세포액
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
세포액은 1965년에 처음 사용된 용어로, 세포를 파괴하여 얻은 액체나 온전한 세포 내 세포질의 액체상을 지칭한다. 세포액은 물, 용해된 이온, 작은 분자, 단백질 등으로 구성되며, 세포 부피에서 세포질이 차지하는 비율은 생물종에 따라 다르다. 세포질은 세포 내 여러 과정이 일어나는 장소이며, 신호 전달, 세포 분열, 물질 수송 등의 기능을 수행한다. 원핵생물과 진핵생물 모두에서 대사 과정이 일어나며, 동물 세포에서는 단백질 생합성, 해당 과정 등 주요 대사 경로가 진행된다.
더 읽어볼만한 페이지
- 세포해부학 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포해부학 - 세포막
세포막은 세포질을 둘러싸고 세포 내외부 환경을 구분하는 선택적 투과성 막으로, 인지질 이중층과 단백질, 탄수화물로 구성되어 물질 수송, 세포 형태 유지, 세포 간 신호 전달, 세포 접합 등의 기능을 수행하며, 유동 모자이크 모델로 설명된다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.
| 세포액 | |
|---|---|
| 개요 | |
| 정의 | 세포 내에 존재하는 액체 성분 |
| 구성 요소 | 물, 이온, 작은 분자, 거대 분자 (예: 단백질) |
| 위치 | 세포막 내, 세포 소기관 외부 |
| 기능 | 세포 내 물질 운반, 생화학 반응 장소 제공 |
| 관련 용어 | 세포질 세포기질 |
| 물리화학적 특성 | |
| 조성 | 주로 물, 단백질, 이온, 작은 분자로 구성 |
| 농도 | 세포 종류와 상태에 따라 다름 |
| pH | 일반적으로 중성에 가까움 |
| 점성 | 세포 종류와 상태에 따라 다름 |
| 기능 | |
| 물질 운반 | 세포 내 물질 이동 통로 역할 |
| 대사 | 많은 대사 경로가 일어나는 장소 제공 |
| 세포 신호 전달 | 신호 전달 물질의 확산 및 반응 장소 제공 |
| 세포 형태 유지 | 세포골격과 상호작용하여 세포 형태 유지 |
| 효소 활성 조절 | 이온 농도 및 pH 변화를 통해 효소 활성 조절 |
| 관련 연구 | |
| 연구 분야 | 세포 생물학, 생화학, 분자 생물학 |
| 연구 방법 | 현미경 관찰 생화학적 분석 분자생물학적 기법 |
| 중요성 | 세포의 기본적인 기능 이해에 필수적 |
2. 정의
"세포액"이라는 용어는 1965년 H. A. 래디에 의해 처음 도입되었다. 초기에는 세포를 파괴하고 모든 불용성 성분을 초원심분리로 펠릿화하여 생성된 액체를 지칭했다.[3][15] 이러한 가용성 세포 추출물은 세포질의 가용성 부분과 동일하지 않으며, 일반적으로 세포질 분획이라고 불린다.[4]
현재 "세포액"이라는 용어는 온전한 세포 내 세포질의 액체상을 지칭하는 데 사용된다.[4] 여기에는 세포 소기관 내에 포함된 세포질의 모든 부분이 제외된다.[22] "세포액"이라는 단어가 세포 추출물과 온전한 세포 내 세포질의 가용성 부분을 모두 지칭하는 데 사용될 수 있어 혼동될 수 있기 때문에, "수성 세포질"이라는 구절은 살아있는 세포의 세포질의 액체 내용을 설명하는 데 사용되었다.[15]
이전에는 그 본질이 잘 이해되지 않았기 때문에(원형질 참조), 세포액에 대해 '''유리질'''[5]을 포함한 다른 용어들이 동의어는 아니지만 사용되었다.[4]
3. 성분
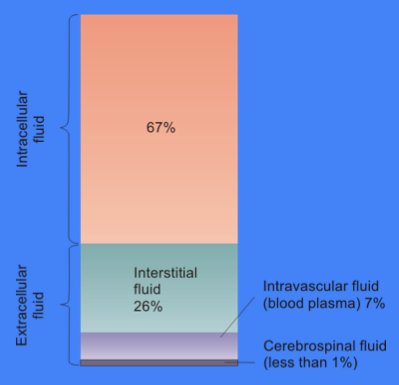
세포액은 대부분 물, 용해된 이온, 작은 분자 및 큰 수용성 분자(예: 단백질)로 구성된다. 비단백질 분자는 대부분 300 Da 미만의 분자량을 갖는다.[7] 이러한 작은 분자의 혼합물은 대사 물질의 다양성이 매우 커 복잡하다. 예를 들어, 식물에서는 최대 20만 개의 서로 다른 작은 분자가 만들어질 수 있지만, 모두가 동일한 종이나 단일 세포에 존재하지는 않는다.[8] 대장균이나 빵 효모와 같은 단일 세포 내 대사 물질 수는 1,000개 미만으로 예측된다.[9][10]
3. 1. 물
세포기질의 대부분은 물이며 일반 세포 전체 용적의 약 70%를 차지한다.[75] 세포기질의 pH는 7.4인 반면,[76] 인간 세포기질 pH는 7.0~7.4 사이이며 세포가 성장할수록 이 수치는 더 상승하는 것이 보통이다.[77]
세포질의 점성은 순수한 물과 거의 같지만, 이 액체를 통한 작은 분자의 확산은 순수한 물에서보다 약 4배 느린데, 이는 주로 세포질 내의 많은 고분자와의 충돌 때문이다.[14] 브라인 슈림프에 대한 연구에서 물이 세포 기능에 미치는 영향을 조사했는데, 세포 내 물의 양이 20% 감소하면 신진대사가 억제되고, 세포가 건조해짐에 따라 신진대사가 점진적으로 감소하며, 물의 수준이 정상보다 70% 낮아지면 모든 대사 활동이 중단되는 것을 확인했다.[15]
물은 생명에 필수적이지만, 세포질 내 물의 구조는 잘 이해되지 않고 있는데, 이는 주로 핵 자기 공명 분광법과 같은 방법이 물의 평균 구조에 대한 정보만 제공하고 미세한 규모의 국소적인 변화를 측정할 수 없기 때문이다. 순수한 물의 구조조차도 물이 수 클러스터와 같은 구조를 수소 결합을 통해 형성할 수 있기 때문에 제대로 이해되지 않고 있다.[19]
세포 내 물에 대한 고전적인 관점은 이 물의 약 5%가 용매화의 물로서 용질이나 고분자에 의해 강하게 결합되어 있으며, 대부분은 순수한 물과 동일한 구조를 가지고 있다는 것이다.[15] 이 용매화의 물은 삼투에 활성화되지 않으며 다른 용매 특성을 가질 수 있으므로, 일부 용해된 분자는 제외되고 다른 분자는 농축될 수 있다.[16][17] 그러나 다른 연구자들은 세포 내 고농도의 고분자의 영향이 세포질 전체에 걸쳐 나타나며, 세포 내 물이 희석된 용액의 물과는 매우 다르게 행동한다고 주장한다.[18] 이러한 아이디어에는 세포가 저밀도 및 고밀도 물 구역을 포함하고 있어 세포의 다른 부분의 구조와 기능에 광범위한 영향을 미칠 수 있다는 제안이 포함된다.[19][20] 그러나, 생체 내 물의 이동성을 직접 측정하기 위한 첨단 핵 자기 공명 방법의 사용은 이러한 아이디어와 모순되는데, 이는 세포 물의 85%가 순수한 물처럼 작용하는 반면, 나머지는 덜 이동하며 고분자에 결합될 가능성이 있음을 시사하기 때문이다.[21]
3. 2. 이온
포유류의 세포기질과 혈액의 일반적인 이온 농도는 다음과 같다.[78][22]
| 이온 | 세포기질 (밀리몰) | 혈액 (밀리몰) |
|---|---|---|
| 칼륨 | 139-150 | 4 |
| 나트륨 | 12 | 145 |
| 염화 이온 | 4 | 116 |
| 탄산수소염 | 12 | 29 |
| 단백질 내 아미노산 | 138 | 9 |
| 마그네슘 | 0.8 | 1.5 |
| 칼슘 | <0.0002 | 1.8 |
세포질 내 다른 이온의 농도는 세포외액의 농도와 상당히 다르다. 세포질은 세포 구조 외부보다 단백질 및 핵산과 같은 하전된 거대분자를 훨씬 더 많이 함유하고 있다.
세포외액과 대조적으로 세포질은 칼륨 이온 농도가 높고 나트륨 이온 농도가 낮다.[25] 이러한 이온 농도의 차이는 삼투압 조절에 매우 중요하다. 만약 이온 수치가 세포 내부와 외부에서 동일하다면 세포 내부의 거대분자 수치가 외부보다 높기 때문에 물이 삼투에 의해 지속적으로 유입될 것이다. 대신 나트륨 이온은 배출되고 칼륨 이온은 Na⁺/K⁺-ATPase에 의해 흡수되며, 칼륨 이온은 농도 기울기에 따라 칼륨 선택 이온 채널을 통해 흐른다. 이러한 양전하 손실은 음의 막 전위를 생성한다. 이러한 전위 차이의 균형을 맞추기 위해 음의 염화물 이온도 선택적 염화물 채널을 통해 세포를 빠져나간다. 나트륨 및 염화물 이온의 손실은 세포 내부의 유기 분자 농도가 높기 때문에 발생하는 삼투 효과를 보상한다.[25]
세포는 베타인이나 트레할로스와 같은 삼투압 보호제를 세포질에 축적함으로써 훨씬 더 큰 삼투압 변화에 대처할 수 있다.[25] 이러한 분자 중 일부는 세포가 완전히 건조되는 것을 견딜 수 있게 하고 유기체가 은생이라고 하는 정지된 생명 상태에 들어갈 수 있게 한다.[26] 이 상태에서 세포질과 삼투압 보호제는 단백질과 세포막을 건조로 인한 손상으로부터 보호하는 유리질 고체가 된다.[27]
세포질 내 칼슘 농도가 낮기 때문에 칼슘 이온은 칼슘 신호 전달에서 2차 전달자로 기능할 수 있다. 호르몬이나 활동 전위와 같은 신호가 칼슘 채널을 열어 칼슘이 세포질로 쏟아져 들어오게 한다.[28] 세포질 내 칼슘의 이러한 갑작스러운 증가는 칼모듈린 및 단백질 키나아제 C와 같은 다른 신호 분자를 활성화시킨다.[29] 염화물 및 칼륨과 같은 다른 이온도 세포질 내에서 신호 기능을 할 수 있지만, 이에 대해서는 아직 잘 알려져 있지 않다.[30]
3. 3. 거대분자
세포막이나 세포골격에 결합되지 않은 단백질 분자들은 세포기질로 용해된다. 세포에서 단백질의 양은 매우 높으며 200mg/ml에 근접하는데 이는 세포기질 용적의 약 20~30%를 차지하는 것이다.[79] 그러나 온전한 세포에서 정확히 얼마나 많은 단백질이 세포질에 용해되어 있는지 측정하는 것은 어렵다. 일부 단백질은 온전한 세포에서 막이나 세포 소기관과 약하게 결합되어 있는 것으로 보이며, 세포 용해 시 용액으로 방출되기 때문이다.[15]세포질 내 이러한 고농도의 거대 분자는 거대 분자 혼잡이라고 하는 효과를 유발하는데, 이는 다른 거대 분자의 유효 농도가 증가하는 현상으로, 그들이 이동할 공간이 적기 때문이다. 이러한 혼잡 효과는 세포질 내 반응의 반응 속도와 화학 평형의 위치 모두에서 큰 변화를 일으킬 수 있다.[31] 이는 여러 단백질이 함께 모여 단백질 복합체를 형성하거나 DNA 결합 단백질이 게놈 내의 표적에 결합하는 경우와 같이 거대 분자의 결합을 선호함으로써 해리 상수를 변경하는 능력에서 특히 중요하다.[36]
4. 구조
세포질의 구성 요소는 세포막에 의해 구획으로 분리되지는 않지만, 무작위로 섞이지 않고 특정 분자들이 세포질 내의 정해진 위치에 국소화될 수 있도록 조직화되어 있다.[37]
작은 분자들은 세포질 내에서 빠르게 확산되지만, 농도 기울기는 여전히 생성될 수 있다. 예를 들어, 열린 칼슘 통로 주변에서 짧은 기간 동안 "칼슘 스파크"가 생성된다.[38] 여러 스파크가 합쳐져 "칼슘 파동"이라는 더 큰 기울기를 형성할 수 있다.[39] 산소 및 아데노신 삼인산과 같은 다른 작은 분자들의 농도 기울기는 미토콘드리아 덩어리 주변에 생성될 수 있지만, 이에 대해서는 덜 알려져 있다.[40][41]
단백질은 단백질 복합체를 형성하기 위해 결합할 수 있으며, 이들은 유사한 기능을 가진 단백질 세트를 포함한다.[42] 이러한 구성은 기질 전달을 가능하게 하여 경로를 더 빠르고 효율적으로 만들고, 불안정한 반응 중간체의 방출을 방지한다.[43][44]
단백질 복합체 중 일부는 세포질의 나머지 부분과 격리된 커다란 중심 공동을 포함한다. 프로테아좀이 그 예시인데,[47] 프로테아좀은 세포질 단백질을 분해하는 프로테아제를 포함하는 속이 빈 배럴을 형성한다. 배럴은 분해를 지시하는 신호(유비퀴틴 태그)가 있는 단백질을 인식하여 단백질 분해 공동으로 공급하는 조절 단백질에 의해 덮여 있다.[48]
세균 미세구획은 다양한 효소를 캡슐화하는 단백질 껍질로 만들어진 단백질 구획이다.[49] 이러한 구획은 일반적으로 100nm~200nm이며 서로 맞물린 단백질로 만들어진다.[50] 카르복시좀은 탄소 고정에 관여하는 효소를 포함하는 잘 알려진 예시이다.[51]
세포 골격은 세포질의 일부는 아니지만, 이 필라멘트 네트워크는 세포 내에서 큰 입자의 확산을 제한한다. 예를 들어, 여러 연구에서 약 25 나노미터 (리보솜 크기)보다 큰 추적자 입자가[52] 세포 가장자리와 핵 근처의 세포질 부분에서 배제되었다.[53][54] 이러한 "배제 구획"은 액틴 섬유 네트워크를 포함할 수 있다. 이러한 미세 구획은 세포질 내에서 큰 구조물의 분포에 영향을 미칠 수 있다.[55]
4. 1. 농도 기울기
작은 분자들이 세포질 내에서 빠르게 확산되더라도, 이 구획 내에서 농도 기울기가 여전히 생성될 수 있다. 이에 대한 잘 연구된 예시로는 열린 칼슘 통로 주변 지역에서 짧은 기간 동안 생성되는 "칼슘 스파크"가 있다.[38] 이 스파크는 직경이 약 2µm이며, 단지 몇 밀리초 동안 지속되지만, 여러 스파크가 합쳐져 "칼슘 파동"이라고 불리는 더 큰 기울기를 형성할 수 있다.[39] 산소 및 아데노신 삼인산과 같은 다른 작은 분자들의 농도 기울기는 미토콘드리아 덩어리 주변의 세포에서 생성될 수 있지만, 이에 대해서는 덜 알려져 있다.[40][41]4. 2. 단백질 복합체
단백질은 단백질 복합체를 형성하기 위해 결합할 수 있으며, 이들은 종종 동일한 대사 경로에서 여러 단계를 수행하는 효소와 같이 유사한 기능을 가진 단백질 세트를 포함한다.[42] 이러한 구성은 한 효소의 생성물이 용액으로 방출되지 않고 경로의 다음 효소로 직접 전달되는 기질 전달을 가능하게 한다.[43] 기질 전달은 효소가 세포질에 무작위로 분포되어 있는 것보다 경로를 더 빠르고 효율적으로 만들 수 있으며, 불안정한 반응 중간체의 방출을 방지할 수도 있다.[44] 광범위한 대사 경로는 서로 긴밀하게 결합된 효소를 포함하지만, 다른 경로는 세포 밖에서 연구하기 매우 어려운 느슨하게 관련된 복합체를 포함할 수 있다.[45][46] 결과적으로 이러한 복합체가 전반적인 대사에 미치는 중요성은 여전히 불분명하다.4. 3. 단백질 구획
단백질 복합체 중 일부는 세포질의 나머지 부분과 격리된 커다란 중심 공동을 포함한다. 이러한 폐쇄된 구획의 한 예는 프로테아좀이다.[47] 여기서 일련의 서브유닛은 세포질 단백질을 분해하는 프로테아제를 포함하는 속이 빈 배럴을 형성한다. 이들이 세포질의 나머지 부분과 자유롭게 섞이면 손상을 일으키므로 배럴은 분해를 지시하는 신호(유비퀴틴 태그)가 있는 단백질을 인식하여 단백질 분해 공동으로 공급하는 일련의 조절 단백질에 의해 덮여 있다.[48]단백질 구획의 또 다른 큰 부류는 다양한 효소를 캡슐화하는 단백질 껍질로 만들어진 세균 미세구획이다.[49] 이러한 구획은 일반적으로 100nm~200nm이며 서로 맞물린 단백질로 만들어진다.[50] 잘 알려진 예는 RuBisCO와 같은 탄소 고정에 관여하는 효소를 포함하는 카르복시좀이다.[51]
4. 4. 세포골격에 의한 조절
세포 골격은 세포질의 일부는 아니지만, 이 필라멘트 네트워크의 존재는 세포 내에서 큰 입자의 확산을 제한한다. 예를 들어, 여러 연구에서 약 25 나노미터 (약 리보솜 크기)보다 큰 추적자 입자가[52] 세포 가장자리와 핵 근처의 세포질 부분에서 배제되었다.[53][54] 이러한 "배제 구획"은 나머지 세포질보다 훨씬 더 조밀한 액틴 섬유 네트워크를 포함할 수 있다. 이러한 미세 구획은 일부 영역에서 큰 구조물(예: 리보솜 및 소기관)을 배제하고 다른 영역에 집중시킴으로써 세포질 내에서 큰 구조물의 분포에 영향을 미칠 수 있다.[55]5. 기능
세포질은 원핵생물에서 대부분의 대사가 일어나는 장소이며,[63] 진핵생물 대사 과정의 상당 부분을 차지한다. 포유류의 경우 세포 내 단백질의 약 절반이 세포질에 위치한다.[64] 효모에서 얻은 데이터에 따르면, 대부분의 대사 과정과 대사 산물이 세포질에서 발생한다.[65] 동물 세포질에서 일어나는 주요 대사 경로는 단백질 생합성, 오탄당 인산 경로, 해당 과정, 포도당 신생성이다.[66]
세포질은 대사 산물을 생성 장소에서 사용되는 곳으로 운반하는 기능을 한다. 아미노산과 같이 물에 용해되는 분자는 세포질을 통해 빠르게 확산된다.[14] 그러나 지방산이나 스테롤과 같은 소수성 분자는 특정 결합 단백질에 의해 세포질을 통해 운반되며, 이 단백질은 분자를 세포막 사이에서 이동시킨다.[59][60] 세포 내 섭취로 유입되거나 분비될 예정인 분자는 소포 내에서 세포질을 통해 운반될 수 있으며,[61] 소포는 운동 단백질에 의해 세포 골격을 따라 이동하는 작은 지질 구체이다.[62]
경로의 위치는 유기체에 따라 다를 수 있다. 예를 들어 지방산 합성은 식물의 엽록체[67][68]와 아피콤플렉사의 아피코플라스트에서 발생한다.[69]
5. 1. 세포 분열
세포질은 여러 세포 과정이 일어나는 곳이다. 이러한 과정에는 신호 전달이 세포막에서 세포 핵[56] 또는 세포 소기관과 같은 세포 내 위치로 전달되는 것이 있다.[57] 이 구획은 세포질 분열이 많이 일어나는 곳이며, 유사분열에서 핵막이 분해된 후에도 일어난다.[58]참조
[1]
웹사이트
Cytoplasmic matrix
http://www.oxfordref[...]
Oxford University Press
2006
[2]
저널
Human Anatomy and Physiology Preparatory Course
https://academicwork[...]
CUNY Academic Works
2021-06-22
[3]
서적
On the direction of pyridine nucleotide oxidation-reduction reactions in gluconeogenesis and lipogenesis
https://books.google[...]
Academic
[4]
서적
Oxford dictionary of biochemistry and molecular biology
Oxford University Press
[5]
저널
Das Protoplasma
https://archive.org/[...]
[6]
저널
Compartmentation of metabolism within mitochondria and plastids
2001-04
[7]
저널
Metabolomics by numbers: acquiring and understanding global metabolite data
http://personalpages[...]
2004-05
[8]
저널
Metabolomics in systems biology
[9]
저널
An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR)
[10]
저널
Genome-Scale Reconstruction of the Saccharomyces cerevisiae Metabolic Network
2003-02
[11]
저널
Cytoarchitecture and physical properties of cytoplasm: volume, viscosity, diffusion, intracellular surface area
http://webusers.phys[...]
[12]
저널
Intracellular pH
1981-04
[13]
저널
Fluorescence ratio imaging microscopy: temporal and spatial measurements of cytoplasmic pH
[14]
저널
Solute and macromolecule diffusion in cellular aqueous compartments
2002-01
[15]
저널
Properties and metabolism of the aqueous cytoplasm and its boundaries
1984
[16]
저널
How crowded is the cytoplasm?
1982-09
[17]
저널
The state of water in biological systems
[18]
저널
Do we underestimate the importance of water in cell biology?
2006-11
[19]
저널
Role of water in some biological processes
1990-12-01
[20]
저널
High and low density water and resting, active and transformed cells
1996-06
[21]
저널
Cell water dynamics on multiple time scales
2008-04
[22]
서적
Molecular cell biology
https://archive.org/[...]
Scientific American Books
[23]
저널
Potassium physiology
1986-04-25
[24]
서적
Principles of Renal Physiology, 5th edition
Springer
[25]
저널
Mechanisms and significance of cell volume regulation
2007-10
[26]
저널
Reversible dehydration of trehalose and anhydrobiosis: from solution state to an exotic crystal?
2001-08
[27]
저널
The role of vitrification in anhydrobiosis
[28]
저널
Elementary and global aspects of calcium signalling
http://www.jphysiol.[...]
1997-03-01
[29]
저널
The protein kinase C family: heterogeneity and its implications
[30]
저널
Intracellular monovalent ions as second messengers
2006-04
[31]
저널
Macromolecular crowding: obvious but underappreciated
2001-10
[32]
저널
Organization of Mammalian Cytoplasm
2003-12
[33]
저널
Whatever happened to the 'microtrabecular concept'?
[34]
저널
The bacterial nucleoid: a highly organized and dynamic structure
[35]
서적
Xenopus Protocols
[36]
논문
Macromolecular crowding and confinement: biochemical, biophysical, and potential physiological consequences
[37]
논문
Functional Taxonomy of Bacterial Hyperstructures
2007-03
[38]
논문
Imaging microdomain Ca2+ in muscle cells
2004-04
[39]
논문
Classes and mechanisms of calcium waves
1993-11
[40]
논문
Intracellular compartmentation of organelles and gradients of low molecular weight species
[41]
논문
The cytoplasm: no longer a well-mixed bag
2001-07-20
[42]
논문
Complexes of sequential metabolic enzymes
[43]
논문
Swinging arms and swinging domains in multifunctional enzymes: catalytic machines for multistep reactions
[44]
논문
Channeling of substrates and intermediates in enzyme-catalyzed reactions
[45]
논문
The tentative identification in Escherichia coli of a multienzyme complex with glycolytic activity
1976-06
[46]
논문
Metabolite transfer via enzyme-enzyme complexes
1986-11
[47]
논문
Molecular shredders: how proteasomes fulfill their role
2003-12
[48]
논문
The ubiquitin-proteasome system
http://www.ias.ac.in[...]
2006-03
[49]
논문
Bacterial Microcompartments
http://www.asm.org/A[...]
Am Soc Microbiol
[50]
논문
Protein-based organelles in bacteria: carboxysomes and related microcompartments
2008-08
[51]
논문
CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution
2003-02
[52]
논문
Construction of low-resolution x-ray crystallographic electron density maps of the ribosome
https://zenodo.org/r[...]
2001-11
[53]
논문
Cytoarchitecture of size-excluding compartments in living cells
http://jcs.biologist[...]
1993-10-01
[54]
논문
Hindered diffusion of inert tracer particles in the cytoplasm of mouse 3T3 cells
1987-07
[55]
논문
Effect of cytoarchitecture on the transport and localization of protein synthetic machinery
1993-06
[56]
논문
Four-dimensional organization of protein kinase signaling cascades: the roles of diffusion, endocytosis and molecular motors
2003-06
[57]
논문
Interorganellar communication
2007-12
[58]
논문
Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle
1995-06
[59]
논문
Cytosolic fatty acid binding proteins catalyze two distinct steps in intracellular transport of their ligands
2002-10
[60]
논문
Sterol and lipid trafficking in mammalian cells
2006-06
[61]
논문
The Croonian Lecture 1999. Intracellular membrane traffic: getting proteins sorted
1999-08
[62]
논문
Principles of cargo attachment to cytoplasmic motor proteins
2002-02
[63]
논문
Principles of macromolecular organization and cell function in bacteria and archaea
[64]
논문
A mammalian organelle map by protein correlation profiling
2006-04
[65]
논문
A consensus yeast metabolic network reconstruction obtained from a community approach to systems biology
2008-10
[66]
서적
Biochemistry
https://archive.org/[...]
W.H. Freeman
[67]
논문
Fatty acid synthesis: from CO2 to functional genomics
2000-12
[68]
논문
Subcellular localization of acyl carrier protein in leaf protoplasts of Spinacia oleracea
1979-03
[69]
논문
Fatty acid biosynthesis as a drug target in apicomplexan parasites
2007-01
[70]
논문
Inside a living cell
1991-06
[71]
의학용어사전
대한의협 의학용어 사전
https://www.kmle.co.[...]
[72]
의학용어사전
대한의협 의학용어 사전
https://www.kmle.co.[...]
[73]
웹인용
Cytoplasmic matrix
http://www.oxfordref[...]
Oxford University Press
2006
[74]
저널
Human Anatomy and Physiology Preparatory Course
https://academicwork[...]
CUNY Academic Works
2021-06-22
[75]
저널
Cytoarchitecture and physical properties of cytoplasm: volume, viscosity, diffusion, intracellular surface area
http://webusers.phys[...]
[76]
저널
Intracellular pH
1981-04-01
[77]
저널
Fluorescence ratio imaging microscopy: temporal and spatial measurements of cytoplasmic pH
[78]
서적
Molecular cell biology
https://archive.org/[...]
Scientific American Books
[79]
저널
Macromolecular crowding: obvious but underappreciated
2001-10-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com