다이어울프
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
다이어울프는 약 10,000년 전 멸종된 북아메리카의 멸종된 개과 동물로, 학명은 Canis dirus 또는 Aenocyon dirus이다. 늑대와 유전적으로 매우 분리된 계통이며, 현생 늑대보다 더 크고 강한 턱과 치악력을 지녔다. 무리 생활을 하며 대형 초식동물을 사냥했을 것으로 추정되며, 북아메리카, 남아메리카, 동아시아 등 넓은 지역에서 화석이 발견되었다. 멸종 원인으로는 기후 변화, 경쟁, 먹이 감소, 근친 교배 등이 복합적으로 작용한 것으로 추정된다.
더 읽어볼만한 페이지
- 고대 개과 - 리카온 세코웨이
- 1858년 기재된 화석 분류군 - 크리코사우루스
크리코사우루스는 쥐라기 후기에 살았던 바다악어의 일종으로, 긴 주둥이와 톱 모양 이빨, 염분 배설 땀샘, 유선형 몸체와 지느러미 꼬리 등 바다 생활에 적응한 특징을 가진다. - 1858년 기재된 화석 분류군 - 하드로사우루스
하드로사우루스는 백악기 후기 북아메리카에서 서식한 몸길이 7~8m의 대형 초식 조각류 공룡으로, 북아메리카 최초의 공룡 화석 기록, 최초의 조립된 공룡 골격, 그리고 뉴저지주의 공식 공룡으로 지정된 특징을 가진다. - 플라이스토세 북아메리카의 포유류 - 스밀로돈
스밀로돈은 플라이스토세 시대 아메리카 대륙에 살았던 멸종된 검치호 속으로, 긴 송곳니와 강력한 앞다리, 튼튼한 체격을 지녔으며 대형 초식동물을 사냥했던 최상위 포식자였으나 약 1만 년 전 멸종했다. - 플라이스토세 북아메리카의 포유류 - 퓨마
퓨마는 아메리카 대륙에 서식하며 다양한 이름으로 불리는 대형 고양잇과 동물로, 넓은 분포와 뛰어난 적응력을 가지며 단독 생활을 하는 사냥꾼이지만, 서식지 감소와 인간 활동으로 위협받고 있고 여러 문화에서 상징적인 의미를 지닌다.
| 다이어울프 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Aenocyon dirus |
| 명명자 | 조지프 라이디(1858) |
| 이명 | Canis primaevus 조지프 라이디(1854) Canis dirus 조지프 라이디(1858) Canis indianensis 조지프 라이디(1869) Canis mississippiensis 조엘 아사프 앨런(1876) Canis nehringi 플로렌티노 아메기노(1902) Canis ayersi Sellards, 1916 Aenocyon dirus nebrascensis Frick, 1930 (nomen nudum) |
| 멸종 여부 | 멸종 |
| 아종 | 멸종 Aenocyon dirus dirus 조지프 라이디(1858) 멸종 Aenocyon dirus guildayi 비외른 쿠르텐(1984) |
| 어원 | 그리스어: αἰνός (ainós, 무시무시한) 그리스어: κύων (kúōn, 개) 라틴어: dīrus (무시무시한) |
| 발음 (영어) | IPA: /ˌaɪnəˈsaɪ.ɒn ˈdaɪrəs/ |
| 생물학적 분류 | |
| 속 | Aenocyon속 |
| 종 | 다이어울프 |
| 형태 및 크기 | |
| 키 | 약 2.255 미터 |
| 몸무게 | 약 60 kg ~ 68 kg |
| 생존 시기 | |
| 시기 | 신생대제4기플라이스토세 후기 - 홀로세 초기 (약 30만 년 전 ~ 1만 년 전) |
| 화석 범위 | 플라이스토세 후기 - 홀로세 초기 (125,000년 전 ~ 9,500년 전) |
| 분포 | |
| |
2. 분류
다이어울프는 오랫동안 겉모습의 유사성 때문에 개속(Canis)에 속하는 ''Canis dirus''로 분류되었으며, 회색늑대(Canis lupus)와 가까운 관계로 여겨졌다. 1918년 존 캠벨 메리엄이 별도의 속인 ''Aenocyon''(δεινός κύων|데이노스 퀴온grc, "끔찍한 늑대")을 제안하기도 했으나, ''Canis'' 속 분류가 오랫동안 유지되었다.
그러나 2021년에 발표된 DNA 분석 연구는 이러한 전통적인 분류에 큰 변화를 가져왔다. 이 연구는 다이어울프가 현존하는 개아족(Canina), 특히 회색늑대와 유전적으로 매우 멀리 떨어져 있으며, 약 570만 년 전에 공통 조상으로부터 갈라져 나온 독자적인 신세계 계통임을 밝혔다.[11][12][13] 이는 다이어울프와 회색늑대 간의 형태학적 유사성이 수렴 진화의 결과일 뿐, 가까운 유연관계를 나타내는 것이 아님을 시사한다. 또한, 다이어울프와 다른 북미 개과 동물(회색늑대, 코요테) 사이의 유전자 혼합 증거도 발견되지 않아 오랜 기간 동안 생식적 격리가 이루어졌음을 뒷받침한다.
이러한 유전적 증거는 1918년 메리엄이 제안했던 ''Aenocyon'' 속 분류의 타당성을 강력하게 지지한다. 따라서 현재 학계에서는 다이어울프를 더 이상 ''Canis'' 속이 아닌, 별도의 ''Aenocyon'' 속에 속하는 ''Aenocyon dirus''로 분류하는 것이 적절하다는 견해가 지배적이다.
2. 1. 분류학적 변천
1850년대부터 미국에서는 멸종된 대형 늑대의 화석 유해가 발견되기 시작했으나, 이들이 모두 같은 종에 속하는지는 즉시 명확하지 않았다. 나중에 ''Aenocyon dirus''와 연관될 첫 번째 표본은 1854년 중반 오하이오 강 근처 인디애나주 이반스빌에서 발견되었다. 화석화된 턱뼈와 어금니는 지질학자 조셉 그랜빌 노우드가 이반스빌 수집가 프랜시스 A. 링크로부터 얻었다. 고생물학자 조셉 레이디는 이 표본이 멸종된 늑대 종을 나타낸다고 판단하고 ''Canis primaevus''라는 이름으로 보고했다. 기준 표본은 필라델피아 자연과학 아카데미에 보관되어 있다. 1857년, 레이디는 네브래스카주의 니오브라라 강 계곡을 탐사하던 중 멸종된 ''Canis'' 종의 척추뼈를 발견했고, 이듬해 ''C. dirus''라는 이름으로 보고했다. ''C. primaevus''(Leidy 1854)라는 이름은 레이디가 영국 박물학자 브라이언 호턴 혹슨이 승냥이에게 이미 ''C. primaevus''라는 이름을 사용했다는 것을 알게 되면서 나중에 ''Canis indianensis''(Leidy 1869)로 변경되었다.
1876년 동물학자 조엘 아사프 앨런은 ''Canis mississippiensis''(Allen 1876)의 유해를 발견하고 이를 ''C. dirus''(Leidy 1858)와 ''Canis indianensis''(Leidy 1869)와 연관시켰다. 그러나 이 세 표본에 대한 정보가 부족했기 때문에, 앨런은 관계를 명확히 할 더 많은 자료가 발견될 때까지 각 표본을 잠정적인 이름으로 유지하는 것이 최선이라고 판단했다.
1908년 고생물학자 존 캠벨 메리엄은 캘리포니아주 로스앤젤레스의 라 브레아 타르 구덩이에서 대형 늑대의 화석 뼈 조각을 대량으로 수집하기 시작했다. 1912년까지 그는 이전에 발견된 표본들과 함께 공식적으로 ''C. dirus''(Leidy 1858)라는 이름으로 인정할 만큼 충분히 완전한 골격을 확보했다. 명명법 규칙에 따라 종의 이름은 해당 종에 적용된 가장 오래된 이름을 사용해야 하므로, 메리엄은 레이디의 1858년 표본명인 ''C. dirus''를 선택했다. 1915년 고생물학자 에드워드 트록셀은 ''C. indianensis''를 ''C. dirus''의 동의어로 선언하며 메리엄의 의견에 동의했다. 1918년, 메리엄은 이 화석들을 연구한 후, 이들을 별도의 속인 ''Aenocyon'' (고대 그리스어 'ainos', '끔찍한'과 'cyon', '개'에서 유래)으로 분류하여 ''Aenocyon dirus''로 명명할 것을 제안했다. 그러나 당시 모든 학자가 이 멸종된 늑대를 ''Canis''과는 별개의 새로운 속에 배치하는 데 동의하지는 않았다.
''Canis ayersi''(Sellards 1916)와 ''Aenocyon dirus''(Merriam 1918)는 1972년 고생물학자 어니스트 룬델리우스에 의해 ''C. dirus''의 동의어로 인식되었다. 1979년, 고생물학자 로널드 M. 노와크는 이전에 제안된 모든 관련 분류군(''C. primaevus'', ''C. indianensis'', ''C. mississippiensis'', ''C. ayersi'', ''Aenocyon dirus'')을 ''C. dirus''의 동의어로 정리했다.
1984년 비욘 쿠르텐은 다이어울프 개체군 내의 지리적 변이를 연구하여 두 개의 아종을 제안했다. 캘리포니아와 멕시코에서 발견된, 상대적으로 짧은 사지와 긴 이빨을 가진 표본에 대해서는 ''Canis dirus guildayi'' (고생물학자 존 E. 길데이를 기리기 위해 명명)를, 북미 아메리카 대륙 분기점 동쪽에서 발견된, 더 긴 사지와 짧은 이빨을 가진 표본에 대해서는 ''Canis dirus dirus''를 제안했다. 쿠르텐은 뉴멕시코주 허미츠 동굴에서 발견된 상악골을 기준 아종 ''C. d. dirus''의 대표 표본으로 지정했다.
2021년, DNA 연구 결과 다이어울프는 현존하는 개아족(Canina)과 비교했을 때 유전적으로 매우 멀리 분기된 계통임이 밝혀졌다. 이 발견은 1918년 메리엄이 제안했던 다이어울프의 별도 속 분류, 즉 ''Aenocyon'' (δεινός κύων|데이노스 퀴온grc, "끔찍한 늑대") 속으로의 분류를 지지한다.[11][12][13] 이 연구에 따르면, 다이어울프는 회색늑대보다는 검은등자칼이나 가로줄무늬자칼과 같은 아프리카의 개과 동물과 더 가까운 관계일 수 있으며, 늑대와의 공통 조상으로부터 약 570만 년 전에 갈라져 나온 신세계 고유의 계통으로 여겨진다.[11][12] 이에 따라 다이어울프를 개속(''Canis'')이 아닌 ''Aenocyon'' 속으로 재분류하자는 제안이 힘을 얻고 있다.
2. 2. 진화
북아메리카에서 개과 동물은 약 4천만 년 전에 처음 등장했으며, 개아과는 약 3,200만 년 전에 나타났다. 개아과 내에서 여우와 같은 그룹과 개와 같은 그룹의 조상은 약 900만 년 전에 분화했다. 이후 약 700만 년 전, 개과 동물은 유라시아와 아프리카로 퍼져나갔고, 유럽에서는 ''Canis'' 속의 초기 구성원이 등장했다. 약 400만~300만 년 전에는 중국에서 최초의 늑대 크기 ''Canis'' 속 동물인 ''C. chihliensis''가 나타나 유라시아와 아프리카 전역으로 확산했으며, 이후 ''Canis'' 속의 다른 구성원들이 북아메리카로 이동했다.다이어울프는 북아메리카에서 진화한 것으로 여겨진다. 그러나 그 정확한 조상 계통에 대해서는 논쟁이 있으며, 현재 두 가지 주요 가설이 경쟁하고 있다.
첫 번째 가설은 화석의 형태학적 분석에 기반하며, 다이어울프가 유라시아에서 기원하여 북아메리카로 확장된 ''Canis'' 속의 한 갈래에서 진화했다고 본다. 이 관점은 주로 다이어울프와 다른 ''Canis'' 속 동물들 간의 골격 구조 유사성에 주목한다.
두 번째 가설은 DNA 증거에 기반하며, 다이어울프가 아메리카 대륙에서 독자적으로 기원했으며, 현존하는 늑대 등이 속한 ''Canis'' 속과는 매우 오래전에 갈라진 별개의 계통이라고 주장한다. 2021년 발표된 핵 DNA 분석 결과는 다이어울프가 약 570만 년 전에 늑대와 같은 개과 동물들과 가장 최근의 공통 조상을 공유했으며, 이후 독자적인 진화 경로를 걸었다는 것을 시사한다. 이 연구는 다이어울프와 회색늑대 사이의 형태학적 유사성이 수렴 진화의 결과일 수 있다고 제안하며, 두 계통 간의 유전자 혼합 증거는 발견되지 않았다고 보고했다. 이 DNA 증거는 다이어울프를 ''Canis'' 속이 아닌 별도의 ''Aenocyon'' 속으로 재분류해야 한다는 주장을 뒷받침한다.
2. 2. 1. 형태학적 증거
화석 증거에 기반한 형태학적 분석은 다이어울프가 유라시아에서 기원한 ''Canis'' 속의 확장을 통해 북아메리카에서 출현했음을 시사한다.다이어울프의 직접적인 조상에 대해서는 여러 가설이 존재한다.
고생물학자 샤오밍 왕, 리처드 H. 테드포드, 로널드 M. 노박 등은 다이어울프가 북아메리카의 대형 늑대였던 ''Canis armbrusteri''(아름브루스터 늑대)에서 진화했다고 주장한다. 노박은 두 종 모두 아메리카 대륙에서 기원했으며, 컴벌랜드 동굴(메릴랜드주)에서 발견된 화석이 ''C. armbrusteri''가 ''C. dirus''로 변화하는 과정을 보여주는 증거라고 보았다. 그는 또한 북아메리카 최초의 늑대로 여겨지는 ''Canis edwardii''가 ''C. armbrusteri''와 ''C. dirus'' 계통과 가깝다고 생각했다. 테드포드는 더 나아가 중국의 초기 늑대인 ''Canis chihliensis''가 ''C. lupus''와 ''C. armbrusteri'' 모두의 조상일 수 있다고 제안했다. 그는 ''C. armbrusteri''가 약 150만 년 전 초기 플라이스토세에 매머드와 함께 아시아에서 베링기아를 통해 북아메리카로 이주해 왔을 것으로 추정했다. 반면, 현재의 회색늑대 ''C. lupus''는 이후 플라이스토세 후기에 베링기아에서 진화하여 최후 빙하기 동안 북아메리카로 들어온 것으로 본다.

한편, 로버트 A. 마틴(1974)은 ''C. armbrusteri''가 사실상 ''C. lupus''와 같은 종이라고 주장했으며, 글로리아 D. 고울렛(1993)은 이를 바탕으로 풍부한 먹이 자원 등 특정 환경 조건 하에서 ''C. lupus''가 더 큰 형태인 ''C. dirus''로 진화(과형태화)했을 수 있다는 가설을 제시했다. 그러나 노박, 쿠르텐, 안날리사 베르타 등 다른 학자들은 ''C. dirus''가 ''C. lupus''에서 직접 파생된 것은 아니라고 반박했다. 2010년 프란시스코 프레보스티는 ''C. dirus''가 ''C. lupus''의 자매 분류군이라고 제안하기도 했다.
화석 기록에 따르면, ''C. dirus''는 약 25만 년 전(YBP) 북아메리카 중부 대륙의 개방된 지형에서 처음 나타나 동쪽으로 확산하며 기존의 ''C. armbrusteri''를 대체한 것으로 보인다. 가장 오래된 ''C. dirus'' 화석 증거 중 일부는 네브래스카주 헤이 스프링스 지역과 사우스다코타주 블랙힐스의 도롱뇽 동굴에서 발견되었다. 특히 도롱뇽 동굴의 화석은 ''Canis cf. C. dirus''로 분류되었는데 (여기서 cf.는 라틴어 'confer'의 약자로, 종 동정이 불확실함을 의미), 우라늄-토륨 연대 측정 결과 약 252,000년 전(YBP)의 것으로 추정된다. 이후 다이어울프는 북미 전역(미국, 캐나다, 멕시코)과 남아메리카 일부(베네수엘라, 에콰도르, 볼리비아, 페루)로 퍼져나갔다. 다이어울프는 후기 플라이스토세부터 초기 홀로세까지(125,000–10,000 YBP) 생존했다. 북미에서는 동부의 ''C. d. dirus''(125,000–75,000 YBP)와 서부의 더 작고 후대의 아종인 ''C. d. guildayi''로 나뉘기도 한다.
남아메리카에서는 후기 플라이스토세 시기의 ''C. dirus'' 화석이 주로 북쪽과 서쪽 해안에서 발견되지만, 아르헨티나 지역에서는 발견되지 않았다. 이 지역에서는 ''Canis gezi''와 ''Canis nehringi''라는 다른 대형 개과 동물이 서식했다. 일부 연구자들은 형태학적 유사성을 근거로 ''C. dirus''와 ''C. nehringi''가 동일한 종이며, 다이어울프가 아메리카 대교환 시기에 북아메리카에서 남아메리카로 이주했다고 결론지었다. 반면, 2018년의 한 연구는 ''Canis gezi''가 ''Canis'' 속이 아닌 세르도키오니나 아족에 속한다고 주장하기도 했다.
2020년에는 중국 북동부에서 다이어울프 화석이 발견되어, 이들이 당시 존재했던 육교인 베링기아를 건너 아시아와 아메리카 대륙을 오갔을 가능성도 제기되었다.[3]
2. 2. 2. DNA 증거
DNA 증거는 다이어울프가 아메리카 대륙에서 기원하여 ''Canis'' 속과 분리된 조상 계통에서 유래되었음을 나타낸다.1992년에는 ''A. d. guildayi''의 골격 유해에서 미토콘드리아 DNA 서열을 추출하여 다른 ''Canis'' 종과의 관계를 비교하려는 시도가 있었다. 그러나 이 유해들은 라 브레아 구덩이에서 나왔고, 뼈 물질에서 타르를 제거할 수 없어 이 시도는 실패했다. 2014년에도 타르 구덩이에서 콜럼비아 매머드의 DNA를 추출하려는 시도가 실패했으며, 연구자들은 아스팔트의 유기 화합물이 라 브레아 구덩이의 모든 고대 표본의 뼈를 침투하여 DNA 샘플 추출을 방해한다고 결론 내렸다.
2021년, 연구자들은 13,000년에서 50,000년 전의 다섯 개 다이어울프 화석에서 추출한 핵 DNA(세포 핵에서)를 시퀀싱했다. 이 서열 분석 결과, 다이어울프는 약 570만 년 전에 늑대와 같은 개과 동물과 가장 최근의 공통 조상을 공유한, 매우 분화된 계통임이 밝혀졌다. 이 연구는 또한 다이어울프와 회색 늑대의 골격 표본을 측정한 결과, 형태학적 유사성은 수렴 진화의 결과이며, 이전에 생각했던 것과는 달리 가까운 진화적 관계를 의미하지는 않는다고 결론 내렸다. 늑대와 같은 개과 동물은 서로 교배하는 것으로 알려져 있지만, 이 연구에서는 다섯 개의 다이어울프 샘플에서 현존하는 북미 회색 늑대, 코요테 또는 그들의 공통 조상과의 유전자 혼합 증거를 발견하지 못했다. 이는 늑대와 코요테 계통이 다이어울프 계통과 별개로 진화했음을 보여준다.
이 연구는 다이어울프 계통이 아메리카 대륙에서 기원했으며, 이러한 지리적 격리가 570만 년 전 분기 이후 어느 정도의 생식적 격리를 가능하게 했다고 제안한다. 코요테, 승냥이, 회색 늑대, 그리고 멸종된 ''Xenocyon''은 유라시아에서 진화하여 후기 플라이스토세 동안 비교적 최근에 북아메리카로 확장되었기 때문에 다이어울프와 혼합되지 않았다. 다이어울프 계통의 오랜 격리는 ''C. armbrusteri''와 ''C. edwardii''를 포함한 다른 아메리카 화석 분류군 역시 다이어울프 계통에 속할 가능성을 시사한다.
이러한 DNA 연구 결과는 과거 일부 학자들이 제안했던, 다이어울프를 기존의 ''Canis''이 아닌 별도의 ''Aenocyon'' 속으로 분류하는 분류학적 제안을 지지한다. 2021년 발표된 게놈 분석 논문에 따르면, 다이어울프는 늑대와 약 570만 년 전에 갈라져 나온 '신세계' 계통이며, 유라시아 늑대보다는 오히려 검은등자칼이나 가로줄무늬자칼과 같은 아프리카 자칼류에 더 가까울 수 있다고 제안되었다.[11][12][13]
3. 형태
다이어울프는 플라이스토세 후기에 북미와 남미 일부 지역에 서식했던 대형 개과 동물이다. 전체적인 크기는 현존하는 가장 큰 회색늑대 아종들과 비슷하거나 약간 더 컸지만, 훨씬 더 육중하고 튼튼한 골격을 지녔다. 평균 체중은 아종에 따라 60kg에서 68kg 정도로 추정되며, 이는 현대 회색늑대의 평균 체중보다 상당히 무거운 편이다.
같은 크기의 회색늑대와 비교했을 때, 다이어울프는 머리가 더 크고 넓었으며, 일부 아종의 경우 다리가 상대적으로 짧고 발이 작았다. 특히 로키 산맥 서쪽의 개체군(''Aenocyon dirus guildayi'')은 동쪽의 개체군(''Aenocyon dirus dirus'')보다 사지가 짧아, 회색늑대만큼 빠른 달리기에 적합하지 않았을 수 있다.
두개골과 치아는 회색늑대보다 크고 강력했으며, 특히 열육치는 먹이를 자르는 능력이 뛰어났다. 턱 근육이 발달하여 강한 무는 힘을 낼 수 있었고, 송곳니는 크고 튼튼하여 큰 먹이와 싸우는 데 유리했다. 이러한 형태적 특징들은 다이어울프가 거대동물을 포함한 큰 먹이를 사냥하는 데 특화되었음을 시사한다. 완전한 수컷 다이어울프의 음경골은 다른 현존하는 개과 동물들과 뚜렷하게 달라 종 식별에 도움이 되기도 한다.
3. 1. 신체 특징
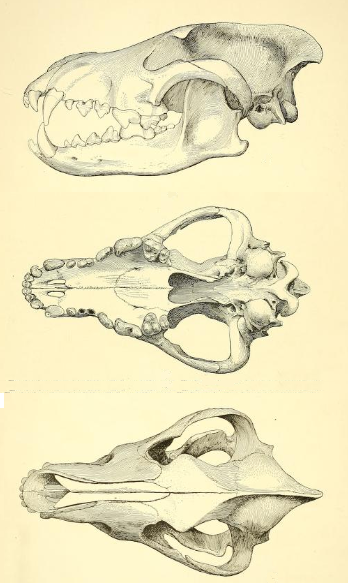
다이어울프의 신장은 대략 1.5m에 체중은 50kg ~ 79kg였다.[18][19] 일부 개체군에서는 수컷의 송곳니가 암컷보다 상당히 커서 번식 경쟁에 사용되었을 가능성이 있지만, 다른 개체군에서는 송곳니의 성적 이형이 거의 나타나지 않아 경쟁이 드물었음을 시사한다.[20]
겉보기에는 회색늑대와 비슷했지만, 다이어울프는 훨씬 크고 튼튼한 체격을 가졌다. 현대의 가장 큰 회색늑대조차 다이어울프의 평균 크기에 불과했으며, 가장 큰 다이어울프는 현존하는 어떤 회색늑대보다도 거대했다. 다이어울프는 현생 회색늑대보다 체중이 약 25% 더 나갔다.[20] 이러한 육중한 체격은 더 큰 거대동물을 사냥하기 위해 필요했던 것으로 보인다.[21] 회색늑대와 비교했을 때 다이어울프의 다리는 상대적으로 짧았고, 두개골은 더 컸다.[22]
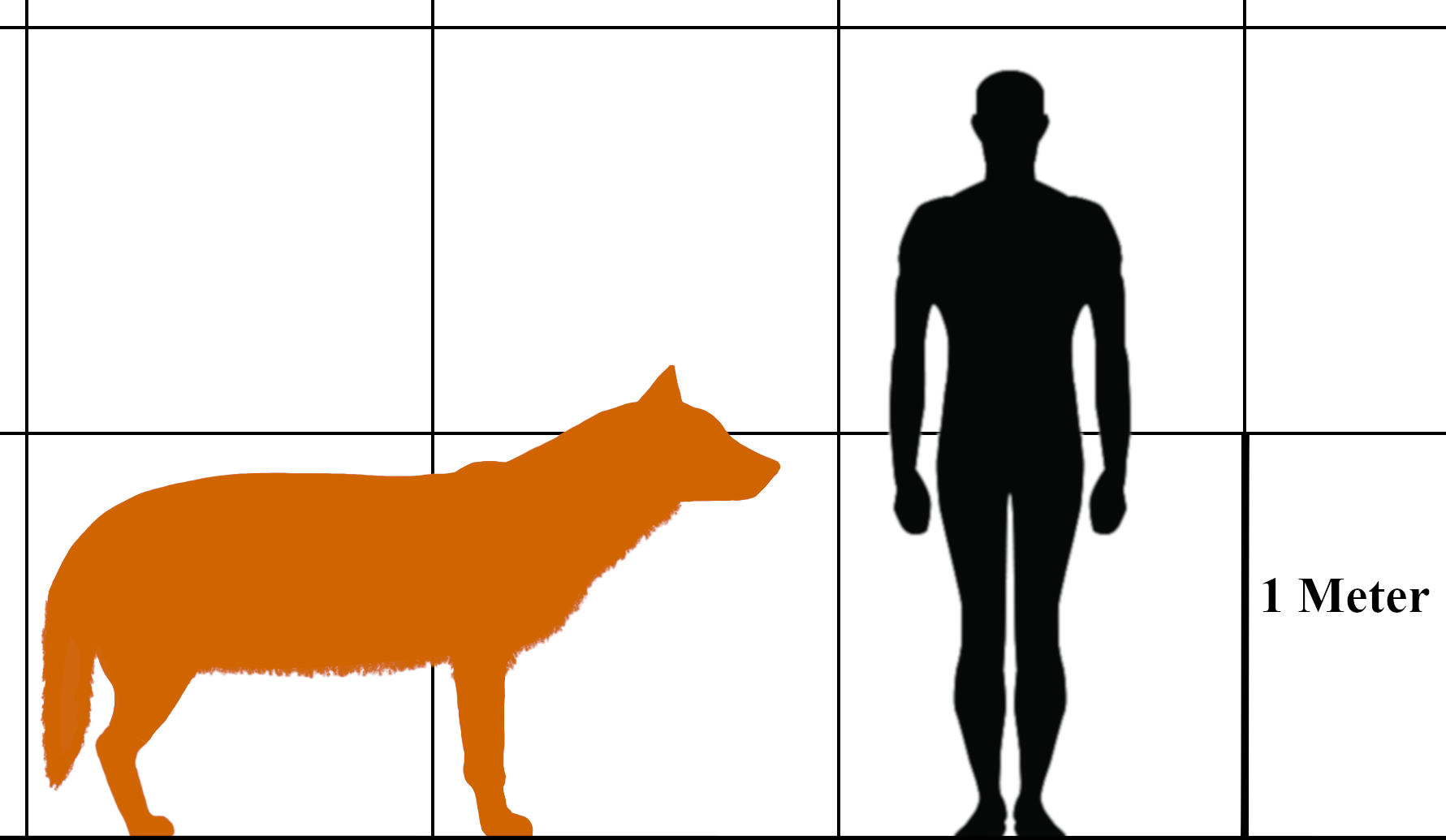
평균적인 다이어울프의 신체 비율은 오늘날 북아메리카에 서식하는 유콘 늑대(''Canis lupus pambasileus'') 와 북서부 늑대(''Canis lupus occidentalis'')와 유사했다. 오늘날 가장 큰 북부 늑대의 어깨 높이는 최대 약 96.52cm이고 몸 길이는 약 175.26cm이다. 랜초 라 브레아에서 발견된 일부 다이어울프 표본은 이보다 작거나 더 컸다.
다이어울프는 같은 크기의 북부 늑대에 비해 발이 작고 머리가 더 컸다. 두개골 길이는 최대 310mm 이상이었으며, 구개, 전두엽, 관골궁이 유콘 늑대보다 넓었다. 이러한 특징 때문에 두개골이 매우 거대해 보였다. 시상 능선은 더 높았고, 이온(occipital torus)은 눈에 띄게 뒤로 돌출되었으며, 비골 뒷부분은 두개골 안쪽으로 비교적 멀리 뻗어 있었다. 랜초 라 브레아에서 발견된 다이어울프의 완전한 골격은 타르 때문에 뼈가 흩어져 찾기 어렵지만, 조립된 척추 일부는 현대 늑대와 유사하며 같은 수의 척추뼈를 가진 것으로 확인되었다.

1984년 골격 연구를 통해 다이어울프의 지리적 차이가 밝혀졌다. 캘리포니아와 멕시코에서 발견된 아종(''Aenocyon dirus guildayi'')과 대륙 분할선 동쪽에서 발견된 아종(''Aenocyon dirus dirus'') 사이에 두개골-치아 특징과 사지 비율에서 차이가 나타났다. ''A. d. guildayi''는 유콘 늑대보다 뒷다리가 8% 짧았는데, 이는 현저히 짧은 경골과 중족골 때문이었다. 앞다리 역시 하위 뼈들이 약간 짧아 전체적으로 짧았다. 상대적으로 가볍고 작은 사지와 거대한 머리를 가진 ''A. d. guildayi''는 늑대나 코요테만큼 달리기에 적합하지 않았다. 반면, ''A. d. dirus''는 ''A. d. guildayi''보다 훨씬 긴 사지를 가졌다. 앞다리는 상완골이 10%, 요골이 15%, 중수골이 15% 더 길어 전체적으로 14% 더 길었고, 뒷다리는 대퇴골과 경골이 10%, 중족골이 15% 더 길어 전체적으로 10% 더 길었다. ''A. d. dirus''의 사지 길이는 유콘 늑대와 비슷했다. 가장 큰 ''A. d. dirus'' 대퇴골은 미주리주 캐롤 동굴에서 발견되었으며 길이는 278mm였다.
| 사지 부위 | A. d. guildayi | 유콘 늑대 | A. d. dirus |
|---|---|---|---|
| 상완골 (앞다리 위쪽) | 218mm | 237mm | 240mm |
| 요골 (앞다리 아래쪽) | 209mm | 232mm | 240mm |
| 중수골 (앞발) | 88mm | 101mm | 101mm |
| 대퇴골 (뒷다리 위쪽) | 242mm | 251mm | 266mm |
| 경골 (뒷다리 아래쪽) | 232mm | 258mm | 255mm |
| 중족골 (뒷발) | 93mm | 109mm | 107mm |
평균 체중은 ''A. d. guildayi''가 60kg, ''A. d. dirus''가 68kg였으며, 일부 표본은 더 컸지만 골격 구조상 110kg를 넘기 어려웠을 것으로 추정된다. 참고로 유콘 늑대의 평균 체중은 수컷 43kg, 암컷 37kg이며, 개체별 체중은 21kg에서 55kg까지 다양하고, 가장 무거운 개체는 79.4kg에 달했다. 이는 평균적인 다이어울프가 가장 큰 현대 회색 늑대와 크기가 유사했음을 보여준다.
완전한 수컷 ''A. dirus''의 유해는 다른 ''Canis'' 표본과 비교하여 때때로 쉽게 식별할 수 있는데, 다이어울프의 음경골 (음경뼈)이 다른 모든 살아있는 개과 동물과 매우 다르기 때문이다. 두개골-미저골 길이는 약 125cm, 꼬리 길이는 약 60cm, 어깨 높이는 약 80cm이다. 현존하는 회색늑대의 대형 아종과 비슷한 크기이지만, 더 묵직한 체형으로 평균 체중은 현존하는 북미 회색늑대의 평균보다 더 무거웠을 것으로 추정된다. ''A. d. dirus''는 ''A. d. guildayi''보다 사지가 길다. 수컷은 현존하는 개과의 종에 비해 현저하게 큰 음경골을 가지고 있었다. 머리 부분은 넓고, 측두창과 관골궁의 확대로 교근에 큰 부착부를 제공했다. 또한, 턱은 튼튼하며, 회색늑대보다 큰 이빨을 가지고 있었다.
3. 2. 치아
다이어울프의 이빨은 크기가 다소 크다는 점을 제외하면 회색늑대의 그것과 유사하며, 이는 초육식성에서 중육식성 활동을 시사한다. 고생물학자 R.M. 노와크(R.M. Nowak)는 다이어울프가 주로 육식을 했지만 부분적으로 잡식성이었을 수도 있다고 보았다.[23] 윗턱 열육치(P4)는 회색늑대보다 크지만, 아랫턱은 비슷하다. 윗 열육치의 날은 회색늑대보다 훨씬 커서 먹잇감을 잡아찢는 능력이 더 뛰어났음을 의미한다. 또한 측두와는 길었고 관골궁은 넓었는데, 이는 회색늑대보다 더 큰 힘을 낼 수 있는 커다란 측두근이 있었음을 시사하며, 실제로 다이어울프의 측두근은 현생 회색늑대보다 훨씬 큰 힘, 즉 더 센 무는 힘을 낼 수 있었다.[20]많은 고생물학자들은 다이어울프가 상대적으로 큰 이빨을 뼈를 부수는 데 사용했을 것이라고 추측한다. 이러한 생각은 화석화된 다이어울프의 이빨 끝이 대부분 닳아 있다는 사실로 뒷받침된다.
개속(Canis) 내에서 다이어울프는 아메리카 대륙에서 가장 진화적으로 발달된 늑대형 종으로 여겨졌다. 다른 모든 ''개속'' 종과 구별되는 특징은 다음과 같다: "후방 소첨두가 있는 P2; 두 개의 후방 소첨두가 있는 P3; 메타스타일, 엔토크리스티드, 엔토코눌리드 및 메타코니드에서 과원추 선반까지 뻗어 있는 횡단 능선이 있는 M1; 엔토크리스티드 및 엔토코눌리가 있는 M2."
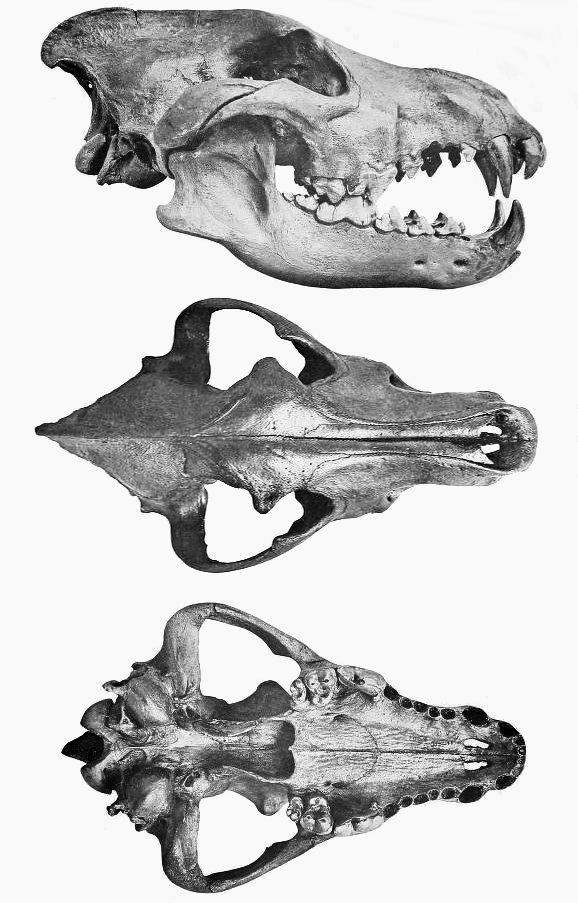
살아있는 포유류와 화석 포식자의 송곳니 물어뜯는 힘을 체중에 맞춰 비교한 연구에 따르면, 태반류 포유류 중 송곳니에서의 물어뜯는 힘(체중 1kg당 뉴턴)은 다이어울프(163)에서 가장 컸다. 이는 종종 자신보다 큰 동물을 잡아먹는 현대의 초육식성 개과 동물인 아프리카 야생 개(142), 회색늑대(136), 승냥이(112), 딩고(108)보다도 강한 수치이다. 카르나시알에서의 물어뜯는 힘도 송곳니와 유사한 경향을 보였다. 포식자가 사냥할 수 있는 먹이의 최대 크기는 생체역학적 한계에 크게 영향을 받는데, 다이어울프의 높은 물어뜯는 힘은 이들이 비교적 큰 동물을 잡아먹었음을 시사한다. 다만, 뼈를 먹는 점박이하이에나의 물어뜯는 힘 등급(117)은 송곳니와 카르나시알에서의 높은 물어뜯는 힘이 반드시 뼈를 먹는 데 필수적이지는 않다는 점을 보여준다.

다이어울프의 두개골 측정과 턱 근육 연구에 따르면, 15가지 측정치 중 4가지를 제외하고는 현대 회색늑대와 큰 차이가 없었다. 주요 차이점은 다이어울프의 위쪽 치아가 더 크고, 특히 P4(윗턱 열육치)는 날이 비교적 크고 무거워 절단 능력이 향상되었다는 점이다. 다이어울프의 턱은 회색늑대보다 상대적으로 더 넓고 큰 측두근을 가지고 있어 더 강한 물어뜯는 힘을 낼 수 있었다. 아래쪽 앞어금니는 회색늑대보다 약간 더 컸고, 아래턱 열육치(m1)는 훨씬 더 크고 절단 능력이 뛰어났다. 다이어울프의 송곳니는 같은 크기의 살아있는 개과 동물의 송곳니보다 굽힘 강도가 더 컸으며, 이는 하이에나나 고양이과 동물의 송곳니와 유사했다. 이러한 특징들은 다이어울프가 회색늑대보다 더 강하게 물 수 있었고, 더 유연하고 둥근 송곳니를 가져 먹이와 싸우는 데 더 잘 적응했음을 나타낸다.
| 치아 변수 | lupus 현대 북미 | lupus 라 브레아 | lupus 베링기아 | dirus dirus 상가모니안 시대 (125,000–75,000 YBP) | dirus dirus 후기 위스콘신(50,000 YBP) | dirus guildayi (40,000–13,000 YBP) |
|---|---|---|---|---|---|---|
| m1 길이 | 28.2 | 28.9 | 29.6 | 36.1 | 35.2 | 33.3 |
| m1 너비 | 10.7 | 11.3 | 11.1 | 14.1 | 13.4 | 13.3 |
| m1 삼각두 길이 | 19.6 | 21.9 | 20.9 | 24.5 | 24.0 | 24.4 |
| p4 길이 | 15.4 | 16.6 | 16.5 | 16.7 | 16.0 | 19.9 |
| p4 너비 | - | - | - | 10.1 | 9.6 | 10.3 |
| p2 길이 | - | - | - | 15.7 | 14.8 | 15.7 |
| p2 너비 | - | - | - | 7.1 | 6.7 | 7.4 |
치아 파절은 육식 동물의 행동과 관련이 깊다. 라브레아 타르 구덩이에서 발견된 36,000년에서 10,000년 전 사이의 대형 육식 동물 화석 연구에 따르면, 다이어울프, 코요테, 아메리카 사자, 스밀로돈의 치아 파절률은 5~17%로, 현대 포식 동물 10종의 0.5~2.7%보다 훨씬 높았다. 이러한 높은 파절률은 모든 치아에서 나타났지만, 송곳니의 파절률은 현대 육식 동물과 비슷했다. 다이어울프는 현대 회색늑대에 비해 앞니가 더 자주 부러졌는데, 이는 먹이를 먹을 때 뼈에 더 가깝게 앞니를 사용했음을 시사한다. 멕시코와 페루에서 발견된 다이어울프 화석에서도 유사한 파절 패턴이 나타난다. 1993년 연구에서는 홍적세 육식 동물에서 치아 파절 빈도가 높은 것이 반드시 더 큰 사냥감을 사냥했기 때문은 아니라고 보았다. 대신 먹이 가용성이 낮아지면서 육식 동물 간의 경쟁이 심화되었고, 이로 인해 더 빨리 먹게 되면서 뼈를 더 많이 섭취하게 되어 치아 파절이 증가했을 수 있다고 제안했다. 약 10,000년 전 먹이가 멸종하면서 잡식성인 코요테를 제외한 이들 홍적세 육식 동물들도 멸종했다.
이후 라 브레아 타르 구덩이 연구에서는 두 시대(15,000년 전과 13,000년 전)의 다이어울프 치아 파절을 비교했다. 그 결과, 15,000년 전 다이어울프는 13,000년 전 다이어울프보다 3배 더 많은 치아 파절을 보였으며, 13,000년 전 다이어울프의 파절률은 현대 육식 동물과 비슷한 수준이었다. 이는 15,000년에서 14,000년 전 사이에 먹이 가용성이 낮거나 경쟁이 더 심했고, 13,000년 전까지 먹이 종이 멸종하면서 포식자 경쟁이 감소하여 다이어울프의 치아 파절 빈도도 감소했음을 시사한다.
육식 동물의 사냥 방식은 아래턱 결합부의 강도를 통해 추론할 수 있다. 단독 사냥꾼은 강한 물기로 먹이를 제압하므로 강한 결합부를 가지는 반면, 무리 사냥꾼은 여러 번 얕게 물어뜯기 때문에 상대적으로 약한 결합부를 가진다. 다이어울프의 아래턱 결합부 강도는 회색늑대나 붉은늑대보다 낮지만, 코요테나 아프리카 사냥개와 매우 유사했다. 이는 다이어울프가 현대 친척들처럼 얕은 물기를 사용했으며, 따라서 무리 사냥꾼이었을 가능성을 시사한다. 또한, 이는 다이어울프가 뼈를 처리할 수는 있었지만 회색늑대만큼 뼈를 부수는 데 특화되지는 않았음을 의미할 수 있다. 후기 홍적세에 다이어울프의 골절 발생 빈도가 현대 친척 수준으로 감소했다는 사실은 경쟁 감소로 인해 다이어울프가 뼈 섭취량이 적은, 즉 자신들에게 더 적합한 먹이 섭취 행동으로 돌아갈 수 있었음을 시사한다.
그러나 라 브레아 타르 구덩이 육식 동물의 치아 에나멜에 대한 치아 미세 마모 연구 결과는 다르게 해석될 여지를 남긴다. 이 연구에 따르면, 다이어울프를 포함한 육식 동물들은 멸종 직전에 먹이 스트레스를 받지 않았으며, 사체 활용 정도(뼈까지 먹는 정도)는 오늘날의 대형 육식 동물보다 오히려 낮았다. 이는 치아 파절이 사냥 행동과 먹이의 크기와 더 관련이 있을 수 있음을 시사한다.
4. 생태
다이어울프는 플라이스토세 후기 북아메리카와 남아메리카 일부 지역에 서식했던 개과의 포식자이다. 마지막 빙하기(약 125,000년 전 ~ 14,500년 전 BP) 동안 번성했으며, 당시의 플라이스토세 거대동물들과 함께 생태계를 이루었다. 특히 캘리포니아주 로스앤젤레스의 라 브레아 타르 웅덩이에서는 수많은 다이어울프 화석이 발견되어 이들의 생태 연구에 중요한 정보를 제공한다. 이 타르 웅덩이에서는 다이어울프 외에도 검치호(스밀로돈), 아메리카사자, 콜럼비아 매머드 등 다양한 동식물 화석이 함께 발견되었다.
다이어울프는 현생 회색늑대보다 크고 무거웠으며, 특히 더 강력한 치악력과 튼튼한 치아를 가지고 있었다. 이는 다이어울프가 자신보다 큰 먹이를 사냥하는 데 적응했음을 보여준다. 동위원소 분석 결과, 다이어울프는 주로 말, 들소, 낙타, 땅늘보와 같은 대형 초식동물을 먹이로 삼았으며, 때로는 해안가에 떠밀려온 고래 사체를 먹기도 했다. 먹이를 두고 검치호, 아메리카사자 등 다른 대형 포식자들과 경쟁했다.
라 브레아 타르 웅덩이에서 다이어울프와 검치호 화석이 대량으로 발견된 점은 이들이 사회적 동물이었음을 시사한다. 무리를 지어 사냥하고 먹이를 먹다가 타르 웅덩이에 빠진 먹이 주변에 모여들어 함께 함정에 빠졌을 가능성이 있다. 다이어울프는 성적 이형성이 거의 없어 현생 늑대처럼 일부일처제를 이루며 무리 생활을 했을 것으로 보인다.
서식지는 북아메리카의 평원, 초원, 산림 지역부터 남아메리카의 건조한 사바나까지 광범위했으며, 동아시아에서도 화석이 발견되었다. 주로 대형 초식동물이 풍부한 개방된 저지대 환경을 선호한 것으로 보인다. 플라이스토세 말기의 급격한 기후 변화는 서식 환경과 먹이 가용성에 영향을 미쳤으며, 이는 신체 크기 변화나 치아 마모 및 파절 양상에서도 확인된다. 결국 다이어울프는 약 1만 년 전 제4기 대멸종 시기에 다른 거대동물들과 함께 멸종했다.
4. 1. 서식지
다이어울프의 서식지는 매우 다양했다. 북아메리카에서는 평원, 초지, 산림 지대에 걸쳐 서식했으며, 남아메리카에서는 건조한 사바나 환경에서 발견되었다. 주로 해발 2255m 이상의 고지대를 선호했다.[20] 다이어울프 유해가 발견된 장소들은 주로 먹잇감인 대형 초식동물과 함께 개방된 저지대 환경이었음을 시사한다.대부분의 다이어울프 서식지 연대는 생층서학을 통해 추정되지만, 아스팔트 퇴적층에서는 이 방법의 신뢰도가 떨어진다. 일부 지역에서는 방사성 탄소 연대 측정이 사용되었는데, 예를 들어 라 브레아 구덩이에서 발견된 다이어울프 표본들의 연대는 13,000년 전(YBP)부터 37,000년 전(YBP)까지 다양하게 측정되었다. 미주리주 파우더 밀 크릭 동굴(Powder Mill Creek Cave) 표본은 약 13,170년 전(YBP)으로 측정되었다.
다이어울프가 번성했던 시기는 "빙하기"라고도 불리는 마지막 빙하기(약 125,000년 전 ~ 14,500년 전 BP)와 겹친다. 이 시기는 플라이스토세 후기에 해당하며, 마지막 빙기 극대기(약 33,000년 전 ~ 26,500년 전 BP)에 빙상이 최대로 확장되었다. 해빙은 북반구에서 약 19,000년 전 BP, 남극에서 약 14,500년 전 BP에 시작되었다. 북아메리카 북부는 위스콘신 빙기의 빙상 때문에 접근이 어려웠으며, 마지막 빙하기 말에는 플라이스토세 거대동물의 제4기 대멸종이 일어났다.
마지막 빙기 동안(약 60,000년 전 ~ 빙하기 말) 캘리포니아 남부 해안은 오늘날보다 더 시원하고 습했으며, 연평균 기온은 11°C에서 5°C로, 연 강수량은 100cm에서 45cm로 감소했다. 이 지역은 빙하기 동안 동물과 추위에 민감한 식물의 생물 피난처 역할을 했을 것으로 여겨진다. 약 24,000년 전 BP까지 참나무와 관목은 줄고 소나무가 늘어나 개방된 공원 같은 환경을 이루었으나, 14,000년 전 BP 이후로는 침엽수가 줄고 참나무 숲, 관목, 해안 세이지 스크럽 등 현대와 유사한 식생이 증가했다. 샌타모니카 북쪽 지역은 약 28,000~26,000년 전 BP 사이프러스와 소나무가 우세한 해안 세이지 스크럽 환경이었으며, 샌타모니카 산맥은 다양한 식물 군락을 지탱했다. 이는 겨울 강우 패턴은 현재와 비슷하지만, 전반적으로 더 시원하고 습하며 계절 변화가 적었음을 시사한다. 이러한 환경은 다이어울프와 그 경쟁자들의 먹이가 되는 대형 초식동물을 부양했다.
과거 연구에서는 다이어울프의 체형 변화가 기후 변동과 관련이 있다고 보았다. 라 브레아의 네 개 구덩이(각기 다른 시기 대표)에서 나온 다이어울프 두개골과 치아 분석 결과, 시간에 따라 크기, 치아 마모 및 파절, 두개골 및 주둥이 모양이 변했음이 밝혀졌다. 특히 마지막 빙기 극대기 시작 시점과 따뜻한 알레뢰드 진동 시기 종말 무렵 사이에 체구가 작아졌다. 작은 몸집, 더 큰 두개골 기저부, 짧아진 주둥이(모양 및 크기 유형성숙), 더 많은 치아 파절 및 마모 현상에서 식량 스트레스(영양 섭취 감소를 초래하는 식량 부족)의 증거가 나타난다. 17,900년 전 YBP의 다이어울프는 이러한 특징을 모두 보여 식량 스트레스를 겪었음을 시사한다. 28,000년 전 YBP의 다이어울프 또한 어느 정도 이러한 특징을 보였지만, 연구된 늑대 중 가장 컸으며, 이 늑대들 역시 식량 스트레스를 겪었고 이보다 더 이른 시기의 늑대들은 더 컸을 것으로 추정된다. 영양 스트레스는 사체를 더 완전히 소비하고 뼈를 부수기 위해 더 강한 물기 힘을 유발할 수 있으며, 두개골 모양의 변화는 기계적 이점을 개선하기 위한 것으로 보인다. 북미 기후 기록은 단스가르트-외슈거 사건으로 불리는, 급격한 온난화와 점진적인 냉각이 반복되는 빙하기 동안의 주기적 변동을 보여준다. 이러한 주기는 온도 상승과 건조화를 유발했으며, 라 브레아에서는 생태적 스트레스, 즉 식량 스트레스를 야기했을 것이다. 유사한 경향이 회색늑대에서도 발견되었는데, 홀로세 이전 샌타바버라 분지의 회색늑대는 원래 거대하고 튼튼했으며 다이어울프와 수렴 진화했을 가능성이 있지만, 홀로세 초기에 더 가냘픈 형태로 대체되었다.
:
| 변수 | 28,000 YBP | 26,100 YBP | 17,900 YBP | 13,800 YBP |
|---|---|---|---|---|
| 체구 | 가장 큼 | 큼 | 가장 작음 | 중간/작음 |
| 치아 파절 | 높음 | 낮음 | 높음 | 낮음 |
| 치아 마모 | 높음 | 낮음 | 높음 | 낮음 |
| 주둥이 모양 | 짧아짐, 가장 큰 두개골 기저부 | 평균 | 가장 짧음, 가장 큰 두개골 기저부 | 평균 |
| 치열 모양 | 튼튼함 | – | – | 가냘픔 |
| DO 사건 | 3 또는 4번 | 없음 | 부정확한 데이터 | 부정확한 데이터 |


다이어울프 유해는 북아메리카의 평원, 초원, 일부 산림 지역과 남아메리카의 건조한 사바나 등 광범위한 지역에서 발견되었다. 서식 고도는 해수면부터 2255m까지 다양했다. 북아메리카 고위도 지역에서는 유해가 드물게 발견되는데, 이는 온도, 먹이, 또는 서식지 제한 때문일 수 있다. 북위 42도선 이북에서는 오리건주 화석호(125,000–10,000 YBP), 아이다호주 아메리칸 폭포 저수지(125,000–75,000 YBP), 사우스다코타주 샐러맨더 동굴(250,000 YBP), 네브래스카 북부 등에서 확인되지 않은 화석이 보고된 바 있다.
- 미국: 애리조나, 캘리포니아, 플로리다, 아이다호, 인디애나, 캔자스, 켄터키, 미주리, 네브래스카, 뉴멕시코, 오리건, 펜실베이니아, 사우스캐롤라이나, 사우스다코타, 텍사스, 유타, 버지니아, 웨스트버지니아, 와이오밍, 네바다에서 보고되었다. 캘리포니아 북쪽 보고는 확인이 필요하다. ''A. d. dirus'' 아종의 주요 화석지는 로키 산맥 동쪽의 텍사스 프리젠하운 동굴, 미주리 캐롤 동굴, 플로리다 레딕 등이다.
- 멕시코: 아과스칼리엔테스 엘 세다조, 바하칼리포르니아 수르 코몬두 시, 산루이스포토시 엘 세드랄, 멕시코 주 엘 타호 채석장(테키스퀴악 근처), 푸에블라 발세키요, 할리스코 차팔라 호수, 유카탄 롤툰 동굴, 시날로아 포트레시토, 누에보레온 산 호세시토 동굴(아람베리 근처), 소노라 테라파 등에서 발견되었다. 테라파 표본은 ''A. d. guildayi'' 아종으로 확인되었다. 산 호세시토 동굴과 엘 세다조에서 단일 지역 중 가장 많은 개체 유해가 발견되었다.
- 남아메리카: 약 17,000년 전 YBP 유해가 베네수엘라 서부 팔콘주 무아코 및 동부 모나가스주, 페루 탈라라 지방, 볼리비아 타리하주, 칠레 아타카마 사막, 에콰도르 등에서 보고되었다.[7][8] 북아메리카에서 기원하여 안데스 산맥 회랑을 통해 남아메리카로 확산했을 가능성이 제기된다. 이 경로는 빙하기 동안 시원하고 건조하며 개방된 서식지였기에 이동에 유리했을 것으로 추정된다.
- 동아시아: 2020년, 중국 북동부 하얼빈 근처에서 약 40,000년 전 YBP의 다이어울프 턱뼈 화석이 발견되었다. 이는 기존에 북위 42도선 이북의 추운 기후와 빙상이 이동 장벽이었을 것이라는 가설(해당 위도 이북 화석 부재에 근거)에 도전하며, 다이어울프가 먹이를 따라 베링기아를 거쳐 유라시아로 이동했을 가능성을 시사한다.[14]
4. 2. 행동

다이어울프는 사회적인 동물이었을 가능성이 높으며, 거대한 무리를 지어 생활했을 것으로 추정된다. 이러한 추정은 캘리포니아주 로스앤젤레스 근처의 라 브레아 타르 웅덩이에서 4,000구가 넘는 다이어울프 화석이 대량으로 발견된 사실에 근거한다. 이 타르 웅덩이는 약 4만 년 전부터 1만 2천 년 전까지 형성된 끈적한 아스팔트 퇴적물로, 함정에 빠진 동식물 표본이 잘 보존되어 과거 생태계를 연구하는 데 중요한 자료를 제공한다. 타르 웅덩이에는 초식동물이 빠지면 그 사체를 노리는 포식자들이 모여들었고, 이 포식자들 역시 함정에 빠지는 경우가 많았다. 발견된 화석의 비율을 보면 초식동물 1마리당 약 10마리의 육식동물이 있었으며, ''Aenocyon dirus guildayi'' (다이어울프의 아종)는 이곳에서 가장 흔하게 발견되는 육식동물로, 그 다음이 스밀로돈이다. 다이어울프 화석은 회색늑대 화석보다 5배나 많이 발견되었다. 이는 많은 수의 다이어울프가 무리를 지어 함께 사냥하거나 먹이를 먹다가 동시에 타르 구덩이에 빠졌을 가능성을 시사한다. 이러한 사회성은 오늘날의 아프리카 야생 개, 승냥이, 회색늑대처럼 알파 암컷이 이끄는 친족 무리 형태였을 것으로 추정된다.
다이어울프는 강력한 포식자였을 것으로 보인다. 현생 회색늑대보다 29% 더 강한 치악력을 가졌으며, 이를 이용해 먹이를 붙잡고 제압했을 것이다. 큰 몸집과 튼튼한 송곳니는 다이어울프가 무리를 지어 큰 먹이나 거대동물을 주로 사냥했음을 시사한다.[20] 다이어울프의 평균적인 먹이 크기는 300kg 에서 600kg 범위였을 것으로 추정된다.
동위원소 분석을 통해 다이어울프의 식단을 추정할 수 있다. 라 브레아 타르 웅덩이에서 발견된 다이어울프, ''스밀로돈'', 아메리카 사자는 비슷한 먹이를 두고 경쟁했으며, 주된 먹이로는 멸종된 낙타 ''카멜롭스 헤스테르누스'', 멸종된 들소 ''바이슨 안티쿠스'', 난쟁이 영양, 말 ''에쿠스 옥시덴탈리스'', 할란땅늘보 등이 있었다. 약 1만 년 전의 다이어울프 화석 분석 결과, 당시에는 말이 중요한 먹이였고 나무늘보, 마스토돈, 들소, 낙타는 덜 흔한 먹이였던 것으로 나타났다. 이는 다이어울프가 특정 먹이만 고집하는 전문 사냥꾼은 아니었으며, 후기 플라이스토세 말기에는 주변에서 가장 쉽게 구할 수 있는 초식동물을 사냥하거나 그 사체를 먹었음을 보여준다. 또한, 다이어울프는 반추동물을 선호했으며, 먹이가 부족할 때는 다른 먹이로 전환하거나 해안가에 떠밀려온 고래 사체를 먹기도 했다.
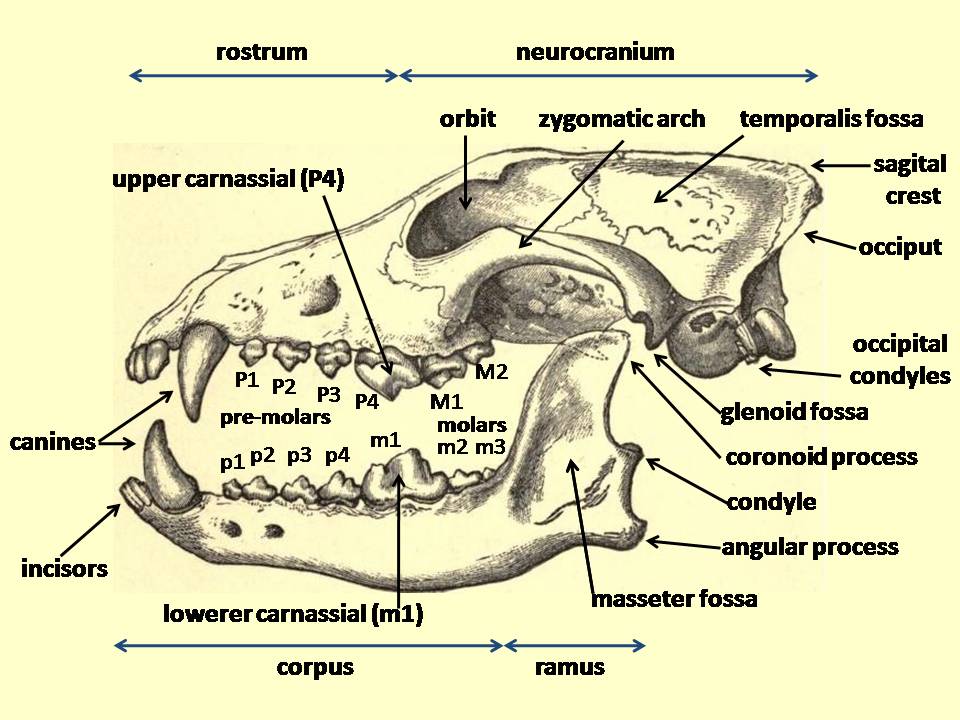
다이어울프의 두개골과 치아는 사냥 방식에 대한 단서를 제공한다. 두개골 형태는 대부분 현대 회색늑대와 유사했지만, 위쪽 어금니(P4)가 더 크고 날카로워 절단 능력이 뛰어났다. 또한, 턱 근육(특히 측두근)이 더 발달하여 회색늑대보다 강한 무는 힘을 낼 수 있었다. 송곳니는 크기가 비슷한 현생 갯과 동물보다 더 튼튼하고 둥근 형태를 가져 먹이와 싸우는 데 유리했다. 아래턱 구조 연구에 따르면, 다이어울프는 무리 지어 사냥하는 동물들처럼 여러 번 얕게 무는 방식을 사용했을 가능성이 높다. 뼈를 부수는 능력에 대해서는 논란이 있는데, 일부 연구에서는 치아 파절 흔적을 근거로 뼈를 먹었을 가능성을 제기하지만, 치아와 두개골 형태 분석 결과 뼈를 전문적으로 부수는 동물은 아니었을 것으로 보인다.[15] 회색늑대처럼 어금니로 골수를 먹었을 수 있지만, 뼈를 부수는 능력은 회색늑대만큼 발달하지 않았을 수 있다.
| 치아 변수 | lupus 현대 북미 | lupus 라 브레아 | lupus 베링기아 | dirus dirus 상가모니안 시대 (125,000–75,000 YBP) | dirus dirus 후기 위스콘신(50,000 YBP) | dirus guildayi (40,000–13,000 YBP) |
|---|---|---|---|---|---|---|
| m1 길이 | 28.2 | 28.9 | 29.6 | 36.1 | 35.2 | 33.3 |
| m1 너비 | 10.7 | 11.3 | 11.1 | 14.1 | 13.4 | 13.3 |
| m1 삼각두 길이 | 19.6 | 21.9 | 20.9 | 24.5 | 24.0 | 24.4 |
| p4 길이 | 15.4 | 16.6 | 16.5 | 16.7 | 16.0 | 19.9 |
| p4 너비 | - | - | - | 10.1 | 9.6 | 10.3 |
| p2 길이 | - | - | - | 15.7 | 14.8 | 15.7 |
| p2 너비 | - | - | - | 7.1 | 6.7 | 7.4 |

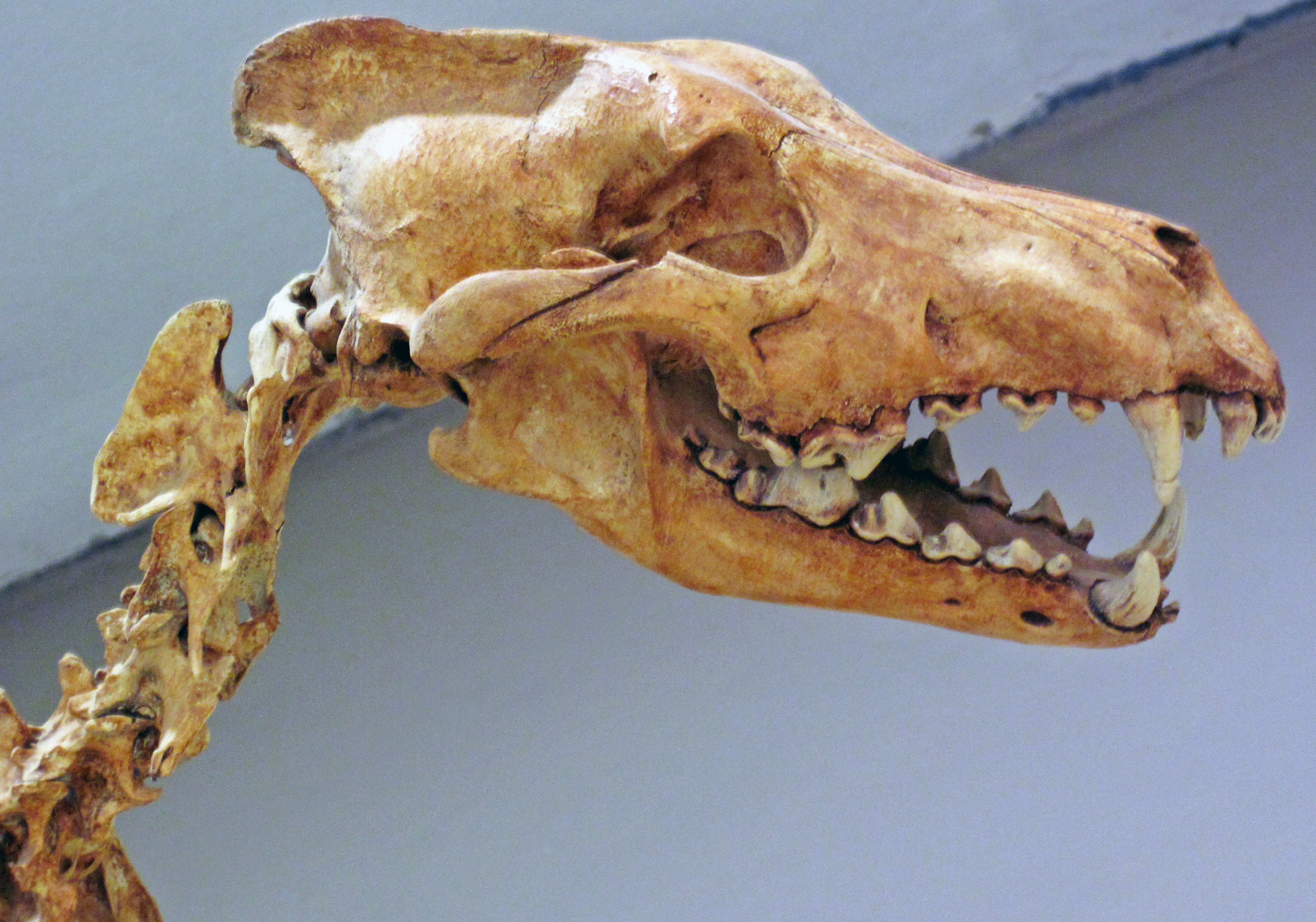
다이어울프 화석에서는 치아 파절(부러짐)이 현생 육식동물보다 훨씬 높은 빈도로 관찰된다. 라 브레아 타르 웅덩이의 다이어울프 화석 연구 결과, 치아 파절률이 5~17%에 달했는데, 이는 현생 포식자들의 0.5~2.7%보다 훨씬 높은 수치이다. 특히 앞니가 자주 부러졌는데, 이는 먹이를 먹을 때 뼈에 더 가깝게 앞니를 사용했음을 시사한다. 높은 치아 파절률의 원인으로는 먹이 가용성이 낮아짐에 따른 육식동물 간의 경쟁 심화가 지목된다. 경쟁이 심해지면 더 빨리 먹이를 먹어야 하고, 이 과정에서 뼈까지 섭취하게 되어 치아가 부러질 위험이 높아진다는 것이다. 실제로 라 브레아의 15,000년 전 다이어울프 화석은 13,000년 전 화석보다 치아 파절률이 3배나 높았는데, 이는 시간이 지나면서 먹이 경쟁이 완화되었음을 시사한다. 하지만 다른 연구에서는 치아 미세 마모 분석 결과, 멸종 직전의 다이어울프가 특별히 먹이 스트레스를 받지는 않았으며 사체 활용도도 현대 육식동물보다 낮았다고 주장하며, 치아 파절이 주로 사냥 행동이나 먹이의 크기와 관련이 있다고 보기도 한다.
다이어울프는 성적 이형성(암수 간의 외형 차이)이 거의 나타나지 않았다. 두개골 길이, 송곳니 크기, 아래 어금니 길이 등을 분석한 결과, 현생 회색늑대와 유사하게 암수 간의 크기 차이가 미미했다. 이는 다이어울프가 일부일처제 관계를 유지하며 생활했을 가능성을 시사한다.[16]
4. 3. 먹이
다이어울프는 현생 회색늑대의 129%에 달하는 강력한 치악력을 가지고 있었으며, 이를 통해 먹이를 붙잡고 억누를 수 있었다. 거대한 몸집과 송곳니로 미루어 볼 때, 다이어울프는 무리 지어 다니는 큰 먹이나 거대동물을 주로 사냥했을 것으로 추정된다. 특정 먹이에만 특화된 사냥꾼은 아니었으며, 당시 풍부했던 대형 동물이면 무엇이든 사냥 대상으로 삼았다.[20] 다이어울프는 대형의, 저항하는 먹이를 사냥하도록 적응된 두개골과 치아를 가진 초육식동물이었다.
동위원소 분석을 통해 라 브레아 지역 다이어울프의 식단을 추정한 결과, 멸종된 낙타인 ''카멜롭스 헤스테르누스'', 멸종된 들소인 ''바이슨 안티쿠스'', "난쟁이" 영양인 ''카프로메릭스 마이너'', 말과의 일종인 ''에쿠스 옥시덴탈리스'', 그리고 북아메리카 초원에 서식했던 할란땅늘보 등을 주로 먹었던 것으로 밝혀졌다. 이들 먹이는 다이어울프뿐만 아니라 검치호(''스밀로돈'')와 아메리카사자(''Panthera atrox'')와도 경쟁 관계에 있었다. 콜럼비아 매머드와 아메리카 마스토돈은 라 브레아 지역에서는 상대적으로 드문 먹이였다.
약 1만 년 전 라 브레아 다이어울프 화석의 동위원소 데이터를 분석한 결과, 당시에는 말이 중요한 먹이였고, 나무늘보, 마스토돈, 들소, 낙타는 식단에서 차지하는 비중이 상대적으로 적었다. 이는 다이어울프가 먹이 전문가는 아니었으며, 플라이스토세 말기에는 가장 쉽게 구할 수 있는 초식동물을 사냥하거나 그 사체를 먹었음을 시사한다. 세드랄, 산 루이스 포토시에서 발견된 표본 연구에 따르면, 다이어울프는 주로 C4 식물을 섭취하는 초식동물과 혼합 식단 초식동물을 먹이로 삼았다.[5] 또한, 다이어울프 뼈의 안정 동위원소 분석 결과, 다른 초식동물보다 바이슨과 같은 반추동물을 선호했으며, 먹이가 부족할 때는 다른 먹이로 전환했고, 때로는 태평양 연안으로 떠밀려온 고래 사체를 먹기도 했다.
다이어울프는 현생 개과 동물 중에서도 높은 수준의 치악력을 가졌다. 살아있는 포유류와 화석 포식자의 송곳니 치악력을 체중 대비로 비교한 연구에 따르면, 다이어울프(163 N/kg)는 조사된 태반류 포유류 중 가장 높은 수치를 기록했다. 이는 현존하는 초육식성 개과 동물인 아프리카들개 (142 N/kg), 회색늑대 (136 N/kg), 승냥이 (112 N/kg), 딩고 (108 N/kg)보다 높은 값이다. 열육치에서의 치악력도 유사한 경향을 보였다. 이러한 강력한 치악력은 다이어울프가 비교적 큰 동물을 사냥했음을 시사한다.
다이어울프의 두개골과 치아는 회색늑대와 유사한 점도 많지만, 몇 가지 중요한 차이점이 있다. 위쪽 어금니 (P4)는 회색늑대보다 크고 무거운 날을 가져 절단 능력이 뛰어났으며, 상대적으로 넓고 큰 측두근 덕분에 더 강한 물어뜯는 힘을 낼 수 있었다. 아래쪽 앞어금니 (m1) 역시 회색늑대보다 훨씬 크고 절단 능력이 뛰어났다. 다이어울프의 송곳니는 크기가 비슷한 현생 개과 동물의 송곳니보다 굽힘 강도가 더 컸는데, 이는 하이에나나 고양이과 동물의 송곳니와 유사한 특징이다. 이러한 특징들은 다이어울프가 회색늑대보다 더 강한 힘으로 물 수 있었고, 먹이와 싸우는 데 더 잘 적응했음을 보여준다.
| 치아 변수 | lupus 현대 북미 | lupus 라 브레아 | lupus 베링기아 | dirus dirus 상가모니안 시대 (125,000–75,000 년 전) | dirus dirus 후기 위스콘신 빙기(50,000 년 전) | dirus guildayi (40,000–13,000 년 전) |
|---|---|---|---|---|---|---|
| m1 길이 | 28.2 | 28.9 | 29.6 | 36.1 | 35.2 | 33.3 |
| m1 너비 | 10.7 | 11.3 | 11.1 | 14.1 | 13.4 | 13.3 |
| m1 삼각두 길이 | 19.6 | 21.9 | 20.9 | 24.5 | 24.0 | 24.4 |
| p4 길이 | 15.4 | 16.6 | 16.5 | 16.7 | 16.0 | 19.9 |
| p4 너비 | - | - | - | 10.1 | 9.6 | 10.3 |
| p2 길이 | - | - | - | 15.7 | 14.8 | 15.7 |
| p2 너비 | - | - | - | 7.1 | 6.7 | 7.4 |
라 브레아 타르 웅덩이에서 발견된 다수의 다이어울프 화석은 이들이 사회적인 동물이었음을 시사한다. 타르 웅덩이에 빠진 초식동물 사체에 이끌려왔다가 함께 갇힌 것으로 추정되며, 이는 다이어울프가 무리를 지어 사냥하고 먹이를 먹었을 가능성을 높인다. 다이어울프의 큰 몸집과 사회적 습성을 고려할 때, 먹이 크기는 300kg에서 600kg 범위였을 것으로 추정된다. 이는 현생 늑대 무리가 500kg 정도의 무스를 사냥하는 것과 비교될 수 있다.
다이어울프의 치아 파절률은 과거 식습관과 생존 환경에 대한 단서를 제공한다. 라 브레아에서 발견된 다이어울프 화석의 치아 파절률은 현생 육식동물보다 훨씬 높았는데, 특히 앞니의 파절률이 현대 회색늑대보다 높았다. 이는 다이어울프가 먹이를 먹을 때 뼈에 더 가깝게 앞니를 사용했음을 시사한다. 높은 치아 파절률은 뼈를 자주 섭취했거나, 먹이 경쟁이 심해 사체를 급하게 먹어야 했던 상황을 반영할 수 있다. 일부 연구에서는 이를 근거로 다이어울프가 뼈를 으깨 먹는 습성이 있었고, 시체 청소부였을 가능성을 제기하기도 했다.
그러나 다이어울프가 뼈를 부수는 데 특화된 동물이었는지에 대해서는 논란이 있다. 아래턱 구조 분석 결과, 다이어울프의 아래턱 결합부는 회색늑대보다 약했으며, 이는 뼈를 부수는 능력보다는 무리 지어 사냥하며 여러 번 얕게 무는 방식에 더 적합했음을 시사한다. 또한, 후기 플라이스토세로 갈수록 다이어울프의 치아 파절률이 감소하는 경향이 나타났는데, 이는 먹이 경쟁이 완화되면서 뼈 섭취량이 줄어드는, 다이어울프에게 더 적합한 먹이 섭취 행동으로 돌아갔기 때문일 수 있다. 최근의 치아 미세 마모 연구에서는 멸종 직전의 다이어울프가 심각한 먹이 스트레스를 겪지 않았으며, 사체 활용도(뼈 섭취 포함)가 오늘날의 대형 육식동물보다 낮았다고 분석했다. 이는 높은 치아 파절률이 반드시 뼈를 주식으로 삼았다는 의미는 아니며, 사냥 행동이나 먹이 크기와 더 관련이 있을 수 있음을 시사한다.
플라이스토세 말기의 기후 변화는 다이어울프의 먹이 환경과 신체적 특징에도 영향을 미쳤다. 라 브레아 지역의 다이어울프 화석을 시기별로 분석한 결과, 기후 변동과 맞물려 다이어울프의 몸 크기, 두개골 및 주둥이 모양, 치아 마모 및 파절 정도가 변화했음이 밝혀졌다. 특히 약 17,900년 전과 같이 기후가 급격하게 변동하며 건조해졌던 시기에는 몸집이 작아지고, 두개골 기저부가 넓어지며 주둥이가 짧아지는 등 식량 스트레스의 증거가 나타났다. 이러한 변화는 먹이 부족 상황에서 사체를 더 완전히 소비하고 뼈를 부수기 위해 더 강한 물기 힘을 내도록 적응하는 과정에서 나타났을 수 있다.
:
| 변수 | 28,000 년 전 | 26,100 년 전 | 17,900 년 전 | 13,800 년 전 |
|---|---|---|---|---|
| 체구 | 가장 큼 | 큼 | 가장 작음 | 중간/작음 |
| 치아 파절 | 높음 | 낮음 | 높음 | 낮음 |
| 치아 마모 | 높음 | 낮음 | 높음 | 낮음 |
| 주둥이 모양 | 짧아짐, 가장 큰 두개골 기저부 | 평균 | 가장 짧음, 가장 큰 두개골 기저부 | 평균 |
| 치열 모양 | 튼튼함 | – | – | 가냘픔 |
| DO 사건 | 3 또는 4번 | 없음 | 부정확한 데이터 | 부정확한 데이터 |
4. 4. 경쟁

다이어울프가 북아메리카에 나타나기 직전, 아시아의 댕견과 아프리카들개의 조상인 ''Xenocyon'' 아속의 개가 먼저 들어왔다. 하지만 화석 기록상 이들은 드물었고, 새로 등장한 다이어울프와의 경쟁에서 밀려난 것으로 보인다.
다이어울프는 당시 북아메리카의 다른 여러 대형 포식자들과 먹이를 두고 경쟁해야 했다. 안정 동위 원소 분석 결과, 다이어울프는 검치호(Smilodon fatalis) 및 아메리카사자(Panthera atrox)와 같은 먹이를 사냥하며 직접적으로 경쟁했던 것으로 밝혀졌다. 또한 다이어울프는 스밀로돈이 먹다 남긴 사체를 이용했을 가능성도 제기된다.[15] 이 외에도 거대한 몸집의 단두곰(Arctodus simus), 현생 퓨마(Puma concolor), 플라이스토세 코요테(Canis latrans), 그리고 오늘날의 회색늑대보다 더 크고 강건했던 플라이스토세 회색늑대(Canis lupus) 등 다양한 포식자들이 공존했다. 또한, 이들 포식자는 비슷한 먹이를 사냥했던 인간과도 경쟁했을 가능성이 있다.
특히, 플라이스토세 후기에는 베링 육교 지역에 서식하던 베링 늑대(Canis lupus)가 로렌타이드 빙상과 코딜레란 빙상 사이의 통로를 통해 남쪽으로 이동했을 수 있다. 와이오밍주 빅혼 산맥의 내추럴 트랩 동굴(Natural Trap Cave)에서 발견된 25,800년에서 14,300년 전 사이의 늑대 화석이 이를 뒷받침한다. 베링 늑대는 거대 동물 사냥에 특화되어 있었으며, 다이어울프의 직접적인 경쟁자였다. 다이어울프 화석은 북위 42도선 이북에서는 발견되지 않는데, 이 지역은 베링 늑대가 남하하여 활동했을 가능성이 있는 영역이다. 베링 늑대 역시 다이어울프처럼 플라이스토세 말기에 멸종했다.
한편, 동유라시아로 진출한 다이어울프는 해당 지역의 지배적인 포식자였던 동굴하이에나(Crocuta crocuta ultima)와 경쟁했을 것으로 추정된다. 강력한 동굴하이에나와의 경쟁 때문에 유라시아 지역 다이어울프의 개체 수가 매우 적었고, 이것이 다른 화석 유적지에서 다이어울프 화석이 드물게 발견되는 이유일 수 있다.
5. 멸종
다이어울프는 플라이스토세 말기에 북아메리카와 남아메리카의 다른 많은 거대동물들과 함께 사라졌다. 이는 약 12,700년 전 시작된 제4기 대량멸종 사건의 일부로, 당시 44kg 이상의 무게를 가진 포유류 90개 속이 멸종했다.
멸종의 정확한 원인은 여전히 논의 중이지만, 단일 원인보다는 여러 요인이 복합적으로 작용한 결과로 여겨진다. 주요 원인으로는 다이어울프가 의존했던 거대 초식동물의 멸종, 마지막 빙하기 이후의 급격한 기후 변화, 그리고 회색늑대나 새로 등장한 인류와의 경쟁 등이 복합적으로 거론된다. 최근의 유전학 및 형태학 연구는 다이어울프가 회색늑대와는 다른 독자적인 진화 경로를 걸었으며, 이로 인해 변화하는 환경과 새로운 경쟁자들에 효과적으로 적응하지 못했을 가능성을 시사한다.
5. 1. 멸종 시기
다이어울프는 플라이스토세 거대동물 중 가장 번성했던 종 중 하나였으나, 약 10,000년 전 다른 대부분의 북아메리카 거대동물들과 함께 멸종했다.[24][25] 이는 제4기 대량멸종 시기와 일치한다.
플라이스토세 후기(약 300,000년 전)에 회색늑대(''Canis lupus'')가 베링 육교를 건너 북아메리카로 들어오면서 다이어울프와 경쟁하기 시작했다. 약 16,000년 전 마지막 빙기가 끝나고 인간이 북아메리카 대륙에 도착하면서, 다이어울프의 주요 먹이였던 거대 포유류들이 사라지기 시작했다. 이는 기후 변화나 인간의 과도한 착취 때문일 수 있다.[26]
회색늑대나 아메리카붉은늑대보다 몸집이 크고 느렸던 다이어울프는 빨라진 먹잇감을 사냥하기 어려워졌고, 점차 죽은 동물의 사체를 먹는 청소부 역할에 의존하게 된 것으로 보인다. 결국 약 10,000년 전, 다이어울프를 포함한 많은 거대 포유류가 멸종했다.
멸종의 정확한 원인은 여전히 논란 중이며, 여러 요인이 복합적으로 작용했을 가능성이 크다. 주요 가설은 다음과 같다.
- 거대동물 먹이 감소: 다이어울프가 의존했던 거대 초식동물의 멸종. 약 12,700년 전 제4기 대량멸종 동안 44kg 이상 무게의 포유류 90개 속이 사라졌다.
- 기후 변화: 마지막 빙기 이후의 급격한 환경 변화.
- 경쟁: 회색늑대와의 경쟁 심화 및 새로 등장한 인간과의 경쟁. 다이어울프보다 작고 날렵했던 회색늑대가 변화된 환경에 더 잘 적응했을 수 있다.
- 생식적 고립: 다이어울프는 코요테, 딩고, 회색늑대 등 다른 개과 동물과 달리 유라시아에서 건너온 종들과 이종교배가 불가능했을 수 있다. 이로 인해 다른 개과 동물들이 교배를 통해 획득했을 수 있는 질병 저항성 등을 얻지 못했을 가능성이 제기된다.
- 근친 교배 가능성: 라 브레아 타르 구덩이에서 발견된 다이어울프와 검치호 표본에서 골연골증 박리와 유사한 관절 질환이 높은 비율로 발견되었다. 이는 멸종 직전 개체군 크기가 줄어들면서 근친 교배가 늘어났을 가능성을 시사하지만, 추가 연구가 필요하다.[9]
다이어울프의 멸종 시기를 보여주는 가장 최근의 화석 기록은 미주리주 분 카운티의 브륀율프슨 동굴에서 발견된 것으로, 방사성 탄소 연대 측정 결과 약 9,440년 전(YBP)으로 나타났다. 캘리포니아주 랜초 라 브레아에서는 약 9,860년 전(YBP), 라 미라다에서는 약 10,690년 전(YBP)의 유해가 발견되었다. 애리조나주 화이트워터 드로우에서는 약 8,200년 전(YBP)으로 측정된 유해가 있지만, 뼈 탄산염 연대 측정의 신뢰성에 대한 지적도 있다. 이 연대들은 보정되지 않은 값이므로 실제 연대는 더 오래되었을 수 있다. 남아메리카에서는 페루 탈라라에서 약 9,030 ± 240년 전(YBP, 보정되지 않음), 아르헨티나 루한 지역에서는 약 10,000~11,000년 전보다 오래된 층서 단면에서 유해가 발견되었다.[10]
다이어울프의 멸종 과정을 정확히 이해하기 위해서는 더 많은 화석 표본의 연대 측정과 함께, 당시 생물지리학적 분포, 개체 수 변화에 영향을 미친 요인들(다른 종과의 경쟁, 포식-피식 관계, 환경 변화 등)에 대한 종합적인 연구가 필요하다. 특히 다이어울프와 밀접하게 상호작용했던 다른 거대동물들의 정확한 멸종 시기를 밝히는 것이 중요하다.
5. 2. 멸종 원인
다이어울프는 플라이스토세 거대동물 중 가장 번성했던 종 중 하나였으나, 약 10,000년 전쯤 다른 대부분의 북아메리카 거대동물들과 함께 멸종했다.[24][25] 멸종의 정확한 원인은 밝혀지지 않았지만, 여러 요인이 복합적으로 작용한 것으로 추정된다.
플라이스토세 후기(약 300,000년 전)에 회색늑대(''Canis lupus'')가 베링 육교를 건너 북아메리카로 들어오면서 다이어울프와의 경쟁이 시작되었다. 약 16,000년 전, 마지막 빙기가 끝나갈 무렵 인류가 북아메리카 대륙에 도달했고, 이 시기부터 다이어울프의 주요 먹이였던 거대 포유류들이 사라지기 시작했다(제4기 대량멸종). 이는 기후 변화나 인간의 사냥 활동 때문일 수 있다.[26]
다이어울프는 회색늑대나 붉은늑대 등에 비해 몸집이 크고 다리가 짧아 상대적으로 느렸기 때문에, 변화된 환경에서 재빠른 동물들을 사냥하기 어려웠을 것으로 보인다. 점차 사냥보다는 죽은 동물의 사체를 먹는 청소부 역할에 의존하게 되었을 가능성이 있다. 결국 약 10,000년 전, 다이어울프를 포함한 많은 거대 포유류들이 멸종에 이르렀다.
약 12,700년 전 시작된 제4기 대멸종 기간 동안, 몸무게 44kg 이상의 포유류 90개 속이 사라졌다. 대형 육식동물과 청소 동물들의 멸종은 이들이 의존하던 거대 초식동물들의 멸종과 밀접한 관련이 있는 것으로 여겨진다. 거대동물군 멸종의 원인에 대해서는 여전히 논란이 있지만, 기후 변화, 다른 종과의 경쟁, 그리고 새로 등장한 인간 사냥꾼에 의한 과도한 착취 또는 이 요인들의 복합적인 작용 등이 주요 가설로 제시된다.
고대 DNA 분석 결과는 다이어울프가 유라시아에서 건너온 회색늑대, 코요테, 딩고 등 다른 개과 동물들과 유전적으로 상당히 멀리 떨어져 있었으며, 이들 사이의 교배가 불가능했을 가능성을 시사한다. 이러한 생식적 고립 때문에 다이어울프는 회색늑대나 코요테처럼 다른 개과 동물과의 교배를 통해 새로운 환경에 적응하거나 질병에 대한 저항성을 얻기 어려웠을 수 있다. 반면, 북미에서는 다이어울프와 베링기안 늑대가 멸종하고, 상대적으로 덜 육식적이고 몸집이 작은 형태의 늑대(현대 회색늑대의 조상)만이 살아남아 번성했다.
또한, 2023년 연구에서는 랜초 라 브레아 타르 구덩이에서 발견된 다이어울프와 검치호 표본들의 관절 표면에서 높은 비율로 골연골증 박리와 유사한 연골하 결손이 발견되었다. 연구자들은 이것이 현대의 개들에서 근친 교배와 연관되는 것처럼, 멸종 위기에 처했던 다이어울프 개체군 내에서도 근친 교배가 만연했을 가능성을 제기했다. 다만, 이것이 아메리카 대륙 전역의 다이어울프에게 해당되는 현상인지는 추가 연구가 필요하다고 덧붙였다.[9]
다이어울프 화석 중 가장 젊은 연대는 미주리주 분 카운티의 브륀율프슨 동굴에서 발견된 것으로, 약 9,440년 전으로 측정되었다. 캘리포니아주 랜초 라 브레아에서는 약 9,860년 전, 라 미라다에서는 약 10,690년 전의 화석이 발견되었다. 애리조나주 화이트워터 드로우에서는 약 8,200년 전으로 측정된 화석도 있지만, 뼈 탄산염을 이용한 방사성 탄소 연대 측정의 신뢰성에 의문을 제기하는 견해도 있다. 이 연대들은 모두 보정되지 않은 값이므로 실제 연대는 더 오래되었을 수 있다. 남아메리카에서는 페루 탈라라에서 가장 최근의 화석이 약 9,030 ± 240년 전(보정되지 않음)으로 측정되었고, 아르헨티나 루한에서 발견된 ''"C. nehringi"'' (다이어울프의 한 형태로 추정됨)의 가장 최근 화석은 약 10,000~11,000년 전의 층서 단면보다 오래된 것으로 나타났다.[10]
다이어울프의 정확한 멸종 원인을 이해하기 위해서는 더 많은 화석 표본의 정밀한 연대 측정과 함께, 당시의 생물지리학적 분포, 개체군 크기에 영향을 미쳤던 요인들(다른 종과의 경쟁, 포식-피식 관계, 환경 변화 등)에 대한 종합적인 연구가 필요하다. 특히 다이어울프와 밀접하게 상호작용했던 다른 거대동물들의 멸종 시기를 명확히 밝히는 것이 중요하다.
참조
[1]
논문
Notas sobre algunos mamíferos fósiles nuevos ó poco conocidos del valle de Tarija
1902
[2]
문서
[3]
논문
A late Pleistocene fossil from Northeastern China is the first record of the dire wolf (Carnivora: Canis dirus) in Eurasia.
https://linkinghub.e[...]
2020-10
[4]
그림
Rancho la Brea. Restoration by Chas. R. Knight. Mural for Amer. Museum Hall of Man. Coast Range in background, Old Baldy at left
http://geology.cwru.[...]
[5]
논문
Diet and habitat of mesomammals and megamammals from Cedral, San Luis Potosí, México
https://www.cambridg[...]
2024-10-22
[6]
서적
The fauna of Rancho La Brea
https://archive.org/[...]
University of California – Berkeley
[7]
논문
First record of cf. ''Aenocyon dirus'' (Leidy, 1858) (Carnivora, Canidae), from the Upper Pleistocene of the Atacama Desert, northern Chile
[8]
논문
Canids (Caninae) from the Past of Venezuela
https://bioone.org/j[...]
[9]
논문
Subchondral defects resembling osteochondrosis dissecans in joint surfaces of the extinct saber-toothed cat ''Smilodon fatalis'' and dire wolf ''Aenocyon dirus''
2023
[10]
논문
"Stratigraphic range of the large canids (Carnivora, Canidae) in South America, and its relevance to quaternary biostratigraphy"
[11]
뉴스
絶滅オオカミ「ダイアウルフ」、実はオオカミと遠縁だった
https://natgeo.nikke[...]
2021-01-16
[12]
웹사이트
The legendary dire wolf may not have been a wolf at all
https://www.sciencem[...]
2021-01-24
[13]
논문
Dire wolves were the last of an ancient New World canid lineage
https://www.nature.c[...]
2021-01-13
[14]
문서
A late Pleistocene fossil from Northeastern China is the first record of the dire wolf (Carnivora: Canis dirus) in Eurasia
[15]
논문
"Craniofacial morphology and feeding behavior in Canis dirus, the extinct Pleistocene dire wolf"
2006
[16]
논문
"Sexual dimorphism, social behavior, and intrasexual competition in large Pleistocene carnivorans"
2002
[17]
문서
日本産後期更新世の巨大狼化石
http://www.gmnh.pref[...]
群馬県立自然史博物館研究報告
2020
[18]
웹인용
Wolves, Coyotes, and Dogs (Genus Canis)
http://www.museum.st[...]
Museum.state.il.us
2011-10-23
[19]
웹인용
San Diego Natural History Museum Fossil Mysteries Field Guide: Dire Wolf
http://www.sdnhm.org[...]
Sdnhm.org
2011-10-23
[20]
웹인용
Dire Wolf Fact Sheet
http://library.sandi[...]
San Diego Zoo
2012-10-31
[21]
논문
Dire Wolf, Canis dirus (mammalia; Carnivora; Canidae), from the Late Pleistocene (Rancholabrean) of East-Central Sonora, Mexico
http://www.academia.[...]
2012-10-31
[22]
웹인용
Wolves, Coyotes, and Dogs (Genus Canis)
http://www.museum.st[...]
Illinois State Museum
2010-06-21
[23]
서적
Walker's Mammals of the World
Maryland, Johns Hopkins University Press
[24]
서적
Dire Wolf
http://books.google.[...]
2009-09
[25]
서적
American Megafaunal Extinctions at the End of the Pleistocene
http://www.springerl[...]
2013-08-13
[26]
DVD
Prehistoric Predators
National Geographic
2008-02-12
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com