2025. 8. 19. 오후 11:36:45
전 세계 3억 명의 우울한 뇌…국내 연구진, 신호 경로 비밀 풀어 [건강한겨레]
출처: 한겨레 ( 한국 / 한국어 )
광유전학은 빛을 사용하여 유전자 조작된 세포의 활동을 정밀하게 제어하는 생물학 기술이다. 1979년 프랜시스 크릭의 아이디어를 시작으로, 2000년대 초 채널로돕신과 같은 광활성 단백질의 발견과 개발을 통해 발전했다. 광유전학은 신경 회로 연구, 질병 연구 및 치료, 행동 연구, 세포 생물학 연구 등 다양한 분야에 응용되며, 뇌와 같은 복잡한 생체 조직의 기능을 이해하는 데 기여한다. 기술적 과제로는 선택적 발현, 반응 속도, 빛 흡수 스펙트럼, 공간적 반응 등이 있으며, 이 분야의 발전에 기여한 과학자들은 여러 권위 있는 상을 수상했다.
| 광유전학 | |
|---|---|
| 개요 | |
 | |
| 정의 | 빛을 사용하여 뉴런 또는 다른 세포 유형의 활동을 제어하는 생물학적 기술 |
| 역사 | |
| 최초 보고 | 2002년 네이처 생명공학에 발표된 Shimizu-Sato 등의 논문 |
| 본격적인 발전 | 2000년대 후반, 칼 데이세로스 연구팀이 광유전학 기술을 신경과학에 적용하면서 급속히 발전 |
| 원리 | |
| 핵심 요소 | 빛에 반응하는 단백질 (광활성 단백질) 유전자 조작 기술 광 자극 시스템 |
| 광활성 단백질 | 채널로돕신 (ChR2): 청색광에 반응하여 양이온 채널을 열어 세포를 활성화 할로로돕신 (NpHR): 황색광에 반응하여 염화 이온 펌프를 작동시켜 세포를 억제 |
| 작동 방식 | 특정 세포에 광활성 단백질을 발현시킨 후, 빛을 비추어 세포의 활동을 조절 |
| 응용 분야 | |
| 신경과학 | 특정 신경 회로의 기능 연구 행동 조절 메커니즘 규명 기억, 학습, 중독 등 다양한 인지 과정 연구 |
| 질병 연구 | 파킨슨병, 알츠하이머병 등 신경 질환 모델 연구 정신 질환의 신경 생물학적 기초 연구 |
| 행동 연구 | 초파리의 먹이 섭취 행동 조절 Musso, Pierre-Yves; Junca, Pierre; Jelen, Meghan; Feldman-Kiss, Damian; Zhang, Han; Chan, Rachel CW; Gordon, Michael D. (2019-07-19). "Closed-loop optogenetic activation of peripheral or central neurons modulates feeding in freely moving Drosophila". eLife. 8: e45636. doi:10.7554/eLife.45636. ISSN 2050-084X. PMC 6668987. PMID 31322499. 초파리의 후진 보행 운동 회로 조절 Feng, Kai; Sen, Rajyashree; Minegishi, Ryo; Dübbert, Michael; Bockemühl, Till; Büschges, Ansgar; Dickson, Barry J. (2020-12-02). "Distributed control of motor circuits for backward walking in Drosophila". Nature Communications. 11 (1): 6166. doi:10.1038/s41467-020-19936-x. ISSN 2041-1723. PMC 7710706. PMID 33268800. 초파리의 다리 고유수용성 감각 회로 연구 Chen, Chenghao; Agrawal, Sweta; Mark, Brandon; Mamiya, Akira; Sustar, Anne; Phelps, Jasper S.; Lee, Wei-Chung Allen; Dickson, Barry J.; Card, Gwyneth M.; Tuthill, John C. (2021-12-06). "Functional architecture of neural circuits for leg proprioception in Drosophila". Current Biology. 31 (23): 5163–5175.e7. doi:10.1016/j.cub.2021.09.035. ISSN 0960-9822. PMC 8665017. PMID 34637749. 초파리의 중앙 복합체의 기능적 연결체 구축 Franconville, Romain; Beron, Celia; Jayaraman, Vivek (2018-08-20). "Building a functional connectome of the Drosophila central complex". eLife. 7: e37017. doi:10.7554/eLife.37017. ISSN 2050-084X. PMC 6150698. PMID 30124430. |
| 장점 | |
| 높은 시간 및 공간 해상도 | 밀리초 단위의 시간 해상도와 특정 세포 수준의 공간 해상도로 신경 활동 조절 가능 |
| 세포 유형 특이성 | 특정 유전자 발현 세포에만 작용하여 정밀한 제어 가능 |
| 가역성 | 빛 자극을 끄면 세포 활동이 원래 상태로 회복 |
| 단점 | |
| 침습성 | 유전자 조작을 위해 뇌에 직접 바이러스 주입 필요 |
| 빛의 침투 깊이 제한 | 빛이 뇌 조직 깊숙이 침투하기 어려워 적용 범위 제한적 |
| 면역 반응 | 광활성 단백질에 대한 면역 반응 가능성 |
| 기술 발전 | |
| 다양한 광활성 단백질 개발 | 다양한 파장의 빛에 반응하는 단백질, 더 강력한 활성 또는 억제 효과를 가진 단백질 개발 |
| 비침습적 광 자극 기술 개발 | 초음파, 자기장 등을 이용한 광 자극 기술 연구 |
| 광유전학 도구의 정밀성 향상 | 세포 유형 특이적 발현 시스템 개선, 빛 자극 전달 시스템 개선 |
| 참고 문헌 | |
1979년, 프랜시스 크릭은 뇌의 특정 유형 세포만을 제어하고 다른 세포는 거의 변경하지 않는 것이 신경과학의 과제라고 제안했다. 크릭은 빛을 사용하는 기술이 시간적, 공간적 정밀도로 신경 활동을 제어하는 데 유용할 수 있다고 추측했지만, 당시에는 뉴런을 빛에 반응하게 만드는 기술이 없었다.
1990년대 초, LC 카츠와 E. 캘러웨이는 빛이 글루탐산을 언케이지할 수 있음을 보여주었다.[17] 1994년 헤버레와 뷜트는 효모에서 박테리오로돕신을 이용해 빛으로 활성화되는 이온 흐름을 보여주었다.[18] 1995년, 게오르그 나겔 등과 에른스트 밤베르크는 미생물 로돕신(박테리오로돕신 포함)의 이종 발현으로 빛 유도 전류를 확인했다.
2002년 1월, 보리스 제멜만과 게로 미젠뵈크는 ''초파리'' 로돕신 배양 포유류 뉴런을 사용하여 빛으로 뉴런을 제어하는 최초의 유전자 표적 방식을 보고했다.[19] 2003년, 제멜만과 미젠뵈크는 단일 이온성 채널 TRPV1, TRPM8 및 P2X2가 광케이지된 리간드에 의해 게이트되는 뉴런의 빛 의존적 활성화를 위한 두 번째 방식을 개발했다.[20] 2004년부터 크라머와 이사코프 그룹은 트라우너 그룹과 협력하여 유전자 도입 이온 채널과 상호 작용하는 유기 광스위치("가역적으로 케이지된" 화합물)를 개발했다.[21][22] TRPV1 방법론은 이후 여러 연구실에서 실험 동물의 먹이 섭취, 이동 및 행동 회복력을 변경하는 데 사용되었다.[23][24][25] 그러나 뉴런 활동 변경을 위한 빛 기반 접근 방식은 채널로돕신이 더 쉽게 사용 가능해지면서 원래 연구실 밖에서는 적용되지 않았다.[26]
페터 헤게만은 레겐스부르크 대학교에서 녹조류의 광주성 연구 중, 고전적인 G 단백질 결합 로돕신으로 설명할 수 없는 빠른 광전류를 발견했다.[27] 프랑크푸르트 막스 플랑크 연구소의 게오르그 나겔과 협력하여, 조류 ''클라미도모나스 레이나르티''의 단일 유전자가 개구리 난모세포에서 발현될 때 큰 광전류를 생성함을 입증했다.[28] 발현 세포 식별을 위해 조류 단백질의 세포질 꼬리를 형광 단백질 YFP로 대체하여 최초의 일반적 광유전학 도구를 생성했다.[26] 2003년 논문에서 "난모세포 또는 포유류 세포에서 ChR2 발현은 단순 조명으로 세포질 Ca2+ 농도를 증가시키거나 세포막을 탈분극화시키는 강력한 도구"라고 말했다.
스탠포드 대학교 칼 다이서로스는 2004년 7월 초 채널로돕신 발현 뉴런의 빛 활성화를 보여주는 실험 노트를 출판했다.[29] 2005년 8월, 그의 연구실(대학원생 에드 보이든, 펑 장 포함)은 게오르그 나겔과 협력, 게오르그 나겔의 채널로돕신-2(H134R)-eYFP 돌연변이를 사용하여 신경 세포에서 단일 구성 요소 광유전학 시스템의 첫 시연을 발표했다.[30] 이 돌연변이는 게오르그 나겔과 헤게만이 기능적 특성을 처음 규명한 이후 채널로돕신-2의 첫 번째 돌연변이였다.[26]
웨인 주립 대학교 주오-화 판은 실명 시력 회복 연구 중, 뇌와 연결되는 망막 신경절 세포에서 채널로돕신을 시험했다. 판에 따르면, 채널로돕신으로 망막 뉴런의 광학적 활성화를 처음 관찰한 것은 2004년 2월이었다.[31] 이는 다이서로스가 2004년 7월에 처음 관찰하기 5개월 전이었다.[32] 2005년 주오-화 판은 쥐 망막 신경절 세포에서 채널로돕신의 생체 내 형질 전환과 망막 절편 배양에서 광자극에 대한 전기적 반응을 보고했다.[33] 이 접근법은 2021년 보톤드 로스카와 동료들에 의해 인간 환자에게 실현되었다.[88]
2005년 4월, 수사나 리마와 미젠뵈크는 유전자 표적 P2X2 광자극을 사용하여 동물의 행동을 제어하는 첫 사례를 보고했다.[34] 도파민성 뉴런 그룹의 광자극이 초파리에서 특징적인 행동 변화를 유발함을 보였다.
2005년 10월, 린 랜드메서와 스테판 헐리체는 채널로호돕신-2를 사용하여 배양된 해마 뉴런과 발달 중인 닭 척수 회로에서 신경 활동을 제어하는 연구 결과를 발표했다.[35] 또한, 해마 뉴런과 발달 중인 닭 배아에서 세포 내 신호 전달 경로를 동원, 신경 활동을 억제하는 도구로 척추 동물 로돕신(빛 활성화 G 단백질 결합 수용체)을 처음 도입했다.[35]
알렉산더 고트샬크와 게오르그 나겔 그룹은 최초의 ChR2 돌연변이(H134R)를 만들고, 유전자 선택 신경 회로의 빛 자극으로 ''꼬마선충''의 운동 패턴을 유발, 온전한 동물에서 신경 활동 제어에 채널로돕신-2를 처음 사용했다(2005년 12월 발표).[36] 쥐의 경우, 광유전학 도구의 제어된 발현은 1990년대 조 Z. 치엔이 개발한 세포 유형 특이적 Cre/loxP 방식을 사용하여 특정 뇌 영역과 세포 유형을 활성화/억제함으로써 ''생체 내''에서 달성된다.[37][38]
2007년, 보이든과 다이서로스 연구실(게오르그 나겔 그룹과 함께)은 신경 세포 활동의 광유전학적 억제를 동시에 보고했다.[39][40]
2007년, 게오르그 나겔과 헤게만 그룹은 cAMP의 광유전학적 조작을 시작했다.[41] 2014년, 아벨라 등은 곰팡이에서 최초의 로돕신-구아닐릴 사이클라제 유전자를 보고했다. 2015년, 샤이프 등과 가오 등은 로돕신-구아닐릴 사이클라제 유전자의 활성을 특징지었다. 시치앙 가오 등과 게오르그 나겔, 알렉산더 고트샬크는 이를 최초의 8 TM 로돕신으로 확인했다.[42]
광유전학이라는 용어는 2006년에 처음 사용되었다.
광활성화 이온 채널인 채널로돕신2 또는 할로로돕신을 특정 뉴런에 유전자 공학으로 발현시킨 후, 특정 파장의 빛을 조사하여 표적 뉴런을 흥분/억제시킬 수 있다.
광유전학 대상은 저분자량 G 단백질 등으로 확대되고 있다. 2009년 클라우스 한 등은 phototropin을 사용한 PA (photoactivatable)-Rac을 보고했다. Neuroscience 2009년에는 PA-RhoA, PA-Cdc42 등도 만들어지고 있음이 보고되었다.
Neuroscience 2009년, 스탠퍼드 대학교 칼 다이서로스는 채널로돕신2에 GPCR을 융합시킨 광유전학 도구를 발표했다. 빛 자극으로 cAMP, IP3, DAG 등 세컨드 메신저 생성을 국소 제어할 수 있다.
광유전학 연구 방법은 네이처 메서드에 의해 "올해의 방법 2010"으로 선정되었다.[168]
신경과학 분야에서, 빛으로 막 전위를 측정하는 막 전위 이미징과 조합, 기존 전기 생리학적 방법에 대신하는 "빛으로 신경의 전기적 특성을 해명하는 도구"인 광학적 전기 생리학으로도 사용된다.
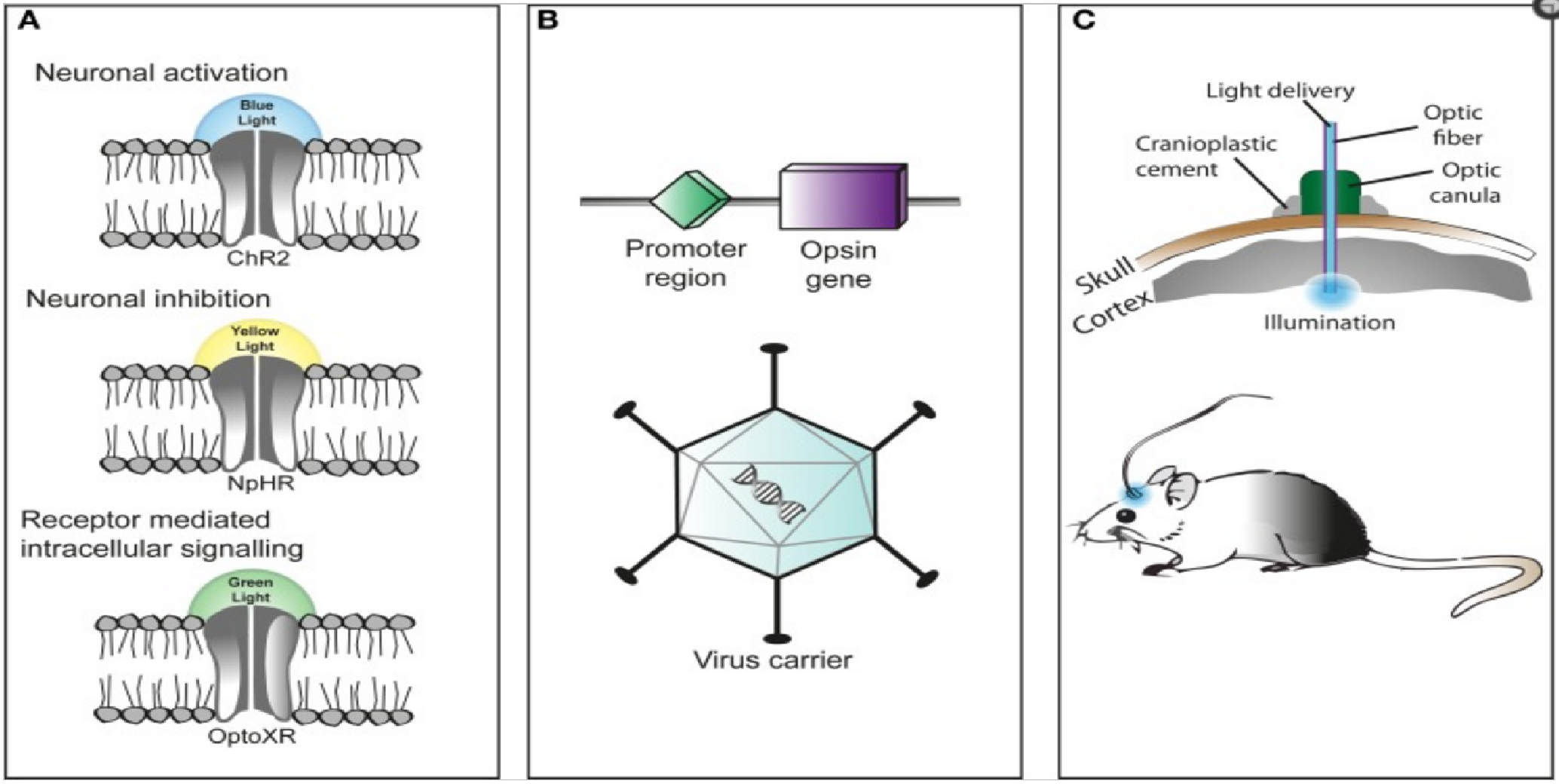
광유전학은 빛에 반응하는 단백질인 옵신, 유전자 전달 기술, 광학 자극 기술, 이 세 가지 주요 요소로 구성된다.
옵신: 광유전학의 핵심 요소로, 빛을 감지하여 세포 내 신호 전달을 유도하는 단백질이다. 채널로돕신은 해조류에서 추출된 단백질로, 빛을 받아 세포를 활성화시키는 역할을 한다. 예를 들어, 채널로돕신을 특정 뉴런에 발현시키면, 빛을 통해 해당 뉴런만 선택적으로 활성화시킬 수 있다.[45]
광유전학적 액추에이터 발현: 광유전학은 특정 뉴런 집단에 광 감응성 프로브를 전달하기 위해 세포 특이적 프로모터나 맞춤형 조건부 활성 바이러스와 같은 유전자 표적화 전략을 사용한다. 무척추동물의 경우, all-trans-retinal (ATR)을 식품에 보충하기도 한다. 미생물 옵신의 주요 장점은 척추 동물에서 외인성 보조 인자를 추가하지 않고도 완전히 기능한다는 것이다.[61]
광 응용 하드웨어: 뇌 깊숙한 곳에서도 특정 세포를 제어할 수 있도록 하는 하드웨어(통합된 광섬유 및 고체 광원 등)가 필요하다. 광섬유 결합 다이오드 기술이 가장 일반적으로 사용되지만,[59][60][61] 쥐 두개골에 이식된 투명하게 수정된 지르코니아로 만들어진 "창"을 새기는 방법도 고안되었다.[62] 대뇌 피질과 같은 뇌 표면 부위를 자극하기 위해 광섬유 또는 LED를 동물의 두개골에 직접 장착할 수 있다.[63] 더 깊은 뇌 부위에 빛을 전달하기 위해 이식된 광섬유가 사용된다.[63] 자유롭게 행동하는 유기체에서 복잡한 행동을 방해받지 않고 연구하기 위해 머리에 장착된 LED에 무선으로 전력을 공급하는 무선 기술도 개발되었다.[64]
광유전학 기술은 실험자의 필요에 따라 유연하게 적용할 수 있다. 양이온 선택 채널로돕신(예: ChR2)은 뉴런을 흥분시키는 데 사용되며, 음이온 전달 채널로돕신(예: GtACR2)은 뉴런 활동을 억제한다. 이러한 도구를 단일 구조체(예: BiPOLES)로 결합하면 조명의 파장에 따라 억제와 흥분을 모두 할 수 있다.[66]
광유전학은 밀리초 단위의 시간적 정밀도를 제공하여, 특정 활동 전위 패턴의 인과적 역할을 정의된 뉴런에서 조사할 수 있도록 한다.[45]
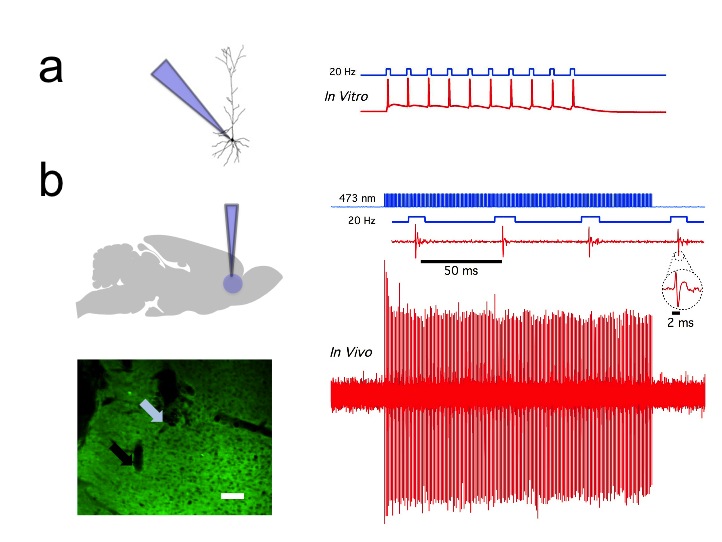
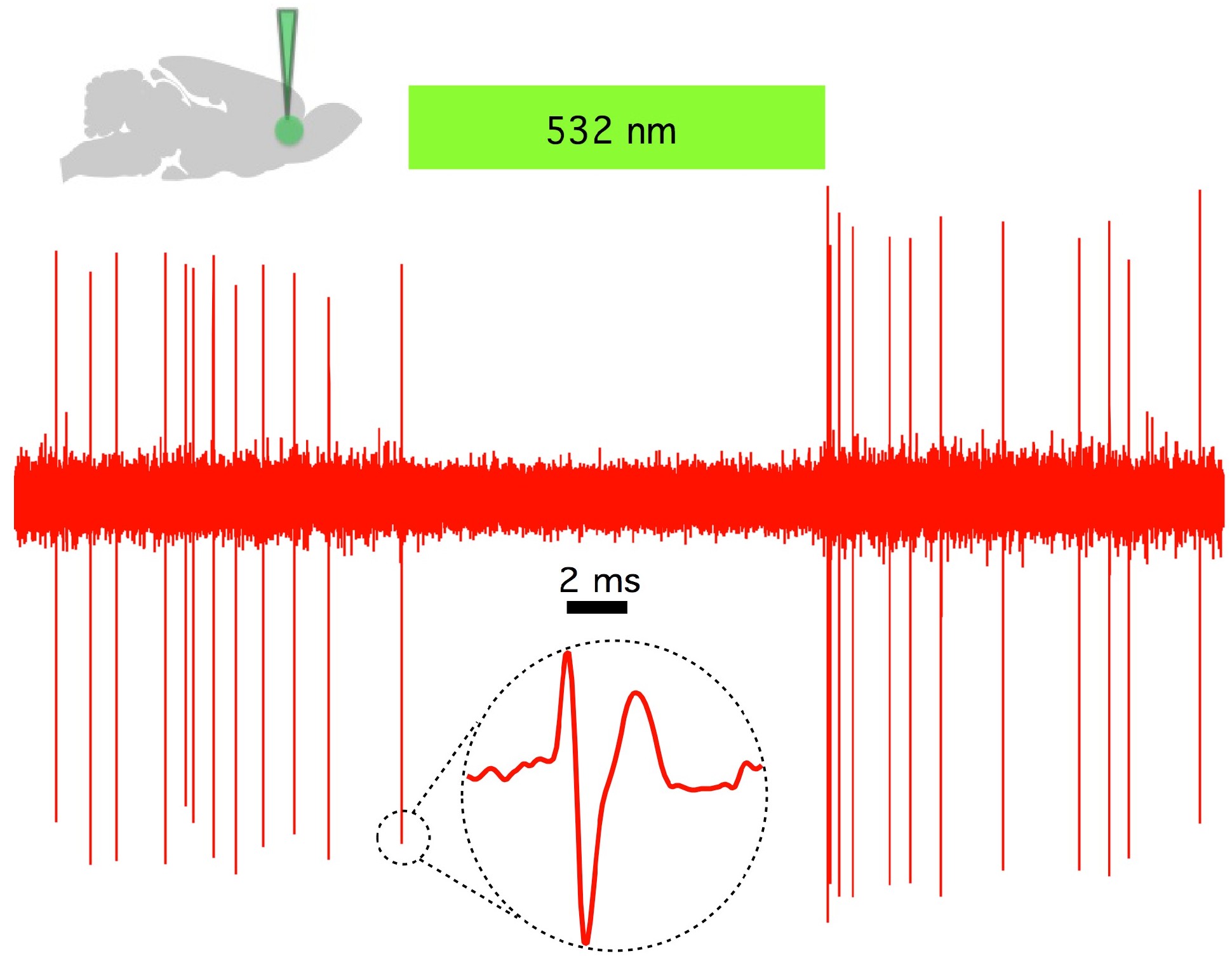


광유전학 기술은 아직 해결해야 할 과제들을 안고 있다. 자유롭게 행동하는 동물의 뇌 심부 특정 유형의 세포를 제어하기 위한 하드웨어(통합된 광섬유나 고체 광원 등)가 필요하다. 2007년에 도입된[169] 광섬유 결합 다이오드 기술이 일반적으로 사용되지만, 매립 전극 없이 광파가 더 깊이 침투하여 개별 뉴런을 자극 또는 억제할 수 있도록 마우스 두개골에 투명한 지르코니아 "창"을 새기는 방법이 개발되었다.[170]
광유전학은 신경과학, 심장학, 세포생물학 등 다양한 분야에서 활용될 가능성이 높은 기술이다.
광유전학은 특정 세포 유형이 생체 조직, 특히 신경 회로의 기능에 어떻게 기여하는지 밝히는 데 중요한 역할을 한다. 예를 들어, 파킨슨병[82][83], 자폐증, 조현병, 약물 남용, 불안, 우울증[50][84][85][86]과 같은 신경 및 정신 질환 연구에 새로운 통찰력을 제공하며, 빛을 이용한 회복 치료법 개발에도 기여하고 있다. 맹인을 위한 실험적 치료법으로는 특수 고글의 빛 패턴으로 자극받는 신경절 세포에 채널 로돕신을 발현시키는 방법이 연구되고 있다.[87][88]
광유전학은 현재 신경 회로, 질병, 행동, 세포 생물학 연구 등에서 활용되고 있다.


광유전학 기술이 뇌 연구에 미치는 강력한 영향은 이 분야의 핵심 인물들에게 수많은 상을 수여함으로써 인정받았다.
2010년, 게오르크 나겔, 페터 헤게만, 에른스트 밤베르크는 와일리 생명 의학상(Wiley Prize in Biomedical Sciences)과 칼 하인츠 베크루츠 상(Karl Heinz Beckurts Prize)을 수상했다.[154][155] 같은 해, 칼 다이서로스는 "행동의 근본이 되는 신경 네트워크의 기능을 연구하기 위한 광유전학적 방법 개발에 대한 선구적인 연구"로 제1회 HFSP 나카소네상(HFSP Nakasone Award)을 수상했다.[156]
2012년, 밤베르크, 다이서로스, 헤게만, 게오르크 나겔은 막스 플랑크 협회(Max Planck Society)로부터 췰히 상(Zülch Prize)을 수상했으며,[157] 미젠뵈크는 "신경 활동을 조작하고 동물의 행동을 제어하는 광유전학적 접근법을 개척한 것"으로 Baillet Latour 건강 상(Baillet Latour Health Prize)을 수상했다.[158]
2013년, 게오르크 나겔과 헤게만은 루이-장테 의학상(Louis-Jeantet Prize for Medicine)을 수상했다.[159] 같은 해, 밤베르크, 보이든, 다이서로스, 헤게만, 미젠뵈크, 게오르크 나겔은 "광유전학의 발명 및 개선"으로 뇌상(The Brain Prize)을 공동 수상했다.[160][161]
2017년, 다이서로스는 "광유전학과 하이드로젤 조직 화학에서의 발견, 그리고 우울증의 신경 회로 기반에 대한 연구"로 엘제 크뢰너-프레제니우스 재단(Else Kröner-Fresenius Foundation) 연구상을 수상했다.[162]
2018년, 이나모리 재단(Inamori Foundation)은 다이서로스에게 "광유전학을 선도"하고 "시스템 신경과학 연구에 혁명을 일으킨" 공로로 교토상(Kyoto Prize)을 수여했다.[163]
2019년, 밤베르크, 보이든, 다이서로스, 헤게만, 미젠뵈크, 게오르크 나겔은 "광유전학의 발명과 개선에 관련된 특별한 공헌"을 인정받아 미국 예술 과학 아카데미(American Academy of Arts and Sciences)로부터 럼포드상(Rumford Prize)을 수상했다.[164]
2020년, 다이서로스는 광유전학과 하이드로젤 조직 화학을 개발한 공로로 네덜란드 왕립 예술 과학 아카데미(Royal Netherlands Academy of Arts and Sciences)로부터 하이네켄상(Heineken Prizes) 의학상을 수상했다.[165] 같은 해, 미젠뵈크, 헤게만, 게오르크 나겔은 쇼상(Shaw Prize) 생명 과학 및 의학 부문을 공동 수상했다.[166]
2021년, 헤게만, 다이서로스, 디터 오스터헬트는 앨버트 래스커 기초 의학 연구상(Albert Lasker Award for Basic Medical Research)을 수상했다.
[1]
논문
A light-switchable gene promoter system
2002-10-01
[2]
논문
Flow of cortical activity underlying a tactile decision in mice
2014-01-01
[3]
논문
Dopaminergic and Prefrontal Basis of Learning from Sensory Confidence and Reward Value
2020-02-01
[4]
논문
Optogenetic stimulation of a hippocampal engram activates fear memory recall
2012-03-01
[5]
논문
Optogenetic Activation of the fruitless-Labeled Circuitry in Drosophila subobscura Males Induces Mating Motor Acts
2017-11-01
[6]
논문
Optogenetic strategies to dissect the neural circuits that underlie reward and addiction
2012-11-01
[7]
논문
Closed-loop optogenetic activation of peripheral or central neurons modulates feeding in freely moving Drosophila
2019-07-19
[8]
논문
Distributed control of motor circuits for backward walking in Drosophila
2020-12-02
[9]
논문
Optogenetic approaches for functional mouse brain mapping
2013
[10]
논문
Light Up the Brain: The Application of Optogenetics in Cell-Type Specific Dissection of Mouse Brain Circuits
2020
[11]
논문
Building a functional connectome of the Drosophila central complex
2018-08-20
[12]
논문
Functional architecture of neural circuits for leg proprioception in Drosophila
2021-12-06
[13]
논문
Optogenetics
2011-01-01
[14]
논문
Insights of the decade. Stepping away from the trees for a look at the forest. Introduction
2010-12-01
[15]
웹사이트
Method of the Year 2010: Optogenetics
https://www.youtube.[...]
2010-12-17
[16]
웹사이트
Optogenetics: Controlling the Brain with Light
http://www.scientifi[...]
Springer Nature America, Inc.
2010-10-20
[17]
논문
The impact of molecular biology on neuroscience
1999-12-01
[18]
논문
Photoactive mitochondria: in vivo transfer of a light-driven proton pump into the inner mitochondrial membrane of Schizosaccharomyces pombe
1994-09-01
[19]
논문
Selective photostimulation of genetically chARGed neurons
2002-01-01
[20]
논문
Photochemical gating of heterologous ion channels: remote control over genetically designated populations of neurons
2003-02-01
[21]
논문
Light-activated ion channels for remote control of neuronal firing
2004-12-01
[22]
논문
Allosteric control of an ionotropic glutamate receptor with an optical switch
2006-01-01
[23]
논문
Genetic control of neuronal activity in mice conditionally expressing TRPV1
2008-04-01
[24]
논문
Transient activation of specific neurons in mice by selective expression of the capsaicin receptor
2012-03-01
[25]
논문
Synaptic modifications in the medial prefrontal cortex in susceptibility and resilience to stress
2014-05-01
[26]
논문
Channelrhodopsin-2, a directly light-gated cation-selective membrane channel
2003-11-01
[27]
논문
Rhodopsin-regulated calcium currents in Chlamydomonas
1991-06-06
[28]
논문
Channelrhodopsin-1: a light-gated proton channel in green algae
2002-06-01
[29]
논문
Optogenetics: 10 years of microbial opsins in neuroscience
2015-09-01
[30]
논문
Millisecond-timescale, genetically targeted optical control of neural activity
2005-09-01
[31]
웹사이트
He may be the rightful inventor of neuroscience's biggest breakthrough in decades. But you've never heard of him
https://www.statnews[...]
2016-09-01
[32]
논문
Optogenetics: 10 years of microbial opsins in neuroscience
2015-08-26
[33]
논문
Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration
2006-04-01
[34]
논문
Remote control of behavior through genetically targeted photostimulation of neurons
2005-04-01
[35]
논문
Fast noninvasive activation and inhibition of neural and network activity by vertebrate rhodopsin and green algae channelrhodopsin
2005-12-01
[36]
논문
Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses
2005-12-00
[37]
논문
Subregion- and cell type-restricted gene knockout in mouse brain
1996-12-00
[38]
논문
Cre-Lox Neurogenetics: 20 Years of Versatile Applications in Brain Research and Counting...
[39]
논문
Multiple-color optical activation, silencing, and desynchronization of neural activity, with single-spike temporal resolution
Public Library of Science
2007-03-00
[40]
논문
Multimodal fast optical interrogation of neural circuitry
2007-04-00
[41]
논문
Fast manipulation of cellular cAMP level by light in vivo
http://edoc.hu-berli[...]
2007-01-00
[42]
논문
Optogenetic manipulation of cGMP in cells and animals by the tightly light-regulated guanylyl-cyclase opsin CyclOp
2015-09-00
[43]
논문
Microbial light-activatable proton pumps as neuronal inhibitors to functionally dissect neuronal networks in C. elegans
[44]
논문
A cholinergic-regulated circuit coordinates the maintenance and bi-stable states of a sensory-motor behavior during Caenorhabditis elegans male copulation
2011-03-00
[45]
뉴스
Optogenetics: Controlling the Brain with Light [Extended Version]
https://www.scientif[...]
2016-11-28
[46]
논문
Synthetic Light-Activated Ion Channels for Optogenetic Activation and Inhibition
2018-00-00
[47]
논문
Potassium channel-based two component optogenetic tool for silencing of excitable cells
2018-11-00
[48]
논문
Improved expression of halorhodopsin for light-induced silencing of neuronal activity
2008-08-00
[49]
논문
eNpHR: a Natronomonas halorhodopsin enhanced for optogenetic applications
2008-08-00
[50]
논문
Cholinergic interneurons control local circuit activity and cocaine conditioning
2010-12-00
[51]
논문
Light-driven activation of beta 2-adrenergic receptor signaling by a chimeric rhodopsin containing the beta 2-adrenergic receptor cytoplasmic loops
2005-02-00
[52]
논문
Temporally precise in vivo control of intracellular signalling
2009-04-00
[53]
논문
Spatiotemporal control of cell signalling using a light-switchable protein interaction
2009-10-00
[54]
논문
A genetically encoded photoactivatable Rac controls the motility of living cells
2009-09-00
[55]
논문
Induction of protein-protein interactions in live cells using light
2009-10-00
[56]
논문
Light modulation of cellular cAMP by a small bacterial photoactivated adenylyl cyclase, bPAC, of the soil bacterium Beggiatoa
2011-01-00
[57]
논문
Natural and engineered photoactivated nucleotidyl cyclases for optogenetic applications
2010-12-00
[58]
논문
Communication in Neural Circuits: Tools, Opportunities, and Challenges
2016-03-00
[59]
논문
An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology
2007-09-00
[60]
논문
Neural substrates of awakening probed with optogenetic control of hypocretin neurons
2007-11-00
[61]
논문
Targeting and readout strategies for fast optical neural control in vitro and in vivo
2007-12-00
[62]
논문
Transparent nanocrystalline yttria-stabilized-zirconia calvarium prosthesis
https://escholarship[...]
2013-11-00
[63]
논문
Fiber photometry in striatum reflects primarily nonsomatic changes in calcium
2022-08-30
[64]
논문
A wirelessly powered and controlled device for optical neural control of freely-behaving animals
2011-08-00
[65]
논문
Optogenetics as a neuromodulation tool in cognitive neuroscience
2013-01-01
[66]
논문
BiPOLES is an optogenetic tool developed for bidirectional dual-color control of neurons
2021-07-26
[67]
논문
Optogenetic interrogation of neural circuits: technology for probing mammalian brain structures
2010-03-00
[68]
서적
Optogenetics: Tools for Controlling and Monitoring Neuronal Activity
2012-09-05
[69]
논문
Optical neural interfaces
2014-07-00
[70]
논문
Making Sense of Optogenetics
2015-07-00
[71]
웹사이트
The Evolution in Freely-Behaving Imaging and Optogenetics Technology
https://www.mightexb[...]
Mightex
2021-06-03
[72]
논문
Deep brain optical measurements of cell type-specific neural activity in behaving mice
2016-04-00
[73]
논문
Deep brain optical measurements of cell type-specific neural activity in behaving mice
2014-06-00
[74]
웹사이트
Current Challenges in Optogenetics
https://www.sfn.org/[...]
2013-00-00
[75]
논문
Optogenetic Stimulation Shifts the Excitability of Cerebral Cortex from Type I to Type II: Oscillation Onset and Wave Propagation
2017-01-00
[76]
논문
Optical induction of synaptic plasticity using a light-sensitive channel
http://www.nature.co[...]
2007-00-00
[77]
논문
Mapping the connectome: multi-level analysis of brain connectivity
2012-05-01
[78]
논문
Nonlinear response characteristics of neural networks and single neurons undergoing optogenetic excitation
2020-09-00
[79]
웹사이트
PyRhO: a virtual optogenetics laboratory
https://github.com/P[...]
[80]
웹사이트
Simulation tool for neural networks and single neurons with light-sensitive channels
https://github.com/j[...]
[81]
논문
Spread of activation and interaction between channels with multi-channel optogenetic stimulation in the mouse cochlea
2023-12-00
[82]
논문
Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry
2010-07-00
[83]
논문
Optical deconstruction of parkinsonian neural circuitry
2009-04-00
[84]
논문
Driving fast-spiking cells induces gamma rhythm and controls sensory responses
2009-06-00
[85]
논문
Parvalbumin neurons and gamma rhythms enhance cortical circuit performance
2009-06-00
[86]
논문
Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning
2009-05-00
[87]
뉴스
Scientists Partially Restored a Blind Man's Sight With New Gene Therapy
https://www.nytimes.[...]
2021-05-24
[88]
논문
Partial recovery of visual function in a blind patient after optogenetic therapy
2021-07-00
[89]
논문
Genetic dissection of an amygdala microcircuit that gates conditioned fear
2010-11-00
[90]
논문
Optical activation of lateral amygdala pyramidal cells instructs associative fear learning
2010-07-00
[91]
논문
Thy1-expressing neurons in the basolateral amygdala may mediate fear inhibition
2013-06-00
[92]
논문
Towards new approaches to disorders of fear and anxiety
2013-06-00
[93]
논문
4-Hz oscillations synchronize prefrontal-amygdala circuits during fear behavior
2016-04-00
[94]
논문
Precise olfactory responses tile the sniff cycle
2011-07-00
[95]
논문
Differential Muscarinic Modulation in the Olfactory Bulb
2015-07-00
[96]
논문
Odor representations in the olfactory bulb evolve after the first breath and persist as an odor afterimage
2013-08-00
[97]
논문
Glutamatergic signaling by mesolimbic dopamine neurons in the nucleus accumbens
2010-05-00
[98]
논문
Optogenetic control of genetically-targeted pyramidal neuron activity in prefrontal cortex
http://precedings.na[...]
2012-04-02
[99]
논문
An Optogenetic Kindling Model of Neocortical Epilepsy
2019-03-00
[100]
논문
Chronic loss of inhibition in piriform cortex following brief, daily optogenetic stimulation
2021-04-00
[101]
논문
Light-induced termination of spiral wave arrhythmias by optogenetic engineering of atrial cardiomyocytes
2014-10-00
[102]
논문
Optogenetics for in vivo cardiac pacing and resynchronization therapies
2015-07-00
[103]
논문
Optogenetic termination of ventricular arrhythmias in the whole heart: towards biological cardiac rhythm management
2017-07-00
[104]
논문
Optogenetic defibrillation terminates ventricular arrhythmia in mouse hearts and human simulations
2016-10-00
[105]
논문
Optogenetics design of mechanistically-based stimulation patterns for cardiac defibrillation
2016-10-00
[106]
논문
Optogenetic stimulation of the auditory pathway
2014-03-01
[107]
논문
Ultrafast optogenetic stimulation of the auditory pathway by targeting-optimized Chronos
2018-12-01
[108]
논문
High frequency neural spiking and auditory signaling by ultrafast red-shifted optogenetics
2018-05-01
[109]
웹사이트
Engineering long-wavelength light-driven ion channels to hear the light. Atlas of Science
https://atlasofscien[...]
2019-11-07
[110]
논문
Optogenetic stimulation of the auditory pathway for research and future prosthetics
2015-10-01
[111]
논문
ReaChR: a red-shifted variant of channelrhodopsin enables deep transcranial optogenetic excitation
2013-10-01
[112]
논문
Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation
2016-02-01
[113]
논문
Independent optical excitation of distinct neural populations
2014-03-01
[114]
논문
Bi-stable neural state switches
2009-02-01
[115]
논문
NEUROSCIENCE. Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics
2015-08-01
[116]
논문
Optogenetic Neuronal Silencing in Drosophila during Visual Processing
2017-10-01
[117]
논문
Optogenetics in a transparent animal: circuit function in the larval zebrafish
2013-02-01
[118]
논문
Optogenetic dissection of descending behavioral control in Drosophila
2018-06-26
[119]
논문
Spatiotemporally precise optogenetic activation of sensory neurons in freely walking Drosophila
2020-04-22
[120]
논문
Two Brain Pathways Initiate Distinct Forward Walking Programs in Drosophila
2020-11-11
[121]
논문
Precise Control of Movement Kinematics by Optogenetic Inhibition of Purkinje Cell Activity
https://www.jneurosc[...]
2014-02-05
[122]
논문
Open Source Tools for Temporally Controlled Rodent Behavior Suitable for Electrophysiology and Optogenetic Manipulations
2018-05-01
[123]
논문
A user's guide to channelrhodopsin variants: features, limitations and future developments
2011-01-01
[124]
논문
Closed-Loop and Activity-Guided Optogenetic Control
2015-04-08
[125]
논문
Closed-loop optogenetic intervention in mice
2013-01-01
[126]
논문
Optogenetically Blocking Sharp Wave Ripple Events in Sleep Does Not Interfere with the Formation of Stable Spatial Representation in the CA1 Area of the Hippocampus
2016-11-19
[127]
논문
Optogenetic control of cellular forces and mechanotransduction
2017-02-01
[128]
논문
Applications of Optobiology in Intact Cells and Multicellular Organisms
2017-10-01
[129]
논문
The development and application of optogenetics
2011-01-01
[130]
Youtube
Method of the Year 2010: Optogenetics
https://www.youtube.[...]
2010-12-17
[131]
웹사이트
optogenetics - Search Results
https://pubmed.ncbi.[...]
2020-02-29
[132]
논문
Lights, cytoskeleton, action: Optogenetic control of cell dynamics
Elsevier Ltd.
2020-10-01
[133]
논문
Optical control of mammalian endogenous transcription and epigenetic states
2013-08-01
[134]
논문
Genetically encoded photoswitching of actin assembly through the Cdc42-WASP-Arp2/3 complex pathway
2008-09-01
[135]
논문
Light-based feedback for controlling intracellular signaling dynamics
2011-09-01
[136]
논문
TULIPs: tunable, light-controlled interacting protein tags for cell biology
2012-03-01
[137]
논문
Optogenetic control of phosphoinositide metabolism
2012-08-01
[138]
논문
Optogenetic protein clustering and signaling activation in mammalian cells
2013-03-01
[139]
논문
Designing photoswitchable peptides using the AsLOV2 domain
2012-04-01
[140]
논문
A genetically encoded photoactivatable Rac controls the motility of living cells
2009-09-01
[141]
저널
Engineering a light-activated caspase-3 for precise ablation of neurons in vivo
2017-09-00
[142]
저널
Engineering extrinsic disorder to control protein activity in living cells
2016-12-00
[143]
저널
Engineering an improved light-induced dimer (iLID) for controlling the localization and activity of signaling proteins
2015-01-00
[144]
저널
LOVTRAP: an optogenetic system for photoinduced protein dissociation
2016-09-00
[145]
저널
Local control of intracellular microtubule dynamics by EB1 photodissociation
Nature Research.
2018-03-00
[146]
저널
Optical control of protein activity by fluorescent protein domains
2012-11-00
[147]
저널
Illuminating cell signalling with optogenetic tools
2014-08-00
[148]
저널
Encoding and decoding cellular information through signaling dynamics
2013-02-00
[149]
저널
Growth factor-induced MAPK network topology shapes Erk response determining PC-12 cell fate
2007-03-00
[150]
저널
Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module
2013-12-00
[151]
저널
Brownian Optogenetic-Noise-Photostimulation on the Brain Amplifies Somatosensory-Evoked Field Potentials
2017-00-00
[152]
저널
Optogenetic noise-photostimulation on the brain increases somatosensory spike firing responses
2018-01-00
[153]
저널
Noisy Light Augments the Na+ Current in Somatosensory Pyramidal Neurons of Optogenetic Transgenic Mice
2020-00-00
[154]
웹사이트
Ninth Annual Wiley Prize in Biomedical Sciences Awarded to Dr. Peter Hegemann, Dr. Georg Nagel, and Dr. Ernst Bamberg
http://eu.wiley.com/[...]
[155]
웹사이트
Karl Heinz Beckurts-Preis 2010
https://www.beckurts[...]
[156]
웹사이트
HFSP Nakasone Award 2010
https://www.hfsp.org[...]
[157]
웹사이트
International Prize for Translational Neuroscience of the Gertrud Reemtsma Foundation (K.J. Zülch Prize until 2019)
https://www.mpg.de/p[...]
[158]
웹사이트
InBev-Baillet Latour International Health Prize
https://www.frs-fnrs[...]
[159]
웹사이트
Louis-Jeantet Prize
https://www.jeantet.[...]
[160]
웹사이트
The Brain Prize 2013
http://www.thebrainp[...]
2013-10-03
[161]
저널
The Brain Prize 2013: the optogenetics revolution
2013-10-00
[162]
웹사이트
Else Kröner Fresenius Prize for Medical Research 2017
http://ekfs.de/en/sc[...]
[163]
웹사이트
2018 Kyoto Prize Laureate Karl Deisseroth
https://kyotoprize.o[...]
[164]
웹사이트
Rumford Prize Awarded for the Invention and Refinement of Optogenetics
https://www.amacad.o[...]
2019-01-30
[165]
웹사이트
2020 Heineken Prize Laureate Karl Deisseroth
https://www.heineken[...]
[166]
웹사이트
2020 Shaw Prize Laureates Miesenböck, Hegemann and Georg Nagel
https://www.shawpriz[...]
[167]
서적
依存症の科学 いちばん身近なこころの病
化学同人
[168]
웹사이트
Method of the Year 2010
http://www.nature.co[...]
[169]
저널
An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology
https://iopscience.i[...]
2007-09-01
[170]
저널
Transparent nanocrystalline yttria-stabilized-zirconia calvarium prosthesis
https://linkinghub.e[...]
2013-11-00
[171]
저널
Sub-millisecond Control of Neuronal Firing by Organic Light-Emitting Diodes
https://www.frontier[...]
2019-10-22
[172]
저널
Next-Generation Optical Technologies for Illuminating Genetically Targeted Brain Circuits
[173]
저널
2010-00-00
[174]
저널
Single-molecule fluorimetry and gating currents inspire an improved optical voltage indicator
2015-00-00
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com