P53
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
p53은 인간의 17번 염색체에 위치한 유전자이자, 다양한 항암 작용 메커니즘을 가진 종양 억제 유전자이다. DNA 손상 시 DNA 복구, 세포 주기 정지, 세포 사멸 유도 등의 기능을 수행하며, 줄기 세포 유지 및 혈관 신생 억제에도 관여한다. p53은 다양한 스트레스 요인에 반응하여 활성화되며, Mdm2 단백질에 의해 분해되어 항상 낮은 수준으로 유지된다. p53 유전자의 돌연변이는 리-프라우메니 증후군과 같은 질병을 유발하며, 다양한 암에서 발견된다. p53은 여러 단백질과 상호작용하며, 유전자 치료를 통해 암 치료에 활용하려는 연구가 진행되고 있다.
더 읽어볼만한 페이지
- 암 억제 유전자 - BRCA1
BRCA1은 DNA 복구, 전사 조절, 세포 주기 조절 등 다양한 기능을 수행하며 유방암 및 난소암과 관련되어, 이 유전자의 돌연변이는 유전성 유방-난소암 증후군을 유발하여 암 위험을 증가시킨다. - 암 억제 유전자 - 종양억제유전자
종양억제유전자는 세포의 비정상적인 성장을 억제하여 암 발생을 막는 유전자로, Rb 유전자, p53 유전자, MSH2 유전자 등이 있으며, 세포주기 조절, 세포 증식 억제, DNA 손상 복구, 세포자멸사 유도 등 다양한 메커니즘을 통해 암 발생을 억제한다. - 노화 관련 단백질 - 텔로머레이스
텔로머레이스는 진핵세포 염색체 말단의 텔로미어 DNA 반복 서열을 연장하는 역전사 효소로, 세포 분열 시 텔로미어 단축을 막아 세포 수명 유지에 중요한 역할을 하며, RNA 구성 요소(TERC)와 역전사 효소(TERT) 복합체로 구성되어 세포 노화, 암, 유전 질환 연구와 관련이 있고, 텔로미어와 텔로머레이스 기능 연구로 엘리자베스 블랙번, 캐럴 W. 그리더, 잭 W. 쇼스택이 2009년 노벨 생리학·의학상을 수상했다. - 노화 관련 단백질 - IGF-1
IGF-1은 성장 호르몬에 의해 간에서 생성되는 폴리펩타이드 호르몬으로, 성장, 대사, 단백질 번역 등 동화 작용에 관여하며, GH-IGF-1 축을 통해 근력 유지, 골격 발달, 뇌 발달 등에 기여하지만, 비정상적인 수치는 질환과 관련될 수 있고 스포츠계에서는 금지 약물로 지정되어 있다. - 세포예정사 - 사이토크롬 c
사이토크롬 c는 진핵생물의 미토콘드리아 내막에 존재하는 전자 전달계 구성 요소이자 세포자멸사 관여 단백질로서, 작은 크기와 보존된 서열 덕분에 분자 계통학 연구에 활용되며 항산화 효소 및 저준위 레이저 치료에서의 역할 등 다양한 응용 가능성이 제시된다. - 세포예정사 - 오토파지
오토파지는 세포 내 구성 성분을 분해 및 재활용하여 항상성을 유지하고, 영양 결핍 시 에너지와 물질을 공급하는 과정으로, 거대 자가포식, 미세 자가포식, 샤페론 매개 자가포식의 세 유형으로 나뉘며, 오스미 요시노리의 노벨상 수상 이후 질병 치료 표적으로 연구가 활발하다.
| P53 | |
|---|---|
| 일반 정보 | |
 | |
| 유전자 기호 | TP53 |
| 유전자 이름 | 종양 단백질 p53 |
| 염색체 | 17p13 |
| 염색체 위치 | 17번 염색체 짧은 팔 13 밴드 |
| 다른 명칭 | BCC7 LFS1 P53 TRP53 |
| 기능 | |
| 주요 기능 | 종양 억제 |
| 관련 질병 | 리-프라우메니 증후군 다양한 암 |
| 상세 정보 | |
| 발견 | 1979년 |
| 발견자 | 데이비드 레인 월터 길버트 아놀드 레빈 로이드 올드 |
| 유전자 정보 | |
| OMIM | 191170 |
| UniProt | P04637 |
| 추가 정보 | |
| 관련 용어 | 텔로미어 유지 DNA 복구 센트로미어 구조 |
2. 유전좌위
사람의 p53 유전자는 17번 염색체 단완(17p13.1)에 존재한다.[4][5][6][7] 이 유전자는 20 kb에 걸쳐 있으며, 비암호화 엑손 1과 10 kb에 달하는 매우 긴 첫 번째 인트론을 가지고 있어, Hp53int1 유전자와 겹쳐 있다. 암호화 서열은 척추동물에서 보존성이 높은 다섯 개의 영역, 주로 엑손 2, 5, 6, 7, 8에 포함되어 있지만, 무척추동물에서 발견되는 서열은 포유류 TP53과 거리가 먼 유사성을 보일 뿐이다.[10] 완전한 게놈 데이터를 사용할 수 있는 대부분의 포유류에서 ''TP53'' 상동 유전자[11]가 확인되었다.
1979년 아놀드 J. 레빈 등이 종양 바이러스 SV40의 대형 T 항원과 결합하는 단백질로 p53을 처음 발견했다.[233] 1989년 레빈과 버트 보겔스타인 등은 p53 유전자가 암 억제 유전자임을 밝혔다.[234] 2015년 미국 의사 협회 저널에는 코끼리가 사람보다 p53 유전자를 많이 가지고 있어 암 발생률이 낮다는 연구 결과가 발표되었다.[235]
p53은 다양한 항암 작용 메커니즘을 가지며, 세포자살, 유전체 안정성 유지, 혈관 신생 억제 등의 역할을 한다.[2]
3. 연구사
2016년 KAIST 김재경 수리과학과 교수팀과 미국 버지니아 공대 연구팀은 미국국립과학원회보(PNAS)에 세포질과 세포핵 내에서 p53 단백질의 양 조절 메커니즘을 밝힌 연구 결과를 발표하였다.[246] 이 연구에서는 생체시계의 주요 유전자인 period와 관련된 period2 단백질이 p53의 양 조절에 관여한다는 사실이 드러났다.
4. 기능
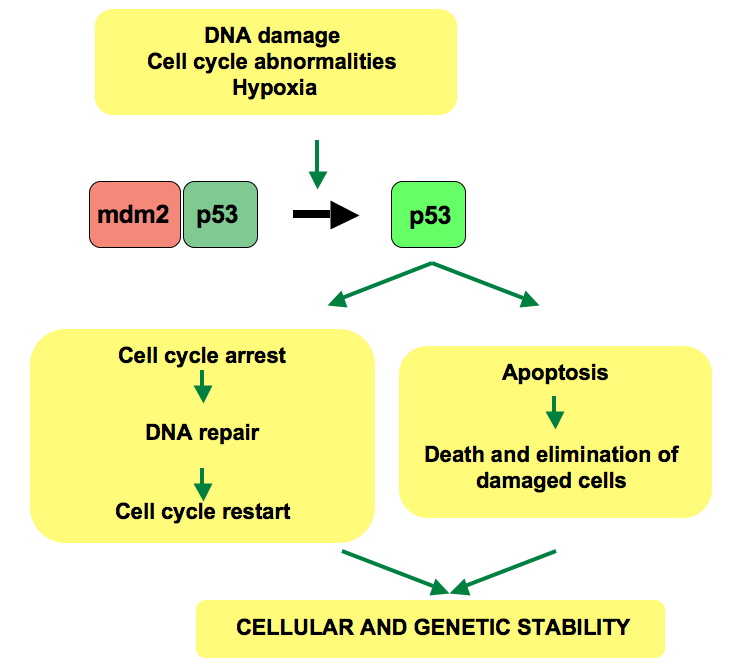
p53은 p21을 비롯한 여러 유전자의 발현을 조절한다. p21은 G1기-S기/CDK 복합체에 결합하여 활성을 억제하고, 세포 주기를 정지시킨다. 돌연변이 p53은 DNA 결합 능력이 손상되어 p21을 통한 세포 주기 정지 기능을 수행할 수 없다.[23]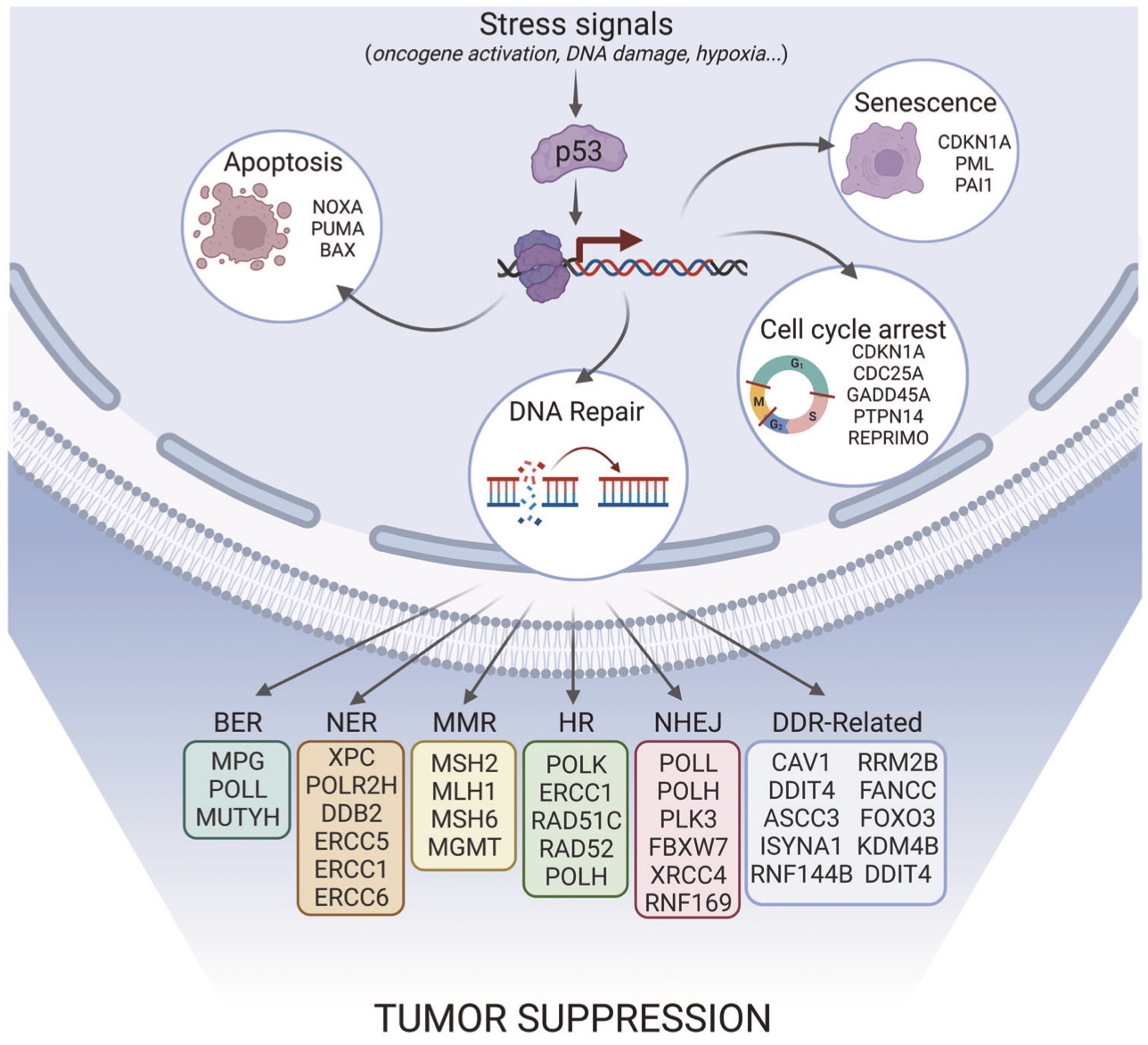
p53은 RB1 경로와 상호작용하며,[24] 백혈병 억제 인자(LIF)를 조절하여 포유류의 착상을 돕는다.[243] 또한, 자외선에 의해 활성화되어 피부를 태양으로부터 보호하는 태닝 반응을 유도한다.[25][26]
p53은 전사 인자로 작용하여 Gadd45, PPM1D, MDM2, p21CIP1/WAF1, BAX, 14-3-3δ 등 다양한 유전자의 발현에 관여한다.
p53은 생체시계의 주요 유전자인 period와 관련된 period2 단백질의 양을 조절한다.[246]
4. 1. 줄기 세포
p53의 수준은 발생 과정과 인간의 남은 생애 동안 줄기 세포 유지에 중요한 역할을 한다.[27]
인간 배아 줄기 세포(hESC)에서 p53은 낮은 비활성 수준으로 유지된다. 이는 p53의 활성화가 hESC의 급격한 분화를 유발하기 때문이다.[28] 연구에 따르면 p53을 제거하면 분화가 지연되고, p53을 추가하면 자발적인 분화가 일어나 p53이 hESC의 분화를 촉진하고 분화 조절자로서 세포 주기에 중요한 역할을 한다는 것을 보여준다. p53이 hESC에서 안정화되고 활성화되면 p21을 증가시켜 더 긴 G1기를 설정한다. 이것은 일반적으로 S기 진입을 없애 G1기에서 세포 주기를 멈추고 분화를 유도한다. 그러나 최근 생쥐 배아 줄기 세포 연구에서는 P53의 발현이 반드시 분화를 유발하지는 않는다는 것을 보여주었다.[29] p53은 또한 miR-34a 및 miR-145를 활성화하여 hESC의 다능성 인자를 억제하여 분화를 더욱 촉진한다.[27]
성체 줄기 세포에서 p53 조절은 성체 줄기 세포 틈새에서 줄기 세포성을 유지하는 데 중요하다. 저산소증과 같은 기계적 신호는 저산소 유도 인자, HIF-1α 및 HIF-2α를 통해 이러한 틈새 세포에서 p53의 수준에 영향을 미친다. HIF-1α는 p53을 안정화시키고, HIF-2α는 p53을 억제한다.[30] p53의 억제는 암 줄기 세포 표현형, 유도 만능 줄기 세포 및 블라스테마 형성과 같은 다른 줄기 세포 역할과 행동에 중요한 역할을 한다. p53 수준이 감소한 세포는 정상 세포보다 훨씬 더 높은 효율로 줄기 세포로 재프로그래밍되는 것으로 나타났다.[31][32] 논문에 따르면 세포 주기 정지 및 세포 사멸의 부재는 더 많은 세포에게 재프로그래밍될 기회를 제공한다. p53의 감소된 수준은 또한 도롱뇽 다리에서 블라스테마 형성에 중요한 측면으로 나타났다.[33] p53 조절은 줄기 세포와 분화된 줄기 세포 상태 사이의 장벽, 그리고 줄기 세포가 기능적인 상태와 암 상태 사이의 장벽 역할을 하는 데 매우 중요하다.[34]
4. 2. 혈관 신생 억제
p53은 조직 수준에서 혈관 신생을 억제하여 항암 효과를 나타낸다.[2] 종양이 성장하기 위해서는 새로운 혈관이 필요하다. p53은 다음의 방법으로 신생 혈관 생성을 억제한다.[36][37]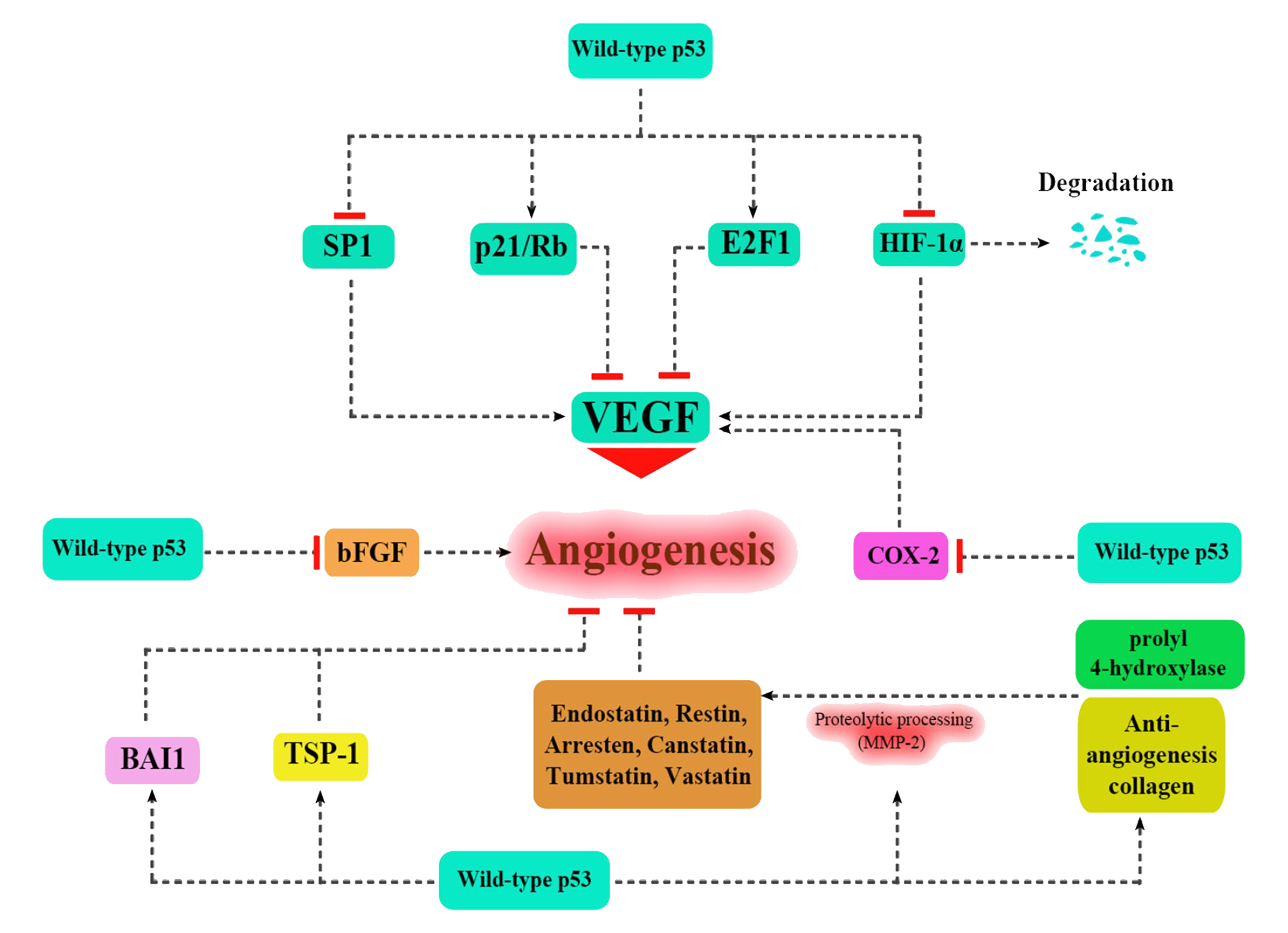
5. 조절
p53은 세포 스트레스 감지기 역할을 한다. p53은 DNA 손상(UV, IR, 과산화수소와 같은 화학 물질에 의해 유도됨), 산화 스트레스,[41] 삼투압 쇼크, 리보뉴클레오티드 고갈, 바이러스성 폐렴[42] 및 조절되지 않는 종양 유전자 발현을 포함한 다양한 스트레스 요인에 반응하여 활성화된다. 활성화는 두 가지 주요 사건으로 특징지어진다. 첫째, p53 단백질의 반감기가 급격히 증가하여 스트레스 받은 세포에서 p53이 빠르게 축적된다. 둘째, 입체 구조 변화는 p53이 이러한 세포에서 전사 조절 인자로 활성화되도록 한다. p53 활성화를 유도하는 중요한 사건은 N-말단 도메인의 인산화이다. N-말단 전사 활성화 도메인은 다수의 인산화 부위를 포함하며, 스트레스 신호를 전달하는 단백질 키나제의 주요 표적으로 간주될 수 있다.
p53의 전사 활성화 도메인을 표적으로 하는 단백질 키나제는 크게 두 그룹으로 나눌 수 있다. 첫 번째 그룹은 MAPK 계열(JNK1-3, ERK1-2, p38 MAPK)에 속하며 막 손상, 산화 스트레스, 삼투압 쇼크, 열 쇼크 등과 같은 여러 유형의 스트레스에 반응한다. 두 번째 그룹(ATR, ATM, CHK1 및 CHK2, DNA-PK, CAK, TP53RK)은 유전자 손상 스트레스에 의해 유발되는 DNA 손상을 감지하고 반응하는 분자 연쇄 반응인 게놈 무결성 검사점에 관여한다. 종양 유전자는 p14ARF 단백질에 의해 매개되는 p53 활성화를 자극한다.
스트레스를 받지 않은 세포에서 p53 수준은 Mdm2(사람에서는 HDM2라고도 함)에 의해 낮게 유지된다.[40] Mdm2는 p53에 결합하여 p53의 작용을 방지하고 세포 핵에서 세포질로 수송한다. Mdm2는 유비퀴틴 연결 효소로 작용하여 유비퀴틴을 p53에 공유 결합시키고, p53을 프로테아좀에 의한 분해 표적으로 삼는다. p53의 유비퀴틴화는 가역적이다. p53이 활성화되면 Mdm2도 활성화되어 피드백 루프를 설정한다. p53 수준은 특정 스트레스에 반응하여 진동 (반복적인 펄스)을 보일 수 있으며, 이러한 펄스는 세포가 스트레스에서 살아남을지 또는 죽을지를 결정하는 데 중요할 수 있다.[43]
MI-63은 MDM2에 결합하여 p53의 기능이 억제된 상황에서 p53을 재활성화한다.[44]
USP7(HAUSP)은 p53에서 유비퀴틴을 절단하여 유비퀴틴화 경로를 통해 프로테아좀 의존적 분해로부터 p53을 보호한다. 이는 종양 유전자 공격에 반응하여 p53이 안정화되는 한 가지 방법이다. USP42 또한 p53의 탈유비퀴틴화를 하며, p53이 스트레스에 반응하는 능력에 필요할 수 있다.[45]
HAUSP는 주로 핵에 국한되어 있지만, 일부는 세포질과 미토콘드리아에서도 발견될 수 있다. HAUSP 과발현은 p53을 안정화시킨다. HAUSP 고갈은 p53 수준의 감소를 초래하지 않고, HAUSP가 Mdm2에 결합하여 탈유비퀴틴화를 하기 때문에 p53 수준을 증가시킨다. 스트레스를 받지 않은 세포에서 HAUSP는 p53보다 Mdm2에 더 나은 결합 파트너이다.
USP10은 스트레스를 받지 않은 세포에서 세포질에 위치하여 세포질 p53의 탈유비퀴틴화를 하고 Mdm2 유비퀴틴화를 역전시킨다. DNA 손상 후 USP10은 핵으로 전위하여 p53 안정성에 기여한다. USP10은 Mdm2와 상호작용하지 않는다.
단백질 키나제에 의한 p53 N-말단 인산화는 Mdm2 결합을 방해한다. Pin1과 같은 다른 단백질은 p53으로 동원되어 p53의 입체 구조 변화를 유도하여 Mdm2 결합을 더욱 방지한다. 인산화는 p300 및 PCAF와 같은 전사 공동 활성 인자의 결합을 허용하며, 이들은 p53의 C-말단을 아세틸화하여 p53의 DNA 결합 도메인을 노출시켜 특정 유전자를 활성화하거나 억제할 수 있도록 한다. Sirt1 및 Sirt7과 같은 탈아세틸화 효소는 p53을 탈아세틸화하여 세포 사멸 억제를 유발할 수 있다.[46] 일부 종양 유전자는 MDM2에 결합하여 그 활성을 억제하는 단백질의 전사를 자극할 수 있다.
히스톤 메틸화와 같은 후성 유전적 표지도 p53을 조절할 수 있다. 예를 들어, p53은 후성 유전적으로 억제된 게놈 영역에서 히스톤에 결합하는 억제성 Trim24 공동 인자와 직접 상호 작용한다.[47] Trim24는 p53이 표적을 활성화하는 것을 방지하지만, 이러한 영역에서만 p53이 주요 표적 유전자에서 히스톤 프로파일을 '판독'하고 유전자 특이적인 방식으로 작용하는 능력을 효과적으로 제공한다.
6. 질병과의 관련성
''TP53'' 유전자가 손상되면 종양 억제 기능이 심각하게 저하된다. ''TP53'' 유전자의 기능적 사본을 하나만 상속받는 사람은 리-프라우메니 증후군이 발생한다. 인간 종양의 50% 이상이 ''TP53'' 유전자의 돌연변이 또는 결실을 포함한다.[48] p53의 손실은 이수배수성 표현형을 초래하는 게놈 불안정성을 생성한다.[49]
''TP53'' 유전자는 돌연변이 유발 물질(화학 물질, 방사선, 또는 바이러스)에 의해 변형될 수 있으며, 이는 통제되지 않는 세포 분열의 가능성을 증가시킨다. 특정 병원체는 ''TP53'' 유전자가 발현하는 p53 단백질에 영향을 미칠 수 있다. 인간 유두종 바이러스(HPV)는 p53 단백질에 결합하여 비활성화하는 단백질인 E6을 암호화한다.
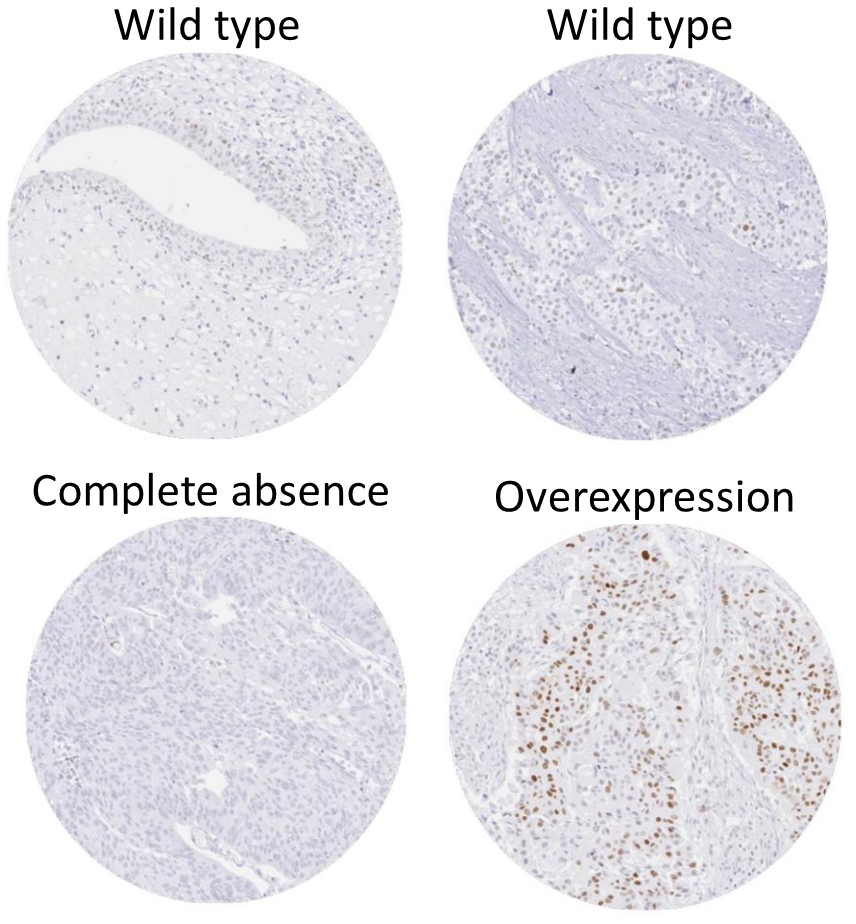
6. 1. p53 변이와 암
p53은 세포자살, 유전체 안정성에 관여하며 혈관 신생을 억제하는 등 다양한 항암 작용 메커니즘을 가진 단백질이다. p53은 DNA 손상 시 DNA 수선 단백질을 활성화하고, 세포주기를 G1/S 단계에서 정지시키며, 손상이 복구 불가능할 경우 세포자살을 유도한다.[239]활성화된 p53은 DNA에 결합하여 miR-34a microRNA를 포함한 여러 유전자를 발현시킨다.[239] WAF1/CIP1에 의해 암호화되는 p21은 G1-S/CDK 및 S/CDK 복합체에 결합하여 세포주기 진행을 억제한다. p53 유전자에 돌연변이가 발생하면 DNA 결합 능력이 저하되어 세포주기 정지 신호를 제대로 전달하지 못하게 된다.[240]
최근 연구에서는 p53과 망막아세포종 단백질(RB1) 경로 간의 연관성이 밝혀졌다.[242] 또한, p53은 백혈병 억제 인자(LIF)를 조절하여 쥐 모델에서 이식을 용이하게 하며, 인간에게도 적용 가능성을 시사한다.[243] p53 발현은 DNA 손상을 유발하는 자외선에 의해 자극될 수 있으며, 이는 선탠을 유발하는 기전 중 하나이다.[244][245]
인간에서는 엑손 4의 코돈 72번 위치에서 아르기닌이 프롤린으로 치환되는 다형성이 흔하게 나타난다. 이 변이와 암 감수성 간의 연관성에 대한 연구는 논란이 있지만, 일부 연구에서는 특정 암(췌장암[13], 유방암[14], 비구강인두암[15], 폐암[16] 등) 위험과의 연관성을 보고하기도 했다. 그러나 대장암[17] 및 자궁내막암[18]과의 연관성은 발견되지 않았다.
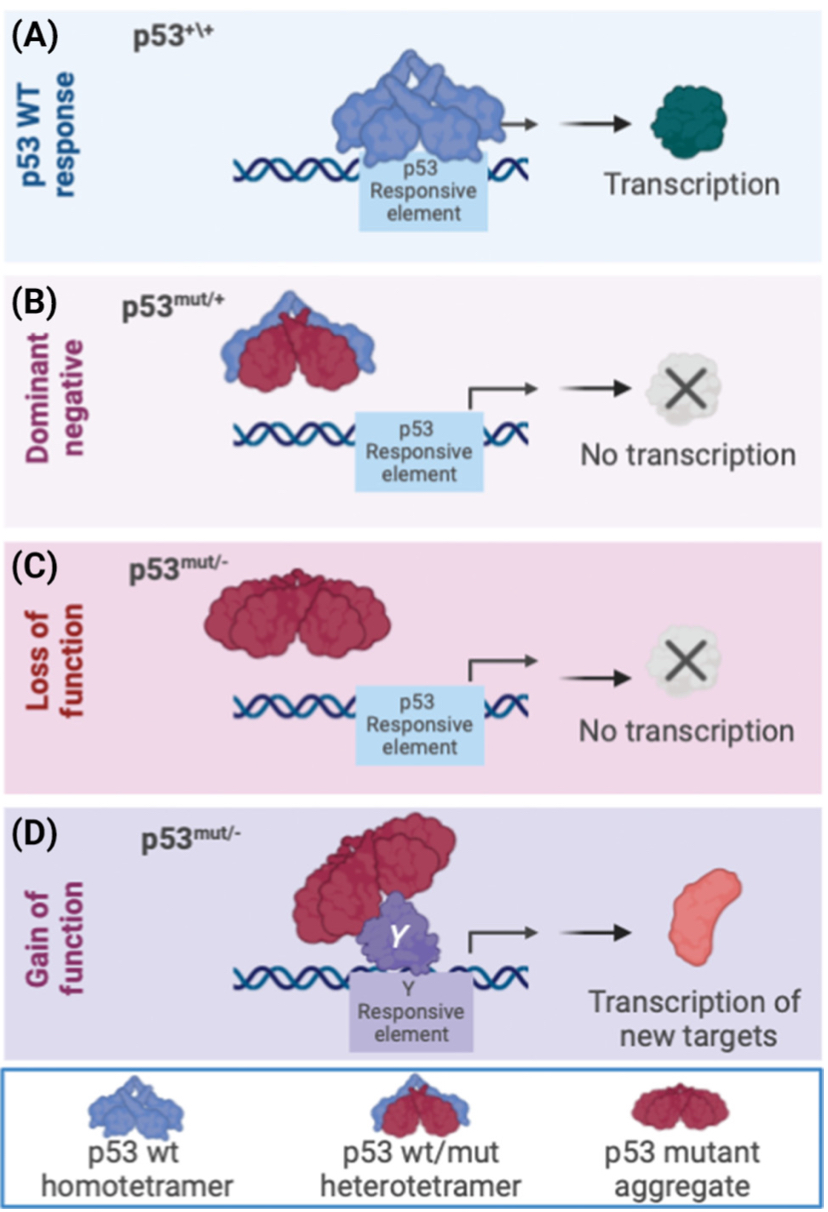
''TP53'' 유전자 돌연변이는 다양한 암 표현형을 유발할 수 있으며, 이는 p53 단백질의 다양한 이소형이 암 예방에 대해 서로 다른 세포 메커니즘을 가지기 때문이다. 돌연변이는 서로 다른 이소형을 생성하여 세포 메커니즘을 방해하고, 암 표현형을 다양하게 만든다.[86][64][65][66] TP53 돌연변이는 에너지 대사에도 영향을 미쳐 유방암 세포에서 해당 작용을 증가시킨다.[67]
p53 단백질과 그 길항제인 Mdm2의 역학 관계는 p53 농도가 시간에 따라 진동함을 보여준다. 이러한 진동은 임상적으로 문서화되고 수학적으로 모델링되었다.[68][69][70]
악성 종양의 절반 이상에서 p53 유전자 변이 또는 결실이 발견되며, 대부분은 점 돌연변이이다. p53 유전자 손상은 세포의 세포자멸사 유도를 어렵게 만든다. 예를 들어, 폐암에서는 담배의 벤조피렌이라는 발암 물질, 간세포암에서는 땅콩 곰팡이가 생산하는 아플라톡신 등이 p53 유전자 변이를 유발한다.
p53 유전자 변이는 항 p53 항체 출현과 상관관계가 있으며, 일본에서는 항 p53 항체 측정 검사가 특정 암(식도암, 대장암, 유방암) 의심 시 건강 보험 적용을 받는다.
7. 유전자 치료
p53 유전자의 기능을 활용하여 암 치료에 응용하려는 연구가 진행되고 있다. 특히 아데노바이러스 등을 이용한 '''p53 유전자''' 도입 치료법은 암세포의 아포토시스를 유도하고, 화학 요법이나 방사선 치료의 효과를 높이는 것을 목표로 한다. p53 유전자에 변이가 있으면 DNA 손상이 발생해도 세포 사멸이 잘 일어나지 않아, p53 유전자 변이를 가진 암은 약물이나 방사선 치료에 저항성을 보인다.[239][240][241][242][243][244][245]
8. 구조
p53은 7개의 도메인을 가지고 있다.
1. 산성 N-말단 전사 활성화 도메인(TAD)은 활성 도메인 1(AD1)이라고도 불리며, 전사 인자를 활성화한다. N-말단은 두 개의 보완적인 전사 활성화 도메인을 포함하며, 주요 도메인은 잔기 1–42에, 부도메인은 잔기 55–75에 위치하며, 여러 전-세포사멸 유전자의 조절에 특이적으로 관여한다.[81]
2. 세포자멸사 활성에 중요한 활성 도메인 2(AD2): 잔기 43–63.
3. 프롤린이 풍부한 도메인은 MAPK를 통한 핵 수출을 통해 p53의 세포사멸 활성에 중요: 잔기 64–92.
4. 중심 DNA 결합 코어 도메인(DBD). 하나의 아연 원자와 여러 아르기닌 아미노산을 포함한다: 잔기 102–292. 이 영역은 p53 코-억제제 LMO3의 결합을 담당한다.[82]
5. 핵 국소화 서열 (NLS) 도메인, 잔기 316–325.
6. 호모-올리고머화 도메인(OD): 잔기 307–355. 사합체화는 ''생체 내'' p53의 활성에 필수적이다.
7. C-말단은 중심 도메인의 DNA 결합 하향 조절에 관여: 잔기 356–393.[83]
SDS-PAGE 분석에 따르면 p53은 53-킬로달톤 (kDa) 단백질이다. 그러나 아미노산 잔기의 질량 합계를 기준으로 한 전체 길이 p53 단백질(p53α)의 실제 질량은 단지 43.7kDa이다. 이러한 차이는 단백질 내의 프롤린 잔기 수가 많아 SDS-PAGE에서 이동 속도가 느려 실제보다 더 무거워 보이는 것 때문이다.[85]
9. 이소형
TP53 유전자는 다양한 스플라이싱 방식을 통해 12개 이상의 서로 다른 p53 이소형(isoform)을 생성한다.[1] 이 단백질들은 3.5에서 43.7 kDa 크기로 다양하다. 2005년에 여러 이소형이 발견되었고, 현재까지 12개의 인간 p53 이소형(p53α, p53β, p53γ, ∆40p53α, ∆40p53β, ∆40p53γ, ∆133p53α, ∆133p53β, ∆133p53γ, ∆160p53α, ∆160p53β, ∆160p53γ)이 확인되었다.[86] 이소형들은 조직 특이적으로 발현되며, p53α는 단독으로 발현되지 않는다.[86]
전체 길이의 p53 이소형은 여러 단백질 도메인으로 구성된다. N-말단에는 아미노 말단 전사 활성화 도메인(TAD 1, TAD 2)이 있고, p53 표적 유전자 발현에 관여한다. 다음으로 프롤린이 풍부한 도메인(PXXP)이 반복되는(P는 프롤린, X는 임의의 아미노산) 프롤린 풍부 도메인이 오며, 이는 p53 매개 세포자멸사에 필요하다.[87] Δ133p53β,γ 및 Δ160p53α,β,γ와 같은 일부 이소형은 이 도메인이 없어 세포자멸사를 매개하지 않는다.[64] 이어서 서열 특이적 결합을 위한 DNA 결합 도메인(DBD)이 온다. C-말단 도메인에는 핵 국소화 신호(NLS), 핵 수출 신호(NES), 올리고머화 도메인(OD)이 있다. NLS와 NES는 p53의 세포 내 위치 조절에 관여하고, OD는 p53의 사합체 형성과 DNA 결합을 돕는다. 이소형들은 일부 도메인이 없을 수 있지만, 대부분 고도로 보존된 DNA 결합 도메인을 공유한다.
이소형은 다양한 메커니즘으로 형성된다. 인트론 9의 다중 스플라이싱은 베타 및 감마 이소형을 생성하여 다른 C-말단을 만든다. 인트론 4의 내부 프로모터 사용은 TAD 도메인과 DBD 일부가 없는 ∆133 및 ∆160 이소형을 유발한다. 코돈 40 또는 160에서의 번역 개시는 ∆40p53 및 ∆160p53 이소형을 생성한다.[86]
p53 단백질의 이소형 특성 때문에, ''TP53'' 유전자 내 돌연변이는 돌연변이된 이소형을 생성하여, ''TP53'' 유전자의 단일 돌연변이가 다양한 암 표현형을 유발할 수 있다.[86][64][65][66]
10. 상호작용
p53은 여러 단백질과 상호작용하는 것으로 알려져 있다. 다음은 p53과 상호작용하는 단백질 목록이다.
| 단백질 | 설명 |
|---|---|
| AIMP2 | |
| ANKRD2 | |
| APTX | |
| ATM | |
| ATR | |
| ATF3 | |
| AURKA | |
| BAK1 | |
| BARD1 | |
| BLM | |
| BRCA1 | |
| BRCA2 | |
| BRCC3 | |
| BRE | |
| CEBPZ | |
| CDC14A | |
| CDK1 | |
| CFLAR | |
| CHEK1 | |
| CCNG1 | |
| CREBBP | |
| CREB1 | |
| 사이클린 H | |
| CDK7 | |
| DNA-PKcs | |
| E4F1 | |
| EFEMP2 | |
| EIF2AK2 | |
| ELL | |
| EP300 | |
| ERCC6 | |
| GNL3 | |
| GPS2 | |
| GSK3B | |
| HSP90AA1 | |
| HIF1A | |
| HIPK1 | |
| HIPK2 | |
| HMGB1 | |
| HSPA9 | |
| 헌팅틴 | |
| ING1 | |
| ING4 | |
| ING5 | |
| IκBα | |
| KPNB1 | |
| LMO3 | |
| MDM2 | |
| MDM4 | |
| MED1 | |
| MAPK9 | |
| MNAT1 | |
| NDN | |
| 핵인 | |
| NUMB | |
| NF-κB | |
| P16 | |
| PARC | |
| PARP1 | |
| PIAS1 | |
| CDC14B | |
| PIN1 | |
| PLAGL1 | |
| PLK3 | |
| PRKRA | |
| PHB | |
| PML | |
| PSME3 | |
| PTEN | |
| PTK2 | |
| PTTG1 | |
| RAD51 | |
| RCHY1 | |
| RELA | |
| Reprimo | |
| RPA1 | |
| RPL11 | |
| S100B | |
| SUMO1 | |
| SMARCA4 | |
| SMARCB1 | |
| SMN1 | |
| STAT3 | |
| TBP | |
| TFAP2A | |
| TFDP1 | |
| TIGAR | |
| TOP1 | |
| TOP2A | |
| TP53BP1 | |
| TP53BP2 | |
| TOP2B | |
| TP53INP1 | |
| TSG101 | |
| UBE2A | |
| UBE2I | |
| UBC | |
| USP7 | |
| USP10 | |
| WRN | |
| WWOX | |
| XPB | |
| YBX1 | |
| YPEL3 | |
| YWHAZ | |
| Zif268 | |
| ZNF148 | |
| SIRT1 | |
| circRNA_014511 |
참조
[1]
논문
Uncovering the role of p53 splice variants in human malignancy: a clinical perspective
2013-12
[2]
논문
The Guardian of the Genome Revisited: p53 Downregulates Genes Required for Telomere Maintenance, DNA Repair, and Centromere Structure
2018-05
[3]
문서
"Gene nomenclature#Vertebrate gene and protein symbol conventions|''italics'' are used to denote the ''TP53'' gene name and distinguish it from the protein it encodes"
[4]
논문
Isolation and characterization of a human p53 cDNA clone: expression of the human p53 gene
1984-12
[5]
논문
Localization of gene for human p53 tumour antigen to band 17p13
1986
[6]
논문
Identification of p53 as a sequence-specific DNA-binding protein
1991-06
[7]
논문
The gene for human p53 cellular tumor antigen is located on chromosome 17 short arm (17p13)
1986-01
[8]
서적
The p53 family
Cold Spring Harbor Laboratory Press
2010
[9]
논문
p53 Isoforms: An Intracellular Microprocessor?
2011-04
[10]
논문
Twenty years of p53 research: structural and functional aspects of the p53 protein
1999-12
[11]
웹사이트
OrthoMaM phylogenetic marker: TP53 coding sequence
http://www.orthomam.[...]
2009-12-02
[12]
논문
TP53 codon 72 polymorphism and cervical cancer: a pooled analysis of individual data from 49 studies
2009-08
[13]
논문
TP53 codon 72 polymorphism is associated with pancreatic cancer risk in males, smokers and drinkers
[14]
논문
P53 gene polymorphisms and breast cancer risk in Arab women
2011-09
[15]
논문
Effects of MDM2 promoter polymorphisms and p53 codon 72 polymorphism on risk and age at onset of squamous cell carcinoma of the head and neck
2011-09
[16]
논문
p53 codon 72 polymorphism and the risk of lung cancer in a Korean population
2011-09
[17]
논문
TP53 codon 72 polymorphism and colorectal cancer susceptibility: a meta-analysis
2011-11
[18]
논문
TP53 Arg72Pro polymorphism and endometrial cancer risk: a meta-analysis
2011-12
[19]
논문
TP53 gene polymorphism: importance to cancer, ethnicity and birth weight in a Brazilian cohort
2011-12
[20]
논문
The polymorphisms of P53 codon 72 and MDM2 SNP309 and renal cell carcinoma risk in a low arsenic exposure area
2011-12
[21]
논문
Decoding p53 tumor suppression: a crosstalk between genomic stability and epigenetic control?
https://www.nature.c[...]
2024-02-20
[22]
서적
Developmental Biology, 10th ed.
Sinauer Associates, Inc. Publishers
[23]
서적
Genes and Disease
United States National Institutes of Health
1998
[24]
논문
p14ARF links the tumour suppressors RB and p53
1998-09
[25]
간행물
Genome's guardian gets a tan started
https://www.newscien[...]
2007-03-17
[26]
논문
Central role of p53 in the suntan response and pathologic hyperpigmentation
2007-03
[27]
논문
p53 regulates cell cycle and microRNAs to promote differentiation of human embryonic stem cells
[28]
논문
Activation of p53 by nutlin leads to rapid differentiation of human embryonic stem cells
2008-09
[29]
논문
Critical role for P53 in regulating the cell cycle of ground state embryonic stem cells
2020-02
[30]
논문
HIF-2α suppresses p53 to enhance the stemness and regenerative potential of human embryonic stem cells
2012-08
[31]
논문
Context-dependent enhancement of induced pluripotent stem cell reprogramming by silencing Puma
2012-05
[32]
논문
A p53-mediated DNA damage response limits reprogramming to ensure iPS cell genomic integrity
2009-08
[33]
논문
Regulation of p53 is critical for vertebrate limb regeneration
2013-10
[34]
논문
p53: the barrier to cancer stem cell formation
2014-08
[35]
논문
Crosslink between p53 and metastasis: focus on epithelial-mesenchymal transition, cancer stem cell, angiogenesis, autophagy, and anoikis
2021-11
[36]
Review
Inhibition of tumor angiogenesis by p53: a new role for the guardian of the genome
2007-11
[37]
논문
p53 inhibits angiogenesis by inducing the production of Arresten
2012-03
[38]
논문
p53 regulates maternal reproduction through LIF
2007-11
[39]
논문
Effects of mycoplasma infection on the host organism response via p53/NF-κB signaling
2018-01
[40]
논문
Targeting mutant p53 for efficient cancer therapy
2018-02
[41]
논문
The in vivo gene expression signature of oxidative stress
2008-06
[42]
논문
Interferon responses in viral pneumonias
2020-08
[43]
논문
p53 dynamics control cell fate
2012-06
[44]
논문
MI-63: a novel small-molecule inhibitor targets MDM2 and induces apoptosis in embryonal and alveolar rhabdomyosarcoma cells with wild-type p53
2009-09
[45]
논문
Regulation of p53 stability and function by the deubiquitinating enzyme USP42
2011-11
[46]
논문
Sirt7 increases stress resistance of cardiomyocytes and prevents apoptosis and inflammatory cardiomyopathy in mice
2008-03
[47]
논문
Readout of histone methylation by Trim24 locally restricts chromatin opening by p53
2023-06
[48]
논문
p53 mutations in human cancers
https://zenodo.org/r[...]
1991-07
[49]
논문
Dissecting p53 tumor suppressor functions in vivo
2002-04
[50]
논문
p53 mutant mice that display early ageing-associated phenotypes
2002-01
[51]
논문
Restoration of p53 function leads to tumour regression in vivo
2007-02
[52]
논문
Visualization and targeted disruption of protein interactions in living cells
[53]
논문
China approves first gene therapy
2004-01
[54]
서적
HIV-1: Molecular Biology and Pathogenesis
[55]
논문
CellLine, a stochastic cell lineage simulator
2007-12
[56]
논문
Thermodynamic stability of wild-type and mutant p53 core domain
1997-12
[57]
논문
Interpretation of P53 Immunohistochemistry in Endometrial Carcinomas: Toward Increased Reproducibility
2019-01
[58]
논문
Altered p53/p16 expression is linked to urothelial carcinoma progression but largely unrelated to prognosis in muscle-invasive tumors.
[59]
논문
P53 overexpression in bladder urothelial neoplasms: new aspect of World Health Organization/International Society of Urological Pathology classification.
https://www.ncbi.nlm[...]
[60]
논문
p53-dependent expression of CXCR5 chemokine receptor in MCF-7 breast cancer cells
2015-03
[61]
논문
Dual targeting of p53 and c-MYC selectively eliminates leukaemic stem cells
2016-06
[62]
뉴스
Scientists identify drugs to target 'Achilles heel' of Chronic Myeloid Leukaemia cells
https://www.myscienc[...]
2016-06-08
[63]
논문
Deciphering the significance of p53 mutant proteins
https://linkinghub.e[...]
2024-07
[64]
논문
p53 Isoforms: An Intracellular Microprocessor?
2011-04
[65]
논문
The relative mRNA expression of p53 isoforms in breast cancer is associated with clinical features and outcome
2014-03
[66]
논문
The p53 isoform Δ133p53β promotes cancer stem cell potential
2015-04
[67]
논문
TP53 mutation hits energy metabolism and increases glycolysis in breast cancer
2016-10
[68]
논문
Oscillations and variability in the p53 system
2006-06
[69]
논문
Explaining oscillations and variability in the p53-Mdm2 system
2008-08
[70]
논문
Mathematical modelling of p53 basal dynamics and DNA damage response
2013-12
[71]
논문
"[Isolation of a plasmid clone containing the mRNA sequence for mouse nonviral T-antigen]"
[72]
논문
Molecular cloning of a cDNA specific for the murine p53 cellular tumor antigen
1983-01
[73]
논문
A single gene and a pseudogene for the cellular tumour antigen p53
[74]
논문
Human p53 cellular tumor antigen: cDNA sequence and expression in COS cells
1985-05
[75]
논문
Chromosome 17 deletions and p53 gene mutations in colorectal carcinomas
1989-04
[76]
논문
The p53 proto-oncogene can act as a suppressor of transformation
1989-06
[77]
논문
Transcriptional activation by wild-type but not transforming mutants of the p53 anti-oncogene
1990-08
[78]
논문
UV irradiation stimulates levels of p53 cellular tumor antigen in nontransformed mouse cells
1984-09
[79]
논문
Control of G1 arrest after DNA damage
1993-12
[80]
논문
Molecule of the year
1993-12
[81]
논문
The requirement for the p53 proline-rich functional domain for mediation of apoptosis is correlated with specific PIG3 gene transactivation and with transcriptional repression
1998-08
[82]
논문
LMO3 interacts with p53 and inhibits its transcriptional activity
2010-02
[83]
논문
The C terminus of p53 family proteins is a cell fate determinant
2005-03
[84]
논문
p53 contains large unstructured regions in its native state
2002-10
[85]
논문
Cell-free translations of proline-rich protein mRNAs
1982-09
[86]
논문
p53 isoforms can regulate p53 transcriptional activity
2005-09
[87]
논문
Definition of the p53 functional domains necessary for inducing apoptosis
2000-12
[88]
논문
AIMP2/p38, the scaffold for the multi-tRNA synthetase complex, responds to genotoxic stresses via p53
2008-08
[89]
논문
BRCA1-BARD1 complexes are required for p53Ser-15 phosphorylation and a G1/S arrest following ionizing radiation-induced DNA damage
2004-07
[90]
논문
Functional interaction of H2AX, NBS1, and p53 in ATM-dependent DNA damage responses and tumor suppression
2005-01
[91]
논문
ATM associates with and phosphorylates p53: mapping the region of interaction
1998-12
[92]
논문
Genetic interactions between atm and p53 influence cellular proliferation and irradiation-induced cell cycle checkpoints
1997-05
[93]
논문
Substrate specificities and identification of putative substrates of ATM kinase family members
1999-12
[94]
논문
A human protein-protein interaction network: a resource for annotating the proteome
2005-09
[95]
논문
ATF3 represses 72-kDa type IV collagenase (MMP-2) expression by antagonizing p53-dependent trans-activation of the collagenase promoter
2002-03
[96]
논문
Suppression of the STK15 oncogenic activity requires a transactivation-independent p53 function
2002-09
[97]
논문
Mitochondrial p53 activates Bak and causes disruption of a Bak-Mcl1 complex
2004-05
[98]
논문
Functional interaction of p53 and BLM DNA helicase in apoptosis
2001-08
[99]
논문
The Bloom syndrome protein interacts and cooperates with p53 in regulation of transcription and cell growth control
2001-12
[100]
논문
Functional and physical interactions between BRCA1 and p53 in transcriptional regulation of the IGF-IR gene
[101]
논문
BRCA1 regulates p53-dependent gene expression
1998-03
[102]
논문
The second BRCT domain of BRCA1 proteins interacts with p53 and stimulates transcription from the p21WAF1/CIP1 promoter
1999-01
[103]
논문
BRCA1 physically associates with p53 and stimulates its transcriptional activity
1998-04
[104]
논문
The BRCA2 gene product functionally interacts with p53 and RAD51
1998-11
[105]
논문
Regulation of BRCC, a holoenzyme complex containing BRCA1 and BRCA2, by a signalosome-like subunit and its role in DNA repair
2003-11
[106]
논문
Physical interaction of tumour suppressor p53/p73 with CCAAT-binding transcription factor 2 (CTF2) and differential regulation of human high-mobility group 1 (HMG1) gene expression
2003-04
[107]
논문
The C-terminal regulatory domain of p53 contains a functional docking site for cyclin A
2000-07
[108]
논문
Downregulation of the cdc2/cyclin B protein kinase activity by binding of p53 to p34(cdc2)
2001-05
[109]
논문
Cisplatin induces p53-dependent FLICE-like inhibitory protein ubiquitination in ovarian cancer cells
2008-06
[110]
논문
Functional interaction between BLM helicase and 53BP1 in a Chk1-mediated pathway during S-phase arrest
2004-09
[111]
논문
Radiation-induced phosphorylation of Chk1 at S345 is associated with p53-dependent cell cycle arrest pathways
[112]
논문
Cyclin G1 has growth inhibitory activity linked to the ARF-Mdm2-p53 and pRb tumor suppressor pathways
2003-01
[113]
논문
MDM2-HDAC1-mediated deacetylation of p53 is required for its degradation
2002-11
[114]
논문
p53 recruitment of CREB binding protein mediated through phosphorylated CREB: a novel pathway of tumor suppressor regulation
2000-07
[115]
논문
Regulation of CAK kinase activity by p53
1998-11
[116]
논문
p53 is phosphorylated by CDK7-cyclin H in a p36MAT1-dependent manner
1997-12
[117]
논문
Regulatory interactions between the checkpoint kinase Chk1 and the proteins of the DNA-dependent protein kinase complex
2003-08
[118]
논문
DNA end-independent activation of DNA-PK mediated via association with the DNA-binding protein C1D
1998-07
[119]
논문
Association of p14ARF with the p120E4F transcriptional repressor enhances cell cycle inhibition
2003-02
[120]
논문
p53 is involved in the p120E4F-mediated growth arrest
2000-01
[121]
논문
MBP1: a novel mutant p53-specific protein partner with oncogenic properties
1999-06
[122]
논문
The double-stranded RNA activated protein kinase PKR physically associates with the tumor suppressor p53 protein and phosphorylates human p53 on serine 392 in vitro
1999-04
[123]
논문
Physical interaction and functional antagonism between the RNA polymerase II elongation factor ELL and p53
1999-06
[124]
논문
p53 Transcriptional activity is mediated through the SRC1-interacting domain of CBP/p300
2002-03
[125]
논문
p300/MDM2 complexes participate in MDM2-mediated p53 degradation
1998-10
[126]
논문
Ordered cooperative functions of PRMT1, p300, and CARM1 in transcriptional activation by p53
2004-06
[127]
논문
Regulation of transcription of the human presenilin-1 gene by ets transcription factors and the p53 protooncogene
2000-11
[128]
논문
p53 modulation of TFIIH-associated nucleotide excision repair activity
http://repub.eur.nl/[...]
1995-06
[129]
논문
Activation of p53 or loss of the Cockayne syndrome group B repair protein causes metaphase fragility of human U1, U2, and 5S genes
2000-05
[130]
논문
A nucleolar mechanism controlling cell proliferation in stem cells and cancer cells
2002-12
[131]
논문
AMF1 (GPS2) modulates p53 transactivation
2001-09
[132]
논문
Direct, activating interaction between glycogen synthase kinase-3beta and p53 after DNA damage
2002-06
[133]
논문
Phosphorylation and hsp90 binding mediate heat shock stabilization of p53
2003-01
[134]
논문
Inhibition of MDM2 by hsp90 contributes to mutant p53 stabilization
2001-11
[135]
논문
Direct interactions between HIF-1 alpha and Mdm2 modulate p53 function
2003-04
[136]
논문
Regulation of tumor angiogenesis by p53-induced degradation of hypoxia-inducible factor 1alpha
2000-01
[137]
논문
Two sequence motifs from HIF-1alpha bind to the DNA-binding site of p53
2002-08
[138]
논문
Stabilization of wild-type p53 by hypoxia-inducible factor 1alpha
1998-03
[139]
논문
Characterization of cells and gene-targeted mice deficient for the p53-binding kinase homeodomain-interacting protein kinase 1 (HIPK1)
2003-04
[140]
논문
Regulation of p53 activity by its interaction with homeodomain-interacting protein kinase-2
2002-01
[141]
논문
Identification and characterization of HIPK2 interacting with p73 and modulating functions of the p53 family in vivo
2002-08
[142]
논문
Interaction with p53 enhances binding of cisplatin-modified DNA by high mobility group 1 protein
2001-03
[143]
논문
HMGB1 interacts with many apparently unrelated proteins by recognizing short amino acid sequences
2002-03
[144]
논문
Hsp70 family member, mot-2/mthsp70/GRP75, binds to the cytoplasmic sequestration domain of the p53 protein
2002-04
[145]
논문
The Huntington's disease protein interacts with p53 and CREB-binding protein and represses transcription
2000-06
[146]
논문
The candidate tumor suppressor ING1b can stabilize p53 by disrupting the regulation of p53 by MDM2
2002-09
[147]
논문
The candidate tumour suppressor p33ING1 cooperates with p53 in cell growth control
1998-01
[148]
논문
Two wobble-splicing events affect ING4 protein subnuclear localization and degradation
2008-10
[149]
논문
p29ING4 and p28ING5 bind to p53 and p300, and enhance p53 activity
2003-05
[150]
논문
The non-ankyrin C terminus of Ikappa Balpha physically interacts with p53 in vivo and dissociates in response to apoptotic stress, hypoxia, DNA damage, and transforming growth factor-beta 1-mediated growth suppression
2002-03
[151]
논문
A role for Hsc70 in regulating nucleocytoplasmic transport of a temperature-sensitive p53 (p53Val-135)
2001-05
[152]
논문
MdmX is a RING finger ubiquitin ligase capable of synergistically enhancing Mdm2 ubiquitination
2002-12
[153]
논문
Isolation and identification of the human homolog of a new p53-binding protein, Mdmx
https://repository.u[...]
1997-07
[154]
논문
RB18A, whose gene is localized on chromosome 17q12-q21.1, regulates in vivo p53 transactivating activity
2000-12
[155]
논문
Identification of RB18A, a 205 kDa new p53 regulatory protein which shares antigenic and functional properties with p53
1997-12
[156]
논문
JNK1, JNK2 and JNK3 are p53 N-terminal serine 34 kinases
1997-11
[157]
논문
Death-associated protein 4 binds MST1 and augments MST1-induced apoptosis
2002-12
[158]
논문
Physical and functional interactions of neuronal growth suppressor necdin with p53
1999-06
[159]
논문
Stress-dependent nucleolin mobilization mediated by p53-nucleolin complex formation
2002-08
[160]
논문
NUMB controls p53 tumour suppressor activity
2008-01
[161]
논문
ARF promotes MDM2 degradation and stabilizes p53: ARF-INK4a locus deletion impairs both the Rb and p53 tumor suppression pathways
1998-03
[162]
논문
Parc: a cytoplasmic anchor for p53
2003-01
[163]
논문
Aprataxin, a novel protein that protects against genotoxic stress
2004-05
[164]
논문
Poly(ADP-ribose) binds to specific domains of p53 and alters its DNA binding functions
1998-05
[165]
논문
Involvement of PIAS1 in the sumoylation of tumor suppressor p53
2001-09
[166]
논문
The human Cdc14 phosphatases interact with and dephosphorylate the tumor suppressor protein p53
2000-01
[167]
논문
Role of Pin1 in the regulation of p53 stability and p21 transactivation, and cell cycle checkpoints in response to DNA damage
2002-12
[168]
논문
The prolyl isomerase Pin1 reveals a mechanism to control p53 functions after genotoxic insults
2002-10
[169]
논문
Enhancement of p53-dependent gene activation by the transcriptional coactivator Zac1
2001-04
[170]
논문
Plk3 functionally links DNA damage to cell cycle arrest and apoptosis at least in part via the p53 pathway
2001-11
[171]
논문
Mammalian Polo-like kinase 3 (Plk3) is a multifunctional protein involved in stress response pathways
2002-09
[172]
논문
PACT: cloning and characterization of a cellular p53 binding protein that interacts with Rb
1997-01
[173]
논문
Prohibitin induces the transcriptional activity of p53 and is exported from the nucleus upon apoptotic signaling
2003-11
[174]
논문
Cellular stress and DNA damage invoke temporally distinct Mdm2, p53 and PML complexes and damage-specific nuclear relocalization
2003-10
[175]
논문
Regulation of p53 activity in nuclear bodies by a specific PML isoform
2000-11
[176]
논문
The function of PML in p53-dependent apoptosis
2000-10
[177]
논문
Proteasome activator PA28 gamma regulates p53 by enhancing its MDM2-mediated degradation
2008-03
[178]
논문
PTEN tumor suppressor regulates p53 protein levels and activity through phosphatase-dependent and -independent mechanisms
2003-02
[179]
논문
Nuclear FAK promotes cell proliferation and survival through FERM-enhanced p53 degradation
2008-01
[180]
논문
Human securin interacts with p53 and modulates p53-mediated transcriptional activity and apoptosis
2002-10
[181]
논문
p53 is linked directly to homologous recombination processes via RAD51/RecA protein interaction
1996-04
[182]
논문
Interaction of p53 with the human Rad51 protein
1997-10
[183]
논문
Pirh2, a p53-induced ubiquitin-protein ligase, promotes p53 degradation
2003-03
[184]
논문
Molecular basis of Pirh2-mediated p53 ubiquitylation
2008-12
[185]
논문
The interaction of p53 with replication protein A mediates suppression of homologous recombination
2004-12
[186]
논문
UV-induced DNA incision and proliferating cell nuclear antigen recruitment to repair sites occur independently of p53-replication protein A interaction in p53 wild type and mutant ovarian carcinoma cells
2001-12
[187]
논문
Ribosomal protein L11 negatively regulates oncoprotein MDM2 and mediates a p53-dependent ribosomal-stress checkpoint pathway
2003-12
[188]
논문
Inhibiting S100B restores p53 levels in primary malignant melanoma cancer cells
2004-08
[189]
논문
Covalent modification of p73alpha by SUMO-1. Two-hybrid screening with p73 identifies novel SUMO-1-interacting proteins and a SUMO-1 interaction motif
2000-11
[190]
논문
p14ARF interacts with DAXX: effects on HDM2 and p53
2008-06
[191]
논문
SWI/SNF complex interacts with tumor suppressor p53 and is necessary for the activation of p53-mediated transcription
2002-06
[192]
논문
A direct interaction between the survival motor neuron protein and p53 and its relationship to spinal muscular atrophy
2002-01
[193]
논문
High-throughput sequencing identifies STAT3 as the DNA-associated factor for p53-NF-kappaB-complex-dependent gene expression in human heart failure
2010-06
[194]
논문
Wild-type p53 binds to the TATA-binding protein and represses transcription
1992-12
[195]
논문
Pax-6 interactions with TATA-box-binding protein and retinoblastoma protein
1999-06
[196]
논문
Tumor suppressor activity of AP2alpha mediated through a direct interaction with p53
2002-11
[197]
논문
Functional interaction between DP-1 and p53
1996-10
[198]
논문
p53 and metabolism: Inside the TIGAR
2006-07
[199]
논문
The interaction between p53 and DNA topoisomerase I is regulated differently in cells with wild-type and mutant p53
1999-08
[200]
논문
Subnuclear distribution of topoisomerase I is linked to ongoing transcription and p53 status
2002-02
[201]
논문
Crystal structure of human 53BP1 BRCT domains bound to p53 tumour suppressor
2002-07
[202]
논문
Comparison of BRCT domains of BRCA1 and 53BP1: a biophysical analysis
2004-03
[203]
논문
The 8-kDa dynein light chain binds to p53-binding protein 1 and mediates DNA damage-induced p53 nuclear accumulation
2005-03
[204]
논문
Structure of the 53BP1 BRCT region bound to p53 and its comparison to the Brca1 BRCT structure
2002-03
[205]
논문
Purification, crystallization and preliminary X-ray analysis of the BRCT domains of human 53BP1 bound to the p53 tumour suppressor
2002-10
[206]
논문
Two cellular proteins that bind to wild-type but not mutant p53
1994-06
[207]
논문
The p53-binding protein 53BP2 also interacts with Bc12 and impedes cell cycle progression at G2/M
1996-07
[208]
논문
Human topoisomerase IIalpha and IIbeta interact with the C-terminal region of p53
2000-02
[209]
논문
TP53INP1s and homeodomain-interacting protein kinase-2 (HIPK2) are partners in regulating p53 activity
2003-09
[210]
논문
p53DINP1, a p53-inducible gene, regulates p53-dependent apoptosis
2001-07
[211]
논문
A TSG101/MDM2 regulatory loop modulates MDM2 degradation and MDM2/p53 feedback control
2001-02
[212]
논문
Supramolecular complex formation between Rad6 and proteins of the p53 pathway during DNA damage-induced response
2003-04
[213]
논문
Associations of UBE2I with RAD52, UBL1, p53, and RAD51 proteins in a yeast two-hybrid system
https://zenodo.org/r[...]
1996-10
[214]
논문
Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1
2002-02
[215]
논문
Identification of c-Cbl as a new ligase for insulin-like growth factor-I receptor with distinct roles from Mdm2 in receptor ubiquitination and endocytosis
2008-07
[216]
논문
The tumour suppressor RASSF1A promotes MDM2 self-ubiquitination by disrupting the MDM2-DAXX-HAUSP complex
2008-07
[217]
논문
CARPs enhance p53 turnover by degrading 14-3-3sigma and stabilizing MDM2
2008-03
[218]
논문
Hedgehog signaling overrides p53-mediated tumor suppression by activating Mdm2
2008-03
[219]
논문
Specific inhibition of Mdm2-mediated neddylation by Tip60
2008-01
[220]
논문
Deubiquitination of p53 by HAUSP is an important pathway for p53 stabilization
2002-04
[221]
논문
USP10 Regulates p53 Localization and Stability by Deubiquitinating p53
2010-02
[222]
논문
The processing of Holliday junctions by BLM and WRN helicases is regulated by p53
2002-08
[223]
논문
p53 Modulates the exonuclease activity of Werner syndrome protein
2001-09
[224]
논문
Hyaluronidase induction of a WW domain-containing oxidoreductase that enhances tumor necrosis factor cytotoxicity
2001-02
[225]
논문
The Ankrd2 protein, a link between the sarcomere and the nucleus in skeletal muscle
2004-05
[226]
논문
Direct interaction of p53 with the Y-box binding protein, YB-1: a mechanism for regulation of human gene expression
2000-12
[227]
논문
YPEL3, a p53-regulated gene that induces cellular senescence
2010-05
[228]
논문
ATM-dependent activation of p53 involves dephosphorylation and association with 14-3-3 proteins
1998-06
[229]
논문
Physical interaction between p53 and primary response gene Egr-1
2001-04
[230]
논문
ZBP-89 promotes growth arrest through stabilization of p53
2001-07
[231]
논문
MiR-34, SIRT1 and p53: the feedback loop
2009-03
[232]
논문
CircRNA_014511 affects the radiosensitivity of bone marrow mesenchymal stem cells by binding to miR-29b-2-5p
2019-05
[233]
논문
Chromosome 17 deletions and p53 gene mutations in colorectal carcinomas
1989-04
[234]
논문
The p53 proto-oncogene can act as a suppressor of transformation
1989-06
[235]
뉴스
ゾウにがんが少ない理由を解明、米研究
https://www.afpbb.co[...]
AFPBB News
2015-10-09
[236]
뉴스
How elephants avoid cancer : Pachyderms have extra copies of a key tumour-fighting gene
http://www.nature.co[...]
2015-10-08
[237]
논문
p53 mutant mice that display early ageing-associated phenotypes
[238]
서적
생명과학개론 9판
[239]
논문
MiR-34a, miR-29c and miR-17-5p are downregulated in CLL patients with TP53 abnormalities
[240]
웹인용
The p53 tumor suppressor protein
http://www.ncbi.nlm.[...]
United States National Institutes of Health
2008-05-28
[241]
논문
MicroRNAs regulate p21(Waf1/Cip1) protein expression and the DNA damage response in human embryonic stem cells.
[242]
논문
p14ARF links the tumour suppressors RB and p53
[243]
논문
p53 regulates maternal reproduction through LIF
[244]
뉴스
Genome's guardian gets a tan started
http://www.newscient[...]
New Scientist
2007-03-17
[245]
논문
Central role of p53 in the suntan response and pathologic hyperpigmentation
[246]
뉴스
잠 못자면 암 발병률 높아지는 원인 밝혀졌다
https://dongascience[...]
동아사이언스
2020-06-04
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com