오록스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오록스는 들소를 의미하는 게르만어 또는 켈트어 단어에서 유래되었으며, 유라시아, 북아프리카, 인도 아대륙에 서식했던 멸종된 소과 동물이다. 과거에는 유라시아 오록스, 아프리카 오록스, 인도 오록스의 세 아종이 있었던 것으로 알려져 있다. 오록스는 플라이스토세 동안 진화했으며, 17세기까지 동유럽에 생존했으나, 과도한 사냥과 서식지 감소, 가축화 등으로 인해 1627년 멸종되었다. 20세기 초부터 헤크 형제에 의해 오록스의 특징을 가진 소 품종을 교배하여 오록스를 되살리려는 시도가 있었으며, 현재 타우루스 소, 타우로스 프로그램, 우루즈 프로젝트 등이 진행되고 있다.
더 읽어볼만한 페이지
- 1827년 기재된 화석 분류군 - 메갈로돈 (이매패류)
메갈로돈 (이매패류)은 신생대 지층에서 주로 발견되는 이매패류로, 페름기에는 발견되지 않았으며 트라이아스기 및 쥐라기 시대의 분포 정보는 알려지지 않았다. - 1827년 기재된 화석 분류군 - 스텝들소
스텝들소는 플라이스토세 유라시아에 살았던 들소의 일종으로, 현생 들소들의 조상으로 여겨지며 남아시아에서 유라시아, 베링 육교를 통해 북아메리카까지 확산되었고, 동굴 벽화나 미라 형태로 발견되며 복제 연구가 진행 중이다. - 고대 소과 - 자이언트들소
자이언트들소는 북아메리카에 서식했던 멸종된 들소로, 스텝들소에서 진화하여 비손 안티쿠스를 낳았으며, 멕시코 남부에서 캐나다 남부에 걸쳐 분포했고, 인류의 사냥 압력, 기후 변화, 환경 변화 등이 멸종 원인으로 추정된다. - 고대 소과 - 스텝들소
스텝들소는 플라이스토세 유라시아에 살았던 들소의 일종으로, 현생 들소들의 조상으로 여겨지며 남아시아에서 유라시아, 베링 육교를 통해 북아메리카까지 확산되었고, 동굴 벽화나 미라 형태로 발견되며 복제 연구가 진행 중이다. - 아시아의 멸종한 포유류 - 시리아야생당나귀
시리아야생당나귀는 서아시아에 서식했던 멸종된 야생 당나귀 아종으로, 무분별한 사냥과 서식지 파괴로 1920년대에 멸종되었으며, 멸종 후 유전자 보존 및 생태적 역할 복원을 위한 노력이 진행 중이다. - 아시아의 멸종한 포유류 - 사우디가젤
사우디가젤은 아라비아 반도에 서식했던 가젤의 한 종류로, 과도한 사냥으로 1980년 야생절멸된 후 2008년 멸종으로 공식 선언되었으며, 도르카스 가젤의 아종으로 분류되기도 했으나 계통 발생 분석 결과 자매 분류군으로 밝혀졌다.
| 오록스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 멸종 시기 | 1627년 |
| 상태 | 절멸 (EX) |
| 학명 | Bos primigenius |
| 학명 명명자 | Bojanus, 1825 |
| 분포 | 유라시아 대륙, 북아프리카 |
| 몸높이 (어깨) | 약 180cm (수컷), 약 155cm (암컷) |
| 뿔 길이 | 약 80cm |
| 로마자 표기 | Aurochs (오록스) |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 목 | 경우제목 |
| 아목 | 소아목 |
| 과 | 소과 |
| 아과 | 소아과 |
| 속 | 소속 |
| 하위 분류 | |
| 아종 | Bos primigenius primigenius (유럽 오록스) Bos primigenius namadicus (인도 오록스) Bos primigenius mauretanicus (북아프리카 오록스) |
2. 명칭
"aur"와 "ur"는 모두 "들소"를 의미하는 게르만어 또는 켈트어 단어이다.[2][3] 고대 고지 독일어에서 이 단어는 "ohso"(소)와 복합어를 이루어 "ūrohso"가 되었고, 이것이 근대 초 "오록스"가 되었다.[25] 라틴어 단어 "urus"는 갈리아 전쟁 이후 들소를 지칭하는 데 사용되었다.[3][4]
칼 린네는 1758년에 폴란드의 야생 소에 대해 ''Bos taurus''라는 학명을 부여했다.[7] 1825년에는 루드비히 하인리히 보야누스가 오록스의 골격과 가축의 골격을 비교하여 ''Bos primigenius''라는 학명을 제안했고, 이는 1827년에 출판되었다.[8][9] ''Bos namadicus''는 1859년 휴 팔코너가 네르부다 퇴적물에서 발견된 소 화석에 사용한 이름이다.[10] 1881년 필리프 토마스는 콘스탄틴 서쪽의 Oued Seguen 근처 퇴적물에서 발견된 화석을 설명하며 ''Bos primigenius mauritanicus''라는 이름을 붙였다.[11]
영어에서 복수형 "aurochsen"의 사용은 독일어 복수형 "Ochsen"과 직접 연결되며, 영어 단수형 "ox"와 복수형 "oxen"과 같은 구분을 유사하게 재현한다. 오록스는 단수와 복수 모두를 나타낼 수 있다.[5][6]
이 동물을 가리키는 명칭으로 영어에서는 aurochs영어나 urus영어가 사용되는데, 전자는 독일어에서, 후자는 라틴어에서 유래했다. 600년경 세비야의 이시도르 대주교에 따르면, urus영어는 산을 의미하는 그리스어 όροςel에서 유래했다. 원래 들소를 가리키는 wisent영어라는 단어도 자주 오용되는데, 이는 들소와 오록스가 매우 오래전부터 혼동되어 온 역사적 배경 때문이다. 율리우스 카이사르가 갈리아 원정 중에 오록스를 "코끼리보다 약간 작고, 모습과 색깔은 소와 매우 비슷하다"고 기록했을 당시에는 들소와 오록스는 각각 bonasusla와 urusla로 명확히 구분되었지만, 약 1세기 후 대 플리니우스의 『박물지』에서는 이미 "사람들이 들소와 오록스를 혼동한다"고 한탄하고 있다. 이러한 경향은 들소와 오록스 모두 개체수가 줄어들면서 가속화되었고, 결국 둘이 다른 동물이라는 사실조차 잊혀지게 되었다.
3. 분류
2003년, 국제 동물 명명 위원회는 야생 종에 대한 ''Bos primigenius'' 이름의 유효성을 인정하여 ''동물학 공식 종명 목록''에 올렸다.[12][13]
전통적으로 알려진 세 아종은 다음과 같다.아종 분포 지역 비고 유라시아 오록스(B. p. primigenius) 유라시아 플라이스토세 거대 동물군의 일부, 동유럽에서 17세기까지 생존.[14] 인도 오록스(B. p. namadicus) 인도 아대륙 [15] 북아프리카 오록스(B. p. mauritanicus) 사하라 사막 북쪽 B. p. opisthonomus라고도 불림.[25][16]
21세기 중국 유전학자들은 미토콘드리아 DNA 증거를 통해 중국 북부의 유라시아 오록스 개체군이 플라이스토세 동안 유전적으로 격리되어 형태적으로 뚜렷하지 않더라도 별도의 아종인 동아시아 오록스(''B. p. sinensis'')로 간주될 수 있다고 발표했다.[2]
플라이스토세 동안 해수면 변화로 인해 지중해 섬에서 최소 두 종의 왜소 아종 오록스가 발달했다.
영어에서는 이 동물을 지칭할 때 aurochs영어나 urus영어를 사용하는데, 전자는 독일어에서, 후자는 라틴어에서 유래했다. 600년경 세비야의 이시도르 대주교는 urus영어가 산을 의미하는 그리스어 όροςel에서 유래했다고 한다. 들소를 가리키는 wisent영어라는 오용도 자주 사용되는데, 이는 들소와 오록스가 매우 오래전부터 혼동되어 온 사실 때문이다. 율리우스 카이사르는 갈리아 원정 중에 오록스를 "코끼리보다 약간 작고, 모습과 색깔은 소와 매우 비슷하다"고 기록했을 당시에는 들소와 오록스는 각각 bonasusla와 urusla로 명확히 구분되었지만, 약 1세기 후 대 플리니우스의 『박물지』에서는 이미 "사람들이 들소와 오록스를 혼동한다"고 한탄하고 있다. 이러한 경향은 들소와 오록스 모두 개체수가 줄어들면서 가속화되었고, 결국 둘이 다른 동물이라는 사실조차 잊혀지게 되었다.
학명으로는, 가축종은 야생종의 학명을 사용한다는 관습에 따라, 가축 소(''Bos taurus'')와 혹소(''Bos indicus'')는 그 조상인 오록스와 같은 ''Bos primigenius''라는 학명을 사용하는 경향이 있다. 가축종을 명확히 가리킬 때는 ''B. p. taurus'' 또는 ''B. p. indicus''로 하거나, 구례대로 ''Bos taurus'' 또는 ''Bos indicus''로 한다. 원래 ''Bos taurus''가 ''Bos primigenius''보다 더 오래된 명칭이므로, 둘이 동일종으로 간주될 경우 학명의 우선권은 ''Bos taurus''에 있었다. 그러나 야생종에 가축종의 학명을 적용하는 것은 큰 혼란을 야기한다고 판단하여, 2003년 국제 동물 명명 규약 위원회가 우선권을 ''Bos primigenius''로 변경했다[127](Opinion 2027).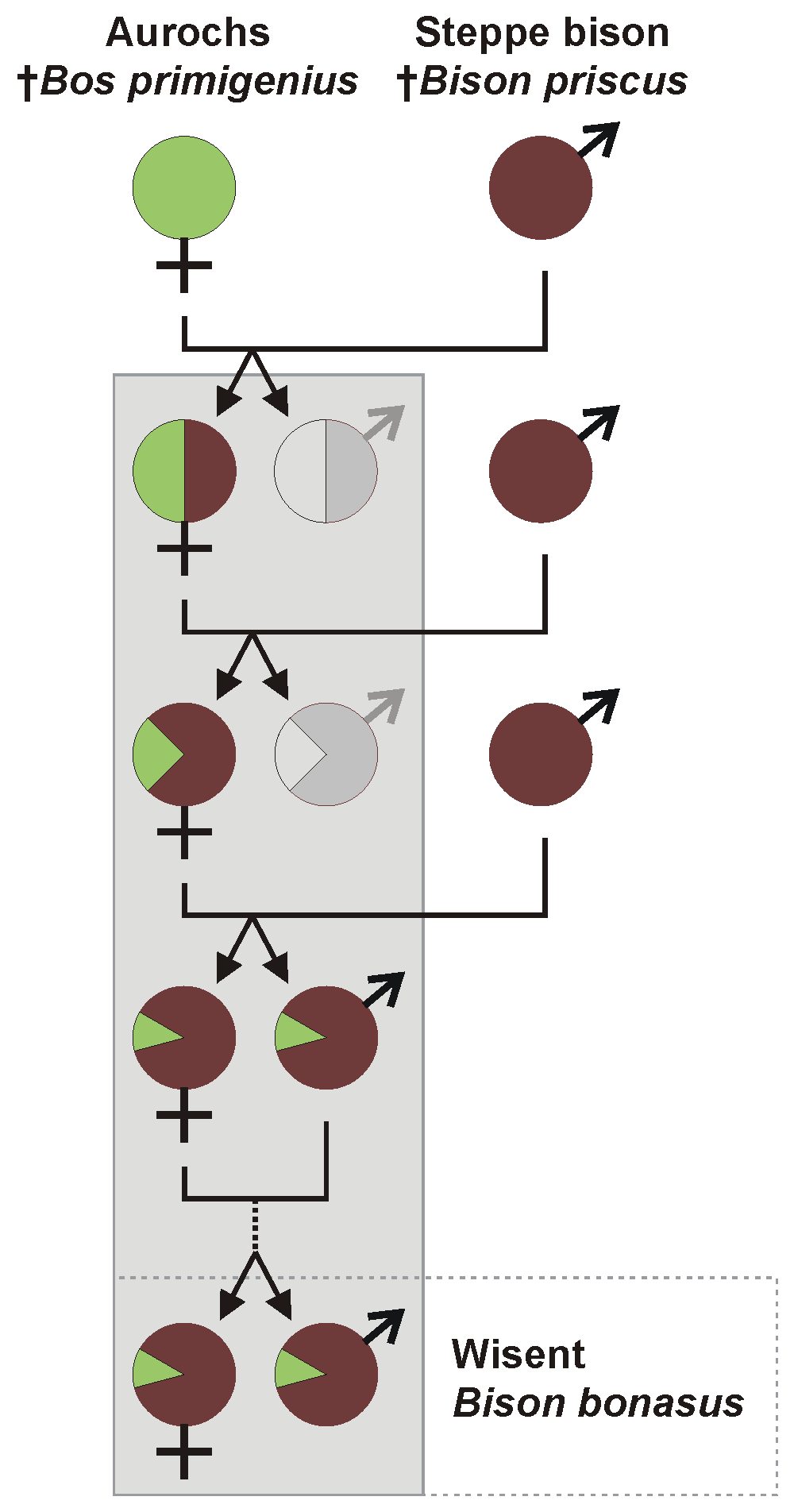
소과의 동물은 자연계에서 다른 종끼리 반복적으로 교배하여 잡종을 만드는 경우가 있어, 각 종의 유전적 계통이나 관계를 명확히 하기 어렵다.[128] 예를 들어 유럽 들소와 코카서스 들소는 북미로 유입된 스텝 들소의 후손이 유라시아 대륙에 복귀한 후, 암컷 오록스와의 사이에서 만들어낸 잡종 "'''힉스 들소'''"(힉스 입자의 영어 이름 Higgs Boson영어을 따서 명명)의 후손이라고 여겨진다.[129][130][131][132]
4. 진화
칼 린네는 1758년 폴란드의 야생 소에 대해 ''Bos taurus''라는 학명을 붙였다.[7] 1825년 루드비히 하인리히 보야누스는 오록스와 가축의 골격 차이를 비교하여 ''Bos primigenius''라는 학명을 제안했고, 이 학명은 1827년에 출판되었다.[8][9] 1859년 휴 팔코너는 네르부다 퇴적물에서 발견된 소 화석에 ''Bos namadicus''라는 이름을 사용했다.[10] 필리프 토마스는 1881년 콘스탄틴 서쪽의 Oued Seguen 근처 퇴적물에서 발견된 화석을 설명하며 ''Bos primigenius mauritanicus''라는 학명을 붙였다.[11]
2003년, 국제 동물 명명 위원회는 ''Bos primigenius''를 동물학 공식 종명 목록에 올려 야생 종에 대한 이 이름의 유효성을 인정했다.[12][13]
전통적으로 세 개의 아종이 역사적인 시기에 존재했던 것으로 알려져 있다.
- 유라시아 오록스(''B. p. primigenius''): 유라시아의 플라이스토세 거대 동물군의 일부였으며 동유럽에서 17세기까지 생존했다.[14]
- 인도 오록스(''B. p. namadicus''): 인도 아대륙에 서식했다.[15]
- 북아프리카 오록스(''B. p. mauritanicus''): 사하라 사막 북쪽에 서식했으며, ''B. p. opisthonomus''라고도 불렸다.[25][16]
21세기에 중국의 유전학자들은 미토콘드리아 DNA 증거를 발표하여 중국 북부의 유라시아 오록스 개체군이 플라이스토세의 상당 기간 동안 유전적으로 격리되었으며, 형태적으로 뚜렷하지 않더라도 별도의 아종인 동아시아 오록스(''B. p. sinensis'')로 간주될 만큼 충분히 독특하다는 것을 뒷받침했다.[2]
플라이스토세 동안 해수면 변화의 결과로 최소 두 종의 왜소 아종의 오록스가 지중해 섬에서 발달했다.
보정과 16종의 소과 종의 화석을 사용한 결과에 따르면, 소족은 약 1170만 년 전에 진화했다.[20] ''Bos''와 ''들소''의 유전 계통은 약 250만 년 전에서 165만 년 전 사이에 소족으로부터 유전적 분기된 것으로 추정된다.[21][22]
추운 플라이스토세 기후는 열린 초원의 확장을 유발하여 대형 초식 동물의 진화를 가능하게 했다.[25] 오록스의 기원은 불분명하며, 아프리카 또는 아시아 기원으로 추정된다. ''Bos acutifrons''는 오록스의 가능한 조상으로 여겨지며, 약 200만 년 전의 초기 플라이스토세로 거슬러 올라가는 화석 두개골이 인도의 시발리크 언덕에서 발굴되었다.[26]
약 78만 년 전으로 거슬러 올라가는 초기 중기 플라이스토세 지층에서 튀니지의 케프 주에서 발굴된 오록스 두개골은 현재까지 가장 오래된 연대 측정된 화석 표본이다. 이 연구의 저자들은 ''Bos''가 아프리카에서 진화하여 중기 플라이스토세 동안 유라시아로 이주했을 수 있다고 제안했다.[27] 중기 플라이스토세 오록스 화석은 사하라의 에르그와 호가르 산맥에서도 발굴되었다.[28]
인도 아종(''Bos primigenius namadicus'')의 화석은 중기 플라이스토세로 거슬러 올라가는 충적 퇴적물에서 남인도에서 발굴되었다.[29] 오록스의 유해는 인도 아대륙 전역의 후기 플라이스토세 유적지에서 흔하다.[103]
유럽에서 가장 초기의 화석은 중기 플라이스토세로 거슬러 올라간다. 역사적으로 오록스가 유럽에서 처음 등장한 것으로 널리 알려진 유적지는 약 60만 년 전의 이탈리아 남부의 노타르치리코 유적지였지만,[30] 2024년 이 유적지에 대한 재조사 결과, 이 지역에 오록스가 존재했다는 증거가 뒷받침되지 않았으며, 현재 오록스의 가장 오래된 기록은 이탈리아 중부의 폰테 몰레 유적지에서 발견되었으며, 약 55만~45만 년 전으로 거슬러 올라간다.[31][32] 오록스는 약 40만 년 전 해양 동위체 단계 11에 영국에 존재했다.[33]
동아시아에서 가장 초기의 오록스 유해는 불확실하지만, 후기 중기 플라이스토세로 거슬러 올라갈 수 있다.[34]
후기 플라이스토세 오록스 화석은 수단의 아파드 23에서 발견되었으며, 이 지역의 기후가 아프리카 습윤기보다 더 습했던 5만 년 전으로 거슬러 올라간다.[35]
가장 최근의 빙하 후퇴 이후, 오록스의 분포는 홀로세 초에 덴마크와 스웨덴 남부로 확장되어 약 1만 2천~1만 1천 년 전으로 거슬러 올라간다.[36]
약 200만 년 전에 인도 주변에서 진화한 것으로 생각된다. 오록스(Bos primigeniusla)는 제4기 초두에 중동으로 분포를 넓혔으며, 유럽에 도달한 것은 약 25만 년 전으로 여겨진다.[133] 홍적세 말기(1만 1000년 전)에는 유럽, 아시아, 북아프리카 등의 넓은 범위에 분포했으며, 1만 5000년 전의 라스코 동굴 벽화에도 오록스가 그려져 있다.
일본 열도에서는 이와테현의 하나이즈미 유적에서 오록스의 화석이 발견되었다.[134] 하지만 2007년 하나이즈미산 척추동물 화석 목록에서는 오록스 기록이 제외되었다.[135]
5. 형태
지그문트 폰 헤르버스타인의 16세기 설명에 따르면, 오록스는 등줄기를 따라 회색 줄무늬가 있는 검은색 털을 가지고 있었다.[37] 1980년대 초까지 오록스의 털 유해는 알려지지 않았다.[40] 묘사에 따르면 북아프리카 오록스는 등에 밝은 안장 무늬가 있었을 수 있다.[41] 송아지는 밤색으로 태어났을 것이며, 어린 수소는 등줄기를 따라 흰색 뱀줄이 있는 검은색으로 변했고, 암소는 붉은 갈색을 유지했다. 암수 모두 밝은 색의 주둥이를 가지고 있었지만, 털색의 변이에 대한 증거는 존재하지 않는다.[25]
많은 원시 소 품종, 특히 남유럽 품종은 오록스와 유사한 털색을 나타내며, 밝은 뱀줄이 있는 수소의 검은색, 옅은 입, 색상의 유사한 성적 이형성을 포함한다.[25][41] 오록스에 대한 역사적 설명에 따르면, 길고 곱슬거리는 이마 털을 가지고 있었지만, 특정 색상을 언급한 내용은 없다.[25]
오록스의 체형과 몸매는 많은 현대 소 품종과 확연히 달랐다. 다리가 상당히 더 길고 가늘어서 어깨 높이가 몸통 길이와 거의 같았다. 큰 뿔이 달린 두개골은 대부분의 소 품종보다 훨씬 크고 길쭉했다. 다른 야생 소과 동물과 마찬가지로, 오록스의 체형은 탄탄했고, 특히 수컷의 경우 목과 어깨 근육이 강하게 발달했다. 따라서 앞부분이 뒷부분보다 컸는데, 이는 유럽들소와 유사하지만, 많은 가축과는 달랐다. 임신한 암소에서도 유방은 작았고 옆에서 거의 보이지 않았다. 이 특징은 다른 야생 소과 동물과 동일하다.[25]
오록스는 홀로세 유럽에서 가장 큰 초식 동물 중 하나였다. 오록스의 크기는 지역에 따라 달랐던 것으로 보이는데, 북유럽의 표본이 남쪽보다 더 컸다. 덴마크와 독일의 오록스는 수컷의 어깨 높이가 155cm에서 180cm였고 암컷은 135cm에서 155cm였으며, 헝가리의 오록스 수컷은 160cm에 달했다.[42] 오록스의 체중은 어느 정도 변동성을 보인 것으로 보인다. 어떤 개체는 약 700kg에 달했고, 후기 중기 플라이스토세의 개체는 최대 1500kg까지 나갔을 것으로 추정된다.[25] 오록스는 수컷과 암컷의 크기에서 상당한 성적 이형성을 보였다.[44]
뿔은 거대하여 길이가 80cm에 달하고 직경은 10cm에서 20cm였다.[41] 뿔은 앞을 향한 주둥이에 대해 60도 각도로 머리에서 자라났으며, 세 방향으로 굽어 있었다. 즉, 밑부분에서 위와 바깥쪽으로 굽었다가, 앞과 안쪽으로 휘어지고, 다시 안과 위로 휘어졌다. 수소 뿔의 굽이는 암소의 뿔보다 더 강하게 나타났다.[25] 뿔심의 밑부분 둘레는 가장 큰 중국 표본에서 44.5cm에 달했고, 프랑스 표본에서는 48cm에 달했다.[49]
6. 생태
어린 오록스 개체는 회색늑대나 큰곰의 먹잇감이었다. 선사시대 유럽, 북아프리카, 아시아에서는 사자, 호랑이, 하이에나 등에게도 잡아먹혔다.[44]
오록스는 주로 겨울에는 작은 무리를 이루었지만, 여름에는 보통 혼자 또는 더 작은 무리로 생활했다.[48] 오록스는 사회적 지위를 얻기 위해 암컷과 수컷 모두 과시와 싸움을 했다.[41] 고치형 치아를 가진 초식 동물로, 풀, 잔가지, 도토리 등을 먹었다.[25] 홀로세 덴마크 오록스의 앞니 중마모 분석에 따르면, 덴마크 선보레알 시대에는 마모가 주된 초식 동물이었지만, 보레알, 대서양, 아보레알 시대에는 혼합된 먹이를 먹는 동물로 변했다.[56] 플라이스토세 영국의 표본에 대한 치아 미세마모 및 중마모 분석 결과, 엄격한 초식 동물보다는 혼합된 먹이를 먹거나 나뭇잎을 먹는 식단을 가졌다.[57]
짝짓기는 9월에 이루어졌고, 송아지는 봄에 태어났다.[48] 수컷들은 격렬한 싸움을 벌였으며, 약토로프 숲의 증거는 서로에게 치명적인 부상을 입힐 수 있다는 것을 보여준다. 가을에 오록스는 겨울을 위해 살을 찌우고 윤기 있는 털을 가졌다. 송아지는 무리를 따라갈 수 있을 만큼 강해질 때까지 어미와 함께 머물렀다. 오록스 송아지는 회색 늑대와 불곰의 포식에 취약했지만, 건강한 성체 오록스는 크고 힘이 세서 대부분의 포식자를 두려워할 필요가 없었다.[25] 역사적 묘사에 따르면, 오록스는 체격에도 불구하고 민첩하고 도발당하면 매우 공격적이었으며, 인간을 두려워하지 않았다.[25] 중기 플라이스토세 유럽에서 오록스는 "유럽 재규어" ''Panthera gombaszoegensis''와 검치호랑이의 먹잇감이었을 가능성이 높고,[58] 후기 플라이스토세 이탈리아에서 동굴 하이에나에 의해 오록스가 소비되었다는 증거가 발견되었다.[59] 사자, 호랑이, 늑대는 홀로세 동안 오록스의 주요 포식자였을 것으로 생각된다.[44]
유럽의 중기 플라이스토세와 초기 후기 플라이스토세의 간빙기 동안 오록스는 검치상아 코끼리, 메르크 코뿔소, 코뿔소, 큰사슴을 포함한 다른 큰 온대 적응 거대 동물 종과 함께 나타났다.[60]
7. 분포 및 서식지
오록스는 전통적으로 세 개의 아종이 역사적인 시기에 존재했던 것으로 알려져 있다.
- 유라시아 오록스(''B. p. primigenius'')는 유라시아의 플라이스토세 거대 동물군의 일부였으며 동유럽에서 17세기까지 생존했다.[14]
- 인도 오록스(''B. p. namadicus'')는 인도 아대륙에 서식했다.[15]
- 북아프리카 오록스(''B. p. mauretanicus'')는 사하라 사막 북쪽에 서식했다.[25]
21세기에 중국의 유전학자들은 중국 북부의 유라시아 오록스 개체군이 플라이스토세의 상당 기간 동안 유전적으로 격리되어 별도의 아종인 동아시아 오록스(''B. p. sinensis'')로 간주될 만큼 충분히 독특하다는 미토콘드리아 DNA 증거를 발표했다.[2]
플라이스토세 동안 해수면 변화의 결과로 최소 두 종의 왜소 아종의 오록스가 지중해 섬에서 발달했다.
약 200만 년 전에 인도 주변에서 진화한 것으로 생각되는 오록스는 제4기 초두에 중동으로 분포를 넓혔으며, 유럽에 도달한 것은 약 25만 년 전으로 여겨진다.[133] 홍적세 말기(1만 1000년 전)에는 유럽, 아시아, 북아프리카 등의 넓은 범위에 분포했으며, 1만 5000년 전의 라스코 동굴 벽화에도 오록스가 그려져 있다.
오록스는 북아프리카, 메소포타미아, 유럽 전역, 폰토스-카스피 스텝, 코카서스, 서쪽의 서시베리아에서 널리 분포했으며, 북쪽으로는 핀란드만과 라도가 호까지 분포했다.[48] 한반도와[51] 일본 열도에서도 화석이 발굴되었다.[52][53]
따뜻한 간빙기 동안 오록스는 유럽 전역에 널리 퍼져 있었지만,[60] 빙하기 동안에는 이베리아 반도, 이탈리아 반도, 발칸 반도의 남부 생물 피난처로 물러났다.[47]
유럽의 풍경은 지난 수천 년 동안 대부분 울창한 숲으로 이루어져 있었을 것이다. 오록스는 호수 주변의 강가 숲과 습지를 사용했을 가능성이 높다.[44] 영국에서 발견된 표본 분석 결과 오록스는 낮고 비교적 평평한 지형을 선호했던 것으로 보인다.[54] 중국에서 오록스 유해와 함께 화석 퇴적물에서 발견된 대부분 작은 관목의 꽃가루는 온대 기후의 풀이 무성한 평원이나 초원이 삼림에 접해 있는 곳을 선호했음을 나타낸다.[49] 또한 열린 초원에서도 살았을 수 있다.[55] 홀로세의 따뜻한 대서양기에 오록스는 남아 있는 개방된 지역과 숲 가장자리로 제한되었으며, 가축과 인간과의 경쟁이 점차 증가하면서 오록스의 쇠퇴로 이어졌다.[56]
8. 인간과의 관계
오록스는 인간과의 관계 속에서 다양한 형태로 나타난다. 하라파와 모헨조다로에서 발견된 인더스 문명의 인장에서는 오록스와 같은 굽은 뿔을 가진 동물을 볼 수 있다.[67][68] 서부 코카서스의 마이코프 문화에서는 오록스 조각상이 만들어졌다.[69]
무레이베트, 할란 체미, 차요뉴 정착지에서 발견된 오록스 뼈와 두개골은 무문 토기 신석기 B 문화에서 음식을 저장하고 공유했음을 보여준다.[65] 약 3,700년 전 레바논 시돈의 묘지에서 발견된 오록스 유해는 수많은 동물, 소수의 인간 뼈, 음식과 함께 묻혀 있었다.[66]
아카드어에서 오록스는 rīmu와 rēmu로 표시되며, 아카드 왕조의 나람-신, 티글라트-필레세르 1세, 살만에세르 3세와 같은 통치자들의 사냥과 관련되어 사용되었다. 메소포타미아에서 오록스는 힘과 성적 능력을 상징했으며, 신 엔릴과 샤마시의 별칭이자 왕 산헤립과 영웅 길가메시의 기량을 나타내는 별칭이었다. 우가리트 문헌에서 야생 황소는 신 바알에게 사냥되고 제물로 바쳐지는 것으로 자주 언급된다.[70] 기원전 6세기에 건설된 바빌론의 이슈타르 문에도 오록스가 묘사되어 있으며,[71] 페르가몬 박물관에서 전시 중인 이슈타르 문 부조에서 확인할 수 있다.
나일강 상류 계곡의 쿠르타에서 발견된 오록스 암각화는 루미네선스 연대 측정법을 사용하여 기원전 19,000~15,000년으로 거슬러 올라가며, 아프리카에서 발견된 가장 오래된 조각품이다.[72] 기원전 20세기 테베 무덤의 부조와 기원전 1175년경 메디네트 하부에 있는 람세스 3세 장제전의 사냥 장면에도 오록스가 등장한다. 후자는 현재까지 고대 이집트 예술에서 오록스를 묘사한 가장 오래된 작품이다.[73]
프랑스 북부 중기 구석기 시대 유적 비아슈-생-바스트에서는 약 24만 년 전 네안데르탈인이 오록스를 도살한 증거(불에 탄 뼈와 절단 자국)가 발견되었다.[74][75] 스페인의 후기 중기 구석기 시대 동굴 쿠에바 데스-쿠비에르타 유적에서는 네안데르탈인이 오록스 두개골을 사냥 전리품으로 보관했던 것으로 추정된다.[76]
프랑스 남부 쇼베와 라스코 동굴의 구석기 시대 동굴 벽화에는 기원전 3만 6천 년에서 2만 1천 년 사이의 오록스가 널리 묘사되어 있다.[77] 칼라브리아 로미토 동굴의 구석기 시대 암각화[78], 이탈리아 레반조 섬 제노베제 동굴[79], 이베리아 반도 그라베티안 문화에서 마들렌 문화에 이르는 동굴[80][81][82] 에서도 오록스 암각화와 그림이 발견되었다.
프랑스, 룩셈부르크, 독일, 네덜란드, 잉글랜드, 덴마크의 다양한 중석기 시대 사냥 및 도살 유적지에서 절단 자국이 있는 오록스 뼈가 발견되었다.[83] 에스토니아 나르바강과 에마요기강 인근 중석기 시대 정착지, 잉글랜드 여러 신석기 시대 유적(구덩이와 소각구)에서도 오록스 뼈가 발견되었다.[84][85]
그리스 바피오에서 발견된 컵에는 미케네 그리스 시대 사람들이 오록스를 잡으려는 사냥 장면이 묘사되어 있다. 황소 한 마리가 사냥꾼 한 명을 땅에 내던지고 다른 한 명을 뿔로 공격한다.[86][87][88] 그리스인과 파이오니아인은 오록스를 사냥하여 거대한 뿔을 전리품, 술잔, 신과 영웅에게 바치는 제물로 사용했다. 사무스, 테살로니카의 필리푸스, 안티파테르가 언급한 마케도니아의 필리포스 5세는 오르빌로스 산기슭에서 오록스를 죽이고 105cm 길이의 뿔과 가죽을 헤라클레스 신전에 바쳤다.[89]
율리우스 카이사르는 ''갈리아 전기''에서 오록스를 묘사했다.[4] 콜로세움과 같은 로마 원형 경기장의 베나티오 쇼에서는 오록스를 포획하여 전시하기도 했다.[90] 로마인들은 오록스 뿔을 사냥 뿔로 자주 사용했다.[25]
니벨룽겐의 노래de에서 지크프리트는 오록스 4마리를 죽인다.[91] 중세 시대에는 오록스 뿔이 술잔으로 사용되었으며, 마지막 황소의 뿔도 포함되었다. 오늘날 많은 오록스 뿔 덮개가 보존되어 있다.[92] 케임브리지 코퍼스 크리스티 칼리지의 오록스 술잔은 17세기에 칼리지 문장으로 새겨졌다.[93] 뿔 사이에 별이 있는 오록스 머리와 기독교 도상학 요소는 수 세기 동안 지속된 몰도바 문장을 나타낸다.[94]
오록스는 화살, 그물, 사냥개로 사냥했으며, 산 오록스의 이마 털을 잘라 만든 허리띠는 여성의 다산을 증가시키는 것으로 여겨졌다. 오록스를 도살할 때 심장에서 추출한 ''심장뼈''는 오록스에게 부여된 신비와 마법의 힘에 기여했다.[25] 동유럽에서는 "오록스처럼 행동한다"(취한 사람이 난폭하게 행동하는 것), "오록스 같은 남자"(크고 힘센 사람)와 같은 표현이 사용되었다.[44]
영어에서 오록스를 가리키는 "aurochs영어"는 독일어에서, "urus영어"는 라틴어에서 유래했다. 600년경 세비야의 이시도르 대주교는 "urus영어"가 산을 의미하는 그리스어 όροςel에서 유래했다고 주장한다. 원래 들소를 가리키는 "wisent영어"라는 오용도 자주 사용되는데, 이는 들소와 오록스가 오랫동안 혼동되어 온 사실에 기인한다. 율리우스 카이사르는 갈리아 원정 중 오록스를 "코끼리보다 약간 작고, 모습과 색깔은 소와 매우 비슷하다"고 기록하며 "bonasusla"(들소)와 "urusla"(오록스)를 명확히 구분했지만, 약 1세기 후 대 플리니우스는 『박물지』에서 "사람들이 들소와 오록스를 혼동한다"고 한탄한다. 이러한 경향은 들소와 오록스 모두 개체수가 줄면서 가속화되었고, 결국 둘이 다른 동물이라는 사실조차 잊혀지게 되었다.
학명과 관련하여, 가축종은 야생종의 학명을 사용한다는 관습에 따라, 한때 라는 학명을 부여했던 가축 소와 학명을 가진 혹소는 그 조상인 오록스와 같은 학명을 사용하는 경향이 있다. 가축종임을 명확히 하기 위해 또는 아종으로 표기하거나, 또는 로 표기하기도 한다. 가축 소에 부여된 와 오록스의 중 더 오래된 명칭은 이므로, 동일종으로 간주될 경우 학명의 우선권은 에 있었다. 그러나 야생종에 가축종 학명을 적용하는 혼란을 막기 위해 2003년 국제 동물 명명 규약 위원회는 에 우선권을 부여했다.[127](Opinion 2027)
8. 1. 가축화
인도 남부 데칸 고원의 후나사기에 있는 아슐리안 지층에서 절단된 자국이 있는 오록스 뼈가 발견되었다.[61] 이스라엘 네셰르 람라 호모 유적지의 중기 구석기 시대 지층에서는 부싯돌로 인해 생긴 절단 자국이 있는 오록스 뼈가 발견되었는데, 이는 약 12만 년 전의 해양 동위원소 단계 5 시기로 거슬러 올라간다.[62]가장 초기에 오록스를 가축화한 것은 비옥한 초승달 지대의 신석기 혁명 시기로, 신석기 시대 농부들이 사냥하고 사육한 소는 기원전 9800년에서 7500년 사이에 점차 크기가 작아졌다. 무레이베트와 괴베클리 테페에서 발견된 오록스 뼈는 시리아 북부의 자데 엘-무가라나 텔 할룰라와 같은 신석기 시대 후기 정착지의 소 뼈보다 크다.[95] 기원전 6천년기에 해당하는 이라크 북부와 이란 서부의 후기 신석기 시대 유적에서도 소 유해가 작지만 더 자주 발견되는데, 이는 할라프 문화 기간 동안 가축화된 소가 비옥한 초승달 지대 중앙 지역에서 수입되었음을 나타낸다.[96]
유전 연구 결과에 따르면, 현대의 털 소(''Bos taurus'')는 약 10,500년 전 아나톨리아 남동부와 시리아 북부에서 길들여진 오록스 80마리에서 유래되었다.[14] 털 소는 다뉴브 강과 지중해 해안을 따라 발칸 반도와 이탈리아 북부로 퍼져나갔다.[97] 오록스 수컷과 초기 가축의 잡종화는 기원전 9500년에서 1000년 사이에 중부 유럽에서 발생했다.[98]
17,000~7,000년 전과 51개 현대 소 품종으로 거슬러 올라가는 이탈리아 오록스 표본의 미토콘드리아 DNA 서열을 분석한 결과, 남부 유럽 소에서 오록스 유전자가 어느 정도 유전자 침투했음이 밝혀졌다. 이는 암컷 오록스가 자유롭게 방목되는 가축과 접촉했음을 나타낸다.[99] 쿠트나 호라 구의 석기 시대 정착지에서 발견된 다양한 크기의 소 뼈는 기원전 3000년에서 2800년 사이에 보헤미아 지역에서 오록스와 가축의 잡종화가 이루어졌음을 보여주는 추가 증거이다.[42]
영국에서 발견된 6,750년 된 오록스 뼈의 전체 게놈 시퀀싱을 81마리 소의 게놈 서열 데이터 및 1,225마리 소의 단일염기 다형성 데이터와 비교한 결과, 영국 및 아일랜드 소 품종이 오록스 표본과 일부 유전적 변이를 공유한다는 사실이 밝혀졌다. 이는 영국 초기 목축업자들이 오록스에서 영국 및 아일랜드 소의 조상으로 지역 유전자 흐름을 담당했을 수 있음을 시사한다.[100] 무르보덴 소 품종은 또한 알프스에서 암컷 유럽 오록스가 가축으로 산발적인 유전자 침투를 보였다.[101] 가축은 중세 시대까지 몸 크기와 뿔 크기가 계속해서 작아졌다.[86]
단일염기 다형성과 공유된 대립 유전자의 비교 분석 결과, 고대 중국에서 동아시아 오록스와 도입된 털 소 사이에 혼합이 발견되었으며, 예를 들어 시마오에서 그러한 흔적이 발견되었다. 이는 자발적인 유전자 침투 또는 가축의 수를 늘리기 위해 다른 오록스 그룹을 포획하는 것을 통해 최대 4,000년 전부터 지역 오록스가 가축에 통합되었음을 시사한다. 같은 연구에서는 특히 티베트 고원 품종에서 동아시아의 오록스와 현대 털 소가 공유하는 파생된 대립 유전자가 감지되었다. 지역 오록스와의 유전자 침투는 새로운 환경에 대한 빠른 적응을 촉진했을 수 있다.[2]
인도 오록스는 10,000~8,000년 전에 가축화된 것으로 생각된다.[102] 파키스탄 메르가르 신석기 유적에서 발견된 오록스 화석은 약 8,000년 전으로 거슬러 올라가며, 인도 아대륙에서의 가축화에 대한 가장 초기의 증거 중 일부를 나타낸다.[103] 암컷 인도 오록스는 북부 인도에서 목축이 확장되는 동안 기원전 5,500년에서 4,000년 사이에 제부(''Bos indicus'')의 유전자 풀에 기여했다. 제부는 처음에 동쪽으로 동남아시아로 퍼져나갔다.[104] 기원전 4,000년 이후 근동에서 제부와 초기 털 소 사이의 잡종화가 4.2킬로년 사건 기간의 가뭄과 일치하여 발생했다.[105] 제부는 약 3,500~2,500년 전에 동아프리카에 도입되었고,[97] 13~14세기에 몽골에 도달했다.[106]
이집트 서부 사막에서 발생한 것으로 여겨지는 세 번째 가축화 사건은 180개 개체를 대표하는 3,196마리의 가축의 유전자 혼합, 유전자 침투 및 이동 패턴 분석 결과로 뒷받침되지 않는다. 그러나 동일한 연구에서는 가축화 이후 근동에서 도착한 아프리카의 털 소와 아프리카의 제부가 아프리카에 들어오기 전에 지역 야생 아프리카 오록스 사이의 광범위한 잡종화를 뒷받침했다.[97] 제부는 고대 이집트를 통해 도입되었고, 아랍의 문화적 영향과 함께 지난 1,400년 동안 서아프리카 전역으로 포괄적으로 확산되기 시작했다. 대부분의 현대 아프리카 소 품종은 제부 소와 가변적인 정도로 잡종화되어 있으며, 체체파리가 있는 서아프리카 지역에서 유전자 침투가 가장 감소했다.[107]
9. 멸종
염기 서열 분석 결과 오록스의 게놈은 16,338개의 염기쌍으로 구성되어 있음이 밝혀졌다.[45] 2024년 연구에 따르면 인도 오록스는 다른 오록스 집단에서 약 300,000~166,000년 전에 분기되었고, 유럽, 중동, 동아시아 오록스 집단은 100,000년 이내에 공통 조상을 공유했다. 후기 홍적세 유럽 오록스는 인도 오록스 및 기타 오록스 분기 이전에 분기된 계통으로부터 약간의 조상 성분을 가졌고, 후기 홍적세 말기에 중동 오록스로부터 상당한 유전자 흐름을 겪었다. 유럽 홀로세 오록스는 주로 최후 빙기 동안 이베리아 반도에 존재했던 오록스의 후손이며, 홀로세 동안 이전에 고립되었던 오록스 집단 간의 혼합도 관찰되었다.[47]
인도 오록스 (''B. p. namadicus'')는 홀로세 기간 중 목축 확장으로 인한 서식지 손실과 가축 제부와의 잡종 교배로 인해 멸종되었다.[104][108] 인도 아대륙에서의 멸종 시기는 불분명하며, 2021년 검토에 따르면 파키스탄 메르가르에서 약 8,000년 전 유해가 홀로세 인도 오록스의 "유일하게 연대 측정되고 신뢰할 수 있게 확인된 증거"일 수 있다고 한다.[103]
아프리카 오록스 (''B. p. mauritanicus'')의 가장 젊은 유해는 약 6,000년 현재 이전 (BP)으로 추정되지만,[110] 일부 저자는 로마 시대까지 생존했을 수 있다고 주장하며, 이는 나일 삼각주의 부토와 파이윰에서 발견된 유해로 확인된다.[43]
중국에서는 오록스가 적어도 3,600 BP까지 생존했다.[111] 유라시아 오록스 (''B. p. primigenius'')는 홀로세 기후 최적기 동안 적어도 7,800년 BP까지 스웨덴 남부에 존재했다.[112] 덴마크에서는 최초로 알려진 오록스의 지역 멸종이 약 8,000–7,500년 BP에 새로 형성된 덴마크 섬에서 해수면 상승 이후 발생했으며, 마지막으로 기록된 오록스는 약 3,000년 BP에 유틀란트 남부에 살았다.[36] 영국에서 가장 늦게 발견된 오록스 화석은 3,245년 BP로, 약 3,000년 전에 멸종되었을 것이다.[113]
과도한 사냥이 시작되어 오록스가 거의 멸종될 때까지 계속되었다. 중앙 유럽에서 오록스의 점진적인 멸종은 9세기에서 12세기 사이에 대규모 숲 벌채와 일치했다.[48] 13세기까지 오록스는 동유럽에 소수만 남아 있었으며, 사냥은 귀족, 나중에는 왕족의 특권이 되었다.[25] 헝가리의 개체수는 적어도 9세기부터 감소했으며 13세기에 멸종되었다.[114][115]
아화석 기록에 따르면, 야생 오록스는 트란실바니아 북서부에서는 14세기에서 16세기까지, 몰다비아 서부에서는 아마도 17세기 초까지 생존했을 수 있다.[116][117] 마지막으로 알려진 오록스 무리는 폴란드 야크토루프 숲의 습지 숲에 살았다. 16세기 중반 약 50마리에서 1601년에는 4마리로 감소했다. 마지막 오록스 암컷은 1627년 자연사했다.[118]
2021년 연구에 따르면 오록스는 적어도 17세기까지 불가리아 북동부에서 생존했을 가능성이 있다.[119] 2020년 소피아에서 발굴된 뿔 심이 오록스에서 나온 것으로 확인되었으며, 이 뿔 심이 발견된 고고학적 층은 17세기 후반 또는 18세기 전반으로 연대가 측정되어 오록스가 그 시기까지 불가리아에서 생존했을 가능성을 시사한다.[120]
오록스는 한때 유라시아 전체와 북아프리카에서 발견되었지만, 서식지 감소, 남획, 가축화 등으로 인해 멸종되었다. 남아시아에서는 역사 시대의 비교적 초기에 사라졌으며, 메소포타미아에서도 페르시아 제국 성립 무렵에는 이미 멸종된 것으로 보인다. 북아프리카에서도 고대 이집트의 종말과 동시에 사라졌다. 중세에는 이미 현재의 프랑스, 독일, 폴란드 등의 숲에서만 발견되었다. 16세기에는 각지에 오록스 사냥 금지 구역이 생겼지만, 이는 제후들이 자신의 취미나 유흥을 위해 사냥할 몫을 확보하기 위해 설치한 것에 불과했기 때문에 사냥감을 모두 잡고 폐쇄되었다. 마지막으로 남은 것은 폴란드 바르샤바 근교의 야크토루프(Jaktorów) 보호 구역이었지만, 밀렵으로 인해 개체 수가 계속 줄어들어 1620년에는 마지막 1마리만 남게 되었다. 그 1마리마저 1627년에 죽음이 확인되어 오록스는 멸종했다.
10. 복원 시도
1920년대 초, 하인츠 헤크는 여러 소 품종을 사용하여 헬라브룬 동물원에서 오록스를 되살리는 번식 프로그램을 시작했다. 그 결과는 헤크 소라고 불린다.[121]
1980년대에 헤크 소는 네덜란드의 간척지인 오스트바르데르플라센에 자연 방목을 위한 오록스 대용으로 방목되었는데, 이는 선사 시대의 풍경을 복원하려는 목적이었다.[122] 2005년과 2010년의 혹독한 겨울 동안 많은 수가 굶어 죽었고, 무간섭 프로젝트는 2018년에 종료되었다.[123]
1996년부터, 헤크 소는 사야게사 소, 키아니나 소, 그리고 소규모로 스페인 투우와 교배되어 더 오록스에 가까운 동물을 만들기 위한 노력이 이루어졌다. 그 결과로 태어난 잡종은 타우루스 소라고 불린다.[124] 타우로스 프로그램과 우루즈 프로젝트는 또 다른 되살림 프로젝트들이다.[122] 그러나 오록스 유사 표현형을 목표로 하는 접근 방식은 오록스 유사 유전자형과 같지 않다.[125]
이후, 1920년대부터 독일인 형제 Lutz Heck|루츠 헤크영어(1892년-1983년)와 Heinz Heck|하인츠 헤크영어(1894년-1982년)는 베를린 및 뮌헨의 동물원에서 현존하는 소 중에서 오록스에 가까운 특징을 가진 것들을 교배시켜 오록스의 모습을 되살리려고 시도했다.[136] 1932년에 작출은 성공했으며, 그 개체의 자손은 현재도 독일의 동물원에서 사육·전시되고 있다. 이 소는 체형이나 성질은 오록스와 비슷하지만 체격은 다소 작으며, 작출에 관여한 당시 동물원장 루츠 헤크(독일인 동물학자)의 성을 따서 "Heck cattle|헤크 캐틀영어"이라고도 불린다. 최근에는 네덜란드에서 유전 정보와 근사종의 교배를 통해 복원하려는 연구가 진행되고 있다.[137]
참조
[1]
IUCN
Bos primigenius
2024-09-27
[2]
서적
Origins: A Short Etymological Dictionary of Modern English
Greenwich House
[3]
서적
A Latin Dictionary
Clarendon Press
[4]
서적
The Gallic Wars by Julius Caesar
Harper & Brothers
[5]
서적
The Cambridge Encyclopedia of the English Language
Cambridge University Press
[6]
서적
Resurrecting Extinct Species
Palgrave Macmillan
[7]
서적
Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis
Laurentii Salvii
1758
[8]
학술지
De Uro nostrate eiusque sceleto commentation
https://archive.org/[...]
[9]
학술지
Corrected date of the first description of aurochs Bos primigenius (Bojanus, 1827) and steppe bison Bison priscus (Bojanus, 1827)
2019
[10]
학술지
Notice of the various species of bovine animals
https://archive.org/[...]
[11]
학술지
Recherches sur les bovidés fossiles de l'Algérie
https://gallica.bnf.[...]
[12]
학술지
Opinion 2027 (Case 3010). Usage of 17 specific names based on wild species which are pre-dated by or contemporary with those based on domestic animals (Lepidoptera, Osteichthyes, Mammalia)
https://archive.org/[...]
[13]
학술지
The naming of wild animal species and their domestic derivatives
https://www.research[...]
2004
[14]
학술지
Modern Taurine Cattle descended from small number of Near-Eastern founders
[15]
서적
In the Light of Evolution
[16]
학술지
Evolution and legacy of East Asian aurochs
2024-11-15
[17]
간행물
Excavations of 2003 at the S. Teodoro cave (North-Eastern Sicily, Italy): preliminary faunistic and stratigraphic data
[18]
간행물
Le faune a mammiferi della Sicilia tra il Tardiglaciale e l'Olocene
[19]
학술지
Insular aurochs (Mammalia, Bovidae) from the Pleistocene of Kythera Island, Greece
[20]
학술지
A multi-calibrated mitochondrial phylogeny of extant Bovidae (Artiodactyla, Ruminantia) and the importance of the fossil record to systematics
[21]
학술지
Incomplete lineage sorting rather than hybridization explains the inconsistent phylogeny of the wisent
[22]
학술지
Tracking of wisent–bison–yak mitochondrial evolution
[23]
학술지
A multi-calibrated mitochondrial phylogeny of extant Bovidae (Artiodactyla, Ruminantia) and the importance of the fossil record to systematics
2013
[24]
학술지
Kouprey (Bos sauveli) genomes unveil polytomic origin of wild Asian Bos
2021
[25]
서적
Retracing the Aurochs: History, Morphology and Ecology of an extinct wild Ox
Pensoft Publishers
[26]
학술지
Bos primigenius Bojanus, 1827 (Mammalia, Bovidae) in Greece: new finds and a revision of the species, with a comparison with body-size variations of aurochs from the Italian Peninsula
[27]
학술지
The early Middle Pleistocene archeopaleontological site of Wadi Sarrat (Tunisia) and the earliest record of Bos primigenius
[28]
서적
Géologie et paléontologie du gisement acheuléen de l'erg Tihodaïne, Ahaggar Sahara Algérien
Memoires du centre de recherches anthlropologiques, prehistoriques et ethnographiques
[29]
학술지
The evolution of the buffaloes, oxen, sheep and goats
[30]
학술지
The Punta Lucero Quarry site (Zierbena, Bizkaia): a window into the Middle Pleistocene in the Northern Iberian Peninsula
2015
[31]
학술지
Climatic and environmental changes of ~100 thousand years: The mammals from the early Middle Pleistocene sequence of Notarchirico (southern Italy)
2024
[32]
학술지
Redefining Ponte Molle (Rome, central Italy): an important locality for Middle Pleistocene mammal assemblages of Europe
2021
[33]
학술지
Terrestrial environments during MIS 11: evidence from the Palaeolithic site at West Stow, Suffolk, UK
2007
[34]
학술지
New fossils of Bos primigenius (Artiodactyla, Mammalia) from Nihewan and Longhua of Hebei, China
[35]
학술지
Wild and Domestic Cattle in the Ancient Nile Valley: Marks of ecological change
[36]
논문
Ancient DNA extracted from Danish aurochs (''Bos primigenius''): genetic diversity and preservation
[37]
논문
Sigmund von Herberstein (1486–1566) und die historischen Konfusionen um Ur und Wisent
https://www.zobodat.[...]
[38]
서적
The ox and its kindred
Methuen &Co. Ltd.
[39]
논문
Update to: "Some late sixteenth-century depictions of the aurochs (''Bos primigenius'' Bojanus, extinct 1627): New evidence from Vatican MS Urb. Lat. 276
https://www.euppubli[...]
[40]
논문
The first hair remains from an aurochs (''Bos primigenius'') and some medieval domestic cattle hair
[41]
서적
Der Auerochs: Das europäische Rind
Lipp Graphische Betriebe
[42]
논문
Aurochs and potential crossbreeding with domestic cattle in Central Europe in the Eneolithic period. A metric analysis of bones from the archaeological site of Kutná Hora-Denemark (Czech Republic)
https://www.research[...]
[43]
논문
Size and size change of the African aurochs during the Pleistocene and Holocene
https://www.research[...]
[44]
논문
History, morphology and ecology of the Aurochs (''Bos primigenius'')
2002
[45]
논문
A complete mitochondrial genome sequence from a mesolithic wild Aurochs (''Bos primigenius'')
[46]
논문
Genome-wide microRNA binding site variation between extinct wild Aurochs and modern cattle identifies candidate microRNA-regulated domestication genes
2017
[47]
논문
The genomic natural history of the aurochs
2024
[48]
서적
Mlekopitajuščie Sovetskogo Soiuza. Moskva: Vysšaia Škola
Smithsonian Institution and the National Science Foundation
[49]
논문
A record of ''Bos primigenius'' from the Quaternary of the Aba Tibetan Autonomous Region
http://www.nau.edu/~[...]
1984
[50]
논문
Ancient DNA reveals evidence of abundant aurochs (''Bos primigenius'') in Neolithic Northeast China
http://bjkg.jlu.edu.[...]
[51]
서적
Mammals of Korea
Magnolia Press
[52]
논문
モノが語る牛と人間の文化 - ② 岩手の牛たち
http://liaj.lin.gr.j[...]
2016-04-06
[53]
논문
First record of Late Pleistocene Bison from the fissure deposits of the Kuzuu Limestone, Yamasuge, Sano-shi, Tochigi Prefecture, Japan
http://www.gmnh.pref[...]
2016-04-06
[54]
논문
A comparative analysis of the habitat of the extinct aurochs and other prehistoric mammals in Britain
2008
[55]
논문
Die Großtierfauna Europas und ihr Einfluss auf Vegetation und Landschaft
[56]
논문
Feeding strategy of the Urus ''Bos primigenius'' Bojanus, 1827 from the Holocene of Denmark
https://www.research[...]
[57]
논문
Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain
2016
[58]
논문
New insights into the Middle Pleistocene paleoecology and paleoenvironment of the Northern Iberian Peninsula (Punta Lucero Quarry site, Biscay): A combined approach using mammalian stable isotope analysis and trophic resource availability modeling
2017
[59]
논문
A spotted hyaena den in the Middle Palaeolithic of Grotta Paglicci (Gargano promontory, Apulia, Southern Italy)
2016
[60]
논문
The Pleistocene easternmost distribution in Eurasia of the species associated with the Eemian ''Palaeoloxodon antiquus'' assemblage
2007
[61]
논문
The faunal background of the stone age cultures of Hunsgi and Baichbal Valleys, Southern Deccan
[62]
논문
Early evidence for symbolic behavior in the Levantine Middle Paleolithic: A 120 ka old engraved aurochs bone shaft from the open-air site of Nesher Ramla, Israel
[63]
논문
Early evidence (ca. 12,000 B.P.) for feasting at a burial cave in Israel
2010
[64]
논문
Gobustan: Rock Art Cultural Landscape
https://www.rockarts[...]
[65]
논문
Private pantries and celebrated surplus: storing and sharing food at Neolithic Çatalhöyük, Central Anatolia
https://www.research[...]
[66]
뉴스
Et si Europe était sidonienne?
http://www.lorientle[...]
2020-01-03
[67]
서적
The Indus civilization
Lovat Dickson & Thompson Ltd.
[68]
서적
Animals in stone: Indian mammals sculptured through time
Brill
[69]
서적
Appropriating Innovations. Entangled Knowledge in Eurasia, 5000-150 BCE
Oxbow Books
[70]
서적
Ritual, Religion, and Reason. Studies in the Ancient World in Honour of Paolo Xella
Ugarit-Verlag
2013
[71]
논문
Foundations of the Earth
Columbia University Press
2014
[72]
논문
First evidence of Pleistocene rock art in North Africa: securing the age of the Qurta petroglyphs (Egypt) through OSL dating
https://www.academia[...]
[73]
논문
''Bos primigenius'' in Ancient Egyptian art – historical evidence for the continuity of occurrence and ecology of an extinct key species
https://epub.uni-bay[...]
[74]
논문
ESR/U-series dating of faunal remains from the paleoanthropological site of Biache-Saint-Vaast (Pas-de-Calais, France)
2015
[75]
논문
Néandertal et le feu au Paléolithique moyen ancien. Tour d'horizon des traces de son utilisation dans le Nord de la France
https://hal.archives[...]
2013
[76]
논문
A symbolic Neanderthal accumulation of large herbivore crania
2023
[77]
논문
From Chauvet to Lascaux: 15,000 years of cave art
[78]
논문
The hunting of large mammals in the Upper Palaeolithic of southern Italy: A diachronic case study from Grotta del Romito
https://www.research[...]
[79]
논문
Palaeolithic art at Grotta di Cala dei Genovesi, Sicily: a new chronology for mobiliary and parietal depictions
https://www.research[...]
[80]
논문
Archäologie und Biologie des Aurochsen
Neanderthal Museum
[81]
논문
Aesthetics and Rock Art III: Symposium
Archaeopress
[82]
논문
Analysis of portable rock art from Foz do Medal (Northwest Iberia): Magdalenian images of horses and aurochs
[83]
논문
Late Mesolithic hunting of a small female aurochs in the valley of the River Tjonger (the Netherlands) in the light of Mesolithic aurochs hunting in NW Europe
[84]
논문
Settlements of coastal Estonia and maritime hunter-gatherer economy
https://www.research[...]
[85]
논문
Where the wild things are: Aurochs and Cattle in England
https://www.research[...]
[86]
논문
On the origin of cattle: How Aurochs became cattle and colonized the World
[87]
논문
The Vapheio Cups: One Minoan and One Mycenean?
[88]
논문
Hands and Tails on the Vapheio Cups
[89]
서적
Birds and Beasts of the Greek Anthology
B. Blom
[90]
서적
The National Cyclopaedia of Useful Knowledge
Little, Brown and Co.
[91]
서적
Das Nibelungenlied und die Klage: Nach der Handschrift 857 der Stiftsbibliothek St. Gallen
Deutscher Klassiker Verlag
[92]
논문
Ancient DNA analysis of Scandinavian medieval drinking horns and the horn of the last aurochs bull
[93]
논문
Cambridge and Cornelimünster
https://journals.ub.[...]
1972
[94]
논문
The comparative study of the state of conservation of two medieval documents on parchment from different historical periods
[95]
논문
The first steps of animal domestication: new archeological approaches
Oxbow Books
[96]
논문
Documenting the initial appearance of domestic cattle in the Eastern Fertile Crescent (northern Iraq and western Iran)
https://scholar.harv[...]
[97]
논문
Domestication of cattle: Two or three events?
[98]
논문
Cattle domestication in the Near East was followed by hybridization with aurochs bulls in Europe
[99]
논문
The origin of European cattle: Evidence from modern and ancient DNA
2006
[100]
논문
Genome sequencing of the extinct Eurasian wild aurochs, ''Bos primigenius'', illuminates the phylogeography and evolution of cattle
2015
[101]
논문
Large-scale mitogenome sequencing reveals consecutive expansions of domestic taurine cattle and supports sporadic Aurochs introgression
[102]
논문
Mitochondrial diversity and the origins of African and European cattle
[103]
논문
Late Quaternary megafaunal extinctions in India: How much do we know?
https://discovery.uc[...]
[104]
논문
Zebu Cattle are an exclusive legacy of the South Asia Neolithic
2010
[105]
논문
Ancient cattle genomics, origins, and rapid turnover in the Fertile Crescent
https://eprints.whit[...]
[106]
논문
Independent mitochondrial origin and historical genetic differentiation in North Eastern Asian cattle
http://webpages.icav[...]
2021-12-22
[107]
웹사이트
Genetic characterization and West African cattle
http://www.fao.org/3[...]
2021-09-20
[108]
서적
India's Wildlife History
Permanent Black
[109]
서적
Aurochs Bos primigenius Bojanus, 1827
https://www.cambridg[...]
Cambridge University Press
2024-11-02
[110]
논문
Late Pleistocene and Holocene mammal extinctions on continental Africa
2014-01-01
[111]
논문
Evolution and legacy of East Asian aurochs
2024-09-01
[112]
서적
Climate Change and Human Responses
Springer
[113]
논문
Five thousand years of livestock in Britain
[114]
논문
Interdisciplinary analysis of an Iron Age Aurochs horn core from Hungary: a case study
[115]
논문
A horn worth blowing? A stray find of Aurochs from Hungary
https://www.research[...]
[116]
논문
Holocene subfossil records of the auroch (''Bos primigenius'') in Romania
[117]
논문
Holocene mammal extinctions in the Carpathian Basin: a review
http://real.mtak.hu/[...]
2017-01-01
[118]
논문
History of the Aurochs (''Bos taurus primigenius'') in Poland
http://agtr.ilri.cgi[...]
[119]
논문
Subfossil vertebrate fauna from Forum Serdica (Sofia, Bulgaria), 16–18th Century AD
https://www.research[...]
[120]
논문
The last ''Bos primigenius'' survived in Bulgaria (Cetartiodactyla: Bovidae)
[121]
논문
The breeding-back of the Aurochs
[122]
논문
From "Nazi cows" to cosmopolitan "ecological engineers": specifying rewilding through a history of Heck cattle
[123]
논문
The Oostvaardersplassen Fiasco
[124]
논문
Ecological substitutes for Wild Horse (''Equus ferus'', Boddaert 1785 = ''E. przewalskii'', Poljakov 1881) and Aurochs (''Bos primigenius'', Bojanus 1827)
[125]
논문
The draft genome of extinct European Aurochs and its implications for de-extinction
[126]
IUCN
Bos primigenius
[127]
논문
Opinion 2027 (Case 3010): Usage of 17 specific names based on wild species which are pre-dated by or contemporary with those based on domestic animals (Lepidoptera, Osteichthyes, Mammalia): conserved
https://www.biodiver[...]
2003-03-31
[128]
논문
Phylogeny of bovine species based on AFLP fingerprinting
[129]
논문
Early cave art and ancient DNA record the origin of European bison
https://www.nature.c[...]
2016-10-18
[130]
웹사이트
The Higgs Bison - mystery species hidden in cave art
https://www.adelaide[...]
The University of Adelaide
2017-01-13
[131]
논문
Genome data on the extinct Bison schoetensacki establish it as a sister species of the extant European bison (''Bison bonasus'' )
2017-01-01
[132]
논문
Hunting the Extinct Steppe Bison (''Bison priscus'' ) Mitochondrial Genome in the Trois-Frères Paleolithic Painted Cave
2015-06-17
[133]
웹사이트
Bos primigenius
https://www.nhm.uio.[...]
Naturhistorisk museum
2011-05-18
[134]
논문
モノが語る牛と人間の文化 ②岩手の牛たち
http://liaj.lin.gr.j[...]
家畜改良事業団
2008-03-25
[135]
간행물
中国黒竜江省ハルビン市周辺のマンモス動物群を訪ねて-中国東北地域の後期更新世哺乳動物群から日本のマンモス動物群を考える
http://www.kasekiken[...]
化石研究会
[136]
웹사이트
知られざる歴史:ナチスは絶滅種の動物を復元するプロジェクトを行い、それを成功させていた。
http://karapaia.com/[...]
2020-07-31
[137]
웹사이트
野生絶滅から1世紀、欧州のバイソン再野生化へ:オランダで4頭を野生復帰、現代に調和した新しい「野生」目指す
https://natgeo.nikke[...]
日経ナショナル ジオグラフィック社
2016-03-28
[138]
논문
Bos primigenius
http://www.iucnredli[...]
IUCN
2018-01-11
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com