원생생물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
원생생물은 동물, 식물, 균류가 아닌 모든 진핵생물을 포괄하는 측계통 분류군이다. 이들은 주로 단세포 생물이지만, 다세포 구조를 형성하기도 하며, 다양한 생활사, 영양 방식, 운동성, 세포 구조를 보인다. 생물 분류 체계의 변천에 따라 원생생물에 대한 정의와 분류는 변화해 왔으며, 현재는 계통발생학적, 기능적 정의를 따른다. 원생생물은 엑스카바타, 디아포레티케스, 아모르페아 등 다양한 그룹으로 나뉘며, 형태학적 특성에 기반한 분류는 종 다양성을 과소평가한다는 한계가 있다. 원생생물은 생태계에서 1차 생산자, 소비자, 분해자, 기생충 등 다양한 역할을 하며, 생지화학적 순환에도 기여한다. 화석 기록을 통해 원생생물의 진화 과정을 추적할 수 있으며, 중원생대와 신원생대에 걸쳐 다양한 원생생물 화석이 발견되었다.
더 읽어볼만한 페이지
- 생물에 관한 - 살아있는 화석
살아있는 화석은 화석 기록에서 오랜 기간 동안 형태적 변화가 거의 없이 나타나는 생물을 의미하며, 진화와 종분화를 이해하는 데 중요한 역할을 한다. - 생물에 관한 - 적응
적응은 생물이 환경에 더 잘 생존하고 번식하도록 돕는 진화 과정으로, 형태, 행동, 생리적 특징 등 다양한 측면에서 유전적 변화, 공진화, 의태 등의 방식으로 나타난다. - 원생생물 - 유공충
유공충은 단단한 껍질을 가진 원생생물로, 껍질 형태와 구성에 따라 분류되며, 해양 환경에서 생물 층서 및 고기후 연구, 석유 탐사 등 다양한 분야에 활용된다. - 원생생물 - 코아노조아
코아노조아는 칼라 구조를 특징으로 하는 깃편모충류와 동물을 포함하는 계통 분류군으로, 동물의 기원과 밀접한 관련이 있으며, 동물 진화 연구에 중요한 연결고리를 제공한다. - 측계통군 - 맹금류
맹금류는 날카로운 발톱과 부리를 가진 사나운 새를 통칭하며, 생태계 최상위 포식자로서 뛰어난 시력과 비행 능력을 갖추고 있으며, 수리, 매, 독수리, 올빼미 등이 대표적이고, 서식지 파괴 등의 위협에 직면해 있다. - 측계통군 - 돌고래
돌고래는 이빨고래류에 속하는 해양 포유류의 총칭으로, 다양한 종이 존재하며 높은 지능과 사회성을 가지지만, 서식지 파괴, 환경 오염 등으로 멸종 위기에 처해 있다.
| 원생생물 - [생물]에 관한 문서 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
| 이름 | 원생생물 |
| 로마자 표기 | Protist |
| 발음 | /ˈproʊtɪst/ |
| 유사 발음 | PROH-tist |
| 분류 상태 | 측계통군 |
| 화석 범위 | 고원생대 2100만 년 전 – 현재 |
| 상위 분류군 | 진핵생물 |
| 하위 분류군 | 슈퍼그룹 |
| 하위 분류 | 아메바류 크루미류 안키로모나드류 디스코바 말라위모나드류 메타모나다 헤미마스티고포라 프로보라 디아포레티케스 |
| 제외 대상 | 동물 균류 배아식물 (육상 식물) |
| 분류 체계 (영문) | |
| 분류 상태 (영문) | paraphyletic |
| 이름 (영문) | Protists |
| 상위 분류군 (영문) | Eukaryota |
| 하위 분류군 (영문) | Supergroups |
| 하위 분류 (영문) | Amorphea CRuMs Ancyromonadida Discoba Malawimonadida Metamonada Hemimastigophora Provora Diaphoretickes |
| 제외 대상 (영문) | Animalia Fungi Embryophyta (land plants) |
| 분류 체계 | |
| 계 | 원생생물계 |
| 학명 | Protista |
| 명명자 | Haeckel, 1866 |
| 하위 분류 이름 | 하위 분류 |
| 하위 분류 | 아메바류 아푸소모나디다 원시색소체생물 (육상식물 포함) 브레비아테아 크루미류 크립티스타 디스코바 합티스타 헤미마스티고포라 말라위모나디다 메타모나스 오피스토콘타 프로보라 SAR 슈퍼그룹 테로네미아 |
| 분류 체계 (원생생물) | |
| 계 (원생생물) | 원생생물계 |
| 분류 (원생생물) | 측계통군 |
| 하위 분류군 (원생생물) | 문 |
| 하위 분류 (원생생물) | 크로말베올라타계 부등편모조류 착편모조류 은편모조류 피하낭류 와편모충류 정단복합체충류 섬모충류 엑스카바타계 유글레나식물 페르콜로조아류 메타모나다 리자리아계 방산충 유공충 사족충류 원시색소체생물 - 일부 홍조식물 - 홍조류 회청조식물 - 기초 원시색소체생물 단편모생물 - 일부 아메바류 코아노조아 기타 다수 및 다양한 분류 |
| 이미지 | |
 | |
| |
2. 생물 분류 체계의 변천
생물을 분류하는 체계는 역사적으로 계속 변화해왔다. 고전적인 린네의 2계 분류 이후 미생물의 발견 등으로 새로운 분류 기준이 필요하게 되었다.
19세기 에른스트 헤켈은 동물과 식물 외에 제3의 계인 원생생물계를 추가하여 3계 분류 체계를 제시했다. 이후 에두아르 샤통은 세포 구조에 따라 생물을 원핵생물과 진핵생물로 나누는 중요한 개념을 도입했다. 허버트 코플랜드는 원핵생물을 모네라계로 독립시켜 4계 분류 체계를 만들었고, 로버트 휘태커는 균류를 독립된 균계로 설정하여 5계 분류 체계를 제안했다. 이 5계 분류 체계는 오랫동안 널리 받아들여졌다.[127]
분자생물학의 발달은 생물 분류에 큰 변화를 가져왔다. 1990년 칼 워즈는 리보솜 RNA(rRNA) 염기서열 분석을 바탕으로 생물을 세균역, 고세균역, 진핵생물역의 3개의 역(Domain)으로 나누는 3역 분류 체계를 제안했다.[126] 이는 기존 모네라계에 속했던 생물들이 실제로는 세균과 고세균이라는 매우 다른 두 그룹으로 나뉜다는 점을 보여주었다.
한편, 토머스 캐빌리어스미스는 세포 구조와 유전 정보를 종합하여 원생생물계를 더욱 세분화하려는 시도로 크로미스타계, 원생동물계 등을 포함하는 6계 또는 8계 분류 체계를 제안하기도 했다.
아래 표는 주요 학자들의 생물 분류 체계 변천 과정을 요약하여 보여준다.
| 린네 (1735) | 헤켈 (1866) | 샤통 (1925) | 코플랜드 (1938) | 휘태커 (1969) | 워즈 외 (1977, 1990) | 캐빌리어스미스 (1993, 1998) | ||
|---|---|---|---|---|---|---|---|---|
| 2계 | 3계 | 2계 | 4계 | 5계 | 6계 | 3역 | 8계 | 6계 |
| (다루지 않음) | 원생생물 (Protista) | 원핵생물 (Prokaryota) | 모네라 (Monera) | 모네라 (Monera) | 진정세균 (Eubacteria) | 세균 (Bacteria) | 진정세균 (Eubacteria) | 세균 (Bacteria) |
| 고세균 (Archaeabacteria) | 고세균 (Archaea) | 고세균 (Archaeabacteria) | ||||||
| 진핵생물 (Eukaryota) | 원생생물 (Protista) | 원생생물 (Protista) | 원생생물 (Protista) | 진핵생물 (Eukarya) | 고동물 (Archezoa) | 원생동물 (Protozoa) | ||
| 원생동물 (Protozoa) | ||||||||
| 유색조식물 (Chromista) | 유색조식물 (Chromista) | |||||||
| 식물 (Vegetabilia) | 식물 (Plantae) | 식물 (Plantae) | 식물 (Plantae) | 식물 (Plantae) | 식물 (Plantae) | 식물 (Plantae) | ||
| 균류 (Fungi) | 균류 (Fungi) | 균류 (Fungi) | 균류 (Fungi) | |||||
| 동물 (Animalia) | 동물 (Animalia) | 동물 (Animalia) | 동물 (Animalia) | 동물 (Animalia) | 동물 (Animalia) | 동물 (Animalia) |
대한민국에서는 1990년대 이후 교육과정 등에서 휘태커의 5계 분류 체계가 주로 사용되었으나, 최근에는 워즈의 3역 분류 체계가 학계의 주류로 받아들여지면서 그 중요성이 강조되고 있다.[126] 생물 분류 체계는 새로운 과학적 발견에 따라 계속해서 수정되고 발전하는 역동적인 분야이다.
2. 1. 린네부터 헤켈까지
고전적으로 생물은 크게 동물계(움직이며 먹이를 구하는 생물)와 식물계(동물이 아닌 생물)의 두 그룹으로 분류되었다. 이러한 2계설에서는 균류 역시 식물계에 포함되었으며, 오랫동안 큰 문제 없이 받아들여졌다.하지만 안톤 판 레이우엔훅이 미생물을 발견하면서 상황은 크게 달라졌다. 이후 많은 연구자들이 다양한 미생물을 찾아냈는데, 이들 중에는 아오미도리처럼 움직이지 않고 광합성을 하여 식물로 분류할 수 있는 것, 짚신벌레처럼 활발히 움직이며 먹이를 먹어 동물로 볼 수 있는 것, 곰팡이처럼 포자를 만들어 균류로 간주할 수 있는 것 등 기존 분류 틀에 맞는 생물도 있었다.
그러나 기존의 동물과 식물이라는 이분법적 분류로는 설명하기 어려운 생물들이 다수 발견되었다. 예를 들어, 유글레나는 광합성을 하면서도 편모로 운동하는 특징을 가졌고, 변형균은 포자를 만드는 균류와 비슷하지만 영양체 상태에서는 아메바처럼 움직이며 먹이를 섭취하는 동물과 유사한 모습을 보였다. 또한, 규조류는 단단한 껍질을 가지고 광합성을 하면서도 이동 능력을 갖추고 있었다.
19세기에 이르러 에른스트 헤켈은 이러한 동물과 식물의 경계에 있는 원시적인 생물들을 묶어 제3의 생물계인 '''원생생물계'''(Protista)를 제안하며 3계설을 제시했다. 헤켈이 처음 원생생물계로 분류한 대상에는 세균, 진균, 단세포 조류, 원생동물, 심지어 해면동물까지 포함되었으나, 이후 연구를 통해 주로 단세포 생물을 중심으로 하는 개념으로 정리되었다.
2. 2. 휘태커의 5계 분류 체계
1959년, 로버트 휘태커는 생물을 모네라계, 원생생물계, 식물계, 균계, 동물계의 5개 계로 나누는 5계설을 제안했다.[127] 이는 생물의 진화 방향을 크게 세 가지로 본 그의 관점에서 출발했다. 첫째는 광합성을 하며 고정 생활을 하는 식물의 방향, 둘째는 운동하며 먹이를 섭취하는 동물의 방향, 셋째는 몸 표면에서 유기물을 흡수하는 균류의 방향이다. 휘태커는 먼저 세포 구조가 다른 원핵생물을 모네라계로 분리하고, 나머지 진핵생물 중에서 앞서 언급한 세 가지 방향으로 잘 발달한 무리를 각각 식물계, 동물계, 균계로 설정했다. 그리고 이 세 가지 방향으로 분화했지만 그 정도가 낮아 명확히 구분하기 어려운 단세포 진핵생물들을 묶어 '''원생생물계'''로 분류했다. 즉, 원생생물계는 특정 계통으로 묶인 것이 아니라 생물의 발달 단계에 따라 분류된 것이었다.처음 휘태커가 제안한 원생생물계는 단세포 생물만을 포함했지만, 이후 휘태커 자신과 린 마굴리스 등에 의해 진핵 다세포 생물까지 포함하도록 재정의되었다. 이 확장된 정의에 따르면, 원생생물은 “배를 만들지 않고, 조직의 분화나 형성이 없으며, 편모를 가질 경우 미세소관이 9+2 구조로 배열되는 진핵생물”을 의미한다. 기존 원생생물계의 학명은 Protista|프로티스타la였으나, 진핵 다세포 생물을 포함하는 확장된 분류에서는 Protoctista|프로토크티스타la라는 학명을 사용하기도 한다.
2. 3. 캐빌리어스미스의 분류 체계
최근 편모 기저 구조 등 세포 내 미세 구조 연구와 유전자 분석을 통한 생물 간 계통 분석 방법이 발전하면서 원생생물 내에서도 계통 관계 연구가 활발히 진행되고 있다. 또한, 기존 세포내 공생설에서 예상하지 못했던 진핵세포 간의 세포내 공생 발견도 분류 체계에 영향을 미쳤다.이러한 연구 결과를 바탕으로 토머스 캐빌리어스미스는 1993년에 원생생물계를 더욱 세분화한 팔계설을 제안했다. 그는 갈조류를 포함한 황색 식물 등 일부 조류군이 진핵세포에서 유래한 엽록체를 가지며, 엽록체가 없는 난균류, 라비린추라류와 함께 특정 편모 구조(앞쪽 깃털 모양, 뒤쪽 채찍 모양)를 공통으로 가진다는 점에 주목했다. 여기에 형태적으로 유사한 하프토조류와 크립토조류를 포함하여 이들을 하나의 계통군으로 묶어 크로미스타계(Chromista)로 독립시켰다.
또한, 미포자충과 같이 미토콘드리아가 없는 일부 단세포 생물들을 미토콘드리아가 공생하기 이전 단계의 원시적인 진핵생물로 보고 아르케조아계(Archezoa)로 분류했다. 기존 원생생물계에서 크로미스타계와 아르케조아계를 제외한 나머지 생물들은 원생동물계(Protozoa)로 분류했다.
그러나 아르케조아계는 학계에서 널리 받아들여지지 않았다. 이들 생물이 미토콘드리아를 가지지 않는 것은 원시적인 특징이 아니라 이차적으로 퇴화한 결과라는 비판이 제기되었고, 캐빌리어스미스 자신도 나중에 이 분류를 철회했다. 이후 그는 1998년에 아르케조아계를 제외한 6계 분류 체계를 다시 제안하기도 했다.
현재도 원생생물을 포함한 생물 분류 체계에 대한 연구와 재검토는 계속 이루어지고 있다.[126]
3. 원생생물의 정의와 특징
원생생물은 주로 단세포이며 현미경으로 관찰해야 할 만큼 작지만, 다양한 모양과 생존 전략을 보이는 진핵생물(세포 안에 핵이 있는 생물)의 다양한 그룹이다. 원생생물은 다양한 생활사, 영양 단계, 운동 방식, 세포 구조를 가진다.[1][2] 대부분 단세포생물이지만, 일부는 군체를 이루거나 다세포 구조를 형성하여 육안으로 볼 수 있는 크기를 가지기도 한다. '원생생물'이라는 용어는 동물, 식물, 균류가 아닌 모든 진핵생물을 아우르는 측계통 분류군으로 정의된다.[117] 이러한 정의 방식 때문에 원생생물은 진핵생물에서 나타나는 거의 모든 생물학적 특성을 포함한다.
역사적으로 원생생물과 다른 세 진핵생물 계(동물, 식물, 균류) 사이의 구분은 명확하지 않았다. 종속영양생물인 원생동물은 동물계의 일부로 간주되었고, 광합성 생물인 조류는 식물계의 일부로 연구되었다. 별도의 원생생물계가 만들어진 후에도 일부 미세 동물(믹소조아류)[14], '하등' 균류(아페리드, 로젤리드, 미크로스포리디아류 등)[3][4], 일부 조류(홍조류, 녹조류)는 각각 원생생물 또는 식물로 분류되기도 했다. 현재는 '원생생물'이 동물, 유배자식물(육상 식물) — 즉, 모든 조류는 이 범주에 속한다 — 및 모든 균류를 특히 배제하는 것으로 합의되었지만, 하등 균류는 여전히 원생생물학자와 균류학자 모두 연구 대상으로 삼는 경우가 많다.
일부 원생생물(양계통 원생생물)은 동물과 식물 또는 균류의 특징을 모두 가져(점균류, 유글레나류 등), 식물학적 명명 규약(ICN)과 동물학적 명명 규약(ICZN) 모두에 따라 이름이 붙여지기도 했다.
원생생물은 다양한 형태를 보이며, 이는 실용적인 분류에 사용되어 왔지만, 대부분 진화적으로 일관된 분지군을 나타내지는 않는다. 주요 형태는 다음과 같다.
- 아메바: 불규칙하고 유연한 형태로, 세포질의 일부인 위족을 뻗어 표면을 따라 기어 다니며 이동한다. 많은 아메바는 맨몸이지만, 피낭성 아메바와 유공충은 소화된 물질이나 주변의 잔해로 세포 주변에 껍질을 만든다. 방산충과 태양충으로 알려진 일부 아메바는 미세소관으로 지지되는 위족이 세포에서 방사형으로 뻗어 나온 특별한 구형 모양을 가지고 있다. 어떤 아메바는 포자를 가진 줄기가 있는 다세포 단계를 생성할 수 있으며, 종종 서로 응집하여 점균류로 알려져 있다. 아메바를 포함하는 주요 분지군은 다양한 점균류와 피낭성 아메바를 포함하는 아메바류와 리자리아(유명한 그룹인 유공충과 방산충뿐만 아니라 일부 피낭성 아메바 포함)이다. 심지어 일부 아메바는 맨눈으로 볼 수 있을 정도로 거대한 크기로 자랄 수 있다.[5][6]
- 편모충: 이 원생생물은 섬모, 운동편모 또는 진핵생물 편모라고 하는 채찍 모양의 부속물 하나 이상을 가지고 있으며, 이를 통해 환경에서 자유롭게 수영하거나 활주할 수 있다. 편모충은 모든 계통에서 발견되며, 모든 살아있는 진핵생물의 공통 조상이 편모충이었음을 반영한다. 일반적으로 두 개의 섬모를 가지고 있지만(프로보라, 텔로네미아, 스트라메노필라, 알베올라타, 오바조아 및 대부분의 굴착류), 많은 수의 섬모를 가진 편모충 그룹도 있다(헤미마스티고포라 및 기타 굴착류). 잘 알려진 섬모충과 기생성 오팔리니드와 같은 일부 그룹은 섬모가 일정하게 움직이는 세포 표면을 가지고 있다. 일부 아메바 그룹은 편모를 유지하여 아메바 편모충이 된다.[10]
- 조류: 이들은 광합성 원생생물이며, 대부분의 주요 분지군에서 전통적으로 ''원생동물''이라고 불리는 종속영양 원생생물과 완전히 섞여서 발견된다. 조류는 단일 편모 또는 구형 세포(크립토모나드, 하프토파이트, 디노플라젤라, 크로메리드, 많은 녹조류, 황갈조류, 유글레나류)에서부터 아메바형 세포(클로라라크니오파이트)에 이르기까지, 그리고 군체 및 다세포 거대 형태(홍조류, 일부 녹조류, 그리고 켈프와 같은 일부 황갈조류)에 이르기까지 가장 다양한 형태의 범위를 보여준다.[11]
- 균류와 같은 원생생물: 여러 분지군의 원생생물은 균류와 유사한 외관을 균사와 같은 구조와 부생영양을 통해 진화시켰다. 이들은 여러 번 진화했으며, 종종 진정한 균류와 매우 거리가 멀다(난균류, 미로균류, 히포키트리드류는 스트라메노필라에; 점균류는 아메바류에; 식물미생물은 리자리아에; 퍼킨소조아는 알베올라타에).[12][13]
- 포자충: 이 범주에는 전통적으로 포자를 통해 번식하는 기생 원생생물(아피콤플렉사류, 미세포자충류, 믹소조아류, 아세토스포리아류)가 포함되었다.[14] 현재는 아피콤플렉사류[15]와 같이 말라리아의 원인인 ''말라리아 원충''에 제한된다.[16]
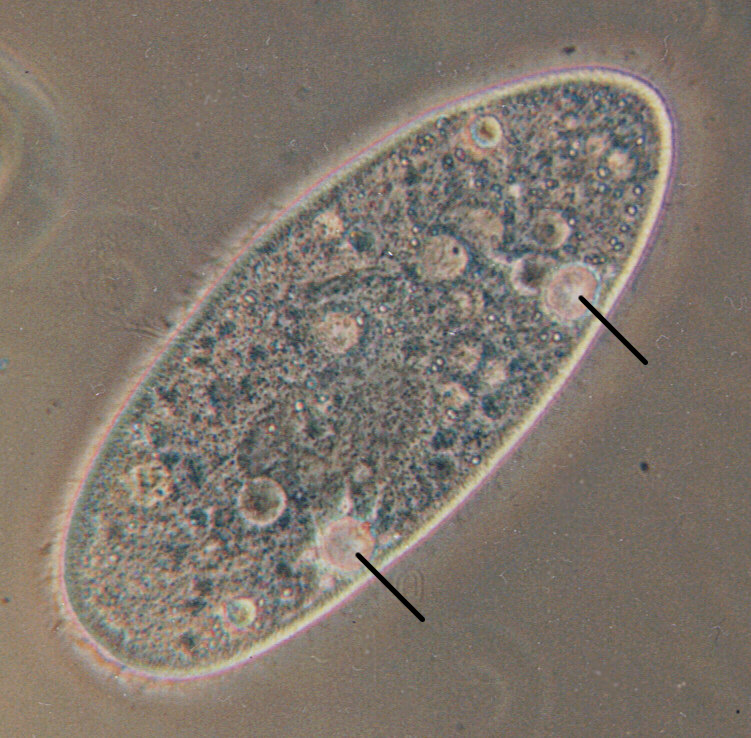
일반적으로 원생생물은 "고등" 진핵생물(동물, 균류 또는 식물)의 세포에서 설명되는 동일한 생리학 및 생화학 원리를 따르는 전형적인 진핵세포이다. 이들은 미토콘드리아를 통해 에너지를 생산하기 위해 산소를 소비하는 호기성 생물이며, 엽록체를 가진 원생생물은 엽록체에서 광합성을 통해 탄소를 고정한다.[17] 그러나 많은 원생생물이 나머지 진핵생물에는 나타나지 않는 다양한 독특한 생리적 적응을 진화시켰다.
- '''삼투조절''': 세포벽이 없는 담수 원생생물은 수축포를 통해 삼투압을 조절할 수 있다. 수축포는 확장기와 수축기를 주기적으로 반복하며 칼륨과 나트륨이 풍부한 액체를 배출하는 특수한 세포소기관이다. 세포가 적응할 때까지 세포가 다른 염도의 배지에 놓이면 이러한 주기는 멈춘다.
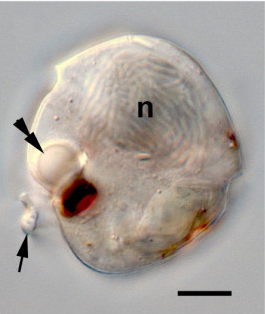
- '''에너지 적응''': 최근 공통 조상 진핵생물은 미토콘드리아를 가지고 산화 대사를 수행하는 호기성 생물이었다. 많은 자유 생활 및 기생성 원생생물 계통은 독립적으로 진화하여 혐기성 또는 미호기성 서식지를 서식하도록 적응하였다. 초기 미토콘드리아를 수소체로 변형시켜 피루브산의 발효를 통해 혐기적으로 ATP를 생성하는 세포소기관이다. 마찬가지로 미호기성 트리파노소마 원생생물에서 발효성 글리코좀은 퍼옥시좀에서 진화하였다.
- '''감각 적응''': 많은 편모충류와 아마도 모든 운동성 조류는 긍정적인 광주성(즉, 빛의 근원을 향해 헤엄치거나 미끄러져 간다)을 보인다. 이를 위해 세 가지 종류의 광수용체 또는 "안점"을 나타낸다. (1) 많은 녹조류, 디노플라젤라 및 크립토파이타에서 발견되는 빛 안테나가 있는 수용체, (2) 불투명한 스크린이 있는 수용체, (3) 포식성 디노플라젤라의 한 그룹인 Warnowiaceae에서 발견되는 세포 내 렌즈가 있는 복잡한 오셀로이드이다. 또한, 일부 섬모충류는 이동하면서 지구의 중력장과 관련하여 스스로의 방향을 정하고( 지구성), 다른 것들은 물 속에 녹아 있는 산소 농도와 관련하여 헤엄친다.
- '''내공생''': 원생생물은 세포에 내공생체를 포함하는 경향이 두드러지며, 이는 새로운 생리적 기회를 만들어냈다. ''Paramecium bursaria''와 그 내공생체 ''클로렐라''와 같이 일부 연관성은 더 영구적이고, 다른 연관성은 더 일시적이다. 많은 원생생물은 포획된 엽록체, 엽록체-미토콘드리아 복합체, 심지어 조류의 안점을 포함하고 있다. 제노좀은 섬모충류에서 발견되는 세균 내공생체이며, 때로는 혐기성 섬모충류 내부에서 메탄 생성 역할을 한다.
원생생물은 일반적으로 환경 조건이 좋으면 무성생식을 하지만, 기아나 열 충격과 같은 스트레스 조건에서는 유성생식하는 경향이 있다. 산화 스트레스는 DNA 손상을 초래하며, 원생생물의 유성생식 유도에 중요한 요인으로 보인다.[19]
진핵생물은 15억 년 전 이상 진화 과정에서 출현했다.[20] 가장 초기 진핵생물은 원생생물이었다. 유성생식은 다세포 진핵생물에서 널리 퍼져 있지만, 최근까지 유성생식이 진핵생물의 원시적이고 기본적인 특징일 수 있다는 것은 의외였다. 이러한 견해의 주된 이유는 진핵생물 계통수에서 일찍 갈라져 나온 특정 병원체 원생생물에서는 유성생식이 없는 것처럼 보였기 때문이다. 그러나 감수분열과 유성생식의 출현 이전의 것으로 여겨졌던 이러한 "초기 분기" 원생생물(예: ''람블편모충'' 및 ''질편모충'') 중 상당수는 현재 유성 진핵생물에 존재하는 일련의 감수분열 유전자를 가지고 있기 때문에 감수분열과 감수분열 재조합이 가능한 조상으로부터 유래한 것으로 알려져 있다.[21][22] 이러한 감수분열 유전자 대부분은 모든 진핵생물의 공통 조상에 존재했을 가능성이 높으며,[23] 이 조상은 선택적(비의무적) 유성생식이 가능했을 가능성이 크다.[24]
이러한 견해는 2011년 아메바에 대한 연구에 의해 더욱 뒷받침되었다. 아메바는 무성 생물로 간주되어 왔지만, 이 연구는 대부분의 아메바형 계통이 조상적으로 유성생식을 하고 있으며, 대부분의 무성생식 그룹은 최근에 독립적으로 발생했을 가능성이 높다는 증거를 제시하고 있다.[25] 20세기 초에도 일부 연구자들은 아메바에서 크로미디아(세포질 내에 자유롭게 존재하는 염색질 과립)와 관련된 현상을 유성생식으로 해석했다.[26]
일반적으로 발견되는 원생생물 병원체, 예를 들어 ''톡소플라스마 곤디''는 다양한 동물(이차 또는 중간 숙주)에 감염되어 무성 생식을 할 수 있지만, 일차 또는 최종 숙주(예: 이 경우 고양잇과 동물, 예를 들어 집고양이)에서만 유성 생식을 할 수 있다.[27][28][29]
일부 종, 예를 들어 ''말라리아 원충'',은 매우 복잡한 생활사를 가지고 있으며, 그중 일부는 유성 생식을 하고 다른 일부는 무성 생식을 한다.[30] 그러나 자연에서 유성 생식이 서로 다른 ''말라리아 원충'' 균주 사이의 유전자 교환을 얼마나 자주 일으키는지는 불분명하며, 기생성 원생생물의 대부분의 개체군은 다른 종과 유전자를 거의 교환하지 않는 클론 계통일 수 있다.[31]
''라이슈마니아'' 속의 병원성 기생 원생생물은 무척추동물 매개체에서 유성 생식 주기를 수행할 수 있는 것으로 나타났으며, 이는 트리파노소마에서 일어나는 감수분열과 유사하다.[32]
단세포 생물 외에도, 다세포이지만 매우 작아서 미생물로 취급되는 것이 많지만, 갈조류(갈조식물문: 다시마 등), 홍조류(홍조식물문: 우뭇가사리, 김 등)처럼 대형이 되는 것도 있다. 또한, 세포성 점균처럼 단세포로 독립하여 영양을 섭취하는 시기와 다세포의 자실체를 형성하는 시기 모두를 생활사에 가지는 생물도 포함되어 있다.
원생생물계에는 다음과 같은 생물이 포함된다.
원생생물은 수중이나 수분이 많은 토양에 서식하고 있다. 육상에서도 햇볕이 잘 드는 곳이나 바위 위 등 건조가 심한 곳에서도, 지의류처럼 다른 생물과 공생하거나, 건조할 때는 휴면하여 물이 있을 때만 활동하는 등의 방법으로 생활하는 것이 있다. 다른 생물에 기생하여 생활하는 종도 있다. 동물의 장 등에도 특수한 것이 서식하고 있지만, 기생의 경우, 공생 관계가 있는 경우, 불명확한 경우 등 다양하다. 장내나 사니층의 내부는 유기물이 풍부하고 산소가 매우 적다. 이것을 식물 출현 이전의 지구 환경에 가깝다고 보고, 그러한 조건에서 서식했던 생물의 현재 서식지라고 보는 견해도 있다.
4. 원생생물의 다양성

전통적으로 형태학적 특징에 기반하여 종을 구분하는 방법은 원생생물의 실제 종 다양성을 심각하게 과소평가하는 경향이 있다. 현재까지 기록된 원생생물 종의 수는 약 26,000종에서 76,000종 이상[42]으로 추정되는데, 이는 역사적으로 잘 연구된 식물, 동물, 균류의 다양성에 비해 매우 적은 숫자이다. 예측되는 총 종 수는 14만 종에서 160만 종까지 연구마다 크게 다르지만, 이러한 예측은 종종 주관적인 추정에 기반한다.
그러나 최근 환경 DNA 바코딩과 같은 분자 기술의 발달로 이전에는 알려지지 않았던 수많은 원생생물 계통이 발견되면서, 원생생물의 광대한 다양성이 드러나고 있다. 이러한 연구 결과들은 현재까지 발견된 진핵생물 유전자 서열이나 조작적 분류 단위(OTU, Operational Taxonomic Units)의 대부분이 원생생물에서 유래하며, 그 수가 식물, 동물, 균류를 합친 것보다 훨씬 많다는 것을 보여준다. 이 때문에 오늘날 원생생물은 진핵생물 다양성의 대부분을 차지하는 것으로 여겨진다.
원생생물의 진화적 관계와 계통은 분자 계통 유전학, 전체 게놈 및 전사체 시퀀싱, 그리고 편모 장치나 세포골격과 같은 미세 구조에 대한 전자 현미경 연구를 통해 점차 밝혀지고 있다. 새로운 주요 원생생물 계통과 생물 다양성이 계속 발견됨에 따라, 생명의 진핵생물 계통수는 지속적으로 수정되고 있다. 가장 최근의 진핵생물 분류 체계는 계, 문, 강, 목 등 전통적인 분류 계급을 공식적으로 사용하지 않고, 대신 유연관계에 기반한 분지군(clade)만을 인정하는 경향이 있다. 이는 장기적으로 분류 체계를 더 안정적으로 유지하고 업데이트하기 쉽게 만들기 위함이다. 이 새로운 분지학적 체계에서 원생생물은 비공식적인 '상위 그룹'(supergroup)으로 불리는 여러 큰 가지로 나뉜다. 예를 들어, 대부분의 광합성 진핵생물은 식물을 포함하는 아르케플라스티다와 TSAR 상위그룹 등을 포함하는 디아포레티케스(Diaphoretickes) 분지군에 속하며, 동물과 균류는 아메바류 등과 함께 아모르페아(Amorphea) 상위 그룹에 속한다. 원시적인 세포 구조를 가진 다양한 그룹은 집합적으로 엑스카바타(Excavata)로 분류되기도 한다.
4. 1. 주요 원생생물 그룹 (분지군)
원생생물은 매우 다양한 형태를 나타내며, 과거에는 이러한 형태적 특징을 기준으로 실용적인 분류가 이루어졌다. 그러나 이러한 분류 대부분은 진화적으로 일관된 분지군을 나타내지 않으며, 여러 형태는 서로 다른 계통에서 독립적으로 진화한 경우가 많다. 주요 형태는 다음과 같다.- 아메바: 불규칙하고 유연한 몸 형태를 가지며, 세포질의 일부인 위족을 뻗어 표면을 기어 다니며 이동하는 것이 특징이다. 대부분은 세포벽 없이 맨몸 상태이지만, 피낭성 아메바나 유공충처럼 소화된 물질이나 주변 환경의 잔해를 이용하여 세포 주변에 껍질을 만드는 종류도 있다. 방산충이나 태양충과 같은 일부 아메바는 미세소관으로 지지되는 가늘고 긴 위족이 세포체에서 방사형으로 뻗어 나오는 독특한 구형 대칭 형태를 보인다. 일부 아메바는 여러 세포가 모여 포자를 형성하는 줄기 구조를 만드는 다세포 단계를 거치기도 하는데, 이들은 흔히 점균류로 알려져 있다. 아메바 형태는 아메바류(점균류, 피낭성 아메바 등 포함)와 리자리아(유공충, 방산충 등 포함) 등 여러 분지군에서 나타난다. 일부 아메바는 맨눈으로도 관찰 가능할 정도로 거대한 크기로 성장하기도 한다.[5][6]
- 편모충: 채찍 모양의 부속기관인 섬모 또는 편모를 하나 이상 가지고 있어, 이를 이용해 물속을 자유롭게 헤엄치거나 표면을 따라 활주하며 이동한다. 편모충은 거의 모든 주요 진핵생물 분지군에서 발견되며, 이는 모든 현존 진핵생물의 공통 조상이 편모충이었을 가능성을 시사한다. 일반적으로 두 개의 편모를 가지는 경우가 많지만(프로보라, 텔로네미아, 스트라메노필라, 알베올라타, 오바조아 및 대부분의 굴착류), 많은 수의 편모를 가진 그룹도 존재한다(헤미마스티고포라 및 기타 굴착류). 섬모충이나 기생성 오팔리니드처럼 세포 표면 전체가 섬모로 덮여 조화롭게 움직이는 그룹도 있다. 일부 아메바 그룹은 편모를 유지하여 아메바 편모충 단계를 거치기도 한다.[10]
- 조류: 광합성을 하는 원생생물을 통칭하는 용어이다. 조류는 특정 분지군에 국한되지 않으며, 대부분의 주요 원생생물 분지군 내에서 종속영양 원생생물(전통적인 '원생동물')과 함께 발견된다. 조류는 단세포 편모충 형태(크립토모나드, 하프토파이트, 디노플라젤라, 크로메리드, 많은 녹조류, 황갈조류, 유글레나류)나 구형 세포 형태에서부터 아메바 형태(클로라라크니오파이트), 군체 형태, 그리고 홍조류, 일부 녹조류, 켈프와 같은 일부 황갈조류처럼 복잡한 다세포 거대 형태에 이르기까지 매우 다양한 형태적 범위를 보여준다.[11]
- 균류 유사 원생생물: 여러 독립적인 분지군의 원생생물들이 균류와 유사하게 균사와 같은 구조를 형성하고 부생영양 방식으로 살아가는 형태로 진화했다. 이들은 겉모습은 균류와 비슷하지만, 실제로는 진정한 균류와 계통적으로 매우 거리가 먼 경우가 많다. 예를 들어 난균류, 미로균류, 히포키트리드류는 스트라메노필라에 속하며, 점균류는 아메바류에, 식물미생물은 리자리아에, 퍼킨소조아는 알베올라타에 속한다.[12][13]
- 포자충: 전통적으로는 포자를 통해 번식하는 기생성 원생생물들(아피콤플렉사류, 미포자충류, 점액포자충류, 아세토스포리아류)을 포괄하는 범주로 사용되었다.[14] 현재는 주로 아피콤플렉사류[15](예: 말라리아 원인균인 ''말라리아 원충''[16]) 등으로 범위가 좁혀졌다.
4. 1. 1. 엑스카바타 (Excavata)
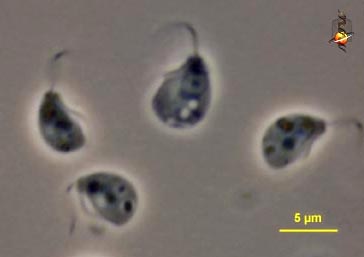
엑스카바타(Excavata)는 대부분 편모를 가진 다양한 원생생물을 포함하는 그룹으로, 호기성 및 혐기성 포식자에서 광영양 생물과 종속영양 생물에 이르기까지 다양하다.[34] '엑스카바타'라는 이름은 세포의 배면 홈(ventral groove)이라는 공통적인 특징을 나타내는데, 이 홈은 현탁 섭식에 사용되며 최근 공통 조상 진핵생물에 존재하는 조상형질로 여겨진다.[35] 엑스카바타는 디스코바(Discoba), 메타모나다(Metamonada), 말라위모나디다(Malawimonadida) 세 개의 분지(clade)로 구성되며, 각 분지에는 특징적인 배면 홈을 가진 자유생활 영양 편모류인 '전형적인 엑스카바타'가 포함된다.[36] 대부분의 계통 분석에 따르면, 이 그룹은 측계통군(paraphyletic)이며, 일부 분석에서는 진핵생물 계통수의 뿌리를 메타모나다 내부에 위치시키고 있다.[37]디스코바(Discoba)는 야코비다(Jakobida), 유글레노조아(Euglenozoa), 퍼콜로조아(Percolozoa) 세 가지 주요 그룹을 포함한다. 야코비다는 약 20종의 작은 그룹으로, 두 개의 섬모를 가진 자유생활 종속영양 편모류이며, 주로 현탁 섭식을 통해 박테리아를 섭취한다. 대부분은 수생 호기성 생물이며, 일부 혐기성 종은 해양, 기수 또는 담수에서 발견된다.[38] 이들은 박테리아와 유사한 미토콘드리아 게놈으로 가장 잘 알려져 있다. 유글레노조아는 매우 다양한 생활 방식을 가진 2,000종 이상[39]의 풍부한 편모류 그룹으로, 자유생활 종속영양(삼투영양 및 포식영양)[34] 및 광합성 유글레나(euglenid)(예: 녹조류에서 유래한 엽록체를 가진 유글레노피타(euglenophyte)), 자유생활 및 기생성 키네토플라스티드(kinetoplastid)(예: 트리파노소마(Trypanosoma)), 심해 혐기성 심비온티다(Symbiontida), 그리고 찾기 어려운 디플로네미다(diplonemid) 등이 포함된다.[40] 퍼콜로조아는 약 150종으로, 아메바, 편모류 및 아메바편모충을 포함하며, 복잡한 생활 주기를 가지고 있으며, 일부 점균류(아크라시드(acrasid))도 포함된다.[41] 유글레노조아와 퍼콜로조아 두 분지는 자매 분류군이며, 원반 모양의 미토콘드리아 크리스타를 참조하여 디스키크리스타타(Discicristata)라는 이름으로 통합되어 있다. ''츠쿠바모나스 글로보사(Tsukubamonas globosa)''종은 자유생활 편모류이며, 디스코바 내에서의 정확한 위치는 아직 확정되지 않았지만, 야코비다보다는 디스키크리스타타와 더 가까운 관계일 가능성이 높다.[38]
메타모나다(metamonad)(Metamonada)는 완전히 혐기성(anaerobe) 또는 미호기성(microaerophilic) 원생동물, 주로 편모류(flagellate)인 문이다. 일부는 흰개미와 같은 동물의 위장관(gastrointestinal tract) 공생체이고, 일부는 자유생활성이며, 일부는 기생성이다. 여기에는 포르니카타(Fornicata), 파라바살리아(Parabasalia), 프리아크소스틸라(Preaxostyla) 세 가지 주요 분지가 포함된다. 포르니카타(>140종)[42]는 두 개의 세포핵(cell nucleus)을 가진 디플로모나드(diplomonad)(예: 인간의 잘 알려진 기생충인 ''지아르디아(Giardia)'' 속)와 여러 작은 그룹의 자유생활, 공생 및 기생 원생생물(예: ''카르페디에모나스(Carpediemonas)'', 레토르타모나드(retortamonad))를 포함한다. 파라바살리아(>460종)[42]는 혐기성, 대부분 내생성 유기체의 다양한 그룹으로, 작은 기생충(인간 병원체인 ''트리코모나스 바기날리스(Trichomonas vaginalis)''와 같음)에서 나무를 먹는 흰개미와 바퀴벌레에서 발견되는 수많은 편모와 핵을 가진 거대한 장 공생체에 이르기까지 다양하다. 프리아크소스틸라는 변형된 미토콘드리아를 가진 혐기성 및 내생성 옥시모나드(oxymonad)와 전형적인 엑스카바타 형태를 가진 자유생활 미호기성 박테리아 포식 편모류 ''트리마스틱스(Trimastix)''와 ''파라트리마스틱스(Paratrimastix)'' 두 속을 포함한다.[43] 최근에 기술되었고 독특한 세포 구조를 가진 두 속의 혐기성 편모류인 ''바르텔로나(Barthelona)''와 ''스콜리오모나스(Skoliomonas)''는 포르니카타와 밀접한 관련이 있다.[44]
말라위모나드(malawimonad)(Malawimonadida)는 담수 또는 해양 현탁 섭식 박테리아 포식 편모류[45]로, 전형적인 엑스카바타 모양을 가지고 있으며, 야코비다와 일부 메타모나드와 유사하지만 대부분의 분석에서 계통적으로 가깝지 않은 작은 그룹(3종)이다.
'''엑스카바타의 대표적인 속'''
4. 1. 2. 디아포레티케스 (Diaphoretickes)
디아포레티케스(Diaphoretickes)는 진핵생물의 주요 그룹 중 하나로, 거의 모든 광합성 진핵생물을 포함하는 방대한 분류군이다. 이 그룹은 크게 TSAR 상위 분류군, 아르케플라스티다(Archaeplastida), 하프티스타(Haptista), 크립티스타(Cryptista)로 구성된다.=== TSAR 상위 분류군 ===
TSAR 상위 분류군은 스트라메노파일, 알베올라타(Alveolata), 리자리아(Rhizaria)라는 세 가지 주요 분지군을 포함하는 거대 그룹이다. 이 중 스트라메노파일은 다음과 같은 특징을 가진다.
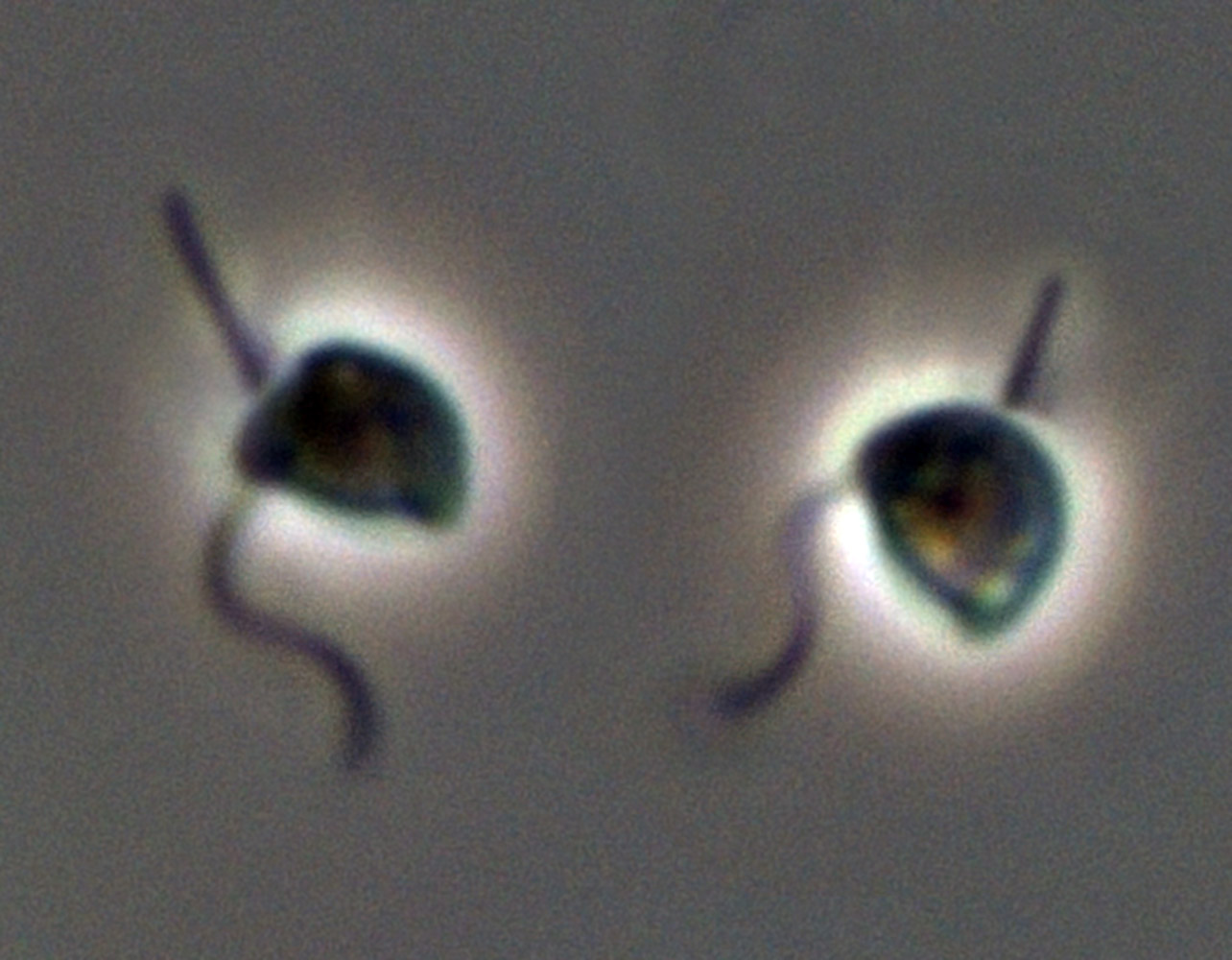
==== 스트라메노파일 (Stramenopiles / Heterokonta) ====
스트라메노파일(Heterokonta)은 두 개의 서로 다른 편모를 가지는 것이 특징인데, 그중 하나는 짧고 짚처럼 생긴 털(마스티고네메)을 많이 가지고 있다. 이 그룹은 광합성을 하는 생물 한 분류군과 다양한 환경에 존재하는 수많은 종속영양 생물 분류군을 포함한다. 스트라메노파일은 일반적으로 잘 뒷받침되는 두 개의 분류군, 비기라(Bigyra)와 기리스타(Gyrista)를 포함하지만, 비기라의 단계통성은 의문시되고 있다.[47] 비기라와 기리스타 두 분류군 모두 외부에서 분기하는 것은 수수께끼 같은 종속영양성 편모충인 단일 종 ''플라티술쿠스 타르두스(Platysulcus tardus)''이다.[47] 종속영양성 스트라메노파일의 다양성의 상당 부분은 아직 특징이 밝혀지지 않았으며, MAST(MArine STramenopiles)로 알려진 유전자 서열 계통에서 거의 전적으로 알려져 있다.[47] 그중 몇 종만이 기술되었다.[48][49]
===== 기리스타 (Gyrista) =====
기리스타 문은 광합성 생물인 황갈조류(Ochrophyta) 또는 부등편모조류(Heterokontophyta)(23,000종 이상)[39]를 포함하며, 이들은 홍조류에서 유래한 엽록체를 포함하고 있다. 여기에는 다양한 구조와 형태를 포함하는 많은 조류 계통이 있다. 가장 다양한 세 가지 황갈조류 강은 다음과 같다.
- 규조류: 단세포 또는 군체 생물로 실리카 세포벽(프러스툴)으로 둘러싸여 있으며, 다양한 모양과 장식을 보인다. 전 세계적으로 생성되는 산소의 상당 부분을 담당하고, 해양 식물 플랑크톤의 상당 부분을 구성한다.
- 갈조류: 사상체 또는 '진정한' 다세포(분화된 조직 포함) 대형 조류로서, 다시마 숲과 같은 많은 온대 및 한랭 해양 생태계의 기초를 이룬다.[50]
- 황금색 조류: 대부분 담수 서식지에 존재하는 단세포 또는 군체 편모충이다.[51]
기리스타 내부에서 황갈조류의 자매 분류군은 주로 삼투영양성(osmotrophic)이고 사상체인 의사균류(1,200종 이상)[52]이며, 세 가지 뚜렷한 계통을 포함한다.
- 난균류 또는 물곰팡이: 기생성이며, 감자 역병을 일으키는 ''피토프토라 인페스탄스(Phytophthora infestans)''가 대표적이다. 의사균류 종의 대부분을 포함한다.
- 히포키트리드: 균류와 같은 생활 방식을 유지하는 다양성이 덜한 비기생성 그룹이다.
- 비기로모나드: 세균을 잡아먹거나 진핵생물을 잡아먹는 포식성 그룹이다.[47]
작은 그룹의 육질편모충류-유사 종속영양성 아메바인 방사충류는 황갈조류 내부 또는 황갈조류의 자매 분류군으로서 불확실한 위치를 가지고 있다.[53]
===== 비기라 (Bigyra) =====
거의 연구되지 않은 비기라 문은 독점적으로 종속영양 생물의 집합체이며, 그 대부분은 자유생활을 한다. 여기에는 다음 그룹들이 포함된다.
- 미로균류: 단세포 아메바형 포식성 생물, 혼합영양 생물, 그리고 영양분을 흡수하고 이동하기 위해 점액 네트워크를 만드는 균류와 같은 사상체 종속영양 생물뿐만 아니라 일부 기생충과 몇몇 피낭성 아메바(암피트레미다)도 포함된다.
- 비코소에시드: 세균을 섭취하는 포식성 편모충이다.
- 플라시도조아: 비코소에시드와 밀접하게 관련되어 있으며, 심해 호염성 플라시디데아와 같은 몇몇 그룹의 종속영양성 편모충과 장내 공생 생물인 오팔리나타를 포함한다. 오팔리나타에는 인간 기생충인 ''블라스토시스티스(Blastocystis)''와, 수많은 핵과 편모를 가진 거대 세포로 구성된 매우 특이한 오팔리나류(원래 섬모충류로 잘못 분류됨)가 속한다.[47]
'''스트라메노파일의 대표적인 속'''

4. 1. 3. 아모르페아 (Amorphea)
아모르페아는 아메바류(Amoebozoa)와 오바조아(Obazoa)를 포함하는 분류군이다.아메바류는 불규칙하고 유연한 형태를 가지며, 세포질의 일부인 위족을 뻗어 이동하는 특징을 가진다. 이 분지군은 다양한 점균류와 피낭성 아메바 등을 포함한다. 점균류는 여러 아메바가 서로 뭉쳐 포자를 가진 다세포 단계를 형성하기도 하며, 피낭성 아메바는 주변 물질로 세포를 감싸는 껍질을 만든다.
오바조아는 일반적으로 두 개의 섬모(편모)를 가지는 편모충 그룹의 일부이다.
4. 1. 4. 고아 그룹 (Orphan groups)
많은 소규모 계통들은 기존의 주요 상위 분류군 중 어느 곳에도 명확히 속하지 않아 계통적 위치가 불분명하며, 데이터가 제한적이고 잘 알려지지 않은 경우가 많아 종종 '고아 그룹'(orphan groups)이라고 불린다.이들 중 일부, 예를 들어 '''CRuMs''' 혈통, '''Malawimonadida''', '''Ancyromonadida''' 등은 아모르페아와 관련이 있는 것으로 보인다. 또 다른 그룹인 '''Hemimastigophora'''(10종)와 '''Provora'''(7종) 등은 SAR, 아르카에플라스티다, 압티스타, 크립티스타를 포함하는 거대 분류군인 Diaphoretickes와 관련이 있거나 그 안에 속하는 것으로 추정된다. 잘 알려지지 않은 원생생물 종인 ''Meteora sporadica''는 이들 고아 그룹 중 후자 두 그룹(Hemimastigophora, Provora)과 더 가까운 관계일 가능성이 제기되었다.[33]
5. 원생생물의 생태
원생생물은 전 세계 현대 생태계에서 필수적인 존재이다. 지구 역사의 대부분 동안 모든 생태계의 유일한 진핵생물 구성 요소였으며, 광범위한 기능적 다양성을 진화시켜 중요한 생태적 역할을 수행한다. 원생생물은 1차 생산자, 여러 영양 단계의 중간자, 주요 조절 기생충 또는 기생벌, 그리고 다양한 공생 파트너로서 생태계 유지에 필수적이다.
5. 1. 서식지 다양성
원생생물은 전 세계 현대 생태계에서 필수적인 존재이다. 또한 지구 역사의 대부분 동안 모든 생태계의 유일한 진핵생물 구성 요소였으며, 이는 광범위한 기능적 다양성을 진화시켜 그 중요한 생태적 의미를 설명해 준다.원생생물은 거의 모든 서식지에 풍부하고 다양하게 존재한다. 지구 생물량에 4기가톤(Gt)을 차지하는데, 이는 동물(2Gt)의 두 배이지만 전체의 1% 미만이다. 원생생물, 동물, 고세균(7Gt), 균류(12Gt)를 합쳐도 지구 생물량의 10% 미만이며, 식물(450Gt)과 세균(70Gt)이 대부분을 차지한다. 환경 DNA 조사를 통해 확인된 원생생물의 다양성은 모든 표본 환경에서 매우 크지만, 대부분은 아직 기술되지 않았다.
가장 풍부한 원생생물 군집은 토양에 나타나며, 다음으로 해양, 그리고 마지막으로 담수 서식지(대부분 플랑크톤의 일부)에 나타난다. 담수 원생생물 군집은 토양 및 해양 플랑크톤보다 더 높은 "베타 다양성"(즉, 표본 간의 이질성이 높음)을 특징으로 한다. 높은 다양성은 극심한 홍수를 통해 다른 서식지의 생물을 모집하는 수문학적 역동성의 결과일 수 있다.[94] 토양 서식 원생생물 군집은 생태적으로 가장 풍부하며, 이는 퇴적물 내 물의 복잡하고 역동적인 분포로 인해 매우 이질적인 환경 조건이 만들어지기 때문일 수 있다. 끊임없이 변화하는 환경은 군집의 일부만 활동하도록 촉진하고 나머지는 비활성 상태를 유지하며, 이 현상은 원핵생물뿐만 아니라 원생생물에서도 높은 미생물 다양성을 촉진한다.
원생생물은 주로 수중이나 수분이 많은 토양에 서식한다. 육상에서도 햇볕이 잘 드는 곳이나 바위 위 등 건조가 심한 곳에서는 지의류처럼 다른 생물과 공생하거나, 건조할 때는 휴면하여 물이 있을 때만 활동하는 등의 방법으로 생활하는 것이 있다. 다른 생물에 기생하여 생활하는 종도 있다. 동물의 장 등에도 특수한 것이 서식하고 있지만, 기생의 경우, 공생 관계가 있는 경우, 불명확한 경우 등 다양하다. 장내나 사니층의 내부는 유기물이 풍부하고 산소가 매우 적은데, 이를 식물 출현 이전의 지구 환경에 가깝다고 보고 그러한 조건에서 서식했던 생물의 현재 서식지라고 보는 견해도 있다.
5. 2. 1차 생산자
원생생물은 전 세계 현대 생태계에서 필수적인 존재이며, 특히 1차 생산자로서 중요한 역할을 담당한다.미세한 광영양성 원생생물, 즉 미세조류는 거의 모든 수생 환경에서 생물량과 1차 생산에 크게 기여하며, 시아노박테리아와 함께 식물플랑크톤을 구성한다. 해양 식물플랑크톤 중 크기가 2 μm 미만인 피코플랑크톤과 2–20 μm 사이인 나노플랑크톤은 프림네시오피테류, 펠라고피테류, 프라시노피테류와 같은 여러 조류가 우점한다. 반면, 크기가 5 μm보다 큰 식물플랑크톤은 규조류와 디노플라젤라류가 주로 차지한다. 담수 식물플랑크톤에서는 황금색 조류, 크립토피테류, 그리고 디노플라젤라류가 가장 풍부한 그룹이다. 이러한 미세조류는 전 세계 1차 생산량의 거의 절반을 책임지며,[96] 박테리아, 고세균 및 더 높은 영양 단계의 생물들(동물플랑크톤, 어류)이 필요로 하는 에너지, 유기물, 그리고 지방산과 같은 필수 영양소의 주요 공급원이다.[95] 해양에서 이들의 풍부함은 온도나 햇빛보다는 무기 영양소의 이용 가능성에 의해 크게 좌우된다. 따라서 육지로부터 영양이 풍부한 물이 흘러드는 연안 해역이나, 영양이 풍부한 심해수가 표면으로 올라오는 지역, 즉 북극해의 용승대와 대륙붕을 따라 가장 풍부하게 분포한다.[96] 담수 서식지에 사는 대부분의 광영양성 원생생물은 혼합영양성, 즉 광합성을 하면서 동시에 다른 유기물을 섭취하는 방식으로 살아가며, 오직 섭취에만 의존하는 종속영양생물은 상대적으로 덜 풍부하다.
대형조류(홍조류, 녹조류, 갈조류)는 식물플랑크톤과 달리 일반적으로 바닥에 붙어 살기 위한 고착점이 필요하다. 이 때문에 이들의 해양 분포는 주로 암반 기질이 있는 연안 해역으로 제한된다. 대형조류는 많은 초식 동물, 특히 저서성 동물에게 먹이를 제공하고 포식자로부터 숨을 수 있는 피난처 역할을 한다. 일부 해조류 군집은 해수 표면을 떠다니며, 함께 살아가는 다른 유기체들에게 피난처를 제공하고 넓은 지역으로 퍼져나가는 분산 수단이 되기도 한다.[97][98]
광영양성 원생생물은 수생 환경뿐만 아니라 토양에서도 풍부하게 발견된다. 토양 조류가 지구 탄소 순환에 기여하는 정도는 아직 정확히 밝혀지지 않았지만, 이전에 생각했던 것보다 더 큰 역할을 할 가능성이 있다. 토양에 주로 서식하는 조류는 스트라메노필류(규조류, 황갈조류, 유스티그마토피테류)와 아르케플라스티드(녹조류)이다. 토양에서 디노플라젤라류와 하프토피테류의 환경 DNA가 발견되기도 했지만, 살아있는 형태는 관찰되지 않았다.
5. 3. 소비자
섭취영양형 원생생물은 모든 생태계에서 가장 다양한 기능적 그룹이다. 이들은 주로 세르코조아류 (담수와 토양에서 우점), 방산충류 (해양에서 우점), 비광합성 스트라메노필류 (해양보다 토양에서 풍부), 그리고 섬모충류로 대표된다.식물플랑크톤과 동물플랑크톤 사이의 일반적인 구분과 달리, 많은 해양 플랑크톤은 혼합영양 원생생물로 구성되어 있다. 이들의 중요성과 풍부함(모든 해양 환경 DNA 서열의 약 12%)은 크게 과소평가되어 왔다. 혼합영양 생물의 존재는 계절적 풍부도와 특정 혼합영양 유형에 따라 다르다.
- 구성적 혼합영양 생물: 부영양성 얕은 서식지부터 빈영양성 아열대 해역까지 거의 모든 해양 조건에서 존재하며, 대부분 유광층을 지배한다. 이들은 박테리아 포식의 대부분을 차지하며, 유해조류 대발생을 일으키기도 한다.
- 비구성적 혼합영양 생물 (색소체 보유): 유사한 생지리학적 분포와 낮은 풍부도를 가지며, 대부분 부영양성 연안 해역에서 발견된다. 일반적인 섬모충류는 유광층에서 섬모충 군집의 최대 절반을 차지할 수 있다.
- 내생 공생 혼합영양 생물: 압도적으로 가장 널리 퍼져 있고 풍부한 비구성적 유형으로, 모든 혼합영양 서열의 90% 이상을 차지하며 대부분 방산충류이다.
토양의 먹이그물에서 원생생물은 박테리아와 균류 모두의 주요 소비자이며, 이들은 더 높은 영양 단계로 영양소 흐름의 두 가지 주요 경로를 형성한다. 글리소모나드류와 세르코모나드류와 같은 아메바편모충류는 가장 풍부한 토양 원생생물 중 하나이다. 이들은 편모와 위족을 모두 가지고 있어, 토양 입자 사이를 탐색하기에 적합한 다양한 형태를 보인다. 껍질 있는 아메바 역시 토양 환경에 적응했으며, 껍질은 건조로부터 보호하는 역할을 한다.
박테리아를 섭식하는 원생생물은 암모니아(NH3) 형태로 질소를 배출하여 식물과 다른 미생물이 이용할 수 있도록 하는 중요한 역할을 한다. 전통적으로 배양 기술의 한계로 인해 원생생물은 주로 박테리아를 섭식하는 것으로 여겨졌지만, 실제로는 많은 원생생물(예: 뱀파이렐리드류, 세르코모나드류, 벌거숭이 아메바류, 껍질 있는 아메바류, 작은 편모충류)이 균류뿐만 아니라 선충류와 같은 일부 동물까지 포함한 다양한 토양 진핵생물을 섭식하는 잡식성이다. 토양 내에서 박테리아를 섭식하는 원생생물과 균류를 섭식하는 원생생물은 비슷한 생물량을 가진다.[99]
5. 4. 분해자
네크로파지(죽은 생물체의 분해)는 주로 세균과 균류가 담당하지만, 원생생물 역시 특수한 용해성 효소를 가진 분해자로서 중요한 역할을 수행한다.[100] 이들의 역할은 아직 완전히 밝혀지지 않았지만, 생태계에서 빼놓을 수 없는 부분이다.토양 환경에서는 균류와 유사한 원생생물 및 점균류들이 분해 활동을 한다. 대표적으로 난균류, 점균류, 아크라시드류 등이 있으며, 이들은 삼투영양생물이나 부생영양생물로서 죽은 유기물을 분해하여 영양분을 얻는다. 해양이나 기수 환경에서는 트라우스트키트리드류(라비린툴미세테스류)가 잘 알려진 분해자로, 죽은 식물이나 동물 조직 등 다양한 유기물을 분해한다. 또한, 여러 종류의 섬모충류나 껍질을 가진 아메바류도 죽은 동물을 분해하는 역할을 한다. 일부 핵생아메바류 아메바는 죽거나 손상된 세포만을 선택적으로 섭취하기도 한다.
하지만 위에 언급된 대부분의 원생생물 분해자는 살아있는 먹이도 섭취하는 겸용 네크로파지이다. 반면, 얕은 습지에 사는 조류를 먹는 세르코조아과 Viridiraptoridae처럼 오직 죽은 조류만을 먹이로 삼는 전문적인 네크로파지도 존재한다. 이러한 분해자들은 환경을 정화하고 다른 살아있는 미생물이 이용할 수 있도록 영양분을 방출하는 중요한 역할을 담당한다.[100]
5. 5. 기생충과 병원체
일부 원생생물은 동물, 식물, 또는 다른 원생생물에 기생하여 중요한 병원체 역할을 한다.[105][106]동물에게 기생하는 대표적인 예로는 ''Plasmodium''(말라리아 원충) 속의 5종이 있는데, 이들은 사람에게 말라리아를 일으키며, 다른 많은 종들은 다른 척추동물에게 유사한 질병을 유발한다. 식물 기생의 예로는 난균류(oomycete)인 ''Phytophthora infestans''(피토프토라 인페스탄스)가 있으며, 이는 감자에 역병을 일으켜 큰 피해를 준 바 있다.[102][103][104] 사람을 감염시킬 수 있는 원생생물은 약 100종에 달하는 것으로 알려져 있다. 2013년에는 원생동물 감염을 치료하기 위해 바이러스를 이용하는 바이로테라피의 가능성이 제기되기도 했다.[107][108]
기생성 원생생물은 생태계에서도 중요한 역할을 한다. 해양 및 토양 환경 DNA 분석 결과, 기생성 원생생물이 전체의 약 15~20%를 차지하는 것으로 나타났다. 반면 담수에서는 약 5% 정도인데, 이는 키트리디오미케스가 비슷한 생태적 지위를 차지하기 때문일 수 있다. 해양 시스템에서는 숙주를 죽이는 기생포식자(예: Syndiniales)가 더 풍부하며, 담수 생태계에서는 Perkinsea와 Syndiniales(알베올라타)가 주요 기생포식자이다. 담수에서 숙주를 죽이지 않는 진정한 기생생물은 주로 난균류, 정단복합체충류(Apicomplexa), 어포자충류(Ichthyosporea)이다. 토양 생태계에서는 동물에 기생하는 정단복합체충류와 식물에 기생하는 난균류, 플라스모디오포라강(plasmodiophorids)이 주요 진정한 기생생물이다. 특히 신열대구 삼림 토양에서는 정단복합체충류가 진핵생물 다양성의 상당 부분을 차지하며 작은 무척추동물의 기생충으로서 중요한 역할을 한다.[101]
기생성 원생생물 중 일부는 매우 복잡한 생활사를 가진다. 예를 들어, ''톡소플라스마 곤디''는 다양한 동물(중간 숙주)에서 무성 생식을 하지만, 고양잇과 동물(최종 숙주, 예: 집고양이)에서만 유성 생식을 할 수 있다.[27][28][29] ''말라리아 원충'' 역시 유성 생식과 무성 생식을 모두 포함하는 복잡한 생활사를 거친다.[30] 그러나 자연 상태에서 서로 다른 ''말라리아 원충'' 균주 간에 유전자 교환이 얼마나 자주 일어나는지는 아직 불분명하며, 대부분의 기생성 원생생물 개체군은 유전적으로 거의 교환되지 않는 클론 계통일 가능성이 있다.[31] ''라이슈마니아'' 속의 병원성 기생 원생생물은 무척추동물 매개체 내에서 트리파노소마와 유사하게 감수분열을 포함하는 유성 생식 주기를 수행할 수 있는 것으로 밝혀졌다.[32]
5. 6. 생지화학적 순환
원생생물은 전 세계 현대 생태계에서 필수적인 존재이다. 또한 원생생물은 지구 역사의 대부분 동안 모든 생태계의 유일한 진핵생물 구성 요소였으며, 이는 광범위한 기능적 다양성을 진화시켜 그 중요한 생태적 의미를 설명해 준다. 원생생물은 1차 생산자로서, 여러 영양 단계의 중간체로서, 주요 조절 기생충 또는 기생벌로서, 그리고 다양한 공생의 파트너로서 필수적이다.해양 원생생물은 생지화학적 순환, 특히 탄소 순환에 근본적인 영향을 미친다. 식물플랑크톤으로서, 이들은 모든 육상 식물을 합친 것만큼 많은 탄소를 고정한다. 토양 원생생물, 특히 피낭아메바류는 껍질의 생물광물화를 통해 숲의 나무만큼 많은 규소 순환에 기여한다.
6. 원생생물의 진화와 화석 기록
원생생물의 진화적 관계는 분자 계통 유전학, 전체 게놈 및 전사체의 염기서열 분석, 그리고 편모 장치와 세포골격에 대한 전자 현미경 연구를 통해 밝혀졌다. 새로운 주요 원생생물 계통과 생물 다양성이 계속 발견되면서 생명의 진핵생물 계통수는 극적인 변화를 겪었다. 가장 최근의 진핵생물 분류 시스템은 공식적인 분류 계급(계, 문, 강, 목 등)을 사용하지 않으며, 대신 관련 유기체의 클레이드(clade)만을 인정하는 방식이다. 이는 장기적으로 분류를 더 안정적으로 만들며 업데이트를 용이하게 한다. 이 새로운 분지학적 체계에서 원생생물은 비공식적으로 상위 그룹(supergroup)이라 불리는 다양한 가지로 나뉜다. 대부분의 광합성 진핵생물은 식물을 포함하는 상위 그룹인 아르케플라스티다(Archaeplastida)와 텔로네미아(Telonemia), 스트라메노파일(Stramenopiles), 알베올라타(Alveolata), 리자리아(Rhizaria)를 포함하는 TSAR를 포함하는 Diaphoretickes 클레이드에 속하며, 크립티스타(Cryptista) 및 합티스타(Haptista) 문도 포함한다. 동물과 균류는 아메바류(Amoebozoa) 및 기타 여러 원생생물 계통을 포함하는 Amorphea 상위 그룹에 속한다. 원시적인 세포 구조를 가진 다양한 진핵생물 그룹은 집합적으로 엑스카바타(Excavata)로 알려져 있다.
18세기 초부터 "주입 동물"(infusoria, 후에 섬모충류)이라는 용어는 원생생물, 세균 및 작은 무척추동물을 가리키는 데 사용되었다. 18세기 중반, 스웨덴 과학자 칼 폰 린네는 원생생물을 대체로 무시했으나, 덴마크의 오토 프리드리히 뮬러는 최초로 원생생물을 이명법 체계에 도입했다.
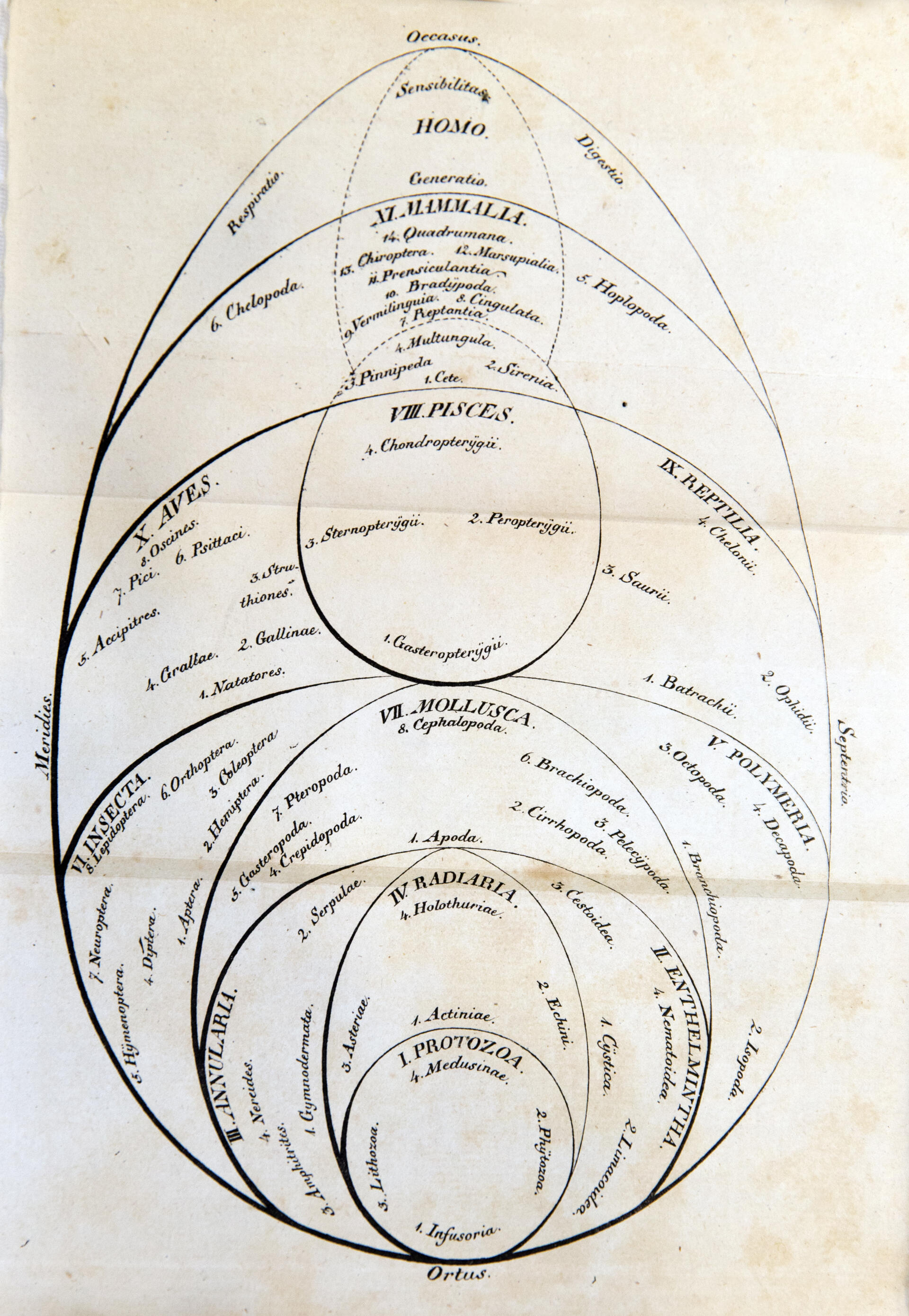
19세기 초, 독일의 자연주의자 게오르크 아우구스트 골드푸스는 원생동물(Protozoa, '초기 동물'이라는 뜻)을 동물계 내의 한 계급으로 제안했으며, 여기에는 섬모충류, 산호, 식물동물(phytozoa, 예: ''크립토모나스''), 해파리 등 매우 다른 네 그룹이 포함되었다. 이후 1845년, 카를 테오도어 폰 지볼트는 최초로 원생동물을 단세포 동물의 문으로 확립했으며, 이를 섬모충류와 근족류(Rhizopoda, 아메바, 유공충 포함)의 두 강으로 나누었다. 그러나 모든 과학자들이 이들을 동물계의 일부로 간주하지 않았고, 19세기 중반에는 미세 유기체를 원생동물(초기 동물), 원생식물(protophyta, 초기 식물), 식물동물(동물 같은 식물), 세균(대부분 식물로 간주) 그룹으로 나누는 등 식물/동물 이분법에 대한 제약이 커졌다. 1858년, 고생물학자 리처드 오언은 최초로 ''원생동물''을 "핵 세포"와 식물 및 동물의 "공통적인 유기적 특징"을 가진 진핵생물의 별개 계로 정의했으나, 해면동물도 여기에 포함시켰다.
1860년, 영국의 자연주의자 존 호그는 '''원생생물'''(Protoctista, '최초로 창조된 존재'라는 뜻)이라는 이름을 자연의 네 번째 계(다른 세 계는 린네의 식물, 동물, 광물)로 제안했다. 여기에는 모든 하등하고 원시적인 유기체(원생식물, 원생동물, 해면동물 포함)가 포함되고, 식물계와 동물계가 합쳐지는 기저에 위치한다.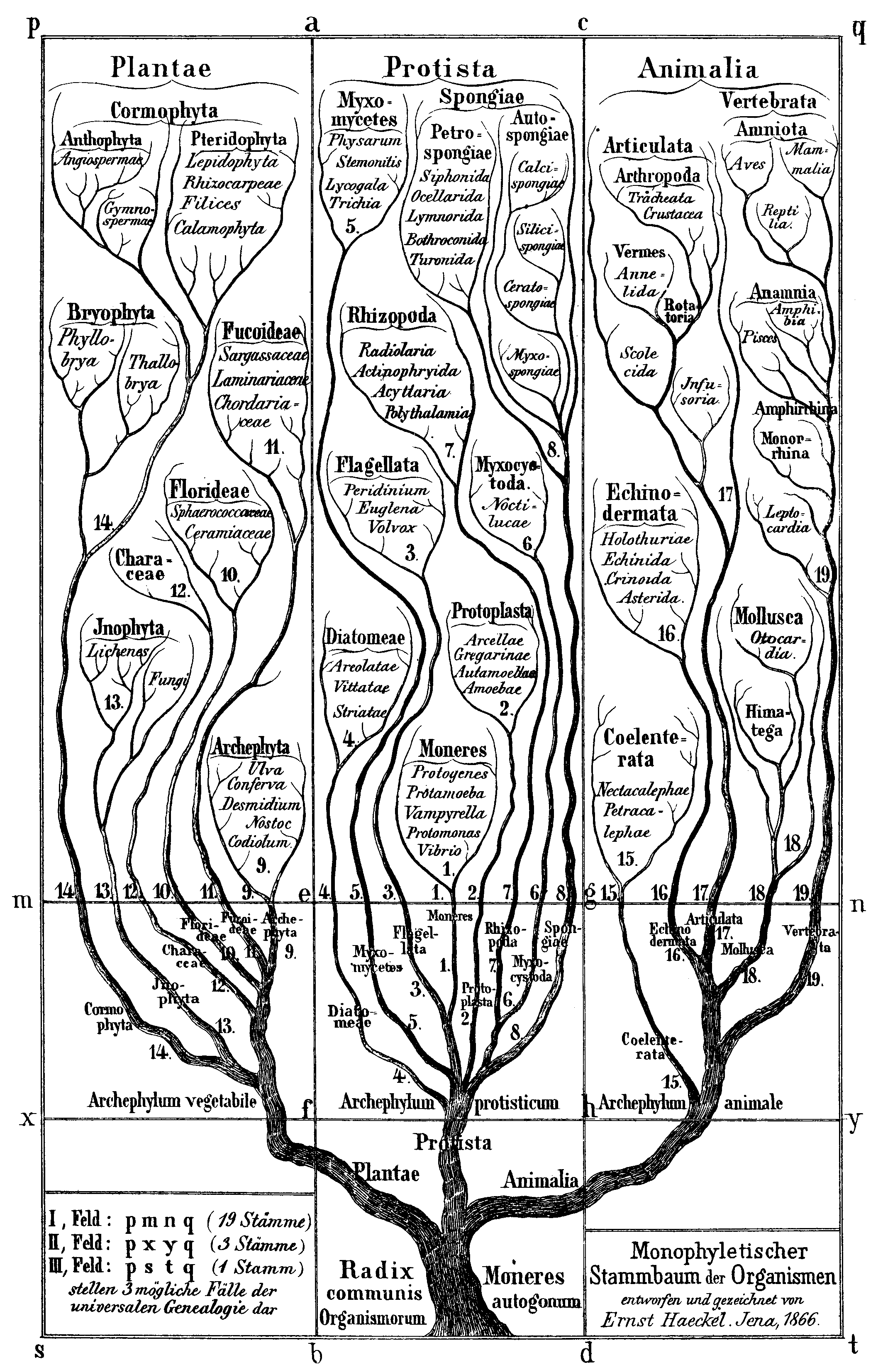
1866년, '원생생물학의 아버지'로 불리는 독일 과학자 에른스트 헤켈은 동물도 식물도 아닌 원시적인 형태를 포함하는 생명의 세 번째 계로서 ''Protistenreich'' ('''원생생물계''')를 제안했다. 이는 동물과 식물의 특징이 혼합된 유기체들을 분류하는 문제를 해결하기 위한 것이었다. 그는 세균과 진핵생물, 단세포 및 다세포 유기체 모두를 원생생물로 분류했다. 독일 동물학자 오토 뷔츠슐리가 섬모충류가 단세포임을 증명할 때까지 헤켈은 섬모충류를 동물계에 두었다. 처음에는 해면동물과 균류를 포함시켰으나, 이후 출판물에서는 원생생물을 주로 단세포 유기체 또는 조직을 형성할 수 없는 군체로 명확히 제한했다. 그는 원생생물의 특징을 유성생식의 부재로, 동물의 특징을 동물 발달의 포배 단계로 보아 원생생물을 진정한 동물과 구분했다. 그는 또한 ''원생동물''과 ''원생식물''이라는 용어를 원생생물의 아계(subkingdom)로 사용했다.
뷔츠슐리는 원생생물계가 너무 다계통군이라고 생각해 세균을 포함하는 것을 거부했다. 그는 원생생물계를 핵을 가진 단세포 동물형 생물만 포함하는 ''원생동물''(protozoa)로 분류했으며, 세균과 ''원생식물''(protophyta)은 별개의 그룹으로 나누었다. 이는 카를 테오도어 폰 지볼트의 오래된 ''원생동물''/''원생식물'' 이분법을 강화했으며, 독일 자연주의자들은 20세기 초까지 이러한 견해를 옹호했다. 그러나 영국 생물학자 C. 클리포드 도벨은 1911년 원생생물이 동물과 식물의 세포 조직과는 매우 다르게 기능한다는 점에 주목했으며, 이를 "무세포성"(acellularity)이라고 부르며 다른 조직을 가진 그룹으로서 원생생물계의 중요성을 강조했다. 그는 원생생물학(protistology)이라는 용어를 만들어 동물학과 식물학에서 독립된 연구 분야로 확립했다.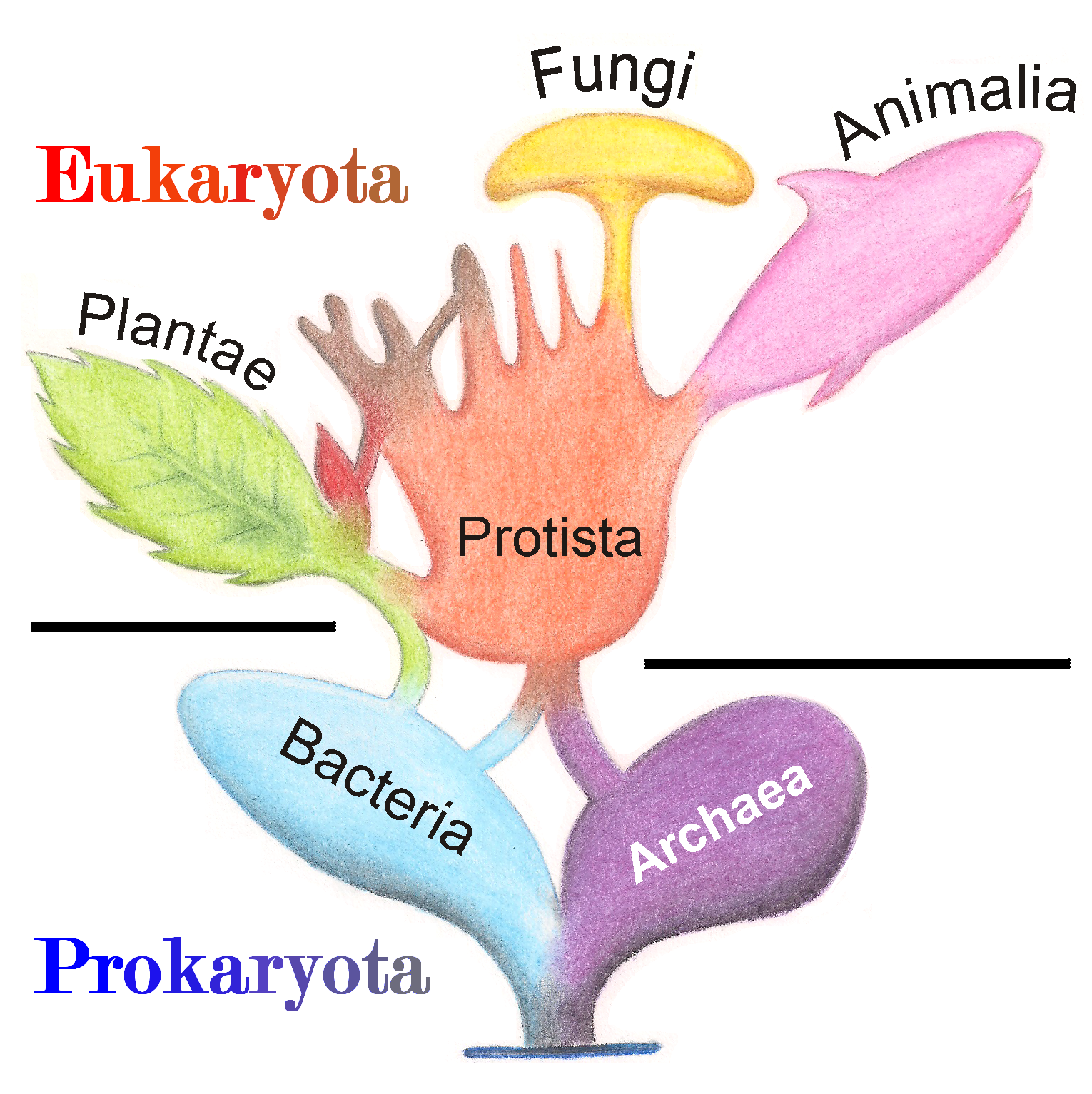
1938년, 미국의 생물학자 허버트 코플랜드는 호그의 분류 체계를 부활시키면서, 헤켈의 ''원생생물''(Protista)에는 세균과 같은 무핵 미생물이 포함되지만 ''원생생물''(Protoctista, "최초로 확립된 존재들")에는 포함되지 않는다고 주장했다. 그의 4계 분류(모네라(Monera), ''원생생물''(Protoctista), 식물계, 동물계)에서 원생생물과 세균은 마침내 분리되었으며, 무핵(원핵생물)과 핵을 가진(진핵생물) 생물의 차이가 인식되었다. 그는 헤켈의 진정한 동물에 대한 배낭포(blastular) 정의를 따라 원생생물을 식물과 확실하게 구분했으며, 배우체(embryophyta, 진정한 식물)를 엽록소 ''a'', ''b'', 카로틴, 크산토필을 가지고 녹말을 생성하는 것으로 정의할 것을 제안했다. 또한 그는 단세포/다세포 이분법이 유효하지 않다는 것을 처음으로 인식했다. 그럼에도 불구하고 그는 홍조류, 갈조류, 그리고 원생동물과 함께 균류를 원생생물계에 남겨두었다.[109] 이 분류는 휘태커의 후속 분류에서 균계, 동물계, 식물계, 원생생물계를 생명의 4계로 정의하는 기초가 되었다.[110]
미국의 식물 생태학자 로버트 휘태커가 1969년에 발표한 인기 있는 5계 분류 체계에서 원생생물계는 진핵생물 중 "단세포이거나 단세포-군체성이며 조직을 형성하지 않는 생물"로 정의되었다. 원핵생물/진핵생물의 구분이 주류가 되면서, 휘태커는[110] 원핵생물인 모네라계와 진핵생물계(동물계(섭취), 식물계(광합성), 균계(흡수) 및 나머지 원생생물계) 사이의 근본적인 생명의 분열을 인식했다.[111]
미국의 진화생물학자 린 마굴리스의 5계 분류 체계에서 "원생생물"(protist)이라는 용어는 미생물에 대해 사용되었으며, 보다 포괄적인 원생생물계(Protoctista 또는 '''원생생물'''(protoctists))에는 켈프, 홍조류, 점균류와 같은 특정한 큰 다세포 진핵생물이 포함되었다.[112] 일부는 마굴리스의 ''원생생물''(protoctist)과 ''원생생물''(protist)이라는 용어를 서로 바꿔 사용해 단세포 및 다세포 진핵생물(전문화된 조직을 형성하지만 다른 전통적인 계에는 속하지 않는 것 포함)을 모두 포함한다.[113]
5계 분류 체계는 20세기 후반 분자계통유전학의 발전으로 원생생물이 동물, 균류, 식물이 진화한 측계통군임이 명확해지기 전까지 받아들여진 분류였다. 이후 3역 분류 체계(세균, 고세균, 진핵생물)가 널리 퍼졌다. 오늘날 원생생물은 공식적인 분류군으로 취급되지는 않으나, 두 가지 방식으로 편의상 일반적으로 사용된다.[117]
- 계통발생학적 정의: 원생생물은 측계통군이다.[114] 원생생물은 동물, 육상식물 또는 균류가 아닌 모든 진핵생물이다.[115] 따라서 과거 원생생물에 포함되었던 균류의 미세포자충류, 키트리디오미세테스, 효모와 같은 많은 단세포 군집과 비단세포성 믹소조아 동물을 제외한다.[116]
- 기능적 정의: 원생생물은 본질적으로 독립된 세포로 존재하거나, 군체로 존재하는 경우에도 조직으로 분화되지 않는 진핵생물이다.[117] 일반적인 용어로는 원생생물이 보이는 비군체성 다세포성 유형의 다양성(예: 칼라목과 같은 집합성 또는 갈조류와 같은 복잡한 다세포성)을 제외한다.[119]
그러나 21세기까지 지속된 전통적인 계급에 기반한 원생생물의 분류가 하나 있다. 영국의 원생동물학자 토마스 캐벌리어-스미스는 1998년부터 6계 분류 체계를 개발했다. 그의 체계는 세균, 동물계, 식물계, 균계, ''원생동물'' 및 ''색소체류''의 6개 계로 구성된다. 그의 분류에서는 측계통군이 분지군보다 우선시된다. ''원생동물''과 ''색소체류'' 두 원생생물 계는 문과 같은 측계통군, 예를 들어 아푸소조아, 에올루카 또는 후편모충류를 포함한다. 또한 홍조류와 녹조류는 진정한 식물로 간주되며, 균류 그룹인 미세포자충류, 로젤리다 및 아펠리다는 문 후편모충류 아래 원생동물로 간주된다. 이 체계는 그의 마지막 출판물이 나온 2021년까지 유지되었다.
6. 1. 중원생대
모든 살아있는 진핵생물(원생생물 포함)은 최근 공통 조상 진핵생물(LECA)로부터 진화했다. 이 조상의 후손은 "왕관군" 또는 "현생" 진핵생물로 알려져 있다. 분자시계 연구에 따르면 LECA는 약 18억 년 전에서 12억 년 전 사이에 출현한 것으로 추정된다. 모든 분자 예측에 따르면, 현생 진핵생물은 10억 년 전 이전에 이미 유성생식이 가능한 다세포 조류나 식세포작용 및 운동성을 가진 단세포 원생생물의 형태로 형태학적, 생태적 다양성을 갖추었다. 그러나 이 시기 현생 진핵생물의 화석 기록은 매우 부족하여 예측된 다양성과는 차이를 보인다.[120]대신 이 시기의 화석 기록에는 "줄기군 진핵생물"이 포함되어 있다. 이 화석들은 알려진 어떤 왕관군에도 속하지 않으며, 아마도 LECA 이전에 갈라져 나온 멸종된 계통일 것으로 보인다. 이들은 중원생대(약 16억 5천만 년 전 ~ 10억 년 전) 화석 기록 전반에 걸쳐 꾸준히 나타난다. 줄기군 진핵생물은 복잡한 세포벽 장식이나 세포막 돌출부처럼 유연한 내막계가 필요한 진핵생물의 특징을 보여준다. 하지만 이들은 세포막 구성에서 왕관 진핵생물과 중요한 차이가 있었다. 왕관 진핵생물은 세포막에 콜레스테롤이나 에르고스테롤과 같은 "왕관 스테롤"을 생성하는 반면, 줄기 진핵생물은 생합성 경로에서 더 일찍 나타나는 원시스테롤을 만들었다.[120]
왕관 스테롤은 생합성 과정에서 더 많은 에너지가 필요하지만, LECA의 후손들에게 몇 가지 진화적 이점을 제공했을 것으로 여겨진다. 왕관 스테롤의 특정 불포화 패턴은 건조와 재수화가 반복되는 환경에서 삼투압 충격으로부터 세포를 보호한다. 또한 왕관 스테롤은 에틸기를 받아들여 지질 사이의 결합력을 강화함으로써 극한의 추위나 더위에 대한 적응력을 높일 수 있다. 생합성 경로의 추가 단계를 통해 세포는 막 내 서로 다른 스테롤의 비율을 조절할 수 있게 되었는데, 이는 더 넓은 온도 범위에서 생존하고 비대칭 세포 분열이나 자외선 노출로 손상된 막을 수리하는 등의 독특한 메커니즘을 가능하게 했다. 더 나아가, 이러한 스테롤이 원생대 변화하는 산소 농도로부터 세포를 보호하는 역할을 했을 것이라는 추측도 있다. 이러한 스테롤 기반 메커니즘 덕분에 LECA의 후손들은 당시 극한 환경에서 살아가며, 건조와 재수화 주기가 반복되고, 고온과 저온의 일교차가 크며, 높은 자외선에 노출되는 생태적 틈새(예: 갯벌, 강, 불안정한 해안선, 지상 토양)로 다양화할 수 있었을 것으로 이론화된다.[120]
반면, 줄기군 진핵생물은 원시스테롤만 생성할 수 있었기 때문에 이러한 적응 메커니즘을 가지지 못했다. 이들은 주로 개방된 해양 환경에 서식했으며, 당시 산소가 부족했던 중원생대 바다에서 번성했던 통성 혐기성 생물이었을 가능성이 높다. 이후 토니안기(신원생대) 동안 지구의 산소 농도가 증가하자, 산소가 풍부한 환경을 선호하는 왕관 진핵생물이 개방 해양으로 진출하면서 경쟁이 심화되었고, 이것이 줄기 진핵생물의 멸종 원인 중 하나였을 수 있다. 또한, 이들의 원시스테롤 기반 세포막은 크라이오게니안기의 "눈덩이 지구" 빙하기와 같은 극심한 추위나 이후의 급격한 온난화 환경에서 생존에 불리했을 수 있다.[120]
6. 2. 신원생대
현생 진핵생물은 풍부한 홍조류의 증가에 힘입어 토니아기(10억~7억 2천만 년 전)에 다량으로 나타나기 시작했다. 현생 진핵생물로 분류되는 가장 오래된 화석은 두 가지 광합성 원생생물에 속한다. 다세포 홍조류인 ''Bangiomorpha''(10억 5천만 년 전)와 녹조류인 ''Proterocladus''(10억 년 전)이다.[120] 다수의 이종영양성 원생생물 화석은 그보다 늦은 약 9억 년 전, 균류의 출현과 함께 나타난다.[120] 예를 들어, 가장 오래된 Amoebozoa 화석은 현생 유공충과 유사한 항아리 모양의 미세화석으로, 8억 년 된 암석에서 발견되었다. 방산충 껍질은 캄브리아기(~5억 년 전) 이후 화석 기록에서 풍부하게 발견되지만, 최근의 고생물학 연구는 일부 선캄브리아기 화석을 방산충의 가장 초기 증거로 해석하기 시작했다.[121][122][123]참조
[1]
논문
Protists push animals aside in rule revamp
2005-11-01
[2]
서적
Introduction to Paleobiology and the Fossil Record
https://archive.org/[...]
Wiley-Blackwell
[3]
논문
Microsporidia: emerging pathogenic protists
2001-01-01
[4]
논문
Morphology, phylogeny, and ecology of the aphelids (Aphelidea, Opisthokonta) and proposal for the new superphylum Opisthosporidia
2014-01-01
[5]
논문
Giant deep-sea protist produces bilaterian-like traces
2008-01-01
[6]
논문
Giant protists (xenophyophores) function as fish nurseries
2019-01-01
[7]
논문
Lynn Margulis replies
1986-01-01
[8]
논문
Undulipodia, flagella and cilia
1980-01-01
[9]
논문
Terminology and nomenclature of the cytoskeletal elements associated with the flagellar/ciliary apparatus in protists
1991-01-01
[10]
Q
[11]
논문
Protist diversity: Novel groups enrich the algal tree of life
2021-01-01
[12]
논문
What has happened to the "aquatic phycomycetes" (sensu Sparrow)? Part II: Shared properties of zoosporic true fungi and fungus-like microorganisms
2018-01-01
[13]
서적
Marine fungi
De Gruyter
2012-01-01
[14]
논문
A newly revised classification of the Protozoa
1980-01-01
[15]
논문
Kingdom Protozoa and its 18 phyla
1993-01-01
[16]
웹사이트
Facts about malaria
https://www.ecdc.eur[...]
2017-06-09
[17]
논문
Evolutionary cell biology of proteins from protists to humans and plants
http://nbn-resolving[...]
[18]
논문
Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences.
2009-05-25
[19]
서적
DNA Repair: New Research
Nova Sci. Publ.
2012-01-01
[20]
논문
Morphological and ecological complexity in early eukaryotic ecosystems
2001-07-01
[21]
논문
A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis
2005-01-01
[22]
논문
Population genetics provides evidence for recombination in Giardia
2007-11-01
[23]
논문
An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis
2007-08-01
[24]
논문
The first sexual lineage and the relevance of facultative sex
1999-06-01
[25]
논문
The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms
2011-07-01
[26]
논문
Chromidia and the binuclearity hypotheses: A review and a criticism
http://jcs.biologist[...]
[27]
웹사이트
CDC – Toxoplasmosis – Biology
https://www.cdc.gov/[...]
2015-03-17
[28]
웹사이트
Cat parasite linked to mental illness, schizophrenia
http://www.cbsnews.c[...]
CBS
2015-06-05
[29]
웹사이트
CDC – About Parasites
https://www.cdc.gov/[...]
[30]
논문
Gametocytogenesis: the puberty of Plasmodium falciparum
2004-07-01
[31]
논문
Are eukaryotic microorganisms clonal or sexual? A population genetics vantage
1991-06-01
[32]
논문
Demonstration of genetic exchange during cyclical development of Leishmania in the sand fly vector
2009-04-01
[33]
논문
''Meteora sporadica'', a protist with incredible cell architecture, is related to Hemimastigophora
2024-01-22
[34]
서적
Brock Biology of Microorganisms
Pearson
2019-01-01
[35]
논문
Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota)
2003-01-01
[36]
논문
The function of the feeding groove of 'typical excavate' flagellates
2023
[37]
논문
An excavate root for the eukaryote tree of life
2023
[38]
서적
Handbook of the Protists
Springer
2017
[39]
논문
How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing
2024
[40]
논문
Euglenozoa: taxonomy, diversity and ecology, symbioses and viruses
2021
[41]
서적
Handbook of the Protists
Springer
2017
[42]
논문
Diversity, Nomenclature, and Taxonomy of Protists
2007
[43]
논문
Genomics of Preaxostyla Flagellates Illuminates the Path Towards the Loss of Mitochondria
2023
[44]
논문
Characterization of ''Skoliomonas'' gen. nov., a haloalkaliphilic anaerobe related to barthelonids (Metamonada)
2024
[45]
논문
Description of Imasa heleensis, gen. nov., sp. nov. (Imasidae, fam. nov.), a Deep-Branching Marine Malawimonad and Possible Key Taxon in Understanding Early Eukaryotic Evolution
December 2020
[46]
논문
On the origin of TSAR: morphology, diversity and phylogeny of Telonemia
The Royal Society
[47]
논문
Integrated overview of stramenopile ecology, taxonomy, and heterotrophic origin
2024
[48]
논문
Kaonashia insperata gen. et sp. nov., a eukaryotrophic flagellate, represents a novel major lineage of heterotrophic stramenopiles
2023
[49]
논문
Phylogenomic position of genetically diverse phagotrophic stramenopile flagellates in the sediment-associated MAST-6 lineage and a potentially halotolerant placididean
2025
[50]
서적
Handbook of the Protists
Springer
2017
[51]
서적
Handbook of the Protists
Springer
2017
[52]
논문
Oomycetes
2018
[53]
논문
An Enigmatic Stramenopile Sheds Light on Early Evolution in Ochrophyta Plastid Organellogenesis
10 April 2022
[54]
논문
A Review for the Special Issue on ''Paramecium'' as a Modern Model Organism
2023
[55]
서적
Handbook of the Protists
Springer
2017
[56]
논문
Colponemids Represent Multiple Ancient Alveolate Lineages
2013
[57]
논문
Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives
2015
[58]
서적
Handbook of the Protists
Springer
2017
[59]
논문
Emerging Parasitic Protists: The Case of Perkinsea
2021
[60]
서적
Handbook of the Protists
Springer
2017
[61]
논문
Plastid Complexity in Dinoflagellates: A Picture of Gains, Losses, Replacements and Revisions
https://www.reposito[...]
2017
[62]
논문
Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria
2018
[63]
논문
Rhizaria
2014
[64]
논문
Diversity and ecology of Radiolaria in modern oceans
2022
[65]
논문
Next-Generation Environmental Diversity Surveys of Foraminifera: Preparing the Future
https://www.journals[...]
2014-10-01
[66]
서적
Handbook of the Protists
Springer
2017
[67]
논문
Evolutionary Relationship of the Scale-Bearing Kraken (incertae sedis, Monadofilosa, Cercozoa, Rhizaria): Combining Ultrastructure Data and a Two-Gene Phylogeny
July 2017
[68]
논문
Aggregative multicellularity evolved independently in the eukaryotic supergroup Rhizaria
2012
[69]
논문
Novel Cultured Protists Identify Deep-branching Environmental DNA Clades of Cercozoa: New Genera ''Tremula'', ''Micrometopion'', ''Minimassisteria'', ''Nudifila'', ''Peregrinia''
https://www.scienced[...]
2011
[70]
논문
Ascetosporea
2019
[71]
논문
Revised Taxonomy and Expanded Biodiversity of the Phytomyxea (Rhizaria, Endomyxa)
2020-01-01
[72]
논문
The Vampyrellid Amoebae (Vampyrellida, Rhizaria)
2022-01-01
[73]
서적
Handbook of the Protists
Springer
2017-01-01
[74]
논문
Rhizarian 'Novel Clade 10' revealed as abundant and diverse planktonic and terrestrial flagellates, including ''Aquavolon'' n. gen.
2018-01-01
[75]
논문
Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista
2015-01-01
[76]
논문
The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins
2012-01-01
[77]
논문
The closest lineage of Archaeplastida is revealed by phylogenomics analyses that include Microheliella maris
2022-01-01
[78]
서적
Handbook of the Protists
Springer
2017-01-01
[79]
논문
Molecular phylogeny, scale evolution and taxonomy of centrohelid heliozoa
2007-01-01
[80]
논문
Centrohelid heliozoans of Ukraine with a description of a new genus and species (Haptista: Centroplasthelida)
2022-01-01
[81]
서적
Handbook of the Protists
Springer
2017-01-01
[82]
논문
Palpitomonas bilix represents a basal cryptist lineage: insight into the character evolution in Cryptista
2014-04-10
[83]
서적
Handbook of the Protists
Springer
2017-01-01
[84]
서적
Advances in Botanical Research
Academic Press
2012-01-01
[85]
서적
Handbook of the Protists
Springer
2017-01-01
[86]
논문
Morphological and molecular characterization of a new member of the phylum Rhodelphidia
2023-01-01
[87]
논문
Non-photosynthetic predators are sister to red algae
https://www.nature.c[...]
2019-08-01
[88]
논문
Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae
2021-01-01
[89]
논문
The genome of Prasinoderma coloniale unveils the existence of a third phylum within green plants
2020-06-22
[90]
서적
Handbook of the Protists
Springer
2017-01-01
[91]
서적
Handbook of the Protists
Springer
2017-01-01
[92]
서적
Handbook of the Protists
Springer
2017-01-01
[93]
논문
New Lineage of Microbial Predators Adds Complexity to Reconstructing the Evolutionary Origin of Animals
2020-01-01
[94]
논문
Freshwater protists: unveiling the unexplored in a large floodplain system
2022-01-01
[95]
논문
Protists are microbes too: a perspective
2009-01-01
[96]
논문
Microalgae
2023-02-06
[97]
논문
Macroalgae
2021-02-07
[98]
서적
Seaweed Biology: Novel Insights into Ecophysiology, Ecology and Utilization
Springer
2012-01-01
[99]
논문
The soil food web revisited: Diverse and widespread mycophagous soil protists
2016-01-01
[100]
논문
Broad- range necrophytophagy in the flagellate ''Orciraptor agilis'' (Viridiraptoridae, Cercozoa) and the underappreciated role of scavenging among protists
2024-11-03
[101]
논문
Parasites dominate hyperdiverse soil protist communities in Neotropical rainforests
2017-01-01
[102]
논문
Not in your usual Top 10: protists that infect plants and algae
2018-04-01
[103]
논문
The Top 10 oomycete pathogens in molecular plant pathology
2015-05-01
[104]
서적
Biology
Pearson Benjamin Cummings
2008-01-01
[105]
간행물
Diseases of protozoa
https://www.int-res.[...]
John Wiley & Sons
1980-01-01
[106]
서적
Systematics of parasitic protozoa
Academic Press
[107]
학술지
Beyond phage therapy: virotherapy of protozoal diseases
2013-07
[108]
학술지
Fleas and smaller fleas: virotherapy for parasite infections
2013-05
[109]
학술지
The Kingdoms of Organisms
[110]
학술지
On the Broad Classification of Organisms
[111]
학술지
New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms
1969-01
[112]
서적
Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth
https://books.google[...]
Academic Press
2009-03-19
[113]
서적
Handbook of the Protists
https://www.springer[...]
Springer International Publishing
2017
[114]
학술지
Protists – A textbook example for a paraphyletic taxon☆
[115]
웹사이트
Protista
http://www.microbewo[...]
microbeworld.org
2016-06-11
[116]
학술지
Actinomyxidies, nouveau groupe de Mesozoaires parent des Myxosporidies
[117]
학술지
The other eukaryotes in light of evolutionary protistology
[118]
학술지
The new higher level classification of eukaryotes with emphasis on the taxonomy of protists
[119]
서적
The Evolution of Multicellularity
CRC Press
2022
[120]
학술지
Lost world of complex life and the late rise of the eukaryotic crown
https://hal.science/[...]
2023
[121]
학술지
New Siliceous Microfossils from the Terreneuvian Yanjiahe Formation, South China: The Possible Earliest Radiolarian Fossil Record
2018-08-14
[122]
학술지
Early Cambrian radiolarians and sponge spicules from the Niujiaohe Formation in South China
2019-09
[123]
학술지
The identification of putative Lower Cambrian Radiolaria
2017-06
[124]
학술지
Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes
[125]
학술지
Microbial predators form a new supergroup of eukaryotes
2022
[126]
서적
人間のための一般生物学
裳華房
2010-03-10
[127]
서적
人間のための一般生物学
裳華房
2010-03-10
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com