로돕신
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
로돕신은 1876년 프란츠 크리스티안 볼에 의해 발견된, 시각에 관여하는 단백질이다. G 단백질 연결 수용체의 일종으로, 망막의 간상체에 존재하며 빛을 감지하는 역할을 한다. 로돕신은 옵신이라는 단백질과 레티날이라는 비타민 A 유도체로 구성되며, 빛을 받으면 레티날이 구조를 바꾸면서 신호 전달을 시작한다. 로돕신 유전자 돌연변이는 망막 색소 변성과 같은 망막 질환을 유발할 수 있다.
더 읽어볼만한 페이지
- 생물 색소 - 피토크롬
피토크롬은 식물의 주요 광수용체로서 적색광과 원적색광을 흡수하여 광가역성을 통해 식물의 생리적 변화를 조절하는 색소 단백질이다. - 생물 색소 - 빌리루빈
빌리루빈은 적혈구 파괴로 생성되는 헴의 대사 산물이며, 간에서 대사 과정을 거쳐 담즙으로 배설되고, 혈액 검사를 통해 농도를 측정하여 질병을 진단하는 데 사용된다. - 감각수용기 - 피토크롬
피토크롬은 식물의 주요 광수용체로서 적색광과 원적색광을 흡수하여 광가역성을 통해 식물의 생리적 변화를 조절하는 색소 단백질이다. - 감각수용기 - 기계수용기
기계수용기는 압력, 진동, 신장과 같은 기계적 자극을 감지하여 전기적 신호로 바꾸는 감각 수용기로, 세포막 변형을 감지하는 이온 채널 형태이며, 촉각, 고유수용성 감각, 청각, 평형 감각 등 다양한 감각 기능에 관여한다. - G 단백질 연결 수용체 - P2Y12
P2Y12 길항제는 P2Y12 수용체에 결합해 혈소판 활성화를 억제하는 항혈소판제로, 클로피도그렐, 프라수그렐, 티카그렐러, 캉그렐러 등이 있으며, 아스피린과 병용하는 이중 항혈소판 치료는 급성 관상 동맥 증후군의 주요 치료법으로 쓰인다. - G 단백질 연결 수용체 - GABA 수용체
GABA 수용체는 신경전달물질 GABA에 의해 활성화되며, 이온 채널형 GABA 수용체와 대사 조절형 GABA 수용체로 구분되고, 신경 흥분 억제, 신경계 발달 등에 중요한 역할을 담당하며, 다양한 질환 및 약물과 관련된다.
| 로돕신 | |
|---|---|
| 기본 정보 | |
 | |
| 다른 이름 | 시각 자색 |
| 기능 | 광수용체 |
| 유전자 | 사람의 경우 RHO |
| 식별 | |
| 심볼 | RHO |
| 유전자좌(Locus) | 염색체 3q22.1 |
| 단백질 데이터 뱅크(Protein Data Bank) | 1U19, 1GZM |
| 앙트르 | '6010' |
| OMIM | '180380' |
| UniProt | P08100 |
| RefSeq (유전자) | NM_000539 |
| RefSeq (단백질) | NP_000530 |
| 위치 (UCSC) | uc003fxx.5 |
| 특징 | |
| 분자량 | 34828.65 Da |
| 아미노산 개수 | 348 |
| 기타 | |
| 상호 작용 | 트랜스듀신 어레스틴 |
| 관련 질병 | 망막 색소 변성증 선천성 야맹증 |
| 외부 링크 | |
| 미쉬켄의 유전자(GeneCards) | RHO |
| 인간 유전자 지도(HGNC) | 10025 |
| 국립 생물 정보 센터(NCBI) | '6010' |
2. 명칭
로돕신은 1876년 프란츠 크리스티안 볼에 의해 발견되었다.[4][5][6] 로돕신이라는 이름은 분홍빛 색깔을 띠기 때문에 "장미"를 뜻하는 고대 그리스어 ῥόδον|로돈grc과 "시각"을 뜻하는 ὄψις|옵시스grc에서 유래되었다.[7] 이 용어는 1878년 독일 생리학자 빌헬름 프리드리히 퀠네 (1837–1900)에 의해 만들어졌다.[8][9]
로돕신은 간상 세포의 외부 분절 원반에서 발견되는 단백질로, 어두운 빛에서의 암순응 시각을 매개한다.[3][13] 로돕신은 녹청색광(~500 nm)을 가장 강하게 흡수하여[14][15] 붉은 자주색으로 보이기 때문에 "시각 자색"이라고도 불린다.
조지 왈드는 로돕신이 레티날과 아포단백질로 구성된 홀로단백질이라는 것을 발견하고 이를 옵신이라고 불렀는데, 오늘날에는 아포-로돕신으로 더욱 좁게 묘사된다.[10] 오늘날 옵신이라는 용어는 망막에 결합하여 빛에 민감한 광수용체가 되는 G 단백질 연결 수용체의 종류, 즉 모든 관련된 단백질을 광범위하게 지칭한다.[16][37][11] 왈드와 그의 동료들이 나중에 닭의 망막에서 요오돕신을 분리하여 최초로 알려진 원추 옵신을 발견했을 때, 그들은 아포-요오돕신을 ''포토옵신''(명소시와의 관련성 때문에), 아포-로돕신을 ''스코토옵신''(암소시에 사용되기 때문에)이라고 불렀다.[12]
3. 구조
로돕신은 다른 옵신과 마찬가지로 G 단백질 연결 수용체(GPCR)이다.[18][19] GPCR은 세포막의 지질 이중층에 내장되어 있으며 리간드에 대한 결합 포켓을 형성하는 7개의 막 횡단 도메인을 가지고 있는 화학 수용체이다.[20][21] 로돕신의 리간드는 비타민 A 기반 발색단인 11-''시스''-레티날이며,[22][23][24][25][26] 이는 세포막에 수평으로 놓여 있고,[27] 공유 결합적으로 일곱 번째 막 횡단 도메인의 라이신 잔기(lys296)에 쉬프 염기를 통해 결합한다.[29][27][30][31]
11-''시스''-레티날은 결합 포켓을 막을 뿐 로돕신을 활성화시키지 않는다. 11-''시스''-레티날이 광자의 빛을 흡수하여 모든 ''트랜스''-레티날로 이성질체화될 때만 활성화되며,[32][33] 이는 수용체를 활성화시키는 형태이다.[34][35] 로돕신에서 형태적 변화(탈색)를 일으켜[34] 광변환 폭포를 활성화시킨다.[36] 따라서 화학 수용체는 광수용체 단백질로 변환된다.[37]
레티날 결합 라이신은 거의 모든 옵신에서 보존되며, 몇몇 옵신만이 진화 과정에서 이를 잃었다.[37] 라이신이 없는 옵신은 로돕신을 포함하여 빛에 민감하지 않다.[38][39][40] 11-''시스''-레티날은 역 작용제이다.
소의 로돕신은 348개의 아미노산을 가지고 있으며, 레티날 결합 라이신은 Lys296이다. 이는 단백질 1차 구조의 아미노산 서열[46]과 단백질 3차 구조가 처음으로 밝혀진 옵신이다.[27] 그 구조는 로돕신 결정에 대한 X선 결정학을 통해 상세히 연구되었다.[47]
로돕신은 네이티브 막 내에서 높은 밀도로 발견되어 광자를 포착하는 능력을 촉진한다. 막 내에 밀집되어 있어 로돕신이 단백질을 포착할 가능성이 더 높다. 그러나 높은 밀도는 G 단백질 신호 전달과 관련하여 단점도 제공하는데, 수용체인 로돕신으로 채워진 혼잡한 막에서는 확산이 더 어려워지기 때문이다.[52]
4. 기능 및 광변환 과정

로돕신은 광변환에서 필수적인 G 단백질 연결 수용체이며, 망막에서 빛 수용기 세포 형성과 빛 인지 초기 단계를 담당한다. G 단백질 결합 수용체의 일종으로 빛에 민감하여 야간 시력을 가능하게 한다. 빛을 받으면 즉시 탈색되며, 회복에는 사람의 경우 약 30분이 걸린다.
로돕신은 옵신이라는 단백질 부분과 레티날(레틴알데히드)이라는 보조 인자로 구성된다. 옵신은 7개의 나선이 묶여 막을 관통하며, 중심 포켓 내에 레티날이 결합한다. 레티날은 망막에서 비타민 A로부터 만들어지므로, 비타민 A 결핍은 야맹증을 유발한다.
11-시스-레티날 (11-''cis''-retinal)이 빛에 의해 올트랜스레티날 (all-''trans''-retinal)로 이성화되면 옵신은 구조 변화를 일으켜, 회합하고 있는 G 단백질을 활성화시켜 세컨드 메신저 캐스케이드를 일으킨다.
로돕신에 빛이 닿으면 시스형 레티날이 안정적인 트랜스형으로 돌아가고, 트랜스형 레티날 분자는 옵신에 수납되지 못하고 떨어져 나가게 된다. 이 변화가 세포 안으로 전달되어, 화학적으로 증폭되어, 빛이 닿았다는 신호가 되어 시신경에 전달된다. 트랜스형 레티날은 다시 이성질화 효소의 작용으로 시스형으로 구부려져 옵신에 수납된다. 이윽고 레티날은 소모되므로, 부족한 분은 레티놀로부터 산화되어 보충된다.
간상체에 있는 로돕신은 녹청색 빛을 가장 흡수하여 적자색으로 보이며, 이 때문에 어두운 곳에서의 시야는 흑백으로 보인다.
left
포토프신은 로돕신과 유사하지만 아미노산 배열이 약간 다르고 최대 흡수 파장이 다른 원추 세포에서 발견되는 색소로, 색각의 근원이 된다. 사람은 로돕신 외에 황록색(photopsin I), 녹색(photopsin II), 청자색(photopsin III)의 빛을 흡수하는 3가지 포토프신을 갖는다.
고세균 중에는 광합성을 하는 박테리오로돕신이 있는데, 로돕신과 마찬가지로 레티날을 가지며 7개의 막 관통 α 나선을 갖지만 G 단백질과는 결합하지 않는다. 조류는 채널로돕신2라는 광 게이트 이온 채널을 가진 옵신을 가지고 있는 것으로 알려져 있다.
망막에는 레티놀을 레티날로 산화시키기 위한 알코올 탈수소 효소가 풍부하게 존재한다. 따라서 메탄올을 마신 경우에는, 망막에서 포름알데히드가 대량으로 만들어지고, 포름알데히드의 독성으로 시세포가 죽어 실명하게 된다[60]。
4. 1. 광변환
로돕신은 광변환에 필수적인 G 단백질 연결 수용체이다. 로돕신에서 레티날의 알데히드기는 단백질의 라이신 잔기의 아미노기와 양성자화된 쉬프 염기(-NH+=CH-) 형태로 공유 결합되어 있다.[28] 로돕신이 빛을 흡수하면 레티날 보조 인자가 11-시스 형태에서 all-trans 형태로 이성질체화되고, 단백질은 이성질체화된 보조 인자의 변경된 형태를 수용하기 위해 일련의 완화를 겪는다. 이 과정에서 형성된 중간체는 조지 월드의 연구실에서 처음 연구되었다.[53]
광 이성질체화 역학은 이후 시간 분해 IR 분광법 및 UV/Vis 분광법으로 연구되었다. 조사 후 200 펨토초 이내에 '''포토로돕신'''이라는 첫 번째 광생성물이 형성되고, 피코초 이내에 왜곡된 all-trans 결합을 가진 '''바토로돕신'''이라는 두 번째 광생성물이 형성된다. 이 중간체는 저온에서 포획하여 연구할 수 있으며, 초기에는 prelumirhodopsin이라고 불렸다.[54] 후속 중간체인 '''루미로돕신'''과 '''메타로돕신 I'''에서, all-trans 레티날과의 쉬프 염기 연결은 양성자화된 상태로 유지되며, 단백질은 붉은색을 유지한다. 신경 흥분을 시작하는 중요한 변화는 메타로돕신 I이 '''메타로돕신 II'''로 전환되는 것과 관련이 있으며, 이는 쉬프 염기의 탈양성자화와 색상이 빨간색에서 노란색으로 바뀌는 것과 관련이 있다.[55]
11-시스-레티날 (11-''cis''-retinal)이 빛에 의해 올트랜스레티날 (all-''trans''-retinal)로 이성화되면 옵신은 구조 변화를 일으켜, 회합하고 있는 G 단백질을 활성화시켜 세컨드 메신저 캐스케이드를 일으킨다. 로돕신에 빛이 닿으면 시스형 레티날이 안정적인 트랜스형으로 돌아가고, 트랜스형 레티날 분자는 옵신에 수납되지 못하고 떨어져 나가게 된다. 이 변화가 세포 안으로 전달되어 화학적으로 증폭되어, 빛이 닿았다는 신호가 되어 시신경에 전달된다. 트랜스형 레티날은 다시 이성질화 효소의 작용으로 시스형으로 구부려져 옵신에 수납된다.
4. 2. 시각 신호 전달
빛 활성화의 산물인 메타로돕신 II는 시각 광전환 세컨드 메신저 경로를 시작한다. 이는 G 단백질 트랜스듀신(Gt)을 자극하여 α 소단위체를 유리시키는 결과를 낳는다. 이 GTP 결합 소단위체는 차례로 cGMP 포스포다이에스터레이스를 활성화한다. cGMP 포스포다이에스터레이스는 cGMP를 가수분해(분해)하여 국소 농도를 낮추므로 cGMP 의존성 양이온 통로를 더 이상 활성화할 수 없게 된다. 이는 광수용체 세포의 과분극을 유발하여 신경 전달 물질의 방출 속도를 변화시킨다.[56][4]
로돕신이 시신경에 신호를 전달하는 메커니즘은 다음과 같다. β-카로틴이 사슬 중간에서 절단되면, 두 개의 트랜스형 레티놀 (알코올형 비타민 A)이 생성된다. 레티놀은 산화되어 레티날 (알데히드)이 된다. 이 트랜스형 레티날은 시스형 레티날로 변화된 후 옵신에 수납된다. 이 상태가 로돕신이다.
로돕신에 빛이 닿으면 시스형 레티날이 안정적인 트랜스형으로 돌아가고, 트랜스형 레티날 분자는 옵신에 수납되지 못하고 떨어져 나가게 된다. 이 변화가 세포 안으로 전달되어 화학적으로 증폭되고, 빛이 닿았다는 신호가 되어 시신경에 전달된다. 트랜스형 레티날은 다시 이성질화 효소의 작용으로 시스형으로 구부려져 옵신에 수납된다. 이후 레티날은 소모되므로, 부족한 분은 레티놀로부터 산화되어 보충된다.
망막에는 레티놀을 레티날로 산화시키기 위한 알코올 탈수소 효소가 풍부하게 존재한다. 따라서 메탄올을 마시면 망막에서 포름알데히드가 대량으로 만들어지고, 포름알데히드의 독성으로 시세포가 죽어 실명하게 된다.[60]
4. 3. 비활성화 및 재생
메타 II는 로돕신 키나아제와 아레스틴에 의해 트랜스듀신을 활성화시킨 후 빠르게 비활성화된다.[57] 추가적인 광변환이 일어나기 위해서는 로돕신 색소가 재생성되어야 하는데, 이는 올-트랜스-레티날을 11-시스-레티날로 대체하는 것을 의미한다. 이 과정에서 메타 II의 분해는 매우 중요하다. 메타 II가 분해되는 동안, 올-트랜스-레티날과 아포단백질 옵신(아포로돕신)을 결합시키는 시프 염기 결합이 가수분해되어 메타 III이 된다. 막대 외절에서 메타 III은 분리된 올-트랜스-레티날과 옵신으로 분해된다.[57]
메타 II 분해의 두 번째 생성물은 올-트랜스-레티날이 두 번째 결합 부위로 전위된 올-트랜스-레티날 옵신 복합체이다. 메타 II의 분해가 메타 III으로 진행될지, 아니면 올-트랜스-레티날 옵신 복합체로 진행될지는 반응의 pH에 따라 달라지는 것으로 보이며, pH가 높을수록 분해 반응이 메타 III으로 진행되는 경향이 있다.[57]
로돕신에 빛이 닿으면 시스형 레티날이 안정적인 트랜스형으로 돌아가고, 트랜스형 레티날 분자는 옵신에 수납되지 못하고 떨어져 나가게 된다. 이 변화가 세포 안으로 전달되어 화학적으로 증폭되어 빛이 닿았다는 신호가 되어 시신경에 전달된다. 트랜스형 레티날은 다시 이성질화 효소의 작용으로 시스형으로 구부려져 옵신에 수납된다. 레티날은 소모되므로, 부족한 분은 레티놀로부터 산화되어 보충된다.
5. 관련 질환
로돕신 유전자의 돌연변이는 망막 색소 변성 등 다양한 망막 질환의 주요 원인으로 작용한다. 일반적으로 결함이 있는 로돕신은 유비퀴틴과 함께 봉입체에 축적되어 중간 섬유 네트워크를 파괴하고, 세포가 제대로 기능하지 않는 단백질을 분해하는 능력을 손상시켜 광수용체의 세포자멸사를 유발한다.[58] 로돕신의 다른 돌연변이는 주로 구성적 활성화로 인해 X 연관 선천성 야맹증을 유발하며, 이는 돌연변이가 로돕신의 발색단 결합 포켓 주변에서 발생할 때 나타난다.[59] 로돕신과 관련된 몇 가지 다른 병리학적 상태로는 골지체 후 수송 불량, 조절되지 않는 활성화, 막대 외절 불안정 및 아레스틴 결합 등이 있다.[59]
6. 다른 옵신과의 관계
로돕신과 비슷한 옵신에는 포토프신(photopsin) 그룹이 존재한다. 포토프신은 로돕신과 아미노산이 약간 다르며 최대 흡수 파장이 다르다. 이 색소들은 망막의 원추 세포에서 발견되며, 색각의 근원이 된다. 사람은 로돕신 외에 황록색(photopsin I), 녹색(photopsin II), 청자색(photopsin III)의 빛을 최대로 흡수하는 3가지 포토프신을 갖는다.[60]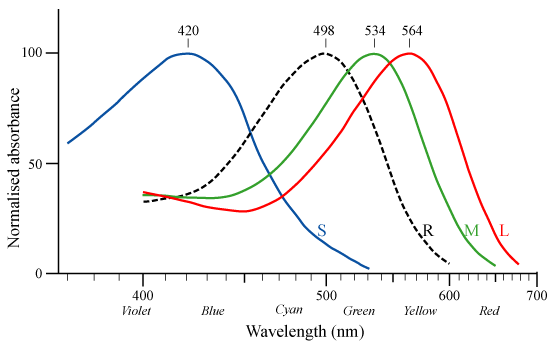
고세균 중에는 광합성을 하는 박테리오로돕신이라는 프로톤 펌프를 발현하는 것이 있다. 박테리오로돕신은 로돕신처럼 레티날을 가지며, 7개의 막 관통 α 나선을 갖지만 G 단백질과는 결합하지 않는다. 조류는 독자적인 광 게이트 이온 채널을 포함한 채널로돕신2라는 옵신을 가지고 있다.[60]
참조
[1]
웹사이트
RHO rhodopsin [Homo sapiens (human)]
https://www.ncbi.nlm[...]
2017-11-16
[2]
서적
Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set)
JAI Press
[3]
서적
Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set)
JAI Press
[4]
서적
Encyclopedia of the Neurological Sciences
https://books.google[...]
Academic Press
2014-04-29
[5]
서적
Photophysiology: General Principles; Action of Light on Plants
https://books.google[...]
Elsevier
2015-09-23
[6]
논문
Zur Anatomie und Physiologie der Retina
https://books.google[...]
1877
[7]
웹사이트
Rhodopsin: History and Etymology for rhodopsin
https://www.merriam-[...]
[8]
논문
Untersuchungen über den Sehpurpur
https://www.biodiver[...]
1878
[9]
논문
Visual purple (sehpurpur)
2008-11
[10]
논문
The photochemical basis of rod vision
1951-12
[11]
논문
Rhodopsin, light-sensor of vision
2023-03
[12]
논문
Iodopsin
1955-05
[13]
웹사이트
Rhodopsin
http://www.britannic[...]
Britannica.com
2016-01-30
[14]
논문
Human rhodopsin
1958-01
[15]
논문
Visual pigments of rods and cones in a human retina
1980-01
[16]
논문
The opsins
[17]
논문
Human cryptochrome exhibits light-dependent magnetosensitivity
2011-06
[18]
논문
G protein involvement in receptor-effector coupling
1988-02
[19]
논문
Fingerprinting G-protein-coupled receptors
1994-02
[20]
논문
Cloning of the gene and cDNA for mammalian beta-adrenergic receptor and homology with rhodopsin
1986-05
[21]
논문
Ligand binding to the beta-adrenergic receptor involves its rhodopsin-like core
1987-03
[22]
논문
Carotenoids and the Vitamin A Cycle in Vision
1934-07
[23]
논문
Hindered Cis Isomers of Vitamin a and Retinene: The Structure of the Neo-B Isomer
1955-07
[24]
논문
The neo-b isomer of vitamin A and retinene
1956-10
[25]
논문
The Synthesis and Configuration of Neo-B Vitamin A and Neoretinine b
1956-06
[26]
논문
HINDERED CIS ISOMERS OF VITAMIN A AND RETINENE: THE STRUCTURE OF THE NEO-b ISOMER
1956-09
[27]
논문
Crystal structure of rhodopsin: A G protein-coupled receptor
2000-08
[28]
논문
Site of attachment of retinal in rhodopsin
1967-12
[29]
논문
The structure of bovine rhodopsin
1983
[30]
논문
Rhodopsin and indicator yellow
1953-03
[31]
논문
Studies on rhodopsin. VIII. Retinylidenemethylamine, an indicator yellow analogue
1955-01
[32]
논문
The Action of Light on Rhodopsin
1958-02
[33]
논문
The mechanism of bleaching rhodopsin
1959-11
[34]
논문
Crystal structure of metarhodopsin II
2011-03
[35]
논문
Molecular basis of visual excitation
1968-10
[36]
논문
Evolution and diversity of opsins
2012-01
[37]
논문
The Gluopsins: Opsins without the Retinal Binding Lysine
2022-08
[38]
논문
Chromophore-Independent Roles of Opsin Apoproteins in Drosophila Mechanoreceptors
2019-09
[39]
논문
Functions of Opsins in Drosophila Taste
2020-04
[40]
논문
Melanopsin triggers the release of internal calcium stores in response to light
2007-03
[41]
논문
Constitutively active mutants of rhodopsin
1992-10
[42]
논문
Synthesis and characterization of a novel retinylamine analog inhibitor of constitutively active rhodopsin mutants found in patients with autosomal dominant retinitis pigmentosa
1997-12
[43]
서적
Pharmacology & Therapeutics of Constitutively Active Receptors
2014
[44]
논문
A comparison of the efficiency of G protein activation by ligand-free and light-activated forms of rhodopsin
1997-12
[45]
논문
Relocating the active-site lysine in rhodopsin and implications for evolution of retinylidene proteins
2013-08
[46]
논문
Rhodopsin and bacteriorhodopsin: structure-function relationships
1982-11
[47]
논문
Photocyclic behavior of rhodopsin induced by an atypical isomerization mechanism
2017-03
[48]
논문
Crystallographic analysis of primary visual photochemistry
2006-06
[49]
논문
Quantum mechanical studies on the crystallographic model of bathorhodopsin
2006-06
[50]
논문
The twisted C11=C12 bond of the rhodopsin chromophore--a photochemical hot spot
2007-09
[51]
논문
Monomeric G-protein-coupled receptor as a functional unit
2005-07
[52]
논문
Rhodopsin Oligomerization and Aggregation
2019-10
[53]
웹사이트
The Nobel Prize in Physiology or Medicine 1967
https://www.nobelpri[...]
Nobel Media AB 2014
2015-12-12
[54]
논문
Pre-lumirhodopsin and the bleaching of visual pigments
1963-03
[55]
논문
Tautomeric Forms of Metarhodopsin
1963-11
[56]
서적
Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set)
JAI Press
[57]
논문
Signaling states of rhodopsin. Formation of the storage form, metarhodopsin III, from active metarhodopsin II
2003-01
[58]
논문
The cellular fate of mutant rhodopsin: quality control, degradation and aggresome formation
https://discovery.uc[...]
2002-07
[59]
논문
Mechanisms of cell death in rhodopsin retinitis pigmentosa: implications for therapy
2005-04
[60]
서적
続・身のまわりの毒
東京化学同人
[61]
간행물
Guest Editorial Essay
Perception
[62]
서적
Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set)
JAI Press
[63]
서적
Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set)
JAI Press
[64]
서적
Encyclopedia of the Neurological Sciences
https://books.google[...]
Academic Press
2014-04-29
[65]
서적
Photophysiology: General Principles; Action of Light on Plants
https://books.google[...]
Elsevier
2015-09-23
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com