표본에서 ''나균''의 산성 염색을 사용하여 확인할 수 있다.[51]
매년 보고되는 나병 발병 건수는 약 25만 건으로, MDT의 사용으로 나병 유병률이 90% 감소했음에도 불구하고 전염 고리가 아직 끊어지지 않았음을 나타낸다.[26] 이는 질병 관리가 아직 필요한 수준에 도달하지 못했음을 분명하게 보여주며, 치료 및 관리를 위한 지속적인 연구의 필요성을 강조한다.[26]
''나균''의 예방 조치는 치료받지 않은 감염된 사람과의 긴밀한 접촉을 피하는 것이다.[52] 실명, 손과 발의 불구, 마비는 모두 치료받지 않은 ''나균''과 관련된 신경 손상의 영향이다.[50] 치료는 이미 발생한 신경 손상을 되돌릴 수 없으므로 조기 치료가 필요하다.[50] 칼메트-게랭 간균 백신은 주요 목표인 결핵 외에도 나병에 대한 다양한 수준의 보호를 제공한다.[53]
10. 1. 항생제 표적
답손은 효소 디히드로프테로에이트 생성효소(DHPS)를 경쟁적으로 억제하여, 핵산 생합성의 필수 성분인 테트라히드로폴레이트의 생성을 감소시킨다.[54] 리팜핀은 DNA 의존성 RNA 중합효소의 β-소단위 결합을 방해하여 mRNA 생산을 분리하고 세포 사멸을 유발한다.[54] 클로파지민의 작용 기전은 완전히 밝혀지지 않았지만, 마이코박테리아 DNA에 결합하는 것으로 보인다.[54] 클로파지민이 마이코박테리아 DNA에 결합하는 것은 쥐에서 ''Mycobacterium leprae''에 대해 약하게 살균 작용을 하는 것으로 입증되었으며, 이것이 나병에 대한 단일 약물 요법에 적합하지 않은 이유이다.[54] 세 가지 주요 약물 중 리팜핀이 답손이나 클로파지민보다 더 강력한 살균력을 가진다.[54]10. 2. 잠재적 항생제 표적
항생제 내성이 증가함에 따라 새로운 항생제 표적을 찾는 것이 중요하다.[55] 나균(Mycobacterium leprae)의 펩티도글리칸 생합성에 필수적인 murC, murD, murE, murF, murG, murY의 6가지 효소는 항생제의 잠재적인 표적이다.[55] 이러한 효소들은 짧은 폴리펩타이드 사슬의 첨가를 촉매하여 세균 세포벽 합성을 돕기 때문에, 이 효소들을 표적으로 삼으면 세균 세포벽 합성을 방지할 수 있다.[55]10. 3. 항생제 내성
항생제 내성은 나균 신규 환자의 약 10%와 재발 환자의 약 15%에서 나타난다.[56]나균의 약제 내성은 항생제 표적 유전자의 변형과 세포벽 투과성 감소로 인해 발생하는 것으로 여겨진다.[57] 유출 펌프의 양과 비교했을 때, 나균은 결핵균(''M. tuberculosis'')보다 유출 펌프를 약 절반 정도 가지고 있다. 결핵균에서 약제 내성과 독성에 기여하는 유출 펌프는 나균이 겪은 게놈 축소 진화 과정에서도 보존되었다.[57]
11. 발견
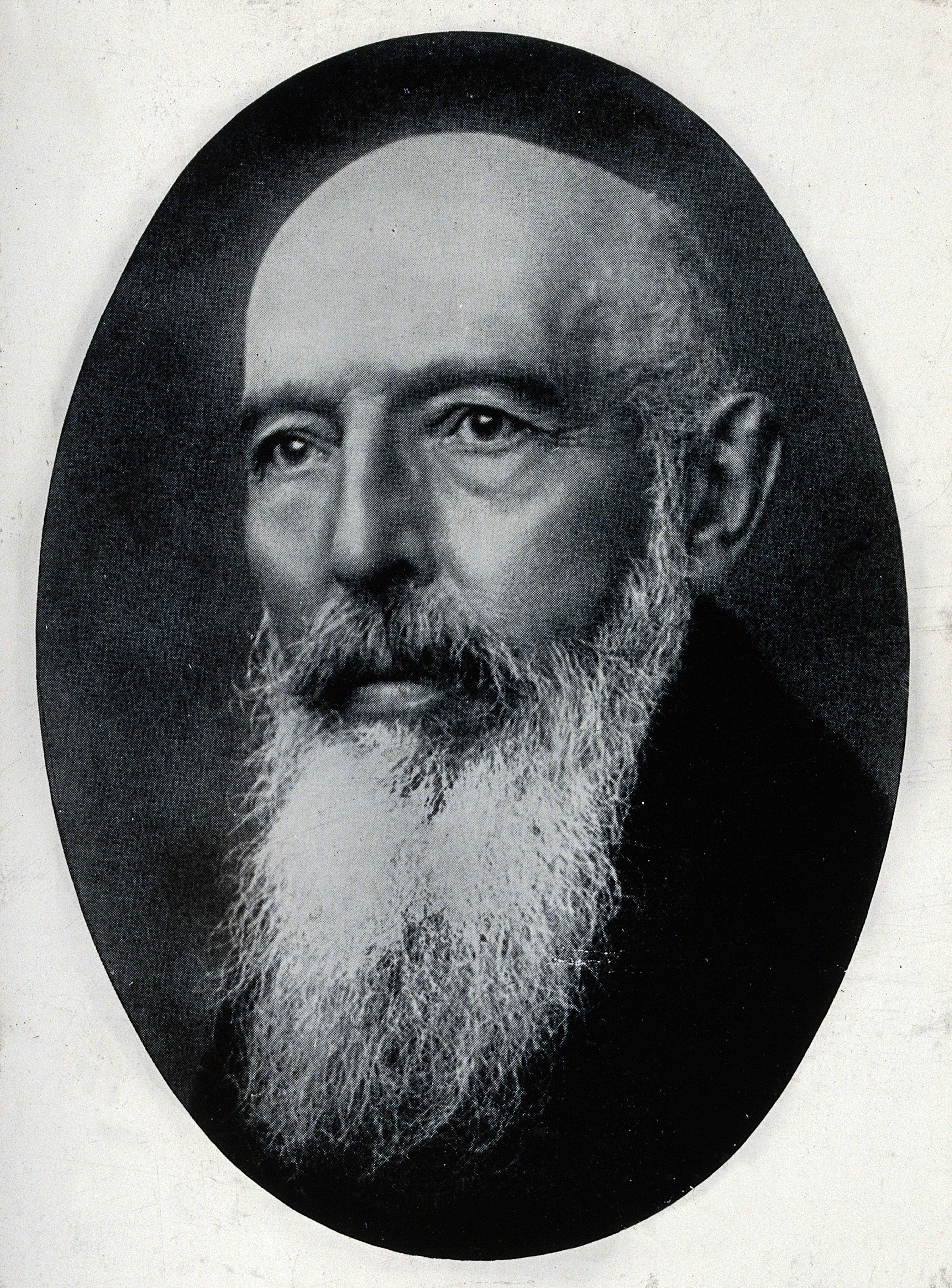
''나균''(''Mycobacterium leprae'')은 1873년 노르웨이 의사 게르하르트 아르마우어 한센에 의해 발견되었으며, 인간에게 질병을 일으키는 원인으로 확인된 최초의 세균이었다.[58] 이는 알베르트 루드비히 지게스문트 네이서에 의해 세균임이 확인되었으며, 네이서는 한센과 발견 우선순위를 두고 논쟁을 벌였다.[59] 한센은 세균을 동물에게 감염시키려는 시도는 실패했다. 1879년, 그는 동의 없이 나종 나병 환자의 조직을 결핵형 감염의 경미한 형태인 33세의 카리 닐스다터의 눈에 주사하여, 베르겐의 나병 병원에서 해고되고 의료 행위가 금지되었다.[60] 이 사건은 한센의 전문적인 명성에 거의 영향을 미치지 않았고, 그는 연구를 계속했다.[61]
참조
[1]
논문
Neuropathic ulcers in leprosy: clinical features, diagnosis and treatment
2022-06
[2]
웹사이트
Mycobacterium Leprae, the Cause of Leprosy
https://microbiology[...]
2014-08-27
[3]
논문
Differential growth of Mycobacterium leprae strains (SNP genotypes) in armadillos
2018-08
[4]
논문
Large-Scale Gene Expression Signatures Reveal a Microbicidal Pattern of Activation in Mycobacterium leprae-Infected Monocyte-Derived Macrophages With Low Multiplicity of Infection
2021
[5]
서적
Baron's Medical Microbiology
University of Texas Medical Branch
[6]
논문
Efficacy of fluorescent microscopy versus modified Fite-Faraco stain in skin biopsy specimens of leprosy cases - a comparative study
2022-05
[7]
논문
Leprosy: clinical and immunopathological characteristics
2022
[8]
웹사이트
Microbiology of M.leprae
https://www.who.int/[...]
[9]
서적
The Prokaryotes
https://zenodo.org/r[...]
Springer
[10]
서적
Infectious diseases
Saunders
1992
[11]
논문
Leprosy as a zoonosis: an update
1988
[12]
논문
Susceptibility and resistance in leprosy: Studies in the mouse model
2021-05
[13]
웹사이트
Mycobacterium leprae
https://www.scienced[...]
[14]
논문
Viable M. leprae as a research reagent
2001-03
[15]
논문
Mycobacterium leprae
[16]
문서
Lahiri R, Adams LB. (2016) "Cultivation and viability determination of ''Mycobacterium leprae''", Chapter 5.3. ''In'' Scollard DM, Gillis TP (ed), International textbook of leprosy, [https://internationaltextbookofleprosy.org].
[17]
논문
Zoonotic Leprosy in the Southeastern United States
2015-12
[18]
논문
Intracellular Mycobacterium leprae Utilizes Host Glucose as a Carbon Source in Schwann Cells
2019-12
[19]
논문
GSMN-ML- a genome scale metabolic network reconstruction of the obligate human pathogen Mycobacterium leprae
2020-07
[20]
논문
Massive gene decay in the leprosy bacillus
2001-02
[21]
논문
Whole-Genome Sequences of Four Clinical Isolates of Mycobacterium tuberculosis from Tamil Nadu, South India
2013-06
[22]
논문
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
1998-06
[23]
논문
Mycobacterium leprae's evolution and environmental adaptation
2019-09
[24]
논문
Massive gene decay in the leprosy bacillus
2001-02
[25]
논문
The genome of Mycobacterium leprae: a minimal mycobacterial gene set
2001-08-03
[26]
논문
Mycobacterium leprae: genes, pseudogenes and genetic diversity
2011-01
[27]
문서
Akama T, Suzuki K, Tanigawa K, et al. Whole-genome tiling array analysis of ''Mycobacterium leprae'' RNA reveals high expression of pseudogenes and noncoding regions. J Bacteriol. 2009;191(10):3321–3327. DOI:10.1128/JB.00120-09
[28]
논문
Detection of RNA expression from pseudogenes and non-coding genomic regions of Mycobacterium leprae
2009-09
[29]
논문
Mycobacterium leprae's evolution and environmental adaptation
2019-09
[30]
논문
Computational genome analyses of metabolic enzymes in Mycobacterium leprae for drug target identification
2010-03-31
[31]
논문
New Insights into the Geographic Distribution of Mycobacterium leprae SNP Genotypes Determined for Isolates from Leprosy Cases Diagnosed in Metropolitan France and French Territories
2015
[32]
웹사이트
'Risk of Exposure {{!}} Hansen''s Disease (Leprosy) {{!}} CDC'
https://www.cdc.gov/[...]
[33]
논문
Mycobacterium leprae transmission characteristics during the declining stages of leprosy incidence: A systematic review
2021-05
[34]
논문
The missing millions: a threat to the elimination of leprosy
2015-04
[35]
논문
Detection of Mycobacterium leprae DNA in soil: multiple needles in the haystack
2019-02
[36]
논문
Zoonotic Leprosy in the Southeastern United States
2015-12
[37]
논문
Evidence of zoonotic leprosy in Pará, Brazilian Amazon, and risks associated with human contact or consumption of armadillos
2018-06
[38]
논문
Insight into the evolution and origin of leprosy bacilli from the genome sequence of Mycobacterium lepromatosis
2015-04
[39]
논문
On the origin of leprosy
https://hal-pasteur.[...]
2019-07-14
[40]
논문
Genome-wide comparison of medieval and modern Mycobacterium leprae
2013-07
[41]
웹사이트
Leprosy (Hansen's disease) – Blue Book – Department of Health and Human services, Victoria, Australia
http://ideas.health.[...]
2015-11-17
[42]
논문
Cell Biology of Intracellular Adaptation of ''Mycobacterium leprae'' in the Peripheral Nervous System
2019-07
[43]
논문
Modulation of the Response to Mycobacterium leprae and Pathogenesis of Leprosy
2022
[44]
논문
Lucio's phenomenon: A systematic literature review of definition, clinical features, histopathogenesis and management
2022
[45]
웹사이트
Leprosy Overview
https://www.webmd.co[...]
2019-11-14
[46]
논문
Armadillos and leprosy: from infection to biological model
2019-09
[47]
간행물
Red squirrel leprosy, The University of Edinburgh
https://scottishsqui[...]
[48]
논문
Chemical probes for tagging mycobacterial lipids
2021-12
[49]
간행물
Guidelines for the diagnosis, treatment and prevention of leprosy.
World Health Organization
2018
[50]
웹사이트
Diagnosis and Treatment {{!}} Hansen's Disease (Leprosy) {{!}} CDC
https://www.cdc.gov/[...]
2019-11-12
[51]
간행물
Cultivation and viability determination of ''Mycobacterium leprae'', Chapter 5.3. ''In'' Scollard DM, Gillis TP (ed), International textbook of leprosy. www.internationaltextbookofleprosy.org.
International textbook of leprosy
2016-09-18
[52]
웹사이트
Leprosy: MedlinePlus Medical Encyclopedia
https://www.nlm.nih.[...]
2015-11-17
[53]
논문
Advances and hurdles on the way toward a leprosy vaccine
2011-11
[54]
간행물
Anti-leprosy drugs: Modes of action and mechanisms of resistance in ''Mycobacterium leprae'', Chapter 5.2. ''In'' Scollard DM, Gillis TP (ed), International textbook of leprosy. www.internationaltextbookofleprosy.org.
International textbook of leprosy
2019-01-25
[55]
논문
Computational genome analyses of metabolic enzymes in Mycobacterium leprae for drug target identification
http://www.bioinform[...]
2022-11-17
[56]
논문
Drug Resistance (Dapsone, Rifampicin, Ofloxacin) and Resistance-Related Gene Mutation Features in Leprosy Patients: A Systematic Review and Meta-Analysis
2022-10
[57]
논문
Insights on ''Mycobacterium leprae'' Efflux Pumps and Their Implications in Drug Resistance and Virulence
2018
[58]
서적
Undersøgelser Angående Spedalskhedens Årsager
https://wellcomelibr[...]
2021-04-09
[59]
논문
Gerhard Hansen Vs. Albert Neisser: Priority For the invention of Mycobacterium Leprae and problems of bioethics
2020-12
[60]
서적
Disease
Quercus
[61]
웹사이트
Armauer Hansen: The Controversy Surrounding his Unethical Human-to-Human Leprosy Transmission Experiment
https://worldneurolo[...]
2020-10-30
[62]
논문
Undersøgelser Angående Spedalskhedens Årsager (Investigations concerning the etiology of leprosy)
[63]
논문
Viable M. leprae as a research reagent
[64]
논문
Whole-Genome Sequences of Four Clinical Isolates of ''Mycobacterium tuberculosis'' from Tamil Nadu, South India
[65]
논문
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
[66]
서적
Sherris Medical Microbiology
https://archive.org/[...]
McGraw Hill
[67]
서적
Baron's Medical Microbiology
University of Texas Medical Branch
[68]
간행물
제33장 한센병
http://nih.go.kr/CDC[...]
국립보건연구원
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com