초항원
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
초항원은 세균이 생성하여 분비하는 독소로, 면역 체계를 과도하게 자극하여 다양한 질병을 유발한다. 이들은 MHC II형 분자에 결합한 후 T 세포 수용체와 결합하여 T 세포의 활성화를 유도하며, 과도한 사이토카인 방출로 인한 사이토카인 폭풍, T 세포의 무반응 또는 소실을 초래한다. 초항원은 또한 염증 반응을 유발하고, 내독소의 효과를 증폭시키며, 당뇨병, 독성 쇼크 증후군 등 다양한 질병과 관련이 있다. 치료는 미생물 제거, 항체 생성 유도, 면역 억제제 사용 등을 포함하며, 초항원 생산은 면역 회피를 위한 진화적 전략으로 여겨진다. 일부 바이러스는 내인성 초항원을 활용하여 면역 체계를 회피하기도 한다.
더 읽어볼만한 페이지
- 독물학 - 중독
중독은 유해 물질 노출로 신체적, 정신적 건강에 해로운 영향을 미치는 상태로, 급성·만성, 외인성·내인성으로 분류되며, 다양한 물질이 원인이 될 수 있으므로 예방을 위해 안전 수칙 준수와 응급 처치가 중요하다. - 독물학 - 중금속
중금속은 과학적 정의가 불분명한 용어로, IUPAC는 모호성과 오해의 소지를 지적하며 독성 및 환경 오염을 유발하고 미나마타병, 이타이이타이병과 같은 중독 사례가 있는 물질이다. - 면역계 - 알레르기
알레르기는 면역 체계가 무해한 물질에 과도하게 반응하여 아토피 피부염, 비염, 천식, 두드러기 등 다양한 증상을 일으키는 질환으로, Gell과 Coombs 분류법에 따라 여러 유형으로 나뉘며, 심각한 경우 아나필락시스를 유발할 수 있고, 유전적, 환경적 요인이 복합적으로 작용하여 발생하며, 알레르겐 회피, 약물, 면역요법 등으로 치료한다. - 면역계 - 면역억제
면역억제는 면역계의 활동을 감소시키는 현상으로, 의도적으로는 장기 이식 거부 반응 방지나 자가면역 질환 치료를 위해 면역억제제를 사용하고, 비의도적으로는 질병으로 인해 발생하며 면역결핍질환을 유발할 수 있다.
| 초항원 |
|---|
2. 구조
초항원은 세균이 세포 내에서 생성하며 감염 시 성숙한 독소 형태로 세포 외부로 분비된다.[54] 여러 초항원 하위 그룹 간에 염기서열은 비교적 잘 보존되어 있으며, 이보다 더 중요한 것은 3차원 구조가 매우 유사하여 기능적인 효과도 유사하다는 점이다.[55][56]
초항원은 먼저 MHC 클래스 II 분자에 결합한 후, T 세포 수용체의 가변 α 사슬 또는 β 사슬과 상호작용한다.[58][56][59] 장독소의 결정 구조는 이들이 특징적인 2개의 도메인 접힘 패턴을 공유하는, 콤팩트하고 타원체 형태의 단백질임을 보여주며, 각 도메인은 주요 조직 적합성 복합체 II (MHC class II) 및 T 세포 수용체 (TCR)에 대한 결합 부위를 갖는다.[8] 초항원은 이 둘을 연결하여 비특이적인 활성화를 유발한다.
장내독소의 결정 구조는 이들이 특징적인 두 개의 도메인 접힘 패턴을 공유하는 작고 타원체 형태의 단백질임을 보여준다. 이들은 각각 MHC 클래스 II 분자와 T 세포 수용체(TCR)에 대한 결합 부위를 갖는다.[57] 이 패턴은 NH2-말단 β 배럴 구형 도메인 (올리고당/올리고뉴클레오티드 접힘으로 알려져 있음), 분자의 중심을 대각선으로 가로지르는 긴 α-나선 및 COOH-말단 구형 도메인으로 구성된다.[5]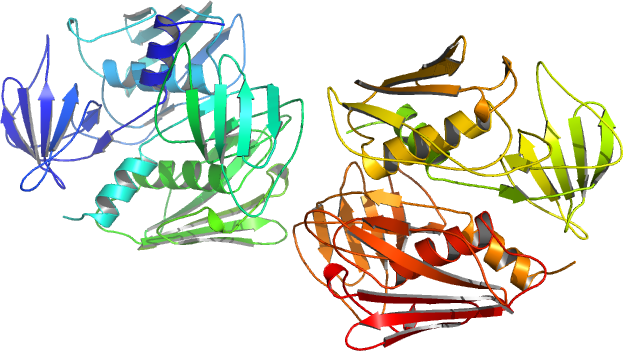
3. 결합
초항원 중 T 세포와 결합하는 영역은 T 세포 수용체의 β 사슬과 상호 작용을 일으킨다. 초항원은 TCR의 β 사슬에 있는 V 절편에 결합하는데, 이 절편은 약 50 종류 밖에 없고, 일부 초항원이 다수의 V 절편에 결합할 수 있기 때문에 1 종류의 초항원으로도 상당히 많은 수의 T 세포를 활성화할 수 있다.
TCR 영역은 개인마다 차이가 있어, 특정 초항원에 대해 일부 사람만 더 강하게 반응하는 경우가 발생한다.
3. 1. MHC Class II
초항원은 HLA-DQ 형태의 분자에 대한 선호도를 보인다.[59] α 사슬에 결합하면 초항원이 T 세포 수용체와 반응할 수 있는 적절한 위치에 놓이게 된다.[56] 드물게, 초항원은 아연 이온 매개 반응을 통해 다형성 MHC 클래스 II의 β 사슬에 결합하는데, 이 결합은 매우 보존된 HLA-DR β 사슬과 세 개의 초항원 잔류물 사이에 이루어진다.[55][56] 결합에서 아연 이온을 사용하면 높은 친화도 반응이 일어난다. 몇몇 포도상구균 초항원은 MHC 분자의 α 사슬과 β 사슬 모두에 결합하여 가교 결합을 형성할 수 있다.[56] 이 기작은 항원 제시 세포에서 사이토카인 발현과 분비를 자극하며, T 세포에 더욱 효과적으로 결합하여 T 세포를 활성화시키는 상호 자극적인 역할을 하는 분자의 생산을 자극한다.[56]
3. 2. T-세포 수용체
초항원의 T 세포 결합 부위는 T 세포 수용체 베타 사슬(β chain)에 있는 가변 부위(Vβ 부위)와 상호 작용한다.[8] 인간 T 세포는 약 50가지 유형의 Vβ 요소로 구성되어 있으며, 일부 초항원은 여러 유형의 Vβ 부위에 결합할 수 있어 T 세포 집단의 상당 부분을 활성화할 수 있다. 이러한 상호작용은 초항원의 그룹에 따라 약간씩 다르다.[8] 사람들마다 T 세포 부위 유형의 차이가 있어, 특정 초항원에 대해 더 강하게 반응하는 사람도 있다.
그룹 I 초항원은 CDR2와 프레임워크 영역에서 Vβ에 접촉한다.[11][12] 그룹 II 초항원은 컨포메이션에 의존하는 메커니즘을 사용하여 Vβ 부위와 상호 작용하는데, 대부분 특정 Vβ 아미노산 측쇄와 관련이 없다. 그룹 IV 초항원은 특정 Vβ 형태의 세 개의 CDR 루프 모두에 관여하는 것으로 나타났다.[11][12]
이러한 상호 작용은 초항원의 작고 큰 도메인 사이의 틈에서 일어나며, 초항원이 TCR과 MHC 사이의 쐐기 역할을 할 수 있게 한다. 이는 항원성 펩티드를 TCR에서 멀어지게 하고 T 세포 활성화의 정상적인 메커니즘을 우회한다.[6][13]
초항원의 생물학적 강도(자극 능력)는 TCR에 대한 친화도에 의해 결정된다. TCR에 대한 친화도가 가장 높은 초항원이 가장 강한 반응을 유도한다.[14] SPMEZ-2는 현재까지 발견된 가장 강력한 초항원이다.[14]
4. T-세포 신호전달
초항원은 MHC와 TCR에 가교결합하여 신호전달체계를 유도한다. 그 결과로 세포의 증식과 사이토카인 생산이 일어난다. 초항원에 의해 활성화된 T세포에서 낮은 농도의 Zap-70는 정상적인 T세포 활성화 신호전달 체계가 제 기능을 못함을 보여준다.[64]
티로신 키나아제에 의해 Lck 보다는 Fyn이 활성화되어 후천적인 면역성결핍 (anergy)의 유도가 이루어진다는 가설이 있다.[65]
단백질인산화효소C 경로와 단백질 타이로신 키나아제 경로가 모두 활성화되면, 염증전 사이토카인의 생성이 상향조절된다.[66]
이런 대안적 신호전달 경로는 칼슘/칼시뉴린 과 Ras/MAPkinase 경로를 약하게 악화시키지만 집중된 면역 반응을 위해 감안한다.[65]
5. 직접적 효과
항원제시세포와 T 세포의 초항원 자극은 주로 Th1 도움 T 세포의 반응에 초점을 둔 염증 반응을 유발한다. 주요 생성물은 IL-1, IL-2, IL-6, TNF-α, 인터페론 감마(IFN-γ), 대식세포 염증 단백질 1α (MIP-1α), MIP-1β, 단핵구 화학유인 단백질 1 (MCP-1) 등이 있다.[66]
이 과도하게 통제되지 않는 사이토카인(특히 TNF-α)의 분비는 신체에 부담을 주어 발진, 발열, 그리고 심한 경우 다중기관 부전, 혼수상태나 사망에 이르게 한다.[59][61]
지속적인 독소 노출에 의해 IL-4와 IL-10이 분비되며, 이에 따른 결과로 활성화 T 세포의 무반응 또는 결실이 나타나 감염 상태에 이른다. IL-4와 IL-10은 인터페론 감마, MHC 클래스 II, 항원제시세포 표면에 상호자극하는 역할을 하는 분자들의 생산을 하향 조절한다. 이러한 효과들은 항원의 자극에 반응하지 않는 기억 세포를 만들게 한다.[67][68]
이러한 효과의 가능한 한 가지 방법은 사이토카인 매개 T 세포 억제와 연관된다. MHC 교차결합은 또한 신호 전달 체계를 활성화시켜 혈액 생성을 억제하고 Fas-매개 세포자멸사(apoptosis)를 상향조절한다.[69]
지속적인 초항원 노출에 따른 또 다른 결과는 인터페론 알파(IFN-α)의 생성이다. 이 사이토카인은 자가면역 유도와 밀접하게 관련되어 있으며,[70] 가와사키병 같은 자가면역 질병은 초항원의 감염에 의한 것으로 알려져 있다.[63]
T 세포의 초항원 활성화는 CD40 리간드의 생성을 일으켜 B 세포에서 IgG, IgM, IgE로의 동기준표본변환(isotype switching)을 활성화시킨다.[71]
요약하자면, T 세포는 자극을 받아 과도한 양의 사이토카인을 생성하고, 그 결과 신체에서는 항상성을 회복하기 위해 사이토카인 매개 T 세포 억제와 활성화된 세포의 삭제가 일어난다. 미생물과 초항원에 의한 독성 효과는 조직과 기관에 피해를 주는 독성 쇼크 증후군과 같은 상태를 유발한다.[71] 초기 염증에서 생존하면 숙주 세포는 아네르기 상태가 되거나 삭제되어 심각한 면역 손상을 일으킨다.
6. 초항원의 항원성과는 독립적(간접적)인 효과
초항원은 세포분열 능력 외에도 감염의 특징적인 증상을 유발할 수 있다.[53], [2]
초항원에 의한 자극은 항원 제시 세포나 T세포 (주로 Th1 헬퍼 T 세포 중심의 염증 반응)를 유도한다. 이 과정에서 IL-1, IL-2, IL-6, TNF-α, IFN-γ, 대식세포 염증성 단백질(MIP-1α와 MIP-1β), 단핵구 주화 인자(MCP-1) 등이 생성된다.[47]
이러한 사이토카인, 특히 TNF-α의 무질서한 방출은 "'''사이토카인 방출 증후군'''"(사이토카인 폭풍)이라고 불리며, 전신에 과도한 부담을 주고 발진, 발열을 일으키며, 심한 경우 다기관 부전, 혼수, 사망에 이르게 한다.[38],[39]
초항원에 장기간 노출되면 IL-10 생성이 증가하여 활성화된 T 세포 결핍이나 에너지를 유발하여 감염으로 이어진다. IL-10은 IL-2나 MHC 클래스 II 분자, 항원 제시 세포 표면의 공동자극 분자 생산을 억제한다. 이러한 영향으로 항원 자극에 반응할 수 없는 기억 세포만 남게 된다.[48],[49]
이러한 면역 세포 불활성화는 IL-10과 같은 사이토카인을 통해 T 세포를 억제하거나, MHC 분자 가교가 조혈계를 억제하는 신호를 활성화하여 Fas를 통한 아폽토시스를 유발하는 방식으로 일어난다.[50] IFN-α도 장기간 초항원 노출의 산물이다. 이것은 자가 면역 관련 사이토카인이며,[51] 가와사키병 같은 자가 면역 질환은 초항원에 의해 유발된다고 알려져 있다.[44]
초항원으로 T 세포가 활성화되면 CD40 리간드 생산이 유도되어 IgM, IgG, IgE로의 아이소타입 스위치(클래스 스위치)가 활성화된다.[52]
T 세포는 초항원에 의해 활성화되면 과도한 사이토카인을 분비하고, 음성 피드백으로 T 세포 억제 및 결손이 발생한다. 미생물 독소와 초항원은 조직, 장기 손상을 유발하는 독성 쇼크 증후군을 일으킨다.[52] 초기 염증 이후, 세포는 에너지나 결손을 일으켜 심각한 면역 부전을 초래한다.
6. 1. 구토
초항원은 감염의 특징적인 증상들을 일으키는데, 그 중 하나는 구토이다.[53] 이러한 효과는 식중독의 경우에 느껴진다. 이는 내열성이 강한 독소를 분비하는 초항원을 만드는 세균에 의해 발생한다. 분자 내에는 위장관계 독성을 나타내는 구별되는 부분이 있다.[53] 이러한 활동은 매우 강력하며, 초항원 20-35μg에 이르는 소량에도 구토를 유발할 수 있다.[59]6. 2. 염증 유발
초항원은 T 세포 자극과 독립적으로 감염 부위에 호중구 보충을 자극할 수 있다. 이는 초항원이 단핵구를 활성화시켜 사이토카인 TNF-α 분비를 자극하고, 백혈구를 감염 부위로 유인하는 부착 분자 발현을 증가시키기 때문이다. 그 결과 폐, 장 조직 등 세균이 대량 서식하는 곳에 염증이 발생한다.[72] 소량의 염증은 자연스럽고 유익하지만, 과도한 염증은 조직 파괴를 일으킬 수 있다.[72]6. 3. 내독소 효과 증폭
초항원은 신체 내독소의 효과를 증폭시키는 능력이 있다.[58] 이는 초항원 감염에 의해 유도된 면역 체계 효율 감소 때문일 수 있다. Schlievert는 초항원과 내독소를 함께 투여하면 그 효과가 최대 50,000배까지 증가하는 것을 입증했다.[9] 내독소와 초항원의 시너지 효과 외에도, 내독소와 초항원의 활성이 동시에 일어나는 "더블 히트(double hit)" 효과는 전형적인 세균 감염에 의한 것보다 훨씬 유해하다.[58] 이는 세균 감염이 있는 환자에서 초항원이 패혈증으로 진행되는 것과도 연관되어 있다.[71],[22]7. 초항원 생산과 관련된 질병들
다음은 초항원 생산과 관련된 질병 목록이다.
황색포도구균과 화농성 연쇄상구균은 대표적인 초항원 생성 세균으로, 20종류 이상의 다양한 초항원을 생산한다. 초항원 유전자는 대부분 플라스미드나 병원성 유전자의 이동성 세균 유전자 근처에 위치하며, 엔테로톡신 유전자군으로 알려진 오페론이 자주 발견된다. 초항원은 많은 수의 T세포 항원 수용체에 결합하여 사이토카인 생산을 촉진하고, 과도하게 활성화된 T세포의 아폽토시스를 유발하여 면역 억제를 초래한다.
초항원 자극은 항원 제시 세포나 T 세포 반응(주로 Th1 헬퍼 T 세포 중심 염증 반응)을 유도하며, IL-1, IL-2, IL-6, TNF-α, IFN-γ 등 다양한 사이토카인이 생성된다.[47] 특히 TNF-α의 과도한 방출은 "사이토카인 방출 증후군"(사이토카인 폭풍)을 일으켜 발진, 발열, 심하면 다기관 부전, 혼수, 사망에 이르게 한다.[38][39] 장기간 초항원 노출은 IL-10 생성을 유발, T세포 결핍 및 에너지를 초래하여 감염으로 이어질 수 있다.[48][49]
IFN-α는 초항원 장기 노출로 생성되는 자가 면역 관련 사이토카인으로,[51] 가와사키병 같은 자가 면역 질환을 유발할 수 있다.[44]
초항원으로 활성화된 T세포는 CD40 리간드 생성을 유도하여 IgM, IgG, IgE로의 아이소타입 스위치 (클래스 스위치)를 활성화한다.[52]
결론적으로, T세포는 초항원에 의해 활성화되어 과도한 사이토카인을 분비하고, 음성 피드백으로 T세포 억제 및 결손이 발생한다. 미생물 독소와 초항원은 조직, 장기 손상을 유발하는 독성 쇼크 증후군을 일으킨다.[52] 초기 염증 이후, 세포는 에너지나 결손으로 심각한 면역 부전에 빠진다.
8. 치료
의학적 치료의 기본적인 목표는 초항원을 생성하는 미생물을 없애는 것이다.[53] 이는 혈관 수축제, 수액 요법과 항생제의 사용을 통해 이루어진다.
신체는 자연적으로 몇몇 초항원에 대한 항체를 만들어 내며, 이러한 효과는 항생제의 B 세포 생성을 자극하여 증폭될 수 있다.[73]
면역 글로불린은 특정 항체를 중화시키거나 T 세포 활성화를 막는 것이 가능하다. 초항원과 유사하게 행동하는 합성 항체와 합성 펩타이드가 만들어져 왔다. 이들은 초항원과 MHC class II 분자의 초항원이 결합하는 부위에 결합해서 T 세포의 반응과 활성화를 막는다.[53]
면역 억제제 또한 T 세포의 활성화와 사이토카인 분비를 막기 위해 사용된다. 코르티코스테로이드는 염증 반응을 줄이기 위해 사용된다.[71]
9. 초항원 생산의 진화
초항원 생산은 미생물이 면역 반응을 효과적으로 붕괴시켜 확인되지 않고 운반 및 전염되게 한다. 이는 T세포가 항체와 초항원에 대해 아네르기 상태를 유도함으로써 일어난다.[64][67] Lussow와 MacDonald는 연쇄상구균 항원을 동물에 전신적으로 노출시켜 이를 입증했는데, 초항원 감염 이후에 다른 항원에 노출될 경우 면역반응을 끌어내는데 실패하는 것을 확인했다.[67] Watson과 Lee의 실험에서는 일반적인 항원자극에 의해 형성된 기억 T 세포가 초항원 자극에 대해서는 아네르기 상태인 것과 초항원 감염 이후 형성된 기억 T세포가 모든 항원 자극에 대해 아네르기 상태인 것을 발견했다.[64] 이러한 현상이 발생하는 기작은 아직 분명히 알려지지 않았다. 초항원의 발현을 조절하는 유전자는 M단백질이나 세균 캡슐 발현 같은 면역 회피 기작 또한 조절한다. 이는 초항원의 생산이 면역체계 침입을 중심으로 진화했다는 가설을 뒷받침한다.[74]
개별 초항원의 도메인 구조를 다른 면역 항체에 결합하는 연쇄상 구균 단백질과 비교했을 때, 각각의 도메인은 각 계보에 속하는 것들과 유사함이 발견됐다. 이러한 유사성은 초항원이 두개의 작은 베타 가닥 모양의 재조합을 통해 진화했음을 보여준다.[75]
10. 내인성 초항원
MLS 외독소(Minor lymphocyte stimulating exotoxins)는 원래 쥐의 흉선 기질 세포에서 발견되었다. 이 독소들은 초항원 유전자에 의해 발현되었으며, 이들은 쥐 유선 종양 바이러스(mouse mammary tumor virus, MMTV)에서 쥐 게놈으로 통합된 것이다. 쥐 게놈에서 이런 유전자의 존재는 쥐가 흉선에서 항원을 표현하도록 하며, 이는 바이러스 유래 초항원의 자극에 감수성이 있는 베타 가변부를 가진 림프구가 음성 선택(negative selecting)을 하는 수단이 된다. 그 결과 이 쥐들은 이후 일생동안 그 바이러스에 대한 감염에 대해 면역을 가지게 된다.[53]
인간 게놈에서는 이와 유사한 내인성 초항원 의존적 선택과정이 밝혀지지는 않았지만, 내인성 초항원은 밝혀졌으며 이것들이 바이러스 감염에서 통합적인 역할을 하는 것으로 의심된다. 예를 들어, 엡스타인-바 바이러스 감염은 감염세포에서 초항원 생산을 일으키는 것으로 알려져 있다. 하지만 바이러스의 게놈에서 독소의 유전자가 발견되지는 않았다. 바이러스는 자기의 초항원이 감염 세포에서 발현되도록 조절하며, 이는 숙주의 면역 체계를 피하는 데 도움을 준다. 비슷한 결과가 광견병, 거대세포바이러스, HIV에서 발견되었다.[53]
참조
[1]
서적
Review of Medical Microbiology and Immunology
McGraw-Hill Education
[2]
논문
Superantigens: microbial agents that corrupt immunity
2002-03
[3]
논문
The structural basis of T cell activation by superantigens
[4]
논문
Streptococcus pyogenes: Insight into the function of the streptococcal superantigens
[5]
논문
Interplay between superantigens and immunoreceptors
2004-04
[6]
논문
Cross-linking of major histocompatibility complex class II molecules by staphylococcal enterotoxin A superantigen is a requirement for inflammatory cytokine gene expression
1995-11
[7]
논문
TCR recognition of peptide/MHC class II complexes and superantigens
2007-08
[8]
논문
Crystal structure of microbial superantigen staphylococcal enterotoxin B at 1.5 A resolution: implications for superantigen recognition by MHC class II molecules and T-cell receptors
1998-03
[9]
논문
Enhancement of host susceptibility to lethal endotoxin shock by staphylococcal pyrogenic exotoxin type C
1982-04
[10]
논문
Staphylococcal and streptococcal superantigens: molecular, biological and clinical aspects
2003-02
[11]
논문
Crystal structure of the streptococcal superantigen SpeI and functional role of a novel loop domain in T cell activation by group V superantigens
2007-04
[12]
논문
Characterization of T cell receptors engineered for high affinity against toxic shock syndrome toxin-1
2005-10
[13]
논문
Three-dimensional structure of the complex between a T cell receptor beta chain and the superantigen staphylococcal enterotoxin B
1998-12
[14]
논문
Conservation and variation in superantigen structure and activity highlighted by the three-dimensional structures of two new superantigens from Streptococcus pyogenes
2000-05
[15]
논문
Defective T cell Receptor-mediated Signal Transduction in Memory CD4 T Lymphocytes Exposed to Superantigen or anti-T cell Receptor Antibodies
2006-08
[16]
논문
Molecular mechanisms for adaptive tolerance and other T cell anergy models
2007-06
[17]
논문
Staphylococcal Enterotoxins: a Purging Experience in Review, Part I
[18]
논문
Differential effects of superantigen-induced "anergy" on priming and effector stages of a T cell-dependent antibody response
1994-02
[19]
논문
Anergy and Cytokine-Mediated Suppression as Distinct Superantigen-Induced Tolerance Mechanisms in Vivo
1999-07
[20]
논문
Induction of negative regulators of haematopoiesis in human bone marrow cells by HLA-DR cross-linking
1999-09
[21]
논문
Interferon-alpha-induced endogenous superantigen. a model linking environment and autoimmunity
2001-10
[22]
논문
The superantigen toxic shock syndrome toxin-1 induces CD40 ligand expression and modulates IgE isotype switching
1996-10
[23]
논문
Induction of acute inflammation in vivo by staphylococcal superantigens I: Leukocyte recruitment occurs independently of T lymphocytes and major histocompatibility complex Class II molecules
1998-06
[24]
논문
New insights into the pathology of nasal polyposis: the role of superantigens and IgE
[25]
논문
Superantigens are critical for Staphylococcus aureus infective endocarditis, sepsis, and acute kidney injury
[26]
논문
Identification of the Antigenic Epitopes in Staphylococcal Enterotoxins A and E and Design of a Superantigen for Human Cancer Therapy
[27]
논문
High-frequency intracellular infection and erythrogenic toxin A expression undergo phase variation in M1 group A streptococci
1998-04
[28]
논문
''Staphylococcus aureus'' enterotoxins: a key in airway disease?
2002-06
[29]
논문
Structural and functional properties of staphylococcal superantigen-like protein 4.
2012-11
[30]
논문
Staphylococcal enterotoxin-like X (SElX) is a unique superantigen with functional features of two major families of staphylococcal virulence factors.
2017-09
[31]
논문
Cutting Edge: Epstein-Barr Virus Transactivates the HERV-K18 Superantigen by Docking to the Human Complement Receptor 2 (CD21) on Primary B Cells
2006-08-15
[32]
논문
Three-dimensional structure of the complex between a T cell receptor beta chain and the superantigen staphylococcal enterotoxin B
http://linkinghub.el[...]
[33]
논문
Superantigens: microbial agents that corrupt immunity
[34]
논문
Enhancement of host susceptibility to lethal endotoxin shock by staphylococcal pyrogenic exotoxin type C
http://iai.asm.org/c[...]
[35]
논문
High-frequency intracellular infection and erythrogenic toxin A expression undergo phase variation in M1 group A streptococci
http://www3.intersci[...]
[36]
DorlandsDict
superantigen
nine/000634520
[37]
논문
Crystal structure of the streptococcal superantigen SpeI and functional role of a novel loop domain in T cell activation by group V superantigens
http://linkinghub.el[...]
[38]
논문
Characterization of T cell receptors engineered for high affinity against toxic shock syndrome toxin-1
http://linkinghub.el[...]
[39]
논문
Staphylococcal and streptococcal superantigens: molecular, biological and clinical aspects
[40]
논문
Streptococcus pyogenes: Insight into the function of the streptococcal superantigens
http://linkinghub.el[...]
[41]
논문
Interplay between superantigens and immunoreceptors
http://www3.intersci[...]
[42]
논문
Cross-linking of major histocompatibility complex class II molecules by staphylococcal enterotoxin A superantigen is a requirement for inflammatory cytokine gene expression
[43]
논문
Crystal structure of microbial superantigen staphylococcal enterotoxin B at 1.5 A resolution: implications for superantigen recognition by MHC class II molecules and T-cell receptors
http://linkinghub.el[...]
[44]
논문
Conservation and variation in superantigen structure and activity highlighted by the three-dimensional structures of two new superantigens from Streptococcus pyogenes
http://linkinghub.el[...]
[45]
논문
Defective T cell receptor-mediated signal transduction in memory CD4 T lymphocytes exposed to superantigen or anti-T cell receptor antibodies
http://linkinghub.el[...]
[46]
논문
Molecular mechanisms for adaptive tolerance and other T cell anergy models
http://linkinghub.el[...]
[47]
논문
Staphylococcal Enterotoxins: a Purging Experience in Review, Part I
[48]
논문
Differential effects of superantigen-induced "anergy" on priming and effector stages of a T cell-dependent antibody response
[49]
논문
Anergy and cytokine-mediated suppression as distinct superantigen-induced tolerance mechanisms in vivo
http://www.jem.org/c[...]
[50]
논문
Induction of negative regulators of haematopoiesis in human bone marrow cells by HLA-DR cross-linking
http://linkinghub.el[...]
[51]
논문
Interferon-alpha-induced endogenous superantigen. a model linking environment and autoimmunity
http://linkinghub.el[...]
[52]
논문
The superantigen toxic shock syndrome toxin-1 induces CD40 ligand expression and modulates IgE isotype switching
http://intimm.oxford[...]
[53]
저널인용
Superantigens: microbial agents that corrupt immunity
2002-03
[54]
저널인용
Streptococcus pyogenes: Insight into the function of the streptococcal superantigens
http://linkinghub.el[...]
[55]
저널인용
Interplay between superantigens and immunoreceptors
http://www3.intersci[...]
2004-04
[56]
저널인용
Cross-linking of major histocompatibility complex class II molecules by staphylococcal enterotoxin A superantigen is a requirement for inflammatory cytokine gene expression
1995-11
[57]
저널인용
Crystal structure of microbial superantigen staphylococcal enterotoxin B at 1.5 A resolution: implications for superantigen recognition by MHC class II molecules and T-cell receptors
http://linkinghub.el[...]
1998-03
[58]
저널인용
Enhancement of host susceptibility to lethal endotoxin shock by staphylococcal pyrogenic exotoxin type C
http://iai.asm.org/c[...]
1982-04
[59]
저널인용
Staphylococcal and streptococcal superantigens: molecular, biological and clinical aspects
2003-02
[60]
저널인용
Crystal structure of the streptococcal superantigen SpeI and functional role of a novel loop domain in T cell activation by group V superantigens
http://linkinghub.el[...]
2007-04
[61]
저널인용
Characterization of T cell receptors engineered for high affinity against toxic shock syndrome toxin-1
http://linkinghub.el[...]
2005-10
[62]
저널인용
Three-dimensional structure of the complex between a T cell receptor beta chain and the superantigen staphylococcal enterotoxin B
http://linkinghub.el[...]
1998-12
[63]
저널인용
Conservation and variation in superantigen structure and activity highlighted by the three-dimensional structures of two new superantigens from Streptococcus pyogenes
http://linkinghub.el[...]
2000-05
[64]
저널인용
Defective T cell Receptor-mediated Signal Transduction in Memory CD4 T Lymphocytes Exposed to Superantigen or anti-T cell Receptor Antibodies
http://linkinghub.el[...]
2006-08
[65]
저널인용
MOLECULAR MECHANISMS FOR ADAPTIVE TOLERANCE AND OTHER T CELL ANERGY MODELS
http://linkinghub.el[...]
2007-06
[66]
저널인용
Staphylococcal Enterotoxins: a Purging Experience in Review, Part I
[67]
저널인용
Differential effects of superantigen-induced "anergy" on priming and effector stages of a T cell-dependent antibody response
1994-02
[68]
저널인용
Anergy and Cytokine-Mediated Suppression as Distinct Superantigen-Induced Tolerance Mechanisms in Vivo
http://www.jem.org/c[...]
1999-07
[69]
저널인용
Induction of negative regulators of haematopoiesis in human bone marrow cells by HLA-DR cross-linking
http://linkinghub.el[...]
1999-09
[70]
저널인용
Interferon-alpha-induced endogenous superantigen. a model linking environment and autoimmunity
http://linkinghub.el[...]
2001-10
[71]
논문
The superantigen toxic shock syndrome toxin-1 induces CD40 ligand expression and modulates IgE isotype switching
http://intimm.oxford[...]
1996-10
[72]
논문
Induction of acute inflammation in vivo by staphylococcal superantigens I: Leukocyte recruitment occurs independently of T lymphocytes and major histocompatibility complex Class II molecules
1998-06
[73]
논문
Identification of the Antigenic Epitopes in Staphylococcal Enterotoxins A and E and Design of a Superantigen for Human Cancer Therapy
[74]
논문
High-frequency intracellular infection and erythrogenic toxin A expression undergo phase variation in M1 group A streptococci
http://www3.intersci[...]
1998-04
[75]
논문
"Staphylococcus aureus" enterotoxins: a key in airway disease?
http://www3.intersci[...]
2002-06
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com