2025. 4. 17. 오전 10:03:00
'소 결핵균' 인체감염 국내 첫 확인…"인수감염 감시 서둘러야"
출처: SBS 뉴스 ( 한국 / 한국어 )
결핵균은 결핵을 일으키는 세균으로, 완전 호기성이며 세포벽의 지질 함량이 높아 특수한 염색법으로 확인된다. 사람의 기침이나 재채기를 통해 공기 감염되며, 대식세포 내에서 번식하여 면역력이 저하된 사람에게 심각한 증상을 유발한다. 결핵균은 다양한 유전자형을 가지며, 항생제 내성으로 인해 치료가 어려워지기도 한다. 진단은 가래 검사, 흉부 X선 촬영 등을 통해 이루어지며, 여러 항생제를 병용하는 화학 요법으로 치료한다. BCG 백신은 소아 결핵 예방에 효과적이나, 성인 폐결핵 예방 효과는 제한적이다. 결핵균은 아프리카에서 기원하여 전 세계로 확산된 것으로 추정된다.
| 결핵균 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Mycobacterium tuberculosis |
| 명명자 | Zopf 1883 |
| 동의어 | 결핵균 (Robert Koch 1882) |
| 과학적 분류 | |
| 도메인 | 세균 (Bacteria) |
| 문 | 방선균문 (Actinobacteria) |
| 강 | 방선균강 (Actinobacteria) |
| 목 | 방선균목 (Actinomycetales) |
| 아목 | 코리네박테리움아목 (Corynebacterineae) |
| 과 | 미코박테리움과 (Mycobacteriaceae) |
| 속 | 미코박테리움속 (Mycobacterium) |
| 종 | 결핵균 (M. tuberculosis) |
| 특징 | |
| 설명 | 결핵을 일으키는 병원성 세균 |
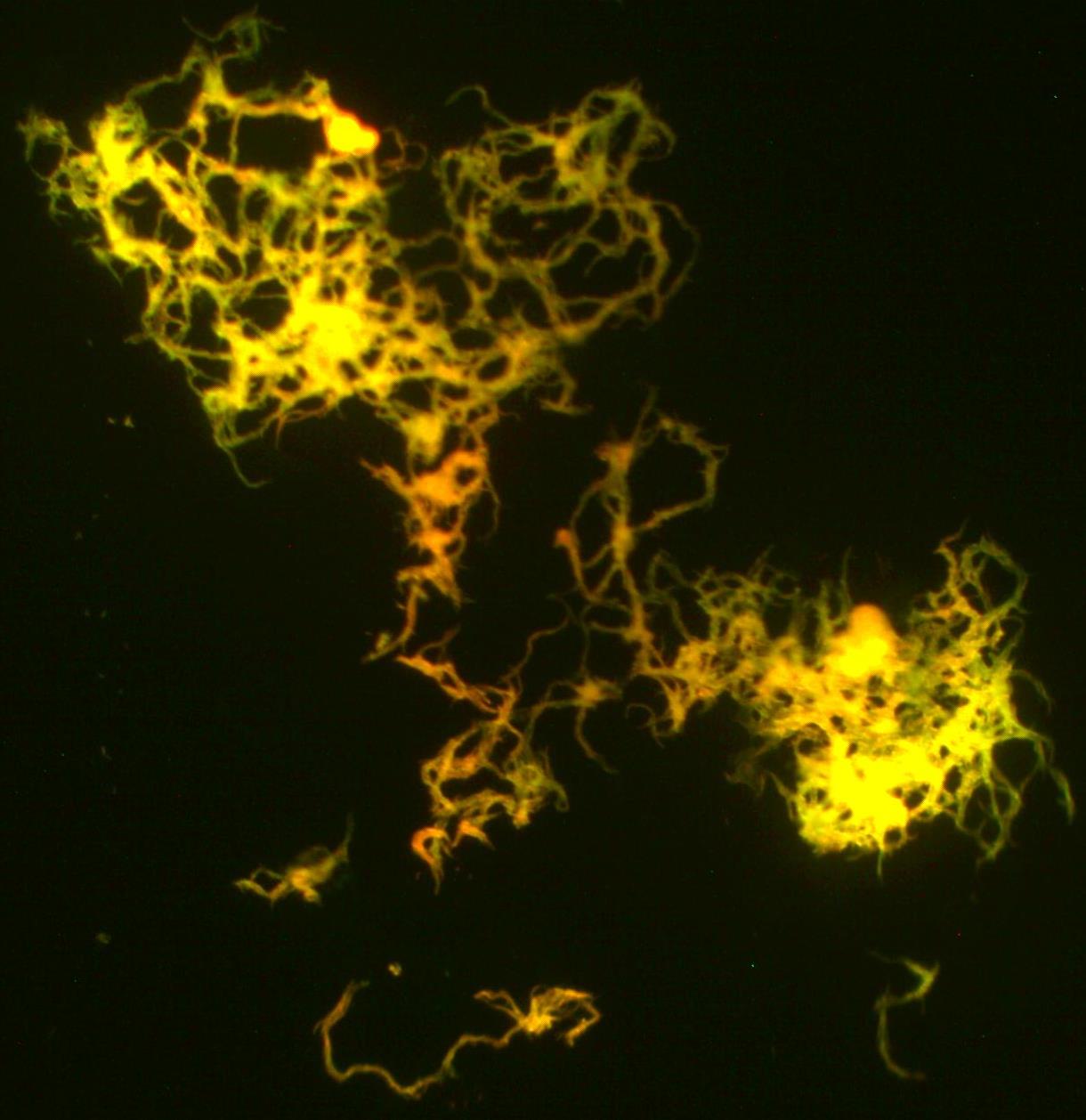
결핵균은 산소를 필요로 하는 절대 호기성 세균이며, 운동성이 없다.[7][8] 세포벽의 지방질 함량이 높아 질-닐슨 염색(항산성 염색)과 같은 특수한 염색 방법을 사용해야 한다. 결핵균은 15~20시간마다 분열하는데, 이는 분 단위로 분열하는 다른 세균들에 비해 매우 느린 속도이다.[9] (예: 대장균은 약 20분마다 분열).
결핵균은 작은 간균 형태이며, 약한 소독제에도 견딜 수 있고 건조한 상태에서도 몇 주 동안 생존할 수 있다. 미콜산과 코드 인자 글리콜리피드와 같은 지질이 풍부한 독특한 세포벽은 이러한 저항성과 발병력의 주요 원인으로, 주요 독성 인자로 작용한다.[9][10]
폐에서 결핵균은 폐포 대식세포에 의해 제거되지만, 세균은 소화되지 않는다. 세균의 세포벽은 코드 인자인 당지질로 만들어져 있으며, 파고솜과 항균 인자가 많은 리소좀의 융합을 억제한다.[18][10] 특히 결핵균은 결합 분자인 초기 엔도솜 자가 항원 1(EEA1)을 차단하는데, 이는 영양소가 채워진 소포의 융합은 막지 않아 결과적으로 세균의 증식이 억제되지 않는다. 또한, 디테르펜인 아이소투베르쿨로시놀의 생산은 파고솜의 성숙을 막고,[19] 반응성 질소 중간체를 중화하여 대식세포의 살해를 회피한다.[20] 최근에는 결핵균이 뉴클레오사이드인 1-투베르쿨로시닐아데노신(1-TbAd)을 분비하여 자신을 덮는데, 이는 제산제 역할을 하여 pH를 중화하고 리소좀의 부기를 유발한다.[21][22]
결핵균 감염에서 PPM1A 수치가 상향 조절되어, PPM1A가 내재적 및 외재적 세포자멸사 경로에 관여하기 때문에 병원체를 제거하기 위한 대식세포의 정상적인 세포자멸사 반응에 영향을 미친다. PPM1A 수치가 증가하면, PPM1A의 발현이 두 개의 세포자멸사 경로를 억제하여, [23] JNK/AP-1 신호 전달 경로를 통해 대식세포의 세포자멸사 경로가 제어된다.[23] 세포자멸사가 억제된 결과로, 결핵균은 안전한 복제 틈새를 제공받아 오랜 시간 동안 잠복 상태를 유지할 수 있다.[24]
면역 세포가 조직적으로 모인 육아종은 결핵 감염의 특징이다. 육아종은 면역 반응을 조절하고 조직 손상을 최소화하지만, 감염 확산에도 도움을 줄 수 있다.[25][26][27][28][29]
결핵균의 돌연변이를 구축하고 개별 유전자 산물의 특정 기능을 테스트하는 능력은 그 발병 기전과 독성 인자에 대한 이해를 크게 발전시켰다.[30] 예를 들어, 코드 인자(트레할로스 디미콜레이트)는 숙주 내 생존을 증가시키는 역할을 한다. 결핵균의 내성 균주는 유전자 돌연변이로 인해 하나 이상의 결핵 약물에 내성을 갖게 된다. 리팜피신 및 아이소니아지드와 같은 기존의 1차 결핵 약물은 세포 내 기생충인 결핵균을 제거하는 데 효율성이 감소했는데, 이는 대식세포 틈새로 효과적으로 침투하지 못하기 때문이다.[31]
JNK는 세포자멸사 경로(내재적 및 외재적) 제어에 중요한 역할을 하며, PPM1A 활성의 기질로도 발견되었다.[32] PPM1A 수치가 결핵균 감염 중에 증가하기 때문에, PPM1A 신호 전달 경로를 억제함으로써, 병원균 방어에서 정상적인 세포자멸사 기능을 복원하여 결핵균에 감염된 대식세포를 죽이는 치료법이 될 수 있다.[23]
결핵균에 감염된 대식세포의 세포자멸사를 복원하는 능력은 현재의 결핵 화학 요법 치료를 개선할 수 있으며, 결핵 약물이 틈새 내 박테리아에 더 잘 접근할 수 있도록 하여[34] 결핵균 감염의 치료 시간을 단축시킬 수 있다.
결핵균(H37Rv 균주)의 유전체는 1998년에 발표되었으며,[44][45] 크기는 4백만 염기쌍이고, 3,959개의 유전자를 가지고 있다. 이 중 40%는 기능이 밝혀졌고, 44%는 기능이 추정되며, 6개의 유사유전자가 있다.
유전체에는 지방산 대사에 관여하는 250개의 유전자가 포함되어 있으며, 이 중 39개는 왁스 코트를 생성하는 폴리케타이드 대사에 관여한다. 이는 병원체 생존에 왁스 코트가 중요하다는 것을 보여준다. 이후 연구에서 지방, 콜레스테롤과 같은 숙주 유래 지질로만 구성된 ''M. tuberculosis''의 지질 대사의 중요성이 입증되었다. 감염된 쥐의 폐에서 분리된 박테리아는 탄수화물보다 지방산을 선호하여 사용하며,[46] ''M. tuberculosis''는 콜레스테롤을 유일한 탄소원으로 성장할 수 있다. 콜레스테롤 사용 경로에 관여하는 유전자는 ''M. tuberculosis'' 감염, 특히 만성 감염 단계에서 중요하다.[47]
코딩 능력의 약 10%는 ''PE''/''PPE'' 유전자 계열이 차지하며, 이 유전자들은 산성, 글리신이 풍부한 단백질을 암호화한다. 이 단백질은 보존된 N-말단 모티프를 가지며, 이 모티프가 결실되면 대식세포 및 육아종에서 성장이 저해된다.[48]
''M. tuberculosis''에서는 9개의 비코딩 sRNA가 특징적으로 나타났으며,[49] 생물정보학 스크린에서 추가로 56개가 예측되었다.[50]
2013년 연구에서는 여러 감수성, 초강력 내성 및 다중 내성 ''M. tuberculosis'' 균주의 유전체를 연구하여 항생제 내성 메커니즘을 조사했다. 그 결과, 새로운 관계와 약물 내성 유전자가 밝혀졌으며, 일부 유전자 및 유전자 간 영역이 여러 약물에 대한 내성에 관여할 수 있음을 시사한다. 약물 내성의 원인으로 제안된 대부분의 유전자는 ''M. tuberculosis'' 발달에 필수적이며, 유전자 간 영역의 역할이 주목할 만하다.[51]
단일 분자 실시간 시퀀싱 및 생물정보학적 분석을 통해 ''M. tuberculosis''에서 3개의 DNA 메틸전달효소가 확인되었다. '''M'''ycobacterial '''A'''denine '''M'''ethyltransferases A (MamA),[52] B (MamB),[53] 및 C (MamC'')[54] ''세 가지 모두 아데닌 메틸전달효소이며, 각 효소는 일부 ''M. tuberculosis'' 임상 균주에서는 기능하지만 다른 균주에서는 기능하지 않는다.''[55][54] 일부 ''M. tuberculosis'' 균주는 MamA에 돌연변이를 가지고 있어 표적 아데닌 염기의 부분적인 메틸화를 유발하는데,[54] 이는 세포 내 확률적 메틸화로 발생한다.[54][57] MamA 돌연변이는 베이징 아계열 ''M. tuberculosis''에서 가장 흔하게 나타난다.[54]'' 메틸화가 유전체 내 일부 위치에서 유전자 발현에 영향을 미치기 때문에,[52] IMM은 표현형 다양성을 유발하고 베이징 아계열의 세계적인 성공에 부분적으로 기여할 수 있다는 가설이 있다.[54]
결핵의 초기 증상은 기침, 가래, 미열, 체중 감소, 식욕 부진 등이며, 다른 질환과 구별하기 어려울 수 있다.[119] 초기 폐결핵은 신열과 마른 기침, 흉부 X-선으로 알아낼 수 있는 이상을 초래할 수 있지만, 대부분의 경우 주 감염은 사람들이 극복할 수 있는 정도이다. 이러한 증상은 자연적으로 해결되기도 하지만, 절반 이상은 재발한다.
폐결핵은 결핵성 흉막염을 일으키기도 하는데, 흉통, 마른 기침, 열 등의 증상이 나타난다. 면역 체계가 약해진 경우, 결핵균 감염은 체내 다른 부분으로 전파될 수 있는데, 이를 속립성 결핵이라고 한다. 속립성 결핵 환자는 열, 체중 감소, 무력감, 식욕 감퇴 등의 증상을 보이며, 드물게 기침과 호흡 곤란을 일으킬 수 있다.
휴면기(비활동성) 결핵은 특정 기간이 지난 후 재발할 수 있으며, 종종 폐 상엽에 발생하고 증세가 심하여 점액이 많이 생기고, 기침에 피를 토하기도 한다.[117] 대부분 환자는 열, 식욕 부진, 이유 없는 체중 감소, 식은땀 등의 증상을 보인다.
감염이 체내 다른 부위로 옮겨가면 감염 부위에 따라 다른 증상이 나타날 수 있다. 복강 감염은 피로, 부어오름, 민감함, 맹장염과 같은 통증을 유발할 수 있다. 배뇨 시 고통은 방광 감염 징후일 수 있다. 어린이의 경우 결핵 감염이 뼈에 영향을 주어 가벼운 부어오름과 통증을 유발할 수 있다. 뇌 감염은 열, 두통, 메스꺼움, 졸림 등의 증상을 유발하며, 치료하지 않으면 혼수상태와 두뇌 손상이 발생할 수 있다.[119] 심막 감염은 열, 목정맥 확장, 호흡 가빠짐을 유발할 수 있다. 신장과 생식계통 감염은 신장 손상, 불임을 초래할 수 있다.
결핵균은 사람의 재채기, 기침 등으로 공기 중에 흩날려 공기 감염(비말핵 감염)을 일으킨다. 결핵균을 들이마셔도 면역이 잠시 균을 가두어 즉시 발병하지 않지만, 방치하면 기침이나 미열이 난다.
결핵의 종류에는 폐결핵, 결핵성 림프절염, 장결핵, 척추 결핵, 결핵성 수막염 등이 있다.
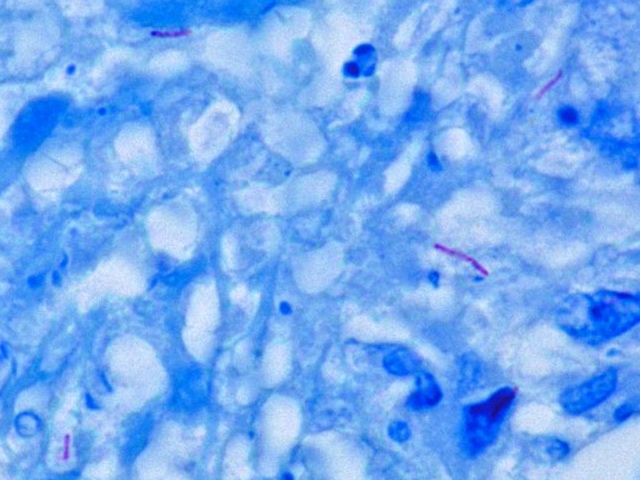
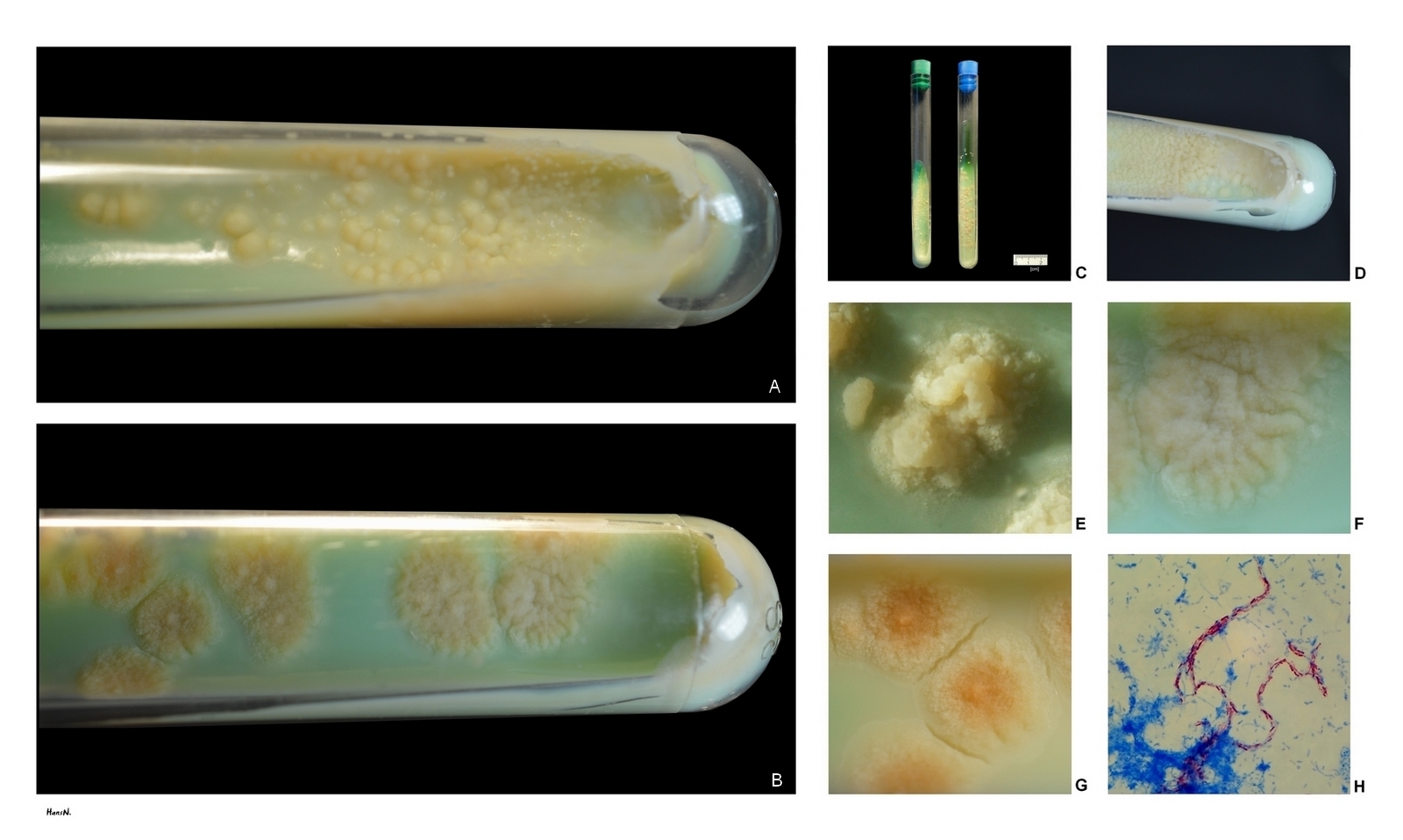
결핵 진단에는 가래 검사가 사용되는데, 균의 수가 적을 수 있으므로 3일 연속 아침에 가래를 채취한다. 표본은 3% 수산화칼륨(KOH)이나 수산화나트륨(NaOH)으로 처리한다. 질-닐슨 염색법을 사용하면 항산성균은 선홍색을 띠며 푸른 배경에서 선명하게 보여 '붉돔'으로 불리기도 한다.[120] 항산성균은 형광 현미경과 오라민-로다민 염색을 사용해 검사하면 금색으로 보인다. 결핵균은 전통적으로 Lowenstein-Jensen medium에서 배양되지만 6~8주 정도 걸려 느리다. 더 빠른 결과는 미들부룩 배지나 BACTEC을 사용하면 얻을 수 있다.
폐결핵의 경우 객담이 배출될 때 객담 도말 염색, 항산균 배양, DNA 중합효소 연쇄 반응(PCR), RNA 증폭(Direct TB 등)을 통해 균의 존재를 확인한다. 항산균 배양에서는 비결핵성 항산균과의 감별을 위해 니아신 시험이 시행된다. 국소 병변에는 각 장기에 맞는 영상 진단·조직 진단을 시행한다. 림프절, 장, 비장, 뼈(소위 카리에스) 등의 감염증도 확인한다. 폐에서는 흉부 X-ray, 흉부 CT, TBLB(경기관지 폐 생검), 폐 생검 등을 시행한다. 장결핵에서는 소화관 내시경, 위 투시, 바륨 관장 등을 시행하고, 림프절에서는 생검을, 카리에스에서는 MRI 등을 시행한다.
혈청을 이용한 검사법으로는 항TBGL 항체 검사가 있고, 말초혈 백혈구를 이용한 검사로는 T-SPOT, 퀀티페론 TB-2G 등이 있다. 투베르쿨린 반응 검사도 여전히 유용한 검사법 중 하나이다.
이러한 이유로 결핵 진단에는 수 시간에서 수 일이 소요된다.
결핵 치료는 통상 외래 환자를 기반으로 관리되며, 주로 약제로 구성된다. 치료는 보통 6~9개월 동안 지속되며, 처음 2개월 동안은 이소니아지드, 리팜핀, 피라진아마이드를 복용하고, 이후 4개월 동안은 약제 감수성 결과에 따라 이소니아지드와 리팜핀, 에탐부톨 또는 스트렙토마이신을 복용한다.[123] 약제 치료 계획은 연구소의 결과에 따라 달라질 수 있다.
증상이 없는 잠복 결핵 감염자의 경우, 항생제 치료는 결핵 발병을 예방하는 데 도움을 준다. 일반적으로 이소니아지드(INH)를 12개월 동안 복용하여 결핵 발병을 예방한다.[124] 그러나 이 약은 임신 중이거나, 간 질환, 알콜중독 등이 있는 경우에는 복용해서는 안 된다. 또한, 여러 부작용이 보고되었으며, 일부는 생명을 위협할 수도 있다. 약제 부작용 중 하나는 말초신경병증으로, 감각이 극도로 둔해지는 증상이다. 이는 이소니아지드와 함께 비타민 B6을 복용하면 예방하거나 완화할 수 있다.
활동성 결핵 환자는 여러 약제를 함께 사용하여 치료한다. 주요 항생제로는 이소니아지드, 리팜피신, 에탐부톨, 피라진아마이드가 사용된다.
주사로 투여하는 스트렙토마이신 또한 사용될 수 있는데, 특히 병세가 심하거나 환자가 약을 제대로 복용하지 못하는 경우에 사용된다.[124]
일반적으로 치료는 수개월 동안 지속되지만, 경우에 따라서는 1년 이상 지속되기도 한다. 치료 성공률은 환자가 처방에 따라 약을 잘 복용하는지에 크게 좌우된다.
1882년 3월 24일, 로베르트 코흐는 결핵균을 처음 발견하였다. 이 발견으로 1905년 노벨 생리학·의학상을 수상했다. 결핵균은 '코흐의 간균'으로도 알려져 있다.[125]
결핵균(''M. tuberculosis'')은 산소를 필요로 하는 세균으로, 운동성이 없다.[7][8] 다른 세균들이 보통 몇 분 단위로 분열하는 것과 달리, 결핵균은 18~24시간마다 분열하여 매우 느리게 증식한다. (대장균은 약 20분마다 분열) 이 세균은 작은 간균 형태이며, 약한 소독제에도 견딜 수 있고, 건조한 상태에서도 몇 주 동안 생존할 수 있다. 미콜산과 코드 인자 글리콜리피드와 같이 지질이 풍부한 독특한 세포벽은 건조에 대한 저항성을 부여하며, 주요 독성 인자로 작용한다.[9][10]
결핵균은 폐에서 폐포 대식세포에 의해 제거되지만 소화되지 않는다. 이는 세균의 세포벽이 식포와 리소좀의 융합을 막기 때문이다. 특히 결핵균은 초기 엔도솜 자기항원 1(early endosomal autoantigen 1, EE1)이라는 결합 분자를 차단하는데, 이로 인해 영양소가 풍부한 소낭은 결합하지만 세균은 소화되지 않아 대식세포 내에서 증식이 억제되지 않는다. 또한 'UreC' 유전자를 통해 식포의 산성화를 막는다.[112]
결핵균의 돌연변이 구성 및 유전자산물 시험 기술은 결핵균의 발병과 독성 요인 이해를 크게 증진시켰다. 분비·배출되는 여러 단백질들이 병 발생에 중요한 역할을 하는 것으로 알려져 있다.[113] 1998년 H37Rv 병균의 유전자 게놈이 출판되었는데,[115] 3959개의 유전자와 4백만 개의 염기쌍으로 구성되어 있으며, 이 중 40%는 역할이 밝혀졌고 44%는 역할이 추측되고 있다. 6개는 거짓 유전자이다.
250개의 유전자는 지방산 대사와 관련되어 있고, 39개는 밀랍같은 외부막을 생산하는 폴리케티드의 대사와 관련되어 있다. 이는 외부막이 병원체의 생존에 있어 유전적으로 중요함을 보여준다. 코딩 용량의 10%는 산성 글리신이 풍부한 단백질을 부호화하는 두 무리의 유전자계가 차지하며, 이 단백질들은 보존된 N말단 모티프를 가지고 있고, 삭제 시 대식세포와 육아종에서의 성장이 어려워진다.[116]
결핵균 분석을 위한 면역크로마토그래픽 혈청학적 분석법도 개발되었다.[122]


결핵균은 미코박테리움과에 속하는 미코박테리움 속 세균으로, 다른 미코박테리움 속 세균과 함께 항산균이라고 불리는 세균군 중 하나이다. 아포, 편모, 협막을 가지지 않으며, 크기는 2-4 x 0.3um인 호기성 간균이다.
세포벽에 미콜산이라고 불리는 지질을 다량 함유하고 있어, 일반적인 그람 염색으로는 염색이 잘 되지 않고 결과가 안정적이지 않아 그람 불균정으로 불리기도 한다. 하지만, 세포벽의 구조와 가온 그람 염색법으로 인해 그람 양성균으로 분류하는 것이 일반적이다. 한편, 매염제를 첨가하여 가온하면서 염색하는 질-넬슨 염색 등의 강력한 방법을 사용하면 염색이 가능할 뿐만 아니라, 일단 염색된 색소액이 탈색되기 어려운 특징을 가지며, 강한 탈색제인 염산 알코올에도 탈색 저항성을 보인다. 이 염색법을 '''항산성 염색'''이라고 부르며, 본법으로 염색되는 미코박테리움 속은 '''항산균''' (acid-fast bacteria, 이 경우 fast는 "퇴색하지 않는", "고정된"의 의미)이라고도 불린다.
| 이름 | 설명 |
|---|---|
| 결핵균 | 사람형 결핵균 |
| 소결핵균 | 소형 결핵균 |
| 아프리카결핵균 | 마이코박테리움 아프리카넘 |
| M. microti | 쥐형 결핵균 |
결핵균군의 4종은 모두 늦은 발육 균군이며, 배지에서의 콜로니 형성에 많은 시간을 필요로 하기 때문에 동정이 어려웠다. 현재는 분자생물학적 방법을 사용하여 유전자 증폭에 의해 동정이 가능하게 되었다.
생화학적 성상 및 병원성의 점에서 4종 각각 차이점을 가진다. 이 중, 결핵균이 결핵의 원인균으로 사람에게 병원성을 나타내는 외에, 소결핵균과 아프리카결핵균이 드물게 사람에게 감염된다. ''M. microti''는 사람에 대한 병원성을 갖지 않는다. 또한 소결핵균을 장기간 계대 배양하여 약독화한 것이 BCG이며, 결핵 예방을 위한 백신으로 이용되고 있다.
결핵균의 항생제 내성은 항생제가 표적으로 하는 유전자의 돌연변이 축적이나 약물의 적정 변화로 인해 발생한다.[42] 전 세계적으로 항생제 내성은 증가하는 추세이며, 특히 다제내성 결핵(MDR-TB)과 광범위 약제내성 결핵(XDR-TB)은 심각한 문제로 대두되고 있다.
결핵균(''M. tuberculosis'')에 감염된 사람들 중 일부만이 질병으로 진행되는 것은 숙주-병원체 상호 작용에 유전적 요소가 있음을 시사한다.[84] 미코박테리아 질환에 대한 멘델 감수성으로 불리는 희귀 질환군은 미코박테리아 감염에 대한 감수성을 증가시키는 유전적 결함을 가진 일부 개인에게서 관찰된다.[84]
초기 사례 연구와 쌍둥이 연구는 유전적 요소가 결핵균에 대한 숙주의 감수성에 중요하다는 것을 보여주었다. 최근의 전체 유전체 연관성 연구(GWAS)는 11p13 및 18q11 위치를 포함한 세 개의 유전적 위험 유전자를 확인했다.[85][86]
결핵균(''M. tuberculosis'')은 세포 내 병원체로서 숙주가 생성하는 항균 독성 라디칼로 인해 다양한 DNA 손상을 입는다. 활성 산소종 및/또는 활성 질소종에 노출되면 산화, 퓨린 제거, 메틸화 및 탈아미노화와 같은 DNA 손상이 발생하여 단일 가닥 및 이중 가닥 절단(DSB)을 유발할 수 있다.[88]
DnaE2 중합효소는 여러 DNA 손상 물질뿐만 아니라 쥐 감염 중에도 결핵균에서 상향 조절된다.[87] 이 DNA 중합효소의 손실은 쥐의 결핵균 독성을 감소시킨다.[87] DnaE2는 오류가 발생하기 쉬운 DNA 복구 중합효소로, 감염 중 결핵균 생존에 기여하는 것으로 보인다.
DSB 복구에는 상동 재조합 복구(HR)와 비상동 말단 연결(NHEJ) 두 가지 주요 경로가 사용된다. 대식세포에 내부화된 결핵균은 이들 경로 중 하나에만 결함이 있어도 생존할 수 있지만, 두 경로 모두 결함이 있으면 생존력이 약화된다.[88] 이는 결핵균이 세포 내에서 활성 산소 및/또는 활성 질소종에 노출될 때 HR 또는 NHEJ에 의해 복구되는 DSB가 형성됨을 의미한다.[88] 그러나 DSB 복구 결핍은 동물 모델에서 결핵균의 독성을 손상시키지는 않는 것으로 나타났다.[89]
결핵균 복합체(MTBC)는 아프리카, 아마도 아프리카의 뿔에서 진화했다.[58][59] 결핵균 복합체는 ''M. tuberculosis'' 외에도, ''M. africanum'', ''M. bovis'', ''M. caprae'', ''M. microti'', ''M. mungi, M. orygis'', 그리고 ''M. pinnipedii'' 등 다양한 동물 종에 감염되는 여러 구성원을 가지고 있다.
결핵균 복합체의 주요 인간 감염 종은 일곱 개의 계통으로 분류된다. 계통 1은 동아프리카-인도 (EAI), 마닐라 균주군 등을 포함한다. 계통 2는 베이징 그룹, 계통 3은 중앙 아시아 (CAS) 균주, 계통 4는 가나와 하를럼 (H/T), 라틴 아메리카-지중해 (LAM) 및 X 균주를 포함한다. 5형과 6형은 ''M. africanum''에 해당하며 주로 서아프리카에서 많이 보인다. 일곱 번째 유형은 아프리카의 뿔에서 분리되었다.[58]
초기 연구에서는 결핵균이 인류와 함께 진화했으며, 최근 공통 조상은 40,000년에서 70,000년 전에 진화했다고 보았다.[63][61] 그러나, 1,000년 된 페루 미라에서 추출한 결핵균 복합체 구성원의 게놈 서열을 분석한 후속 연구는 결핵균 복합체의 공통 조상이 불과 6,000년 전으로 훨씬 더 최근이라는 다른 결론을 제시했다.[64][65] 35개국에서 온 3,000개 이상의 ''M. bovis'' 균주 분석 결과, 이 종은 아프리카에서 기원한 것으로 추정되었다.[66]
BCG 백신은 ''M. bovis''에서 파생되었으며, 소아 결핵과 중증 결핵에 효과적이지만, 오늘날 가장 흔한 형태의 질병인 성인 폐결핵 예방에는 성공률이 제한적이다.[93] 이 때문에 결핵 발병률이 높은 지역에서 주로 사용되며, 감염 위험이 낮아 미국에서는 권장되지 않는 백신이다. 미국에서 이 백신을 접종받으려면 ''M. tuberculosis'' 전문가와 상담 과정을 거쳐 특정 기준을 충족하는 사람에게만 접종된다.[94]
BCG 백신 투여는 "훈련된 면역(trained immunity)"을 유도하는 것으로 나타났으며, 이는 선천 면역 체계의 향상된 반응을 의미한다.[95][96] 적응 면역과 달리 훈련된 면역은 단핵구 및 대식세포와 같은 선천 면역 세포에서 오래 지속되는 변화를 수반하며, 이러한 세포는 감염에 더욱 민감하게 반응하게 된다. 이러한 변화는 히스톤 변형과 같은 후성 유전체 재프로그래밍을 통해 발생하며, 이는 염증성 사이토카인의 생산 증가로 이어진다.
연구에 따르면 BCG 백신 접종과 코로나19에 대한 더 나은 면역 반응 사이에 상관관계가 있을 수 있다.[97]
DNA 백신은 단독으로 또는 BCG와 함께 사용할 수 있다. DNA 백신은 결핵 치료에 사용하고 향후 치료 기간을 단축할 수 있는 잠재력이 충분하다.[98]
[1]
논문
Microbe Profile: Mycobacterium tuberculosis: Humanity's deadly microbial foe
2018-04-01
[2]
서적
Sherris Medical Microbiology : an Introduction to Infectious Diseases
McGraw-Hill
2004
[3]
논문
Is Mycobacterium tuberculosis a closer relative to Gram-positive or Gram-negative bacterial pathogens?
2002-01-01
[4]
논문
Diagnostics for pulmonary tuberculosis
2016-04-01
[5]
논문
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
1998-06-01
[6]
논문
Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv
2002-10-01
[7]
논문
Mycobacteria: bugs and bugbears (two steps forward and one step back)
1999-12-01
[8]
서적
Mandell, Douglas, and Bennett's principles and practice of infectious diseases
Elsevier Saunders
2015
[9]
서적
Medical Microbiology
Elsevier Mosby
[10]
논문
Multiple roles of cord factor in the pathogenesis of primary, secondary, and cavitary tuberculosis, including a revised description of the pathology of secondary disease
https://pubmed.ncbi.[...]
2022-12-16
[11]
웹사이트
Mycobacterium tuberculosis and Tuberculosis
http://textbookofbac[...]
2016-12-24
[12]
서적
Medical Microbiology
University of Texas Medical Branch at Galveston
2017-09-05
[13]
논문
Molecular detection and identification of mycobacterium tuberculosis complex and four clinically important nontuberculous mycobacterial species in smear-negative clinical samples by the genotype mycobacteria direct test
2011-08-01
[14]
논문
Identification of mycobacteria in solid-culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry
2011-05-01
[15]
논문
Structome analysis of virulent Mycobacterium tuberculosis, which survives with only 700 ribosomes per 0.1 fl of cytoplasm
2015-01-28
[16]
논문
Characterization of Mycobacterium orygis as M. tuberculosis complex subspecies
2012-04-01
[17]
웹사이트
How TB Spreads
https://www.cdc.gov/[...]
Center for Disease Control
2018-03-14
[18]
논문
Infection by Mycobacterium tuberculosis promotes human alveolar macrophage apoptosis
1997-01-01
[19]
논문
Edaxadiene: a new bioactive diterpene from Mycobacterium tuberculosis
2009-12-01
[20]
논문
Immune evasion by Mycobacterium tuberculosis: living with the enemy
2003-08-01
[21]
논문
Mycobacterium tuberculosis releases an antacid that remodels phagosomes
2019-09-01
[22]
논문
T(oo)bAd
2019-09-01
[23]
논문
Mycobacterium tuberculosis exploits the PPM1A signaling pathway to block host macrophage apoptosis
2017-02-01
[24]
논문
Alveolar macrophages in pulmonary host defence the unrecognized role of apoptosis as a mechanism of intracellular bacterial killing
2013-11-01
[25]
논문
Revisiting the role of the granuloma in tuberculosis
2012-04-01
[26]
논문
Inflammatory signaling in human tuberculosis granulomas is spatially organized
2016-05-01
[27]
논문
TGFβ restricts expansion, survival, and function of T cells within the tuberculous granuloma
2021-04-01
[28]
논문
The role of the granuloma in expansion and dissemination of early tuberculous infection
2009-01-01
[29]
논문
The Tuberculous Granuloma and Preexisting Immunity
2022-04-01
[30]
서적
Bacterial Secreted Proteins: Secretory Mechanisms and Role in Pathogenesis
Caister Academic Press
[31]
논문
A Macrophage Infection Model to Predict Drug Efficacy Against Mycobacterium Tuberculosis
2016-08-01
[32]
논문
Protein phosphatase 2Calpha inhibits the human stress-responsive p38 and JNK MAPK pathways
1998-08-01
[33]
논문
JNK signaling in apoptosis
2008-10-01
[34]
문서
The ability to restore macrophage apoptosis to M. tuberculosis infected ones could improve the current tuberculosis chemotherapy treatment, as TB drugs can gain better access to the bacteria in the niche (M),
[35]
웹사이트
Tuberculosis – Symptoms and causes
https://www.mayoclin[...]
2019-11-12
[36]
논문
DNA polymorphisms in strains of Mycobacterium tuberculosis analyzed by pulsed-field gel electrophoresis: a tool for epidemiology
1992-06
[37]
논문
Genetic diversity in the ''Mycobacterium tuberculosis'' complex based on variable numbers of tandem DNA repeats
1998-05
[38]
논문
High-resolution minisatellite-based typing as a portable approach to global analysis of Mycobacterium tuberculosis molecular epidemiology
2001-02
[39]
논문
Mycobacterial interspersed repetitive unit typing of Mycobacterium tuberculosis compared to IS6110-based restriction fragment length polymorphism analysis for investigation of apparently clustered cases of tuberculosis
2003-08
[40]
논문
Proposal for standardization of optimized mycobacterial interspersed repetitive unit-variable-number tandem repeat typing of Mycobacterium tuberculosis
2006-12
[41]
논문
Complications in the study of ancient tuberculosis: non-specificity of IS6110 PCRs
[42]
논문
Multidrug-resistant Mycobacterium tuberculosis: molecular perspectives
1998-06
[43]
웹사이트
Drug-resistant TB
https://www.cdc.gov/[...]
Center for Disease Control
2014-04
[44]
논문
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
1998-06
[45]
웹사이트
"''Mycobacterium tuberculosis''"
http://www.sanger.ac[...]
Sanger Institute
2007-03-29
[46]
논문
Biochemical differentiation of Mycobacterium tuberculosis grown in vivo and in vitro
1956-08
[47]
논문
Pathogen roid rage: cholesterol utilization by Mycobacterium tuberculosis
2014
[48]
논문
Microbial pathogenesis of Mycobacterium tuberculosis: dawn of a discipline
2001-02
[49]
논문
Identification of small RNAs in Mycobacterium tuberculosis
2009-08
[50]
논문
Identification of 17 Pseudomonas aeruginosa sRNAs and prediction of sRNA-encoding genes in 10 diverse pathogens using the bioinformatic tool sRNAPredict2
[51]
논문
Genome sequencing of 161 Mycobacterium tuberculosis isolates from China identifies genes and intergenic regions associated with drug resistance
2013-10
[52]
논문
DNA methylation impacts gene expression and ensures hypoxic survival of Mycobacterium tuberculosis
2013-07-04
[53]
논문
Precision methylome characterization of Mycobacterium tuberculosis complex (MTBC) using PacBio single-molecule real-time (SMRT) technology
2016-01
[54]
논문
Drivers and sites of diversity in the DNA adenine methylomes of 93 Mycobacterium tuberculosis complex clinical isolates
2020-10
[55]
논문
Methylation in Mycobacterium tuberculosis is lineage specific with associated mutations present globally
2018-01
[56]
논문
The Epigenomic Landscape of Prokaryotes
2016-02
[57]
논문
Single molecule-level detection and long read-based phasing of epigenetic variations in bacterial methylomes
2015-06
[58]
논문
Significance of the identification in the Horn of Africa of an exceptionally deep branching Mycobacterium tuberculosis clade
[59]
논문
Out-of-Africa migration and Neolithic coexpansion of Mycobacterium tuberculosis with modern humans
2013-10
[60]
논문
Progenitor "Mycobacterium canettii" clone responsible for lymph node tuberculosis epidemic, Djibouti
2014-01
[61]
논문
Genomic insights into tuberculosis
2014-05
[62]
논문
New Mycobacterium tuberculosis Complex Sublineage, Brazzaville, Congo
2017-03
[63]
논문
Origin, spread and demography of the Mycobacterium tuberculosis complex
2008-09
[64]
논문
Armed conflict and population displacement as drivers of the evolution and dispersal of Mycobacterium tuberculosis
2016-11
[65]
논문
Pre-Columbian mycobacterial genomes reveal seals as a source of New World human tuberculosis
2014-10
[66]
논문
An African origin for ''Mycobacterium bovis''
2020-01-31
[67]
논문
Variable host-pathogen compatibility in Mycobacterium tuberculosis
2006-02
[68]
논문
Stable association between strains of Mycobacterium tuberculosis and their human host populations
2004-04
[69]
논문
The role of selection in shaping diversity of natural M. tuberculosis populations
2013-08
[70]
논문
Pre-Columbian mycobacterial genomes reveal seals as a source of New World human tuberculosis
2014-10
[71]
논문
Eighteenth-century genomes show that mixed infections were common at time of peak tuberculosis in Europe
2015-04
[72]
논문
Mycobacterium tuberculosis lineage 4 shaped by colonial migration and local adaptation
2018-10
[73]
논문
Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis
2005-09
[74]
논문
Detection and molecular characterization of 9,000-year-old Mycobacterium tuberculosis from a Neolithic settlement in the Eastern Mediterranean
Public Library of Science (PLoS)
2008-10-15
[75]
논문
Antimicrobial Resistance in Mycobacterium tuberculosis: The Odd One Out
https://discovery.uc[...]
2022-12-23
[76]
서적
Global tuberculosis report 2020.
https://www.who.int/[...]
World Health Organization
2022-04-04
[77]
논문
Trends in burden of multidrug-resistant tuberculosis in countries, regions, and worldwide from 1990 to 2017: results from the Global Burden of Disease study
2021-03
[78]
논문
New insight in molecular detection of Mycobacterium tuberculosis
2024-06-21
[79]
논문
Mutations inside rifampicin-resistance determining region of rpoB gene associated with rifampicin-resistance in Mycobacterium tuberculosis
2018-09
[80]
논문
Drug Resistance Mechanisms in Mycobacterium tuberculosis
2014-07
[81]
논문
MmpL transporter-mediated export of cell-wall associated lipids and siderophores in mycobacteria
2016-09
[82]
논문
MmpL3 is the flippase for mycolic acids in mycobacteria
2017-07
[83]
논문
Cryo-EM structure and resistance landscape of M. tuberculosis MmpL3: An emergent therapeutic target
2021-10
[84]
논문
Mendelian susceptibility to mycobacterial disease: genetic, immunological, and clinical features of inborn errors of IFN-γ immunity
2014-12
[85]
논문
Common variants at 11p13 are associated with susceptibility to tuberculosis
2012-02
[86]
논문
Genome-wide association analyses identifies a susceptibility locus for tuberculosis on chromosome 18q11.2
2010-09
[87]
논문
DnaE2 polymerase contributes to in vivo survival and the emergence of drug resistance in Mycobacterium tuberculosis
2003-04
[88]
논문
Either non-homologous ends joining or homologous recombination is required to repair double-strand breaks in the genome of macrophage-internalized Mycobacterium tuberculosis
2014
[89]
논문
Deficiency of double-strand DNA break repair does not impair Mycobacterium tuberculosis virulence in multiple animal models of infection
2014-08
[90]
웹사이트
Robert Koch and Tuberculosis: Koch's Famous Lecture
http://nobelprize.or[...]
Nobel Foundation
2008-11-18
[91]
서적
Scientific American
https://books.google[...]
Munn & Company
2021-09-10
[92]
웹사이트
Tuberculosis History Timeline
http://www.mycobacte[...]
2010-06-18
[93]
논문
Early BCG vaccination is unrelated to pulmonary immunity against Mycobacterium tuberculosis in adults
2014-10
[94]
웹사이트
'Fact Sheets {{!}} Infection Control & Prevention {{!}} Fact Sheet – BCG Vaccine {{!}} TB'
https://www.cdc.gov/[...]
CDC
2019-11-12
[95]
논문
BCG-induced trained immunity: history, mechanisms and potential applications
2023-02-10
[96]
논문
Trained immunity: A program of innate immune memory in health and disease
2016-04-22
[97]
웹사이트
Tuberculosis vaccine drawing attention in fight against coronavirus
https://english.kyod[...]
2020-04-14
[98]
논문
DNA vaccine construct formation using Mycobacterium-specific gene Inh-A
2022
[99]
문서
코ッホの発見の日に因んで[[世界結核デー]]が制定された。
[100]
논문
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
1998-06
[101]
문서
“Janeways’s Immuno Biologoy” 5th edition by Kenneth Murphy, Fig. 16.22
[102]
문서
“Janeways’s Immuno Biologoy” 5th edition by Kenneth Murphy, p514
[103]
논문
Origin, spread and demography of the Mycobacterium tuberculosis complex
2008-09
[104]
논문
Genomic insights into tuberculosis
2014-05
[105]
논문
Armed conflict and population displacement as drivers of the evolution and dispersal of Mycobacterium tuberculosis
2016-11
[106]
논문
An African origin for ''Mycobacterium bovis''
2020-01-31
[107]
서적
Sherris Medical Microbiology
https://archive.org/[...]
McGraw Hill
[108]
문서
코흐는 이 발견으로 [[1905년]] [[노벨 생리학·의학상]]을 수상했다.
[109]
저널
Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence
[110]
저널
Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv
http://mic.sgmjourna[...]
[111]
서적
Medical Microbiology
Elsevier Mosby
[112]
저널
Vaccines: A souped-up version of BCG
[113]
서적
Bacterial Secreted Proteins: Secretory Mechanisms and Role in Pathogenesis
https://archive.org/[...]
Caister Academic Press
[114]
문서
Mahon, Manuselis, ''Textbook of Diagnostic Microbiology'', p.576
[115]
웹인용
Mycobacterium tuberculosis
http://www.sanger.ac[...]
Sanger Institute
2011-03-20
[116]
저널
Microbial pathogenesis of Mycobacterium tuberculosis: dawn of a discipline
http://linkinghub.el[...]
[117]
웹인용
Tuberculosis Symptoms
http://www.emedicine[...]
2011-04-12
[118]
웹인용
What are the symptoms of tuberculosis?
https://web.archive.[...]
2011-04-12
[119]
웹인용
Tuberculosis
http://www.merck.com[...]
2011-04-12
[120]
저널
Quarantining the noncompliant TB patient: catching the "Red Snapper"
[121]
서적
Brock Biology of Microorganisms
Prentice Hall
[122]
저널
An immunochromatographic serological assay for the diagnosis of Mycobacterium tuberculosis
[123]
웹인용
Tuberculosis Treatment
http://www.emedicine[...]
2011-05-18
[124]
웹인용
How is tuberculosis treated?
https://web.archive.[...]
2011-05-19
[125]
웹인용
Robert Koch and Tuberculosis: Koch's Famous Lecture
http://nobelprize.or[...]
Nobel Foundation
2011-05-19
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com