감수분열
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
감수분열은 유성 생식을 하는 진핵생물의 생식 세포 분열 과정으로, 염색체 수가 절반으로 줄어든 생식 세포(정자, 난자, 홀씨 등)를 생성한다. 1876년 오스카 헤르트비히에 의해 처음 발견되었고, 1905년 J.B. 파머와 J.E.S. 무어에 의해 'meiosis'라는 용어가 처음 사용되었다. 감수분열은 감수분열 I과 감수분열 II로 나뉘며, 각 분열은 전기, 중기, 후기, 말기 단계를 거친다. 감수분열 I에서는 상동 염색체가 분리되고, 감수분열 II에서는 자매 염색분체가 분리된다. 감수분열은 유전적 다양성을 증가시키는데, 상동 염색체 간의 교차와 멘델의 독립의 법칙에 의해 유전자 조합이 다양해지기 때문이다. 감수분열의 오류는 염색체 비분리를 유발하여 다운 증후군과 같은 유전 질환을 초래할 수 있다. 감수분열은 체세포분열과 달리 4개의 딸세포를 생성하며, 생식 세포 생성과 유전적 다양성 확보에 중요한 역할을 한다.
더 읽어볼만한 페이지
- 세포 분열 - 체세포 분열
체세포 분열은 진핵세포에서 하나의 모세포가 유전적으로 동일한 두 개의 딸세포를 생성하는 과정으로, 생물의 성장, 발생, 손상된 세포 대체, 무성 생식에 중요한 역할을 하며 오류 시 질병의 원인이 될 수 있다. - 게놈 - 종분화
종분화는 하나의 종이 두 개 이상의 별개 종으로 갈라지는 진화 과정으로, 유성생식, 이소적·주변적·측지적·동소적 종분화 메커니즘, 생태적·성 선택, 강화, 다배수체 형성, 잡종 종분화, 인위적 종분화 등 다양한 요인과 과정을 통해 발생하며, 점진적 진화와 단속평형설이라는 속도에 대한 논쟁이 존재한다. - 게놈 - 염색체
염색체는 유전자를 포함하는 DNA 가닥으로, 진핵생물에서는 단백질과 결합해 염색질을 형성하고 세포 분열 시 응축된 막대 모양을 나타내며, 원핵생물은 진핵생물보다 단순한 구조를 가지고 종에 따라 수와 구조가 다양하며, 이상이 생기면 유전 질환을 유발할 수 있어 유전 연구에 중요한 역할을 한다. - 1876년 과학 - 1876년 3월 25일 일식
- 1876년 과학 - 1876년 9월 17일 일식
| 감수분열 | |
|---|---|
| 감수 분열 | |
 | |
| 기본 정보 | |
| 유형 | 세포 분열 |
| 관련 과정 | 생식 염색체 |
| 결과 | 배우자 생식 세포 |
| 시작 세포 | 2배체 |
| 최종 세포 | 반수체 |
| 딸세포 수 | 4개 |
| 염색체 수 | 반감 |
| 상세 정보 | |
| 정의 | 감수 분열은 진핵생물의 생식 세포 형성 과정에서 일어나는 특수한 세포 분열이다. |
| 목적 | 유성 생식을 위한 반수체 생식 세포 (정자 또는 난자)를 생성하는 것이다. |
| 과정 | 감수 1분열: 상동 염색체 쌍이 분리되어 두 개의 반수체 세포가 생성된다. 감수 2분열: 각 세포에서 자매 염색 분체가 분리되어 총 4개의 반수체 세포가 생성된다. |
| 유전적 다양성 | 교차: 감수 1분열에서 상동 염색체 간의 유전 물질 교환을 통해 유전적 다양성이 증가한다. 독립적 분류: 염색체가 무작위로 딸세포로 분배되어 추가적인 유전적 다양성을 제공한다. |
| 진화적 중요성 | |
| 기원 | 유성 생식의 진화에 필수적인 요소이다. |
| 다양성 증가 | 교차와 독립적 분류를 통해 유전적 다양성을 증가시킨다. |
| 진화 | 진화적 변화를 위한 재료를 제공한다. |
| 생물학적 중요성 | |
| 생식 | 유성 생식에 필수적인 과정이다. |
| 염색체 수 유지 | 다음 세대의 개체가 부모 세대와 동일한 염색체 수를 유지하도록 한다. |
| 유전적 다양성 | 유전적 다양성을 통해 자손의 적응력과 생존 가능성을 높인다. |
| 오차 | |
| 비분리 | 감수 분열 과정에서 염색체가 제대로 분리되지 못하면 이수성과 같은 염색체 이상을 유발할 수 있다. |
| 인간의 경우 | 감수 분열 오류는 다운 증후군과 같은 유전 질환의 가장 일반적인 원인 중 하나이다. |
| 기타 | |
| 로마자 표기법 | meiosis |
| 언어별 명칭 | 독일어: Meiose 영어: meiosis 그리스어: Μείωσις (Meiosis) |
2. 역사
1876년 독일의 생물학자 오스카 헤르트비히(Oscar Hertwig)가 처음으로 성게 알에서 감수분열을 발견하고 묘사했다.[87] 1883년 벨기에 동물학자 에두아르 반 베네덴(Edouard Van Beneden)은 ''아스카리스'' 속(''Ascaris'') 회충 알에서 감수분열을 염색체 수준으로 다시 묘사했다.[87] 1890년 독일의 생물학자 아우구스트 바이스만(August Weismann)은 생식과 유전 측면에서 감수분열의 중요성을 묘사했는데, 바이스만은 염색체 수가 유지되려면 두배수체 세포 하나가 홑배수체 세포 넷으로 바뀌기 위해서 두 번의 세포 분열이 필요하다고 언급했다.[87] 1911년 미국 유전학자 토머스 헌트 모건(Thomas Hunt Morgan)은 초파리 ''드로소필라 멜라노가스터''(''Drosophila melanogaster'')의 감수분열 과정에서 교차를 발견했으며, 이는 유전 형질이 염색체에서 전달된다는 사실을 확립해주었다.[87]
감수분열은 감수분열 I과 감수분열 II로 나뉘며, 각각 핵분열과 세포질분열로 구성된다. 감수분열을 준비하는 단계는 체세포분열 주기의 사이기와 패턴 및 명칭이 동일하다.[89] 사이기는 다음과 같이 3단계로 나뉜다.
'meiosis'라는 용어는 '줄어듦'을 뜻하는 그리스어 μείωσις에서 유래하였다. J.B. 파머(J.B. Farmer)와 J.E.S. 무어(J.E.S. Moore)는 1905년에 이 용어를 'maiosis'라고 독특하게 번역하여 처음으로 생명과학 분야에서 사용하기 시작했다.[87]
>''플레밍이 '헤테로타입'과 '호모타입'이라고 정의한 두 번의 분열에서 나타나는 핵 변화의 전체 과정을 아울러 우리는 Maiosis나 Maiotic phase라는 용어를 사용할 것을 제안한다''.
이후 코니케(Koernicke; 1905), 그리고 판텔과 드 시네티(Pantel and De Sinety; 1906)는 그리스어 표기 원칙에 따라 철자를 'meiosis'라고 바꾸었다.[88]
일본은 20세기 초반 'meiosis'의 어원을 살려 '수가 줄어드는 세포 분열'이라는 의미에서 '減數分裂'(감수분열)이라는 한자어로 번역했고, 일제 강점기와 그 이후 우리나라가 이 한자어를 그대로 음독하여 표기함으로써 '감수분열'이라는 용어가 만들어졌다.
3. 감수분열의 세포 주기
사이기 이후 감수분열 I이 진행되고, 그 이후 감수분열 II가 진행된다. 감수분열 I 때는 상동 염색체가 분리되어 각 딸세포 2개로 이동하고, 각 염색체는 여전히 2개의 자매 염색분체로 이루어진다. 따라서 이때 염색체 수가 절반으로 감소한다. 감수분열 II 동안에는 자매 염색분체끼리 분리되어 딸염색체가 4개의 딸세포로 각각 나뉘어 들어간다. 두배수체 생물의 경우 감수분열로부터 만들어진 딸세포는 홑배수체이고, 각 염색체 사본은 1개씩만 포함한다. 일부 종들은 감수분열 I과 감수분열 II 사이에 중간기(interkinesis)라는 휴식기를 거친다.
감수분열 I과 II는 체세포분열의 세포 주기와 비슷하게 각각 전기, 중기, 후기, 말기 단계로 나뉜다. 따라서 감수분열은 감수분열 I(전기 I, 중기 I, 후기 I, 말기 I)과 감수분열 II(전기 II, 중기 II, 후기 II, 말기 II)로 구성된다.
감수분열 과정 동안 특정 유전자에서 전사가 활발하게 일어난다.[90][91] 감수분열 단계에 특이적인 mRNA 발현과 더불어, 전반적인 번역 조절(예: 미리 형성된 mRNA의 선택적 사용)도 일어나기 때문에 감수분열 시기 동안 감수분열 단계에 특이적인 단백질 발현이 조절된다.[92] 이러한 전사 및 번역 조절은 감수분열을 수행하는 데 적절하게끔 세포를 전체적으로 재구조화한다.
감수분열 I 때 2가 염색체를 형성하고 있던 상동 염색체가 분리된다. 그 결과 염색분체 쌍을 각각 가지는 2개의 홑배수체 세포가 만들어진다. 배수성이 두배수체에서 홑배수체로 줄어들기 때문에 감수분열 I은 '감소 분열'이라고도 불린다. 감수분열 II는 자매 염색분체가 분리되어 4개의 홑배수체 딸세포를 만들기 때문에 체세포분열과 마찬가지로 '균등 분열'이다.
3. 1. 감수분열 I
감수분열 I은 상동 염색체가 분리되어 염색체 수가 절반으로 줄어드는 '감소 분열'이다. 이때 2가 염색체(4분 염색체)가 형성되고 상동 재조합이 일어난다.[14]
감수분열은 유사분열과 관련된 세포 분열 과정이지만, 다음과 같은 두 가지 중요한 차이점이 있다.
| 감수분열 | 유사분열 | |
|---|---|---|
| 재조합 | 각 염색체 쌍(각 부모로부터 하나씩 받음) 사이의 유전자를 섞어, 모든 배우자에 고유한 유전자 조합을 가진 많은 재조합 염색체를 생성한다. | DNA 손상을 복구해야 할 경우에만 발생한다. 일반적으로 동일한 자매 염색분체 사이에서 발생하며 유전적 변화를 초래하지 않는다. |
| 염색체 수 (배수성) | 부모 세포의 염색체 수의 절반을 가진 유전적으로 고유한 네 개의 세포를 생성한다. | 부모 세포와 동일한 수의 염색체를 가진 유전적으로 동일한 두 개의 세포를 생성한다. |
감수분열은 각 염색체의 두 개의 복사본을 포함하는 이배체 세포(상동 염색체)로 시작한다. 먼저 세포는 DNA 복제를 거치므로 각 상동 염색체는 두 개의 동일한 자매 염색분체로 구성된다. 그런 다음 각 상동 염색체 쌍은 서로 짝을 이루고 상동 재조합을 통해 유전 정보를 교환하여 교차를 유발한다.
감수분열 I에서 방추사에 의해 상동 염색체가 분리되어 딸세포로 나뉜다. 그 후 세포는 DNA 복제 과정 없이 감수분열 II로 진행한다.
이배체 세포에서는 특정 염색체에 대해 매우 유사한 또 다른 염색체가 존재하는데, 이들을 '''상동염색체'''라고 부른다. 상동염색체 중 하나는 모계로부터, 다른 하나는 부계로부터 물려받는다.
감수분열에 앞서 세포는 DNA 복제를 수행하여 DNA량을 두 배로 증가시킨다. 그 결과, 원래 염색체와 같은 배열을 가진 두 개의 염색체가 형성된다. 이러한 동일한 유전 정보를 가진 두 개의 염색체 쌍을 '''자매염색분체'''라고 부른다.
'''감수 제1분열'''에서는 두 개의 염색분체로 구성된 상동염색체들이 서로 접합하여 네 개의 염색분체로 구성된 '''이가염색체'''를 형성한다. 그 후, 각각의 상동염색체(두 개의 염색분체)는 서로 다른 방향으로 분리되고 제1분열이 종료된다.
감수 제1분열 전기에는 상동염색체 간에 '''교차'''가 일어나 일부 배열을 교환한다('''재조합'''). 상동염색체가 교차한 부위에 형성되는 구조를 '''키아즈마'''라고 부른다.
3. 1. 1. 전기 I
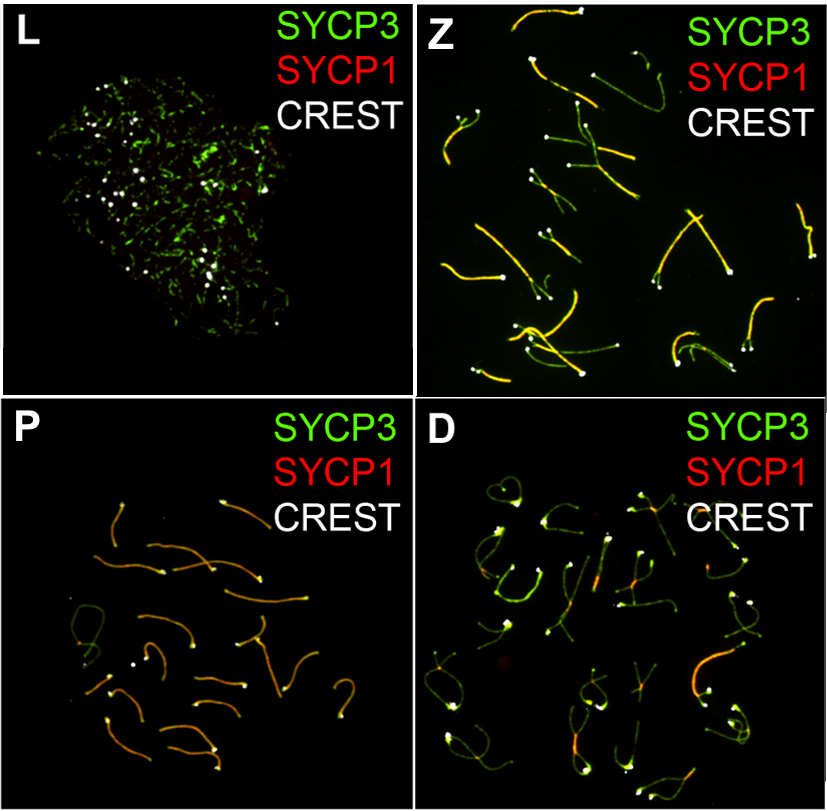

전기 I은 감수분열에서 가장 긴 시기이다(총 14일 동안 진행되는 생쥐의 감수분열 중 전기 I은 13일 동안 지속된다[93]). 전기 I 동안 아버지와 어머니로부터 물려받은 상동 염색체 쌍이 서로 이어지고(시냅스), 유전 정보가 교환되며(상동 재조합으로), 염색체 1쌍당 최소 1개 이상의 교차가 형성된다.[94] 이때 교차점(chiasma)을 통해 교차를 관찰할 수 있다.[95] 교차 과정은 상동 염색체가 안정한 쌍을 형성할 수 있도록 해주며, 이에 따라 첫 감수분열 때 염색체가 정확하게 분리될 수 있도록 해준다. 각 부모로부터 유래하여 복제된 뒤 서로 쌍을 이룬 염색체를 2가 염색체(bivalent; 염색체 2개) 또는 4분 염색체(tetrad; 염색분체 4개)라고 부른다. 전기 I은 염색체의 모양에 따라 가는섬유기, 접합기, 굵은섬유기, 겹섬유기, 이동기의 5단계로 세분화된다.
감수분열은 배우자 형성에서 유전적 다양화를 일으켜 환경 변화에 대한 적응과 진화에 기여한다고 생각된다. 예를 들어 2쌍 4개의 염색체를 가진 생물에서는 22=4가지의 조합을 가진 배우자가 만들어지고, 여기서 얻어지는 차세대는 42=16가지이다. 인간의 경우에는 23쌍의 상동염색체, 총 46개의 염색체를 가지므로, 223=8,388,608(약 840만)가지의 배우자, 8,388,6082=70,368,744,177,664(약 70조)가지의 차세대가 생길 가능성이 있다. 더 나아가 유전적 다양성을 만들어내는 기전에는 염색체의 일부가 교환되는 교차(상동 재조합)가 있다. 교차는 감수 제1분열 시에 일어난다. 분자 기전으로는 DNA의 이중나선 절단이 일어나고, 그것이 DNA 복구에 의해 다시 연결될 때 발생한다.
3. 1. 2. 중기 I
상동 염색체 쌍은 함께 중기판으로 이동한다. 세포 양쪽의 방추 극에서 뻗어 나온 방추부착점 미세관은 상동 염색체 쌍 각각의 방추부착점에 결합하고, 이후 상동 염색체 쌍은 방추를 이등분하면서 적도판을 따라 배열된다.[95] 이때 상동 염색체의 두 방추부착점에 연결된 미세관에 의해 2가 염색체에 균형을 잡아주는 힘이 계속 가해진다. 이러한 부착을 두극 부착(bipolar attachment)이라고 한다. 동일한 적도판을 따라 2가 염색체들이 배열될 때 각 2가 염색체는 중기판에 무작위로 배열되는데, 이는 멘델이 밝힌 독립 유전의 원리를 설명하는 물리적 근거이다.[17] 단백질 복합체인 코히신은 염색체가 복제되는 순간부터 후기 I까지 자매 염색분체를 함께 붙들어둔다.3. 1. 3. 후기 I
방추사가 짧아지면서 상동 염색체(각각 자매 염색분체 쌍으로 이루어짐)를 세포 반대편 극으로 끌어당긴다. 극성 미세관은 길어지면서 중심체를 더 멀리 밀어내고, 그 결과 세포는 중심으로부터 길어진다.[95] 체세포분열과 다르게, 염색체 팔에 결합된 코히신만 분해되고, 중심절을 둘러싸는 코히신은 슈고신(Shugoshin; '수호신'의 일본어)이라는 단백질에 의해 보호받으며 그대로 남아 있다.[109] 이 때문에 자매 염색분체는 상동 염색체가 분리되는 순간에 함께 붙어 있을 수 있다.3. 1. 4. 말기 I
염색체가 양극에 도달하면서 감수분열 I이 사실상 끝난다. 각 딸세포는 이제 염색체 수가 절반이지만, 각 염색체는 염색분체 쌍으로 이루어져 있다. 방추사를 구성하던 미세관은 사라지고, 새로운 핵막이 각 홑배수체 세트를 둘러싸며 생성된다. 염색체는 다시 염색질로 풀린다. 그러나 경우에 따라 핵막은 다시 형성되지 않을 수도 있고, 염색체도 사이기만큼 완전히 풀리지 않는다.동물 세포의 경우 핵막을 집으면서, 식물 세포의 경우 세포벽을 형성하면서 세포질분열이 일어나고, 그 결과 2개의 딸세포 형성이 완료된다. 그러나 세포질분열이 완전히 완료되지 않으면 딸세포끼리 '세포질다리'로 연결되면서 감수분열 II가 끝날 때까지 딸세포는 서로 세포질을 공유한다.[110] 자매 염색분체는 말기 I 동안에도 붙어 있다.
세포는 휴식하는 시기인 중간기(interkinesis; 사이기 II라고도 부름)에 돌입할 수도 있다. 이 시기에는 DNA 복제가 일어나지 않는다.
3. 2. 감수분열 II
감수분열 II는 두 번째 감수분열로, 자매 염색분체가 분리되는 ''균등 분열''이다. 이 과정은 체세포분열과 비슷하지만, 유전적 결과는 근본적으로 다르다. 감수분열 I에서 생성된 두 개의 홑배수체 세포로부터 총 네 개의 홑배수체 세포가 만들어진다. 감수분열 II는 전기 II, 중기 II, 후기 II, 말기 II로 나뉜다.3. 2. 1. 전기 II
핵소체와 핵막이 사라지고, 염색체가 응축하여 짧고 굵어진다. 중심체는 세포 양극으로 이동하고, 두 번째 감수분열을 하기 위해 방추사를 배열한다.[111]3. 2. 2. 중기 II
중기 II에서는 각 중심체로부터 뻗어 나온 방추사가 각 염색체의 방추부착점에 부착된다. 새로 형성되는 적도판은 감수분열 I의 적도판과 비교했을 때 90° 회전된 상태이다.[111]3. 2. 3. 후기 II
중심절 주변에 남아 있던 코히신이 더 이상 슈고신의 보호를 받지 못하게 되면서 분해되어, 자매 염색분체가 분리된다. 이때 자매 염색분체였던 것은 이제 딸염색체가 되어 서로 반대편 극으로 이동한다.[109]3. 2. 4. 말기 II
핵막이 다시 형성되고 염색체는 풀리면서 길어진다. 방추체는 해체된다. 세포질분열이 일어나 홑배수체 염색체 세트를 가지는 네 개의 딸세포가 생성된다.[109] 이로써 감수분열이 완료된다.4. 감수분열의 기원과 기능
감수분열은 진핵생물의 유성 생식 과정에서 핵심적인 역할을 한다. 이 과정은 체세포분열과 유사하지만, 몇 가지 중요한 차이점을 보인다. 다음 표는 감수분열과 체세포분열의 주요 차이점을 비교한 것이다.
| 특징 | 감수분열 | 체세포분열 |
|---|---|---|
| 재조합 | 각 쌍의 염색체 사이에서 유전자를 섞어 고유한 유전적 조합을 가진 재조합 염색체를 만든다. | DNA 손상 복구가 필요한 경우에만 일어나며, 일반적으로 유전자 변화가 없다. |
| 염색체 수 | 모세포 염색체의 절반을 가진 4개의 딸세포를 생성한다. | 모세포와 똑같은 수의 염색체를 가진 2개의 딸세포를 생성한다. |
감수분열은 상동 염색체를 가지는 두배수체 세포에서 시작하여, DNA 복제를 거친 후 상동 재조합을 통해 유전 물질을 교환한다. 이 과정에서 교차가 일어나 상동 염색체 사이에 물리적인 연결이 나타나기도 한다. 첫 번째 감수분열에서는 상동 염색체가 분리되고, 두 번째 감수분열에서는 DNA 복제 없이 자매 염색분체가 분리되어 총 4개의 홑배수체 세포가 생성된다. 암컷 동물의 경우, 이 과정에서 약간의 변형을 통해 하나의 큰 난자와 2~3개의 작은 극체가 생성된다.
이러한 재조합 과정은 각 염색분체가 부모의 유전 정보가 섞인 새로운 조합을 가지게 하여, 자손은 부모와 유전적으로 다르게 만든다. 또한 각 생식자는 온전한 부모의 염색체와 재조합된 염색체의 조합으로 이루어져 유전적 다양성을 더욱 증가시킨다. 이러한 유전적 다양성은 자연 선택의 원동력 중 하나이다.
감수분열은 아키아나 박테리아에서는 일어나지 않으며, 이들은 이분법을 통한 무성 생식을 한다. 그러나 수평 유전자 전달이라는 과정을 통해 유전 물질을 교환하고 재조합하기도 한다.
감수분열은 진핵생물에서 공통적으로 나타나며, 단세포 생물인 효모에서부터 사람과 같은 다세포 생물까지 다양한 생물에서 관찰된다. 진핵생물은 약 22억 년 전에 원핵생물로부터 진화했으며, 초기 진핵생물은 단세포 생물과 유사했을 것으로 추정된다.
감수분열 동안 DNA 재조합은 돌연변이와 함께 유전적 다양성을 일으키는 주요 원인이며, 이는 유리한 대립유전자 조합을 만들어낸다. 감수분열은 독립 분리의 법칙과 교차를 통해 생식자의 유전적 다양성을 높인다.[112][113]
유전적 재조합은 DNA 복구 과정으로 볼 수 있으며, 감수분열 중에는 자손에게 전달되는 유전체 DNA 손상을 복구하는 적응으로 간주된다.[44][45] 실험에 따르면 감수분열의 주요 이점은 상동 재조합을 통한 DNA 손상의 생식세포 복구이다.[46] 예를 들어, 과산화수소 처리나 열충격은 효모나 녹조류에서 감수분열을 유도하며, 이는 산화 스트레스로 인한 DNA 손상 복구와 관련이 있을 수 있다.[47][48][49]
감수분열은 환경 변화에 대한 적응과 진화에 기여하는 유전적 다양성을 만들어낸다. 예를 들어, 2쌍의 염색체를 가진 생물은 16가지, 인간은 약 70조 가지의 다양한 자손을 생성할 수 있다. 또한, 교차는 감수 제1분열에서 DNA 이중나선 절단 후 복구를 통해 일어나며 유전적 다양성을 더욱 증가시킨다.
감수분열에서 염색체가 정상적으로 분배되지 않으면 염색체 비분리가 일어나 다운 증후군과 같은 염색체 이상 질환의 원인이 된다.
4. 1. 전기 I 정지
암컷 포유류와 조류는 난모세포가 미래의 배란에 필요한 모든 것들을 포함한 채로 태어나며, 이 난모세포는 감수분열 전기 I에 정지된 상태에 있다.[114] 사람 여성의 경우, 난모세포는 임신 3~4달 사이 태아 시기 때 형성되며, 태어날 때부터 이미 가지고 있다. 이렇게 전기 I이 정지된 단계(연장기; dictyate)는 수십 년 동안 지속될 수 있으며, 4개의 지놈 사본이 난모세포에 존재하고 있다. 난모세포가 지놈 사본이 4개인 상태로 정지되어 있으면, 여분의 유전 정보를 사용해서 생식계 DNA에 손상이 일어났을 때 복구할 수 있다.[80] 이 복구 과정은 상동 재조합 복구 메커니즘으로 나타난다.[80][115] 전기 I 때 정지된 난모세포는 외부 요인에 의한 이중가닥 절단과 같은 DNA 손상을 효율적으로 복구할 수 있는 능력이 매우 뛰어나다. DNA 복구 능력은 여성 생식계에서 중요한 품질 관리 메커니즘으로 작동하고, 여성의 생식 능력을 결정한다.[80]5. 감수분열의 발생
감수분열은 체세포분열과 유사하지만, 다음과 같은 두 가지 중요한 차이점이 있다.
| 특징 | 감수분열 | 체세포분열 |
|---|---|---|
| 재조합 | 각 쌍의 상동 염색체 사이에서 유전자를 섞어 모든 생식자가 고유한 유전적 조합을 가지는 재조합 염색체를 만든다. | DNA 손상 복구가 필요한 경우에만 일어나며, 일반적으로 동일한 자매 염색분체 사이에서 재조합이 일어나 유전적 변화가 나타나지 않는다. |
| 염색체 수 | 유전적으로 서로 다른 4개의 딸세포를 생성하며, 각 딸세포는 모세포 염색체의 절반만큼의 염색체를 가진다. | 유전적으로 똑같은 2개의 딸세포를 생성하며, 각 딸세포는 모세포 염색체와 똑같은 수의 염색체를 가진다. |
감수분열은 각 염색체의 사본을 2개씩 가지는 이배체 세포에서 시작한다. 먼저 DNA 복제를 통해 각 상동 염색체는 동일한 자매 염색분체 2개로 이루어진다. 이후 각 상동 염색체 세트는 상동 재조합을 통해 유전 물질을 서로 교환하며, 이때 종종 상동 염색체 쌍 사이에 물리적인 연결(교차)이 나타난다. 첫 번째 감수분열에서 방추사에 의해 상동 염색체가 분리된다. 그 이후 세포는 DNA 복제를 추가로 하지 않고 바로 두 번째 분열을 진행한다. 이때 자매 염색분체가 분리되면서 총 4개의 홑배수체 세포가 생성된다. 암컷 동물은 이 패턴과 약간 다른 방법을 사용하여 감수분열 결과 큰 난자 1개와 작은 극체 2~3개를 생성한다.
재조합으로 인해 각 염색분체는 어머니와 아버지의 유전 정보가 서로 섞인 새로운 조합을 포함할 수 있기 때문에 자손은 부모와 유전적으로 다르다. 뿐만 아니라 각 생식자는 재조합이 일어나지 않은 온전한 어머니 염색체, 온전한 아버지 염색체, 재조합 염색체 조합으로 이루어질 수도 있다. 이렇게 유성 생식으로부터 형성되는 유전적 다양성은 자연 선택이 작동할 수 있는 원동력 중 하나다.
감수분열은 진핵생물에서 하나의 세포가 2개의 동일한 딸세포로 분열하는 체세포분열의 메커니즘을 많이 이용한다. 일부 식물과 균류, 원생생물의 감수분열은 홀씨(포자)를 만드는데, 홀씨는 수정 없이도 스스로 분열하면서 성장할 수 있는 홑배수체 세포이다. 브델로이데아 목(Bdelloidea)과 같은 일부 진핵생물은 감수분열을 수행할 능력이 없는 대신 처녀생식을 통해 생식할 능력을 획득하였다.
감수분열은 고균이나 세균에서는 일어나지 않으며, 이들은 일반적으로 이분법을 통해 무성 생식한다. 그러나 수평적 유전자 전달이라는 과정을 통해 '유성' 과정을 수행할 수도 있는데, 이 경우 한 박테리아나 고균으로부터 다른 박테리아나 고균으로 DNA가 전달되고, 서로 다른 세포에서 유래한 DNA 분자가 재조합된다.
5. 1. 생활 주기에서의 감수분열
감수분열은 유성 생식과 관련하여 진핵생물의 생활 주기에서 나타나며, 체세포분열에 의한 성장, 감수분열, 수정 과정이 주기적으로 반복된다. 다세포 생물의 경우, 생명체가 성장할 때 두배수체와 홑배수체 전환 사이에 중간 단계가 존재한다. 생활 주기의 특정 단계에서 종자세포는 생식자를 만든다. 체세포는 생명체의 몸을 구성하고, 생식자 생성에는 관여하지 않는다.홑배수체 상태와 두배수체 상태가 교대로 나타나면서 감수분열과 수정이 주기적으로 반복된다. 생물은 생활 주기의 대부분을 두배수체 상태로 보낼 수도 있고(''두배수체 생활 주기''), 홑배수체 상태로 보낼 수도 있고(''홑배수체 생활 주기''), 둘 다로 보낼 수도 있다(''홑두배수체 생활 주기'', 여기에는 홑배수체 상태와 두배수체 상태의 2가지 구별되는 시기가 존재함). 이 관점으로 보면, 유성 생식을 이용하는 생활 주기는 생물 시기의 위치에 따라 세 종류로 구별된다.
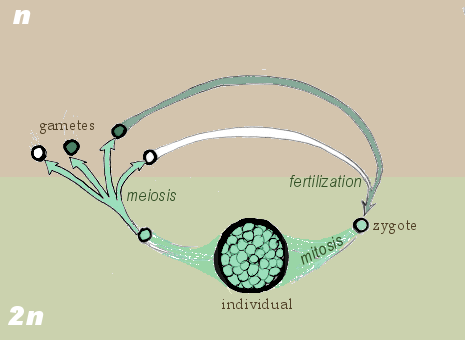
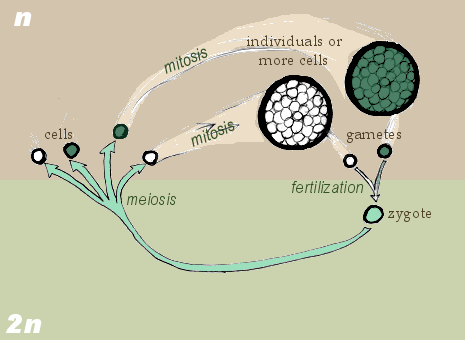
- '''두배수체 생활 주기'''(생식자 만들기 전에 감수분열): 생물체는 두배수체이고, 접합자(zygote)라고 불리는 두배수체 세포로부터 성장한다. 사람도 두배수체 생활 주기로 살아간다. 생물의 두배수체 생식계 줄기세포는 감수분열을 통해 홑배수체 생식자(수컷은 정자, 암컷은 난자)를 만들며, 생식자는 수정하여 접합자를 형성한다. 두배수체 접합체는 다시 체세포분열을 반복하면서 생명체로 성장한다.
- '''홑배수체 생활 주기'''(접합자 만든 이후 감수분열): 생명체는 홑배수체이고, 생식자(gamete)라고 불리는 하나의 홑배수체 세포가 증식하고 분화함으로써 발생한다. 서로 다른 성을 가진 두 생명체에서 나온 홑배수체 생식자는 두배수체 접합자를 형성한다. 접합자는 만들어진 즉시 감수분열을 하여 4개의 홑배수체 세포를 생성한다. 홑배수체 생식자는 다시 체세포분열을 하며 성장해서 하나의 생명체를 만든다. 많은 균류와 많은 원생동물이 홑배수체 생활 주기로 살아간다.
- '''홑두배수체 생활 주기'''(홀씨 감수분열이나 중간 감수분열): 생명체는 홑배수체 상태와 두배수체 상태를 서로 교대한다. 결과적으로 이 주기는 세대 교대라고 알려져 있다. 두배수체 생물의 생식계 세포는 감수분열을 통해 홑배수체 홀씨(spore)를 만든다. 홀씨는 체세포분열을 통해 증식해서 홑배수체 생명체로 성장한다. 이후 홑배수체 생명체의 생식자가 다른 홑배수체 생명체의 생식자와 서로 합쳐지면 다시 두배수체 생명체가 된다. 이처럼 홑두배수체 생활 주기는 두배수체 생활 주기와 홑배수체 생활 주기의 융합으로 여겨질 수 있다.[116]
5. 2. 식물과 동물에서의 감수분열
동물은 감수분열을 통해 생식자를 직접 만든다. 그러나 육상 식물과 일부 조류는 세대 교대를 통해 홑배수체 홀씨(포자)를 생성하며, 이 홀씨는 체세포분열을 거쳐 홑배수체 배우체로 발달한다. 배우체는 추가적인 감수분열 없이 직접 생식자를 만든다. 동물과 식물 모두 생식자가 융합하여 염색체 수를 원래대로 회복한다.[117]5. 3. 포유류에서의 감수분열
암컷의 경우 난모세포에서 감수분열이 일어난다. 각 일차난모세포는 감수분열을 통해 두 번 나뉘는데, 각 분열마다 불균등하게 나뉜다. 첫 번째 분열에서는 딸세포 하나와 그보다 훨씬 작은 극체 하나가 만들어지는데, 극체는 두 번째 분열을 진행할 수도 있고 안 할 수도 있다. 감수분열 II에서 딸세포가 두 번째로 분열하면서 두 번째 극체와 하나의 홑배수체 세포를 생성하는데, 이 세포는 거대해져서 난자가 된다.[118] 그러므로 암컷의 일차 난모세포가 감수분열을 하면 하나의 성숙한 난자와, 1~2개의 극체가 만들어진다.암컷은 감수분열 시기 중간에 감수분열이 중단된다. 성숙한 난모세포는 감수분열 I의 전기 I 때 정지해 있고, 난포세포라는 체세포 보호 껍질에 싸여 휴면 상태에 놓인다. 각 월경 주기가 시작될 때, 뇌하수체 앞엽에서 분비되는 난포 자극 호르몬은 약간의 난포를 난포발생이라는 과정으로 성숙하게 만든다. 이 과정 동안 난모세포는 감수분열을 다시 시작하여 감수분열 II의 중기 II까지 계속한 뒤, 배란 직전에 중기 II에서 다시 멈춘다. 사람의 난포발생 과정 동안 주로 하나의 난포만 선택되어 발생하고, 나머지 난포는 폐쇄된다. 암컷의 감수분열은 난자발생이라는 과정을 거쳐 진행되는데, 연장기(dictyate) 단계라고 알려진 긴 시간 동안 감수분열이 중단된다는 점과, 중심체 도움 없이 진행된다는 점에서 일반적인 감수분열과 다르다.[118][119]
수컷은 고환의 정세관에서 정자발생 과정을 통해 감수분열이 일어난다. 정자발생 동안 일어나는 감수분열은 정모세포라는 특정 종류의 세포에서만 일어나며, 정모세포는 성숙하여 정자가 된다. 원시종자세포의 감수분열은 암컷보다 한참 늦은 사춘기 때 일어난다. 사춘기 이전까지 수컷의 고환 조직은 감수분열을 촉진하는 물질인 레티노산을 분해시킴으로써 감수분열을 억제한다. 사춘기가 되면 정세관에 있는 버팀세포(세톨리 세포)가 스스로 레티노산을 만들기 시작하면서 감수분열이 진행된다. 레티노산에 대한 감수성은 나노스(nanos)와 DAZL라고 불리는 단백질에 의해서도 조절된다.[120][121] 레티노산 생산 효소의 유전자 기능상실 연구를 통해 레티노산은 출생 이후 정조세포의 분화를 촉진하여 정모세포가 정자가 되기 위해 필요하다는 사실이 밝혀졌지만, 레티노산은 감수분열이 시작되는 동안 필요한 것은 아니다.[124]
암컷 포유류는 원시종자세포가 배아의 난소로 이동한 이후 즉시 감수분열을 시작한다. 일부 연구는 원시 콩팥(중간콩팥)에서 유래한 레티노산이 암컷 배아 난소에 있는 난조세포의 분화를 촉진시키고, 수컷 배아 정소 조직은 레티노산을 분해함으로써 감수분열을 억제한다는 사실을 밝혔다.[122] 그러나 레티노산 생산 효소의 유전자 기능상실 연구는 레티노산이 배아 발생 동안 진행되는 암컷 감수분열이나[123] 출생 이후 시작되는 수컷 감수분열[124] 둘 다에 필요하지 않다는 사실을 밝혔다.
5. 4. 편모류에서의 감수분열
대부분의 진핵생물은 감수분열 동안 총 2번 분열하지만(때로는 교차 없이), 바퀴벌레 ''Cryptocercus''의 장에 서식하는 파라바살리드류나 옥시모나드류 같은 일부 편모류는 감수분열 동안 총 1번 분열하는 매우 드문 감수분열을 진행한다.6. 사람 유전과 질병에서의 역할
감수분열 과정에서 생성되는 새로운 DNA 조합은 돌연변이와 함께 중요한 유전적 변이의 원천이며, 이는 유익할 수 있는 새로운 대립 유전자 조합을 만든다. 감수분열은 두 가지 방식으로 배우자의 유전적 다양성을 생성한다. (1) 독립 분리의 법칙. 제1중기 동안 상동 염색체 쌍의 중기판을 따라 독립적인 배향과 제2중기에서 자매 염색분체의 배향, 그리고 제1 및 제2 후기에 상동 염색체와 자매 염색분체의 분리는 각 딸세포(그리고 궁극적으로 배우자)에 염색체가 무작위적이고 독립적으로 분포되도록 한다.[38] (2) 교차. 제1전기에 상동적인 재조합에 의한 상동 염색체 영역의 물리적 교환은 염색체 내에 새로운 유전 정보 조합을 만든다.[39]
23쌍의 인간 염색체 간 재조합은 실제 염색체뿐만 아니라 각 염색체의 일부까지 재분배하는 역할을 한다. 여성의 재조합은 남성보다 약 1.6배 더 많이 일어나는 것으로 추정된다. 또한, 평균적으로 여성의 재조합은 동원체에서 더 높고, 남성의 재조합은 텔로미어에서 더 높다. 평균적으로 100만 bp(1 Mb)는 1 cMorgan(cm = 재조합 빈도 1%)에 해당한다.[60]
감수분열에서 염색체가 정상적으로 분배되지 않는 현상은 염색체 비분리라고 불린다. 염색체 비분리는 염색체 수가 다른 배우자를 만들어내고, 다운 증후군 등 염색체 이상 질환의 원인이 된다.
6. 1. 비분리
감수분열 I에서 염색체가 정상적으로 나뉘거나, 감수분열 II에서 자매 염색분체가 나뉘는 현상을 '분리'라고 한다. 그러나 분리가 정상적으로 일어나지 않을 경우 '비분리'라고 부른다.[127] 비분리 결과 생식자는 특정 염색체 수보다 더 많거나 더 적은 염색체를 가지게 되고, 이는 세염색체증이나 홑염색체증을 일으키는 흔한 메커니즘이다. 비분리는 세포 생식 시기인 감수분열 I이나 감수분열 II 때 일어날 수도 있고, 체세포분열 동안 일어날 수도 있다.대부분 홑염색체증이나 세염색체증을 가진 사람 배아는 생존하지 못한다. 그러나 21번 염색체처럼 작은 염색체의 세염색체증과 같은 일부 비정배수성은 생존할 수 있다. 이런 비정배수성의 표현형은 심각한 발생에서부터 무증상까지 다양하다. 의학적으로 아래와 같은 예들이 있다.
- 다운 증후군 - 21번 염색체의 세염색체증
- 파타우 증후군 - 13번 염색체의 세염색체증
- 에드워드 증후군 - 18번 염색체의 세염색체증
- 클라인펠터 증후군 - 남성에서 X 염색체가 추가로 있음 - XXY, XXXY, XXXXY 등
- 터너 증후군 - 여성에서 X 염색체가 하나 없음 - XO
- 트리플 X 증후군 - 여성에서 X 염색체가 추가로 있음
- 제이콥스 증후군 - 남성에서 Y 염색체가 추가로 있음
사람 난모세포에서 비분리가 일어날 가능성은 여성의 나이가 증가함에 따라 커지는데,[62] 이는 시간이 지나면서 코히신이 손실되기 때문이다.[63]
7. 체세포분열과 비교
| 감수분열 | 체세포분열 | |
|---|---|---|
| 최종 결과 | 일반적으로 4개의 딸세포, 각각 모세포 염색체 수의 절반을 가짐 | 2개의 딸세포, 모세포와 염색체 수가 똑같음 |
| 기능 | 유성 생식을 하는 진핵생물에서 생식자를 생성함 (두배수체 생활주기) | 세포 증식, 성장, 재생, 무성 생식 |
| 일어나는 위치 | 거의 모든 진핵생물 (동물, 식물, 진균, 원생생물);[130][131] 생식샘에서, 생식자가 되기 전 (두배수체 생활주기); 접합자가 된 이후 (홑배수체 생활주기); 홀씨(포자)가 되기 전 (홑두배수체 생활주기) | 모든 진핵생물의 모든 증식하는 세포에서 |
| 단계 | 전기 I, 중기 I, 후기 I, 말기 I, 전기 II, 중기 II, 후기 II, 말기 II | 전기, 전중기, 중기, 후기, 말기 |
| 모세포와 유전적으로 동일한가? | 아니오 | 예 |
| 교차가 일어나는가? | 예, 일반적으로 상동 염색체의 각 쌍 사이에서 일어남 | 매우 드묾 |
| 상동 염색체가 짝을 이루는가? | 예 | 아니오 |
| 세포질분열 | 말기 I과 말기 II 때 일어남 | 말기 때 일어남 |
| 중심절 분리 | 후기 I 때 안 일어나고 후기 II 때 일어남 | 후기 때 일어남 |
8. 분자 조절
감수분열의 분자 조절 메커니즘은 아직 완전히 밝혀지지 않았지만, 여러 요인이 관여하는 것으로 알려져 있다. 성숙촉진인자(maturation promoting factor) (MPF)는 개구리 난모세포의 감수분열에서 특정 역할을 하는 것으로 보이며, MeiRNA는 진균류의 일종인 ''Schizosaccharomyces pombe''에서 감수분열을 시작하도록 돕는다.[132]
효모의 CEP1 유전자 산물은 감수분열 I 동안 염색체가 짝을 이루도록 돕는 역할을 할 수 있다고 제안되었다.[133] 감수분열 재조합은 Spo11 단백질에 의해 촉매되는 이중 가닥 절단에 의해 매개되며, Mre11, Sae2, Exo1 단백질도 DNA 절단과 재조합에 관여한다. DNA 절단 후에는 상동인 부분에서 재조합이 일어나는데, 이는 이중 할리데이 이음(dHJ) 경로나 합성 의존적 가닥 어닐링(SDSA) 방법 중 하나로 이루어진다(SDSA 방법에서는 교차가 일어나지 않는다).[134]
감수분열에도 체크포인트가 존재하여, ''Schizosaccharomyces pombe''에서는 Rad 단백질, Mek1(FHA 카이네이스 도메인 포함), Cdc25, Cdc2 단백질, 그리고 아직 알려지지 않은 인자들이 체크포인트를 형성할 것으로 생각된다.[135]
척추동물의 난자발생에서는 세포정지인자(cytostatic factor) (CSF)가 감수분열 II로의 전환을 조절한다.[133]
참조
[1]
웹사이트
4.1: Meiosis
https://bio.libretex[...]
2021-05-29
[2]
웹사이트
Definition of Reduction division
https://www.medicine[...]
2021-05-29
[3]
서적
Biological Science
Pearson
2011
[4]
논문
To err (meiotically) is human: the genesis of human aneuploidy
2001-04-01
[5]
웹사이트
Interactive Tree of Life
http://itol.embl.de/
2011-07-23
[6]
논문
Evolutionary origin of recombination during meiosis
[7]
논문
Sex is not a solution for reproduction: the libertine bubble theory
2011-06-01
[8]
논문
On the Maiotic Phase (Reduction Divisions) in Animals and Plants
https://www.biodiver[...]
1904-02-01
[9]
논문
Meiosis and mitosis: a terminological criticism
https://books.google[...]
1985
[10]
웹사이트
Mitosis
http://biology.clc.u[...]
University of Cincinnati
2012-10-27
[11]
논문
Regulation of meiotic gene expression in plants
2014-08-01
[12]
논문
Unified single-cell analysis of testis gene regulation and pathology in five mouse strains
2019-06-01
[13]
논문
High-resolution view of the yeast meiotic program revealed by ribosome profiling
2012-02-01
[14]
서적
2005
[15]
논문
Genetic analysis of chromosome pairing, recombination, and cell cycle control during first meiotic prophase in mammals
2006-06-01
[16]
논문
Meiotic Recombination: The Essence of Heredity
2015-10-01
[17]
서적
2005
[18]
서적
Principles of Genetics
Wiley
2008-12-01
[19]
서적
Lewin's Genes X
https://archive.org/[...]
Jones & Barlett Learning
2009-11-01
[20]
논문
Recombination, Pairing, and Synapsis of Homologs during Meiosis
2015-05-01
[21]
논문
Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis
2007-07-01
[22]
논문
Meiosis, genetic recombination, and sexual reproduction
https://www.nature.c[...]
2008
[23]
논문
Meiotic chromosomes in motion: a perspective from Mus musculus and Caenorhabditis elegans
2019-09-01
[24]
논문
Gene conversion: mechanisms, evolution and human disease
2007-10-01
[25]
논문
Meiosis: the chromosomal foundation of reproduction
2018-07-01
[26]
서적
Thompson & Thompson genetics in medicine
Elsevier
2015-05-21
[27]
논문
Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes
2007-08-01
[28]
논문
Human oocytes. Error-prone chromosome-mediated spindle assembly favors chromosome segregation defects in human oocytes
2015-06-01
[29]
논문
Complete kinetochore tracking reveals error-prone homologous chromosome biorientation in mammalian oocytes
2011-08-01
[30]
서적
Chromosomes and Cell Reproduction
W.H. FREEMAN AND CO
2009
[31]
논문
Structure and functions of stable intercellular bridges formed by incomplete cytokinesis during development
2011-01-01
[32]
웹사이트
BioCoach Activity: Concept 11: Meiosis II: Metaphase II
http://www.phschool.[...]
Pearson
2018-02-10
[33]
논문
A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis
2005-01-01
[34]
논문
Population genetics provides evidence for recombination in Giardia
2007-11-01
[35]
논문
Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector
2009-04-01
[36]
논문
The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms
2011-07-01
[37]
논문
The first sexual lineage and the relevance of facultative sex
1999-06-01
[38]
논문
On the origins of the Mendelian laws
1984-01-01
[39]
논문
Inherited differences in crossing over and gene conversion frequencies between wild strains of Sordaria fimicola from "Evolution Canyon"
2001-12-01
[40]
논문
A molecular cell biology toolkit for the study of meiosis in the silkworm Bombyx mori
2023-05-01
[41]
논문
The transformation of the Synaptonemal Complex into the 'elimination chromatin' in Bombyx mori oocytes
1977-04-01
[42]
논문
Why is meiosis arrested?
1998-09-01
[43]
논문
Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health
2020-05-01
[44]
논문
Genetic damage, mutation, and the evolution of sex
1985-09-01
[45]
서적
Molecular Genetics of Development
1987
[46]
논문
The interacting pathways for prevention and repair of oxidative DNA damage
2003-10-01
[47]
논문
Sexual reproduction as a response to H2O2 damage in Schizosaccharomyces pombe
1989-04-01
[48]
논문
Heat shock elicits production of sexual inducer in Volvox
1986-01-01
[49]
논문
Sex as a response to oxidative stress: a twofold increase in cellular reactive oxygen species activates sex genes
2004-08-01
[50]
서적
An Introduction to Phycology
https://books.google[...]
Wiley
2009-07-08
[51]
서적
Stern's Introductory Plant Biology
McGraw Hill
[52]
서적
Chapter 7 - Regulation of Mammalian Oocyte Maturation
https://www.scienced[...]
Academic Press
2024-12-15
[53]
논문
Positioning to get out of meiosis: the asymmetry of division
[54]
논문
The contradictory information on the distribution of non-disjunction and pre-division in female gametes
2006-11-01
[55]
논문
Germ cell-intrinsic and -extrinsic factors govern meiotic initiation in mouse embryos
2008-12-01
[56]
논문
Nanos2 suppresses meiosis and promotes male germ cell differentiation
2008-02-01
[57]
논문
Retinoid signaling determines germ cell fate in mice
2006-04-01
[58]
논문
Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signalling
2011-01-01
[59]
논문
Two functionally redundant sources of retinoic acid secure spermatogonia differentiation in the seminiferous epithelium
2019-01-01
[60]
논문
Genome and Gene Structure
2019-01-01
[61]
논문
Meiosis
2013-01-01
[62]
논문
Effect of maternal age on autosomal trisomies
1980-07-01
[63]
논문
Age-related decrease of meiotic cohesins in human oocytes
2014-05-01
[64]
웹사이트
How Cells Divide
https://www.pbs.org/[...]
Public Broadcasting Service
2012-12-06
[65]
논문
Meiosis in protists. Some structural and physiological aspects of meiosis in algae, fungi, and protozoa
1976-03-01
[66]
논문
Meiosis in protists: recent advances and persisting problems
[67]
논문
Progesterone induced maturation in oocytes of Xenopus laevis. Appearance of a 'maturation promoting factor' in enucleated oocytes
https://www.scienced[...]
1973-10-01
[68]
논문
Cell cycle dynamics of an M-phase-specific cytoplasmic factor in Xenopus laevis oocytes and eggs
1984-04-01
[69]
논문
Molecular control of meiosis
https://www.cell.com[...]
1995-07-01
[70]
서적
A New Understanding on the Regulation of Oocyte Meiotic Prophase Arrest and Resumption
https://link.springe[...]
Springer International Publishing
2024-12-15
[71]
저널
Prophase I arrest and progression to metaphase I in mouse oocytes: comparison of resumption of meiosis and recovery from G2-arrest in somatic cells
https://academic.oup[...]
2010-09-01
[72]
저널
Oocyte Maturation: The Coming of Age of a Germ Cell
2005
[73]
저널
Differential Function and Expression of ''Saccharomyces cerevisiae'' B-type Cyclins in Mitosis and Meiosis
1993-04-01
[74]
저널
Molecular regulation of the mitosis/meiosis decision in multicellular organisms
2011-08
[75]
저널
Control of meiotic gene expression in Saccharomyces cerevisiae
1994
[76]
저널
Molecular mechanisms underlying the mitosis–meiosis decision
https://link.springe[...]
2007-07-01
[77]
저널
Regulatory mechanisms in meiosis
1993-04
[78]
저널
Mechanism and regulation of meiotic recombination initiation
2014-10
[79]
저널
Regulation of meiotic progression by the meiosis-specific checkpoint kinase Mek1 in fission yeast
2003-01
[80]
웹인용
4.1: Meiosis
https://bio.libretex[...]
2019-10-01
[81]
웹인용
Definition of Reduction division
https://www.medicine[...]
[82]
서적
Biological Science
https://archive.org/[...]
Pearson
2011
[83]
저널
To err (meiotically) is human: the genesis of human aneuploidy
2001-04
[84]
웹인용
Interactive Tree of Life
http://itol.embl.de/
[85]
저널
Evolutionary origin of recombination during meiosis
2010
[86]
저널
Sex is not a solution for reproduction: the libertine bubble theory
2011-06
[87]
간행물
s.v.
http://www.oed.com/v[...]
1905
[88]
서적
Meiosis and mitosis: a terminological criticism
https://books.google[...]
1985
[89]
웹인용
Mitosis
http://biology.clc.u[...]
University of Cincinnati
2012-10-27
[90]
저널
Regulation of meiotic gene expression in plants
2014-08
[91]
저널
Unified single-cell analysis of testis gene regulation and pathology in five mouse strains
2019-06
[92]
저널
High-resolution view of the yeast meiotic program revealed by ribosome profiling
2012-02
[93]
저널
Genetic analysis of chromosome pairing, recombination, and cell cycle control during first meiotic prophase in mammals
2006-06
[94]
저널
Meiotic Recombination: The Essence of Heredity
2015-10
[95]
참고문헌
2005
[96]
서적
Principles of Genetics
Wiley
2008-12
[97]
서적
Lewin's Genes X
https://archive.org/[...]
Jones & Barlett Learning
2009-11
[98]
저널
Recombination, Pairing, and Synapsis of Homologs during Meiosis
2015-05
[99]
저널
Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis
2007-07
[100]
저널
Meiosis, genetic recombination, and sexual reproduction
https://www.nature.c[...]
2008
[101]
저널
Meiotic chromosomes in motion: a perspective from Mus musculus and Caenorhabditis elegans
2019-09
[102]
저널
Gene conversion: mechanisms, evolution and human disease
2007-10
[103]
저널
Meiosis: the chromosomal foundation of reproduction
2018-07
[104]
저널
Recombination in the human Pseudoautosomal region PAR1
2014-07
[105]
서적
Thompson & Thompson genetics in medicine
2015-05-21
[106]
저널
Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes
2007-08-00
[107]
저널
Human oocytes. Error-prone chromosome-mediated spindle assembly favors chromosome segregation defects in human oocytes
2015-06-00
[108]
저널
Complete kinetochore tracking reveals error-prone homologous chromosome biorientation in mammalian oocytes
2011-08-00
[109]
서적
Chromosomes and Cell Reproduction
W.H. FREEMAN AND CO
2009-00-00
[110]
저널
Structure and functions of stable intercellular bridges formed by incomplete cytokinesis during development
2011-01-00
[111]
웹인용
BioCoach Activity: Concept 11: Meiosis II: Metaphase II
http://www.phschool.[...]
Pearson
2018-02-10
[112]
저널
On the origins of the Mendelian laws
1984-01-01
[113]
저널
Inherited differences in crossing over and gene conversion frequencies between wild strains of Sordaria fimicola from "Evolution Canyon"
2001-12-00
[114]
저널
Why is meiosis arrested?
1998-09-00
[115]
저널
Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health
2020-05-00
[116]
서적
An Introduction to Phycology
https://books.google[...]
John Wiley & Sons
2009-07-08
[117]
서적
Introductory Plant Biology
McGraw HIll
[118]
저널
Positioning to get out of meiosis: the asymmetry of division
https://semanticscho[...]
[119]
저널
The contradictory information on the distribution of non-disjunction and pre-division in female gametes
2006-11-00
[120]
저널
Germ cell-intrinsic and -extrinsic factors govern meiotic initiation in mouse embryos
2008-12-00
[121]
저널
Nanos2 suppresses meiosis and promotes male germ cell differentiation
2008-02-00
[122]
저널
Retinoid signaling determines germ cell fate in mice
2006-04-00
[123]
저널
Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signalling
2011-01-00
[124]
저널
Two functionally redundant sources of retinoic acid secure spermatogonia differentiation in the seminiferous epithelium
2019-01-00
[125]
저널
Genome and Gene Structure
2019-01-01
[126]
저널
Meiosis
2013-01-01
[127]
저널
Effect of maternal age on autosomal trisomies
1980-07-00
[128]
저널
Age-related decrease of meiotic cohesins in human oocytes
2014-05-00
[129]
웹인용
How Cells Divide
https://www.pbs.org/[...]
Public Broadcasting Service
2012-12-06
[130]
저널
Meiosis in protists. Some structural and physiological aspects of meiosis in algae, fungi, and protozoa
1976-03-00
[131]
저널
Meiosis in protists: recent advances and persisting problems
[132]
저널
Molecular regulation of the mitosis/meiosis decision in multicellular organisms
2011-08-00
[133]
저널
Regulatory mechanisms in meiosis
1993-04-00
[134]
저널
Mechanism and regulation of meiotic recombination initiation
2014-10-00
[135]
저널
Regulation of meiotic progression by the meiosis-specific checkpoint kinase Mek1 in fission yeast
2003-01-00
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com