돌말류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
돌말류는 클로로필을 함유한 단세포 조류로, 다양한 해양 및 담수 환경에 널리 분포하며, 규산질로 이루어진 독특한 세포벽(프러스툴)을 가진다. 1703년 최초의 삽화가 발견된 이후, 찰스 다윈의 연구를 통해 아름다움과 기능적 특징이 주목받았으며, 현재는 해양 생태계 연구와 환경 변화 연구에 활용된다. 돌말류는 중심 규조류와 깃털 규조류로 나뉘며, 무성 생식과 유성 생식을 통해 번식한다. 이들은 해양에서 규소와 탄소 순환에 중요한 역할을 하며, 규조토로 불리는 껍질은 산업적으로, 나노기술 재료로 활용된다. 또한 법의학적 증거로도 사용되며, 한국에서도 활발한 연구가 진행되고 있다.
더 읽어볼만한 페이지
- 돌말류 - 위배선규조류
위배선규조류는 김발돌말목, 민부채돌말목 등을 포함하는 조류 분류군이다. - 돌말류 - 체돌말류
체돌말류는 체돌말강을 포함하는 분류군으로, 조류 정보 사이트의 분류 체계에 따라 Archaegladiopsophycidae, 수염돌말아강, 체돌말아강으로 나뉜다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.
| 돌말류 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 분류 | |
| 학명 | Bacillariophyceae |
| 명명자 | Dangeard, 1933 |
| 이명 | Diatomea Dumortier, 1821 Diatomophyceae Rabenhorst, 1864 Bacillariae Haeckel, 1878 Bacillariophyta Engler & Gilg, 1919 |
| 하위 분류군 | Bacillariophycidae Coscinodiscophycidae Fragilariophycidae |
| 생물학적 특징 | |
| 화석 기록 | 쥐라기 - 현재 |
| 어원 | 그리스어 διατομή (diatomé, "절단, 분리")에서 유래 |
| 서식지 | 해양, 수로, 토양 |
| 역할 | 규조류는 전 세계 산소 생산량의 약 20%를 담당한다. |
| 추가 정보 | |
| 관련 링크 | https://www.livescience.com/46250-teasing-apart-the-diatom-genome.html https://diatoms.org/what-are-diatoms https://www.kcl.ac.uk/sspp/departments/geography/people/academic/drake/Research/The-Sahara-Megalakes-Project/Lake-Megachad.aspx https://www.smithsonianmag.com/science-nature/gas-guzzlers-1-106582382/ http://www.ucmp.berkeley.edu/chromista/diatoms/diatommm.html https://blogs.nasa.gov/earthexpeditions/2018/09/04/the-inner-space-of-the-subarctic-pacific-ocean/ |
2. 역사
1703년, 영국의 한 익명의 학자가 왕립학회 회보에 규조류 ''Tabellaria''에 대한 최초의 그림을 게재하면서 규조류의 존재가 처음으로 알려졌다.[166]
규조류 세포는 다양한 소기관과 독특한 구조를 가지고 있다. 대부분의 규조류는 황갈색 엽록체를 가지고 있는데, 이 엽록체는 4개의 세포막으로 둘러싸여 있으며 푸코산틴과 같은 생물학적 색소를 포함하는 이형편모조류의 특징을 나타낸다. 엽록체는 광합성이 일어나는 장소이다. 일반적으로 편모는 없지만, 중심 규조류의 수컷 배우자는 편모를 가지며, 이는 다른 이형편모조류와 유사한 구조를 보인다.[31]
3. 세포 구조
규조류는 "바다의 보석" 또는 "살아있는 오팔"이라고 불릴 정도로 광학적 특성이 뛰어나다.[32] 이러한 구조적 착색은 의사소통, 위장, 열 교환, 자외선 보호 등과 관련이 있을 것으로 추정된다.[33]
규조류 세포는 주로 실리카로 구성된 정교하고 단단하지만 다공성인 세포벽인 프러스툴을 가지고 있다.[75] 이 규질 벽[34]은 다양한 기공, 능선, 미세한 가시, 가장자리 융기 및 융기로 패턴화되어 있어 속과 종을 구분하는 데 사용된다.
세포는 두 개의 반쪽, 즉 ''하위 갑''과 ''상위 갑''으로 구성되어 있으며, 각각은 평평한 판(밸브)과 가장자리를 연결하는 거들 밴드를 포함한다. 하위 갑은 상위 갑보다 약간 작다. 규조류 형태는 다양하며, 일반적으로 원형이지만 삼각형, 사각형, 타원형 등 다양한 모양을 가질 수 있다.

규조류는 프러스툴 모양에 따라 '''중심 규조류'''와 '''깃털 규조류''' 두 그룹으로 나뉜다.

3. 1. 세포 내 구조
규조류는 모두 광합성을 하는 독립 영양 생물로, 세포 내에 1개 또는 여러 개의 엽록체를 가지고 있다. 엽록체는 4중막 구조이며, 홍조류에서 유래한 것으로 추정된다. 광합성 색소로는 클로로필 a, c1, c2를 가지며, 보조 색소로는 카로티노이드인 푸코산틴, 디아톡산틴, 디아디노산틴, β-카로틴 등을 포함하여 황갈색을 띤다.[105]
세포핵은 세포 중심 부근에 위치하며, 그 주위를 발달된 액포가 둘러싸고 있다. 미토콘드리아는 관 모양의 크리스테 구조를 가지며, 세포질 내에 분산되어 있다.
규조류는 스트라메노필에 속하지만, 일반적인 세포는 편모가 없고 중심 규조류의 유주 세포에만 편모가 존재한다. 유주 세포의 편모는 깃털 모양이며, 미세소관 배열은 일반적인 9+2 구조가 아닌 9+0 구조이다.
3. 2. 세포 외 구조
규조류는 단세포 생물로서 다채로운 세포 외 기질을 가진다. 규조류 대부분은 껍질에 입술 모양 돌기(labiate process)라는 속이 빈 관을 가지고 있으며, 여기서 점액성 물질을 세포 밖으로 분비한다. 특히 중심 규조류 중 플랑크톤으로 생활하는 것들은 점성이 높은 점액이 유기 돌기(strutted process)를 형성하여 세포에서 방사상으로 뻗어 나와 해양에서 세포의 침강을 억제하고 유광층에 머무르게 한다.
깃털 규조류 일부 종은 껍질에 세로 홈(raphe)을 가지고 있으며, 홈의 축 방향으로 기질 위를 미끄러지듯이 운동한다(1-25μm/s 정도).[38] 이 세로 홈 안쪽의 세포막 바로 아래에는 액틴 필라멘트가 배치되어 있으며, 세로 홈에서 점액 섬유가 분비된다.
고착성이며 군체를 이루는 깃털 규조류는 세포 밖에 한천질을 분비하여 수지상 자루를 형성해 군체를 이루기도 한다.
4. 껍질 (Frustule)
규조류는 주로 실리카로 구성된 정교하고 단단하지만 다공성인 세포벽인 프러스툴을 만든다.[75] 이 규질 벽[34]은 다양한 기공, 능선, 미세한 가시, 가장자리 융기 및 융기로 패턴화될 수 있으며, 이 모든 것을 사용하여 속과 종을 구분할 수 있다.
세포 자체는 두 개의 반쪽으로 구성되어 있으며, 각각 본질적으로 평평한 판 또는 밸브와 가장자리를 연결하는 거들 밴드를 포함한다. 한쪽 반쪽인 '하위 갑'은 다른 쪽 반쪽인 '상위 갑'보다 약간 작다. 규조류 형태는 다양하며, 세포 모양은 일반적으로 원형이지만, 일부 세포는 삼각형, 사각형 또는 타원형일 수 있다.
규조류는 프러스툴의 모양에 따라 ''중심 규조류''와 ''깃털 규조류''의 두 그룹으로 나뉜다. 깃털 규조류는 양쪽 대칭이며, 각 밸브에는 라페를 따라 틈새가 있고, 껍질은 일반적으로 이러한 라페에 평행하게 길쭉하다. 중심 규조류는 방사형 대칭이며, 상위 밸브와 하위 밸브(''상위 갑''과 ''하위 갑'')로 구성되어 있다.
4. 1. 껍질의 구조
규조류의 껍질(프러스툴, Frustule)은 주로 실리카로 구성된 정교하고 단단하지만 다공성인 세포벽이다.[75] 이 규질 벽[34]은 다양한 기공, 능선, 미세한 가시, 가장자리 융기 및 융기로 패턴화될 수 있으며, 이 모든 것을 사용하여 속과 종을 구분할 수 있다.세포 자체는 두 개의 반쪽으로 구성되어 있으며, 각각 본질적으로 평평한 판 또는 밸브와 가장자리를 연결하는 거들 밴드를 포함한다. 한쪽 반쪽인 '하위 갑'은 다른 쪽 반쪽인 '상위 갑'보다 약간 작다. 마치 도시락과 뚜껑처럼, 큰 외피(epitheca) 안에 작은 내피(hypotheca)가 조합된 구조로 되어 있다. 껍질 면(valve face)과 띠 면(girdle face)으로 구분되며, 광학 현미경으로는 주로 껍질 면이 관찰된다.
규조류 형태는 다양하다. 세포의 모양은 일반적으로 원형이지만, 일부 세포는 삼각형, 사각형 또는 타원형일 수 있다. 껍질에는 포문(areola)이라고 불리는 미세한 구멍이나, 그것이 배열된 조선(stria)이 다수 존재하며, 이들의 수와 배치는 분류 형질로 사용된다.
4. 2. 규산 대사 (Silicification)
규조류 세포는 프러술이라고 알려진 독특한 규산질 세포벽 안에 들어 있다.[39] 세포벽을 구성하는 생물기원 규소는 생합성에 의해 세포내에서 중합된 규산 단량체에 의해 합성되며, 이 물질은 세포 외부로 배출되어 벽에 추가된다. 규조류 껍질 형성에는 규산(Si(OH)₄) 섭취가 필수적이다.[40]규조류에서는 예로부터 껍질 형성 기구와 규산 대사 기구에 관한 연구가 진행되어 왔다. 1999년에는 ''Cylindrotheca fusiformis''의 껍질에서 규산을 과립상으로 응집시키는 단백질인 실라핀(Silaffin)이 정제되었다. 또한, 규산의 세포 내 대사와 관련된 유전자도 ''Thalassiosira''나 ''Nitzschia''와 같은 대표적인 속에 의해 다수 보고되고 있다.
껍질 형성에는 규산 섭취가 필수적이지만, 이와 경쟁적으로 섭취되는 이산화게르마늄(GeO₂)을 규조류 배양에 첨가하면 껍질이 형성되지 않을 뿐만 아니라 규조류는 사멸한다. 이산화 게르마늄은 규산을 이용하는 조류의 생육을 상당히 선택적으로 차단하는 작용이 있으므로, 이들의 번식을 원하지 않는 배양을 할 때 위력을 발휘한다.
4. 3. 화석
규조류의 껍질(frustule)은 재질 덕분에 퇴적물 속에서 잘 보존되어 미화석으로 대량 산출된다. 규조류 껍질이 대부분을 차지하는 퇴적물은 규조토라고 불린다.[126] 가장 오래된 규조류 화석은 독일 북부의 1억 8500만 년 전 지층에서 발견되었다.[118]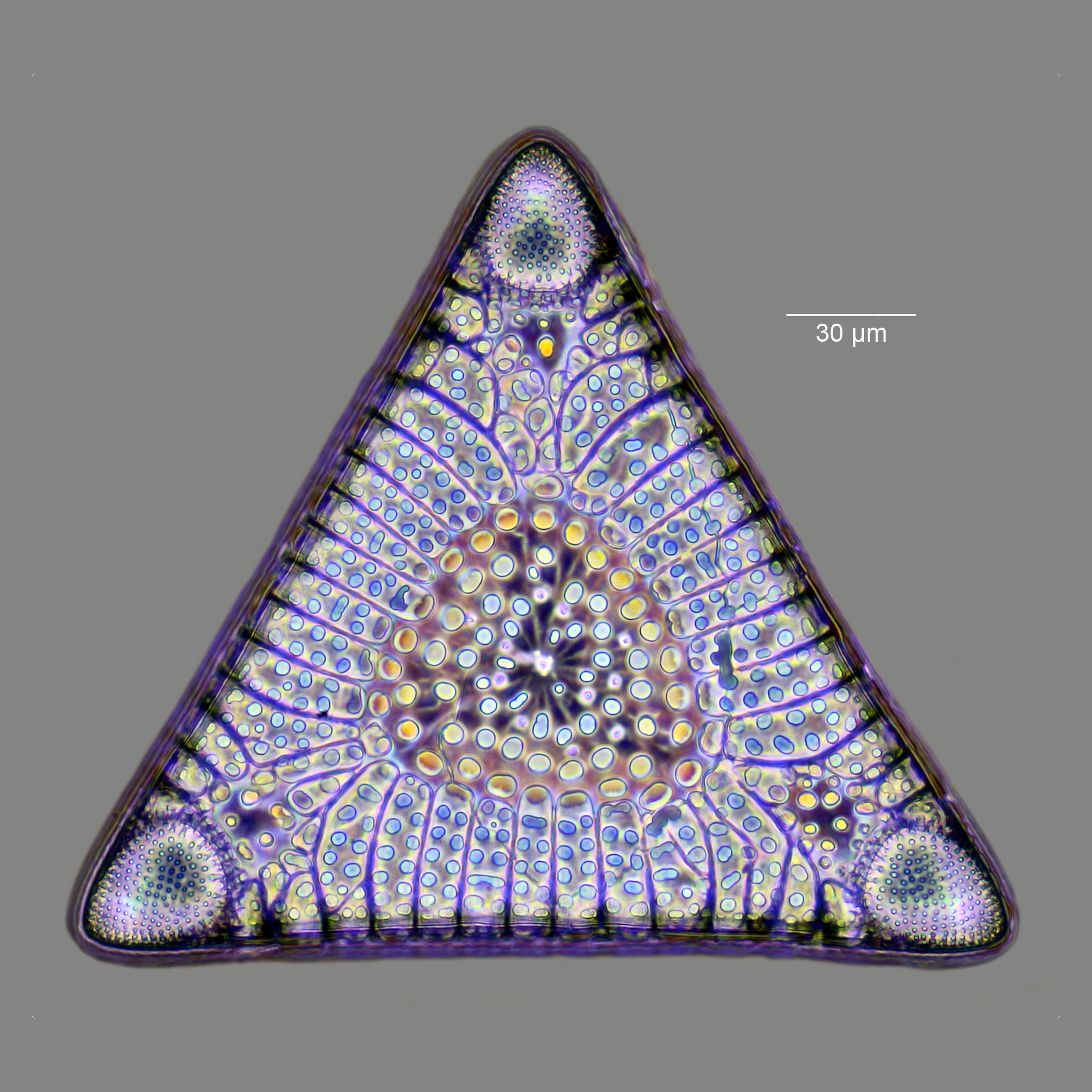
규조류 화석은 고환경 복원 및 지질학적 연대 측정에 중요한 자료로 활용된다. 특히 호수 퇴적물에 보존된 규조류는 수심과 염도의 변동을 겪는 폐쇄 분지 호수의 제4기 기후 고환경 재구성에 널리 사용된다.[126] 해양 규조류의 생물대는 지구 자기 극성 시간 척도에 맞춰 보정된 곳(예: 남극해, 북태평양, 동부 적도 태평양)에서 규조류 기반 연대 추정이 10만 년 이내로 가능하다. 하지만 신생대 규조류 군집의 전형적인 연대 해상도는 수십만 년이다.[126]
고기후학에서 과거 기후를 연구하기 위해 대리 데이터를 사용하는데, 돌말류의 동위 원소 기록인 δ13C, δ18O, δ30Si (δ13C돌말류, δ18O돌말류, 및 δ30Si돌말류)가 고기후 대리 지표로 사용된다.[127]
백악기의 규조류 기록은 제한적이지만, 최근 연구에 따르면 규조류 유형의 점진적인 다양화가 이루어졌다. 백악기-고생대 멸종 사건은 해양 생물에 큰 영향을 미쳤지만, 규조류는 비교적 작은 영향을 받았다. 해양 규조류의 대량 멸종은 신생대 동안 관찰되지 않았지만, 해양 규조류 종 집합체의 비교적 빠른 진화적 전환 시기는 고생대–에오세 경계 부근,[140] 에오세–올리고세 경계에서 발생했다.[141]
5. 생식
돌말류는 무성생식과 유성생식 두 가지 방법으로 번식한다. 무성생식은 주로 분열법을 통해 이루어진다. 세포의 위·아래 껍데기가 갈라지면서 핵과 원형질이 두 개로 나뉘고, 각각의 껍데기 안쪽에 새로운 껍데기를 만들어 두 개의 새로운 개체가 생성된다.
유성생식에는 두 가지 방법이 있다. 하나는 증대 포자를 형성하는 것이다. 분열을 계속하면 세포 크기가 작아지는데, 접합을 통해 증대 포자를 만들어 크기를 회복한다. 이러한 생식법은 우상규조류에서 흔히 볼 수 있다. 다른 하나는 정자와 난자가 합쳐지는 방법이다. 정자는 겉보기에는 1개의 편모를 가진 것처럼 보이지만, 실제로는 길고 짧은 2개의 편모를 가지고 있다. 몸 밖으로 나온 긴 편모는 깃꼴 구조를 가진다. 정자와 난자에 의한 생식은 대부분 중심규조류에서 볼 수 있다.
바다와 호수에 서식하는 특정 세균 종은 유기 조류 물질을 분해하기 위해 가수분해 효소를 사용하여 죽거나 살아있는 돌말류의 규산 용해 속도를 가속화할 수 있다.[69][70]
5. 1. 무성 생식
돌말류는 주로 이분법으로 무성생식을 한다. 세포 분열은 껍질 안에서 진행되며, 두 개의 딸세포는 모세포의 외각과 내각 안쪽에 각각 새로운 반쪽 껍질을 형성한다. 따라서 모세포의 반쪽 껍질은 각각 하나씩 딸세포에 분배된다.[105] 그 결과, 딸세포 중 하나는 모세포와 같은 크기지만, 다른 하나는 내각을 외각으로 사용하기 때문에 모세포보다 작아진다. 이러한 무성 생식을 반복하면 개체군의 평균 크기가 감소한다.[105]5. 2. 유성 생식
무성 생식을 반복하면 세포의 크기가 작아지는데, 이를 회복하기 위해 유성 생식이 일어난다. 중심 돌말류는 감수 분열을 통해 편모를 가진 유주 세포를 만든다. 이 유주 세포는 다른 단상 개체와 합쳐져 증대 포자(auxospore)를 형성한다.[105] 증대 포자는 껍질이 없고, 성장하여 크기를 늘려 일반 세포로 돌아간다. 깃털 돌말류는 감수 분열로 동형 배우자를 만들고, 이들은 접합하여 증대 포자를 형성한다.[105]6. 생태
돌말류는 단세포 식물이며, 현재 약 12,000종 가량이 알려져 있다. 민물이나 바닷물에서 부유 생활(식물 플랑크톤)이나 부착 생활을 하며, 넓은 지역에 걸쳐 광합성을 하고 효율이 높아 많은 유기물을 생산하므로 수계의 매우 중요한 물질 생산자이다. 몸은 규산질로 된 2장의 세포벽으로 둘러싸여 있으며, 각편(껍데기)에는 정교한 무늬가 있는데, 그 배열 방식에 따라 좌우대칭인 우상규조류와 방사대칭인 중심규조류로 나뉜다.
생식에는 무성생식과 유성생식이 있다. 무성생식은 주로 분열법에 의해 이루어진다. 즉, 위·아래 2장의 껍데기가 갈라지면서 핵과 원형질이 2개로 나누어져 각각의 껍데기 안쪽에 새로운 껍데기를 만들어 2개의 새로운 개체가 생긴다. 유성생식에는 증대 포자 형성과 정자와 난자가 합쳐지는 방법, 2가지가 있다. 증대 포자 형성은 분열을 계속하면 세포 크기가 작아지므로, 접합하여 증대 포자를 만듦으로써 크기를 회복하는 방법으로, 우상규조류에서 흔히 볼 수 있다. 정자와 난자에 의한 생식은 대부분 중심규조류에서 볼 수 있다. 정자는 겉보기에 1개의 편모만을 가진 것처럼 보이지만, 실제로는 길고 짧은 2개의 편모가 있는데, 다른 하나는 너무 짧아 몸 속에 묻혀 있어 체표에 드러나지 않는다. 몸 밖으로 나온 긴 편모는 깃꼴구조를 가진다.

6. 1. 분포
규조류는 대양과 담수, 토양과 축축한 표면 등 매우 다양한 환경에서 발견된다.[76] 대부분 열린 물에서 원양 생활을 하지만, 일부는 물-퇴적물 경계면(저서생물)에서 표면 막으로 살거나 습한 대기 조건에서도 서식한다. 특히 대양에서는 1차 생산자로서 중요한 위치를 차지하며, 2003년 연구에서는 전체 해양 1차 생산 유기물의 약 45%를, 2016년 연구에서는 20%를 차지하는 것으로 추정되었다.[72][73] 해양 식물 플랑크톤 종의 공간 분포는 수평 및 수직으로 제한된다.[74][75]담수에서도 중요한 생산자이며, 어류의 먹이로 중요하다. 흐르는 물에서는 벤토스로서 돌 표면에 부착하여 은어 등의 먹이가 된다.
pH 1.2의 강산성 온천에서부터 pH 11의 강알칼리성 호수인 마가디 호와 같은 염기성 호수, 고염 농도의 내륙 호수나 강 부패 수역 등 극한 환경에서도 생육한다. 극지방의 유빙 하부에도 저온성 규조가 생육하며, 해빙 조류(ice algae)라고 불린다. 일부 규조는 유공충과 세포 내 공생하는 경우도 보고된다.
6. 2. 생장
플랑크톤성 돌말류는 담수 및 해양 환경에서 일반적으로 "붐 앤 버스트"(또는 "블룸 앤 버스트") 생활사를 보인다. 상층 혼합층의 조건(영양분과 빛)이 유리할 때(예: 봄), 이들의 경쟁 우위와 빠른 성장률[76]은 식물 플랑크톤 군집을 지배할 수 있게 한다("붐" 또는 "블룸"). 따라서 이들은 종종 기회주의적인 r-전략가로 분류된다(예: 높은 성장률, ''r''로 생태학이 정의되는 유기체).일반적으로 영양분 고갈 시, 조건이 악화되면 규조류 세포는 침강 속도가 증가하고 상부 혼합층에서 벗어난다("파괴"). 이러한 침강은 부력 조절 상실, 규조류 세포를 함께 묶는 점액질 합성 또는 무거운 ''휴면 포자'' 생성에 의해 유도된다. 상부 혼합층에서 벗어나는 것은 규조류를 포식자 개체군과 더 높은 온도(세포 대사를 증가시킴)를 포함하여 성장에 불리한 조건으로부터 제거한다. 더 깊은 물이나 얕은 해저에 도달한 세포는 조건이 다시 유리해질 때까지 휴식을 취할 수 있다. 대양에서는 많은 침강 세포가 깊은 곳으로 손실되지만 피난처 개체군은 수온약층 근처에서 지속될 수 있다.
이러한 휴면 개체군 내의 규조류 세포는 수직 혼합이 그들을 가두면 상부 혼합층으로 다시 들어간다. 대부분의 경우, 이러한 혼합은 상부 혼합층의 영양분을 보충하여 다음 규조류 번식의 무대를 설정한다. 대양(지속적인 용승 지역에서 멀리 떨어진 곳[80])에서 이러한 번식, 파괴, 사전 번식 조건으로의 복귀 주기는 일반적으로 연간 주기로 발생하며, 규조류는 봄과 초여름에만 우세하다. 그러나 일부 지역에서는 여름 성층화의 붕괴와 빛 수준이 여전히 성장에 충분한 동안 영양분 유입으로 인해 가을 번식이 발생할 수 있다. 수직 혼합이 증가하고 겨울이 다가옴에 따라 빛 수준이 감소함에 따라 이러한 번식은 봄에 비해 더 작고 수명이 짧다.
대양에서 규조류(봄) 번식은 일반적으로 규소 부족으로 끝난다. 다른 광물과 달리 규소에 대한 요구 사항은 규조류에만 해당하며, 질소 또는 인 영양분만큼 플랑크톤 생태계에서 효율적으로 재생되지 않는다. 이것은 표면 영양분 농도 지도를 통해 알 수 있다. 즉, 영양분이 기울기를 따라 감소함에 따라 규소가 일반적으로 가장 먼저 고갈된다(일반적으로 질소, 인 순).
6. 3. 생지화학적 역할 (Biogeochemistry)
규조류는 해양에서 규소의 생지화학적 순환 조절에 핵심적인 역할을 한다.[72][85] 돌말류는 성장하면서 표층수에서 용존 규산을 추출하고, 죽으면 수층으로 되돌려 보낸다. 규소의 유입은 바람에 의한 먼지를 통해 상부에서, 강을 통해 해안에서, 해저 퇴적물 재활용, 풍화 작용, 열수 활동을 통해 하부에서 발생한다.[85] 해양의 대부분의 생물기원 규소(생물 생체 광물화에 의해 생성된 규소)는 돌말류에서 나온다.돌말류는 트라이아스기부터 존재했을 수 있지만, 규소 순환의 부상과 "장악" 시기는 더 최근에 일어났다. 현생누대 이전 (5억 4천 4백만 년 전)에는 미생물 또는 무기물 과정이 해양의 규소 순환을 약하게 조절한 것으로 여겨진다.[86][87][88] 그 후, 순환은 방사충과 규질 해면동물에 의해 지배(및 더 강력하게 조절)되는 것으로 보이며, 전자는 동물성 플랑크톤으로, 후자는 주로 대륙붕에서 정착하는 여과 섭식자이다.[89] 지난 1억 년 이내에 규소 순환이 더욱 엄격하게 통제되었으며, 이는 돌말류의 생태적 우세에서 비롯된 것으로 생각된다.
"장악"의 정확한 시기는 여전히 불분명하며, 여러 저자는 화석 기록에 대해 상반된 해석을 하고 있다. 규질 해면동물의 선반에서의 이탈과 같은 일부 증거[90]는 이 장악이 백악기 (1억 4천 6백만 년 전 ~ 6천 6백만 년 전)에 시작되었음을 시사하는 반면, 방사충의 증거는 "장악"이 신생대 (6천 6백만 년 전 ~ 현재)까지 시작되지 않았음을 시사한다.[91]
돌말류는 해양 표층수에서 탄소 수출에 불균형적으로 중요한 역할을 한다.[80][81]
대기 중으로의 인위적인 CO2 배출(주로 화석 연료 연소 및 삼림 벌채로 발생)은 연간 거의 11 기가톤 탄소(GtC)이며, 이 중 거의 2.5 GtC가 표층 해양에 흡수된다. 표층 해수(pH 8.1–8.4)에서 중탄산염 (HCO3-) 및 탄산염 이온(CO32-)은 각각 용존 무기 탄소(DIC)의 거의 90%와 <10%를 차지하며, 용존 CO2(수성 CO2)는 <1%를 차지한다. 해양의 낮은 CO2 농도와 물에서의 느린 확산 속도에도 불구하고 돌말류는 광합성을 통해 연간 10~20 GtC를 고정하며, 이는 그들의 이산화 탄소 농축 기작 덕분이며, 이를 통해 해양 먹이 사슬을 유지할 수 있다. 또한, 유광층에서 생성된 이 유기물의 0.1~1%는 입자 형태로 침강하여 표면 탄소를 심해로 이동시키고 수천 년 이상 동안 탄소 격리를 통해 대기 중 CO2를 격리시킨다. 나머지 유기물은 호흡을 통해 재광물화된다. 따라서 돌말류는 생물학적 탄소 펌프의 주요 구성 요소 중 하나이며, 이는 지구 시스템에서 CO2를 탄소 순환에서 매우 오랫동안 제거할 수 있게 해주는 가장 중요한 생물학적 메커니즘이라고 할 수 있다.[93][92]
돌말류의 특징은 요소 회로이며, 이는 돌말류를 진화적으로 동물과 연결한다. 2011년 Allen et al.은 돌말류가 기능하는 요소 회로를 가지고 있음을 밝혀냈다. 이는 중요한 결과였는데, 왜냐하면 이전에 요소 회로는 돌말류보다 수억 년 전에 출현한 후생동물에서 시작된 것으로 여겨졌기 때문이다. 그들의 연구는 돌말류와 동물이 서로 다른 목적으로 요소 회로를 사용하지만, 동물과 식물이 그렇지 않은 방식으로 진화적으로 연결되어 있음을 보여주었다.[95]
광합성 생물에서 종종 간과되지만, 미토콘드리아 역시 에너지 균형에 중요한 역할을 한다. 두 가지 질소 관련 경로가 관련되어 있으며, 이는 암모늄 (NH4+) 영양 조건과 질산염 (NO3-) 영양 조건에서 달라질 수 있다. 돌말류와 다른 일부 조류에서 요소 회로가 존재한다.[96][97][98] 동물에서 요소 회로의 오랫동안 알려진 기능은 아미노산 이화작용에 의해 생성된 과도한 질소를 배설하는 것이며, 광호흡과 마찬가지로 요소 회로는 오랫동안 폐기 경로로 여겨져 왔다. 그러나 돌말류에서 요소 회로는 미토콘드리아와 세포질 사이의 영양소 교환, 잠재적으로 색소체[99]에서 역할을 하는 것으로 보이며, 암모늄 대사를 조절하는 데 도움을 줄 수 있다.[96][97] 이 회로 때문에, 해양 돌말류는 녹조류와 달리 미토콘드리아 요소 수송체를 획득했으며, 실제로 생물정보학에 따르면 완전한 미토콘드리아 GS-GOGAT 회로가 가설로 제시되었다.[97][94]
7. 분류
돌말류는 약 12,000종이 알려져 있지만, 실제로는 10만 종이 넘을 것으로 추정된다.[174] 이는 껍질 형태를 기준으로 분류된 종 외에, 유성 생식 확인 실험을 통해 밝혀진 종들이 있기 때문이다. Sellaphora영어 속과 같은 일부 규조류는 형태가 비슷한 종들끼리만 유성 생식을 하는 것으로 나타났다.[174]
돌말류의 분류는 아직 완전히 정립되지 않았으며, 분자생물학적 연구 결과에 따라 계속 수정되고 있다. 1990년 Round, Crawford 및 Mann은 규조류를 문(Phylum)으로 취급하고 주요 분류 단위를 강(Class)으로 하는 분류 체계를 제안했다. 중심형 규조류는 Coscinodiscophyceae 강으로 유지되었고, 깃털형 규조류는 Fragilariophyceae와 Bacillariophyceae 두 강으로 분리되었다.[174]
2004년, 린다 메들린(Linda Medlin)과 공동 연구자들은 일부 중심형 규조류 목을 Mediophyceae라는 새로운 강으로 분리할 것을 제안했다. 이들은 Mediophyceae가 다른 중심형 규조류보다 깃털형 규조류와 더 가깝다고 보았다.[112][113][114][115] 그러나 이 제안에 대한 이견도 존재한다.
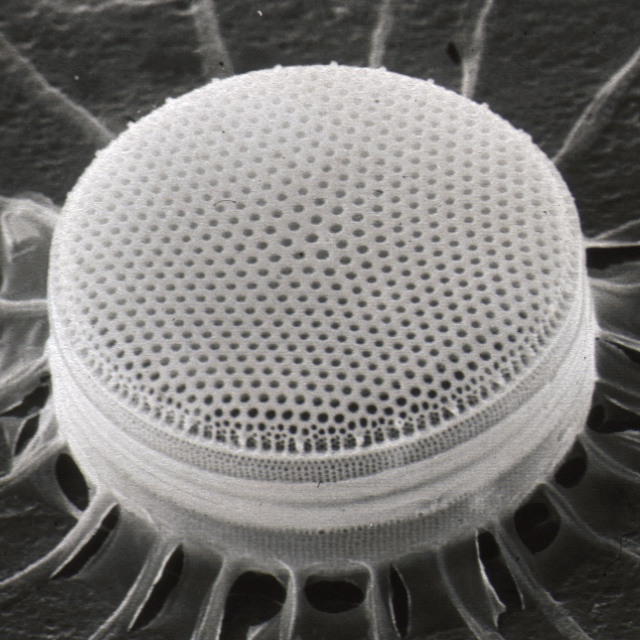


''Stephanodiscus hantzschii''}}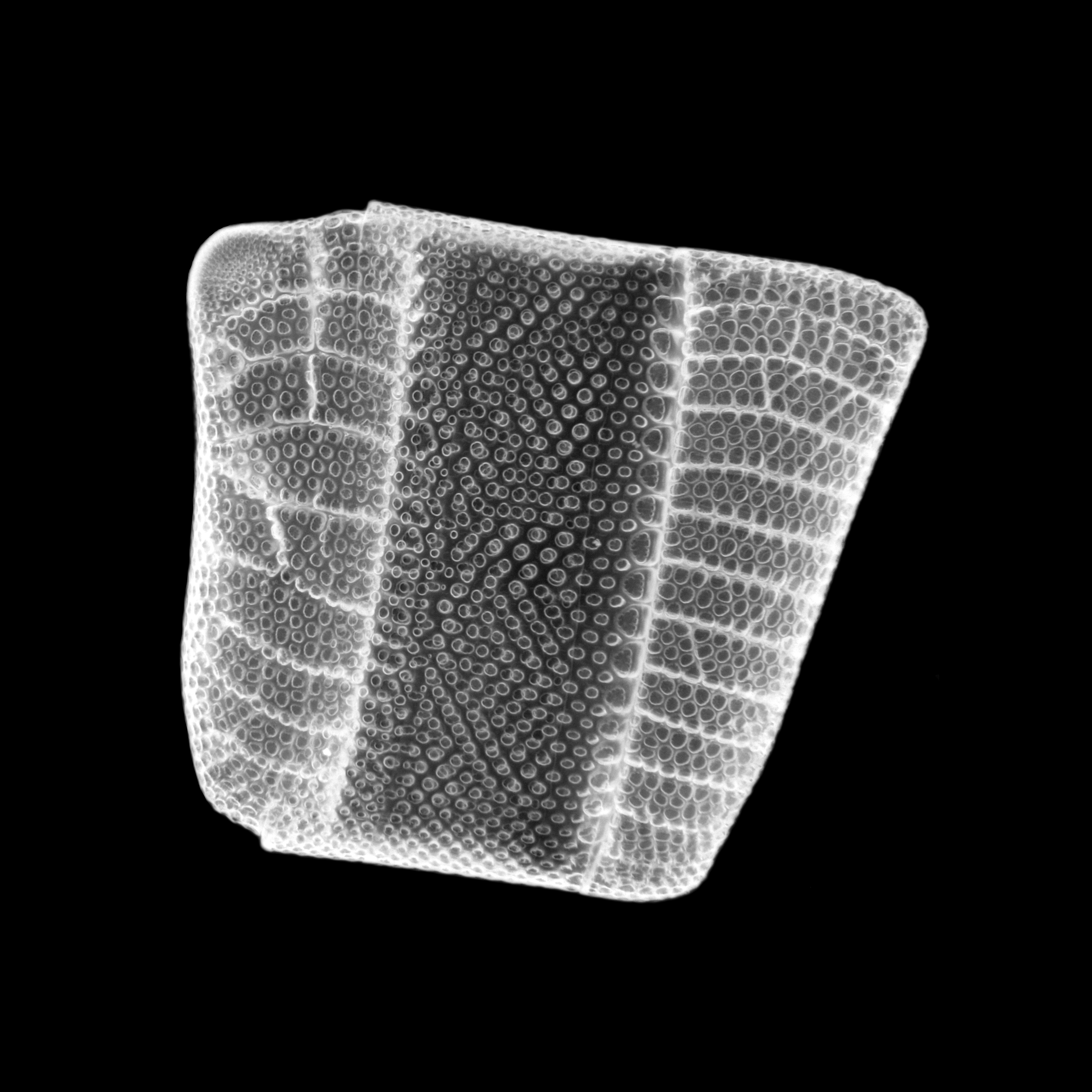
''Isthmia nervosaIsthmia nervosa''}}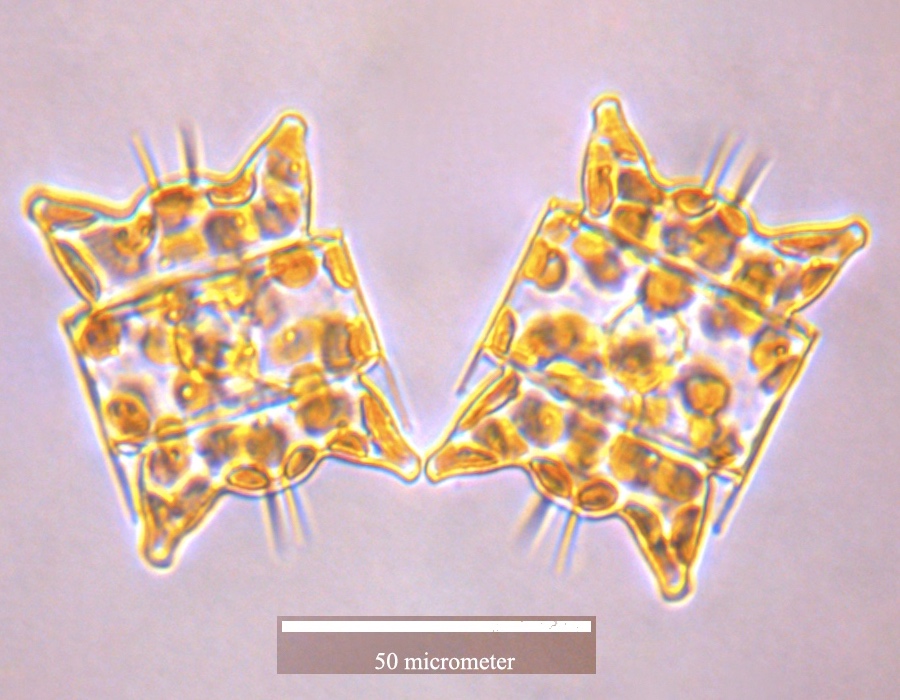
''Odontella aurita''}}
7. 1. 주요 분류군
규조류 분류는 아직 명확하게 정립되지 않았다. 과거에는 껍질 표면 무늬에 따라 중심규조목(Centrales)과 우상규조목(Pennales)으로 분류했다. 그러나 최근 연구 결과에 따라 우상규조목을 세로홈(raphe) 유무에 따라 위배선규조아목(Fragilariophyceae)과 원시배선규조아목(Bacillariophyceae)으로 나눈다.[174]- 중심규조목 (Centrales) 또는 체돌말아강(Coscinodiscophycidae) - 몸의 기본형은 원통 모양이고 껍질 표면 무늬는 방사대칭이다. 여러 개의 색소체를 가지고 있으며, 유성생식은 알과 유영성 정자로 한다.
- 원반규조아목 - 껍질 가장자리에 고리 모양 돌기줄이 1줄 있다 (체모양원반류, 방사원반류 등).
- 원통규조아목 - 껍질에 1개의 돌기부가 있다 (원통돌말류 등).
- 실패돌말아목 - 껍질에 2개 이상의 돌기부가 있다 (실패돌말, 가지돌말 등).
- 우상규조목 (Pennales)
- 위배선규조강(Fragilariophyceae) 또는 위배선규조아강(Fragilariophycidae) - 상하 양 뚜껑껍질에 세로홈이 없다 (띠돌말류 등).
- 원시배선규조강(Bacillariophyceae) 또는 원시배선규조아강(Bacillariophycidae) - 상하 양 뚜껑껍질 또는 한쪽 뚜껑껍질에 세포홈이 있다. (아크난테스류, 수리렐라류, 나비쿨라류, 킴벨라류, 유노티아류, 띠돌말류, 니츠키아류 등).
조류 정보 사이트 AlgaeBase에 따른 돌말류 최신 분류는 아래와 같다.[175]
| 문 | 아문 | 강 | 아강 | 목 |
|---|---|---|---|---|
| 돌말식물문 (Bacillariophyta) | 돌말식물아문 (Bacillariophytina) | 돌말강 (Bacillariophyceae) | 원시배선규조류 (Bacillariophycidae) | 윷돌말목 (Bacillariales) |
| 낟알돌말목 (Cocconeidales) | ||||
| 반달돌말목 (Cymbellales) | ||||
| Dictyoneidales | ||||
| 현돌말목 (Lyrellales) | ||||
| 잎새돌말목 (Mastogloiales) | ||||
| 쪽배돌말목 (Naviculales) | ||||
| 위배선규조류 (Fragilariophycidae) | 활돌말목 (Rhopalodiales) | |||
| 방패돌말목 (Surirellales) | ||||
| Thalassiophysales | ||||
| 눈썹돌말목 (Eunotiales) | ||||
| 김발돌말목 (Fragilariales) | ||||
| 체돌말류 (Coscinodiscophyceae) | - | - | Archaegladiopsophycidae | Archaegladiopsidales |
| 수염돌말아강 (Corethrophycidae) | 수염돌말목 (Corethrales) | |||
| 체돌말아강 (Coscinodiscophycidae) | 별빛돌말목 (Asterolamprales) | |||
| - | 사슬돌말목 (Stephanopyxales) | |||
| - | 점판돌말목 (Stictodiscales) |
2019년, Adl 등[116]은 돌말류 분류를 다음과 같이 제시했다.
- 분기도 돌말류분기도
- * 문 돌말류
- ** 아문 Leptocylindrophytina
강 Leptocylindrophyceae (''Leptocylindrus'')
강 Corethrophyceae (''Corethron'')
- ** 아문 Ellerbeckiophytina (''Ellerbeckia'')
- ** 아문 Probosciophytina (''Proboscia'')
- ** 아문 Melosirophytina (''Aulacoseira'', ''Melosira'')
- ** 아문 Coscinodiscophytina (''Actinoptychus'', ''Coscinodiscus'')
- ** 아문 Rhizosoleniophytina (''Guinardia'', ''Rhizosolenia'')
- ** 아문 Arachnoidiscophytina (''Arachnoidiscus'')
- ** 아문 Bacillariophytina
강 Mediophyceae
* 아강 Chaetocerotophycidae
* 아강 Lithodesmiophycidae
* 아강 Thalassiosirophycidae
* 아강 Cymatosirophycidae
* 아강 Odontellophycidae
* 아강 Chrysanthemodiscophycidae
강 Biddulphiophyceae
* 아강 Biddulphiophycidae
* Biddulphiophyceae incertae sedis (''Attheya'')
강 Bacillariophyceae
* Bacillariophyceae incertae sedis (Striatellaceae)
* 아강 Urneidophycidae
* 아강 Fragilariophycidae
* 아강 Bacillariophycidae
자세한 내용은 돌말류 분류에서 확인할 수 있다.
8. 인간과의 관계
돌말류는 산업, 환경, 법의학 등 다양한 분야에서 인간과 밀접한 관계를 맺고 있다.
8. 1. 산업적 이용
규조토는 돌말류 껍질이 퇴적되어 형성된 광물로, 미세 연마 및 액체 여과에 사용되는 중요한 산업 자원이다.[157]규조토는 천연 나노 물질로 간주되며 다음과 같은 다양한 용도로 활용된다.[158][159][160]
| 활용 분야 |
|---|
| 다양한 세라믹 제품, 건설 세라믹, 내화 세라믹, 특수 산화물 세라믹 |
| 습도 제어 재료 생산 |
| 여과 재료 |
| 시멘트 생산 산업의 재료 |
| 서방형 약물 운반체 생산을 위한 초기 재료 |
| 산업 규모의 흡수 재료 |
| 다공성 세라믹 생산 |
| 유리 산업 |
| 촉매 지지체 |
| 플라스틱 및 페인트의 충전재 |
| 산업용수 정화 |
| 살충제 홀더 |
| 특정 토양의 물리적 및 화학적 특성 개선 |
돌말류의 미세한 껍질 구조는 나노기술의 재료로 제안되었다.[157] 돌말 세포는 다양한 모양과 크기의 껍질을 반복적이고 안정적으로 제조하여, 광학 시스템, 반도체 나노리소그래피, 약물 전달 운송 수단을 포함한 다양한 장치에 사용될 수 있는 마이크로 또는 나노 스케일 구조를 제조할 수 있게 한다.[161][162]
인공 선택을 통해 특정 모양과 크기의 껍질을 생산하는 돌말을 연속 배양하여 나노 스케일 구성 요소를 대량 생산하도록 진화시킬 수 있다.[162] 또한 돌말이 세포벽을 만드는 데 사용하는 이산화규소를 광과민성 이산화 티타늄으로 대체하여 태양 전지의 구성 요소로 사용할 수 있다는 제안도 있었다.[163] 돌말 바이오 연료 생산 태양 전지도 제안되었다.[164]
8. 2. 환경 지표
돌말류의 분해와 부패는 유기물과 무기물(규산염 형태) 퇴적물을 생성하며, 이 무기물 성분은 해저 또는 만(灣) 갯벌의 코어링을 통해 과거 해양 환경을 분석하는 데 사용될 수 있다. 무기물은 점토와 실트의 퇴적물에 포함되어 있으며 이러한 해양 지층의 영구적인 지질 기록을 형성하기 때문이다(규질 연니 참조).8. 3. 법의학적 이용
법의학에서 돌말류 분석은 익사로 인한 사망과 사후 시신이 물속에 담긴 경우를 구별하는 데 주로 활용된다. 실험실 검사를 통해 시신 내 돌말류 존재 여부를 확인할 수 있다.[165] 돌말류의 규산질 골격은 쉽게 부패하지 않아 심하게 부패한 시신에서도 종종 검출된다.[165] 돌말류는 시신 내에 자연적으로 존재하지 않기 때문에, 시신에서 발견된 돌말류가 시신이 발견된 물에서 발견된 종과 동일하다면 익사로 인한 사망을 뒷받침하는 좋은 증거가 될 수 있다.[165] 시신에서 발견된 돌말류 종의 혼합은 주변 물과 같거나 다를 수 있으며, 이는 피해자가 시신이 발견된 장소에서 익사했는지 여부를 나타낸다.[165]참조
[1]
서적
Traite d'Algologie
https://books.google[...]
Paul Lechvalier and Fils, Paris
1933
[2]
웹사이트
Bacillariophyceae
2024-05-09
[3]
서적
Commentationes botanicae. Observations botaniques, dédiées à la Société d'Horticulture de Tournay
http://images.algaeb[...]
Imprimerie de Ch. Casterman-Dieu, Rue de pont No. 10
[4]
서적
Flora europaea algarum aquae dulcis et submarinae
https://archive.org/[...]
Apud Eduardum Kummerum
1864-1868
[5]
문서
Das Protistenreich
http://caliban.mpipz[...]
1878
[6]
서적
Syllabus der Pflanzenfamilien: eine Übersicht über das gesamte Pflanzensystem mit besonderer Berücksichtigung der Medizinal- und Nutzpflanzen, nebst einer Übersicht über die Florenreiche und Florengebiete der Erde zum Gebrauch bei Vorlesungen und Studien über spezielle und medizinisch-pharmazeutische Botanik
Gebrüder Borntraeger Verlag, Berlin
1919
[7]
LSJ
διατομή
[8]
LSJ
διάτομος
[9]
LSJ
διατέμνω
[10]
백과사전
The Compact Oxford English Dictionary
Clarendon Press
[11]
웹사이트
The Air You're Breathing? A Diatom Made That
https://www.livescie[...]
2014-06-11
[12]
웹사이트
What are Diatoms?
Diatoms of North America
2020-01-28
[13]
논문
The Silica Balance in the World Ocean: A Reestimate
[14]
웹사이트
King's College London – Lake Megachad
https://www.kcl.ac.u[...]
2018-05-05
[15]
논문
Fertilizing the Amazon and equatorial Atlantic with West African dust
2010
[16]
웹사이트
Gas Guzzlers
https://www.smithson[...]
2018-05-22
[17]
웹사이트
More on Diatoms
http://www.ucmp.berk[...]
2015-05-20
[18]
서적
Algae: an introduction to Phycology
Cambridge University Press
[19]
논문
Complete genome of a nonphotosynthetic cyanobacterium in a diatom reveals recent adaptations to an intracellular lifestyle
[20]
논문
Global distribution patterns of marine nitrogen-fixers by imaging and molecular methods
2021-07-06
[21]
논문
Functional Relationship between a Dinoflagellate Host and Its Diatom Endosymbiont
https://academic.oup[...]
Oxford Academic
[22]
서적
Freshwater Algae of North America: Ecology and Classification
Academic Press
[23]
논문
Thai amber: insights into early diatom history?
2020
[24]
NASA
The Inner Space of the Subarctic Pacific Ocean
https://blogs.nasa.g[...]
NASA Earth Expeditions
2018-09-04
[25]
논문
Recent decadal trends in global phytoplankton composition
[26]
논문
Quantitative 3D-imaging for cell biology and ecology of environmental microbial eukaryotes
[27]
논문
Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation
American Geophysical Union (AGU)
[28]
논문
The species concept in diatoms
Informa UK Limited
[29]
논문
The diatom system: ideas on phylogeny
[30]
논문
Morphological and transcriptomic evidence for ammonium induction of sexual reproduction in Thalassiosira pseudonana and other centric diatoms
Public Library of Science (PLoS)
2017-07-07
[31]
웹사이트
Diatoms
https://www.ucl.ac.u[...]
2002
[32]
논문
Biomimetics of photonic nanostructures
[33]
논문
The Glass Menagerie: Diatoms for novel applications in nanotechnology
[34]
웹사이트
Glass in Nature
http://www.cmog.org/[...]
The Corning Museum of Glass
2013-02-19
[35]
서적
An Illustrated Guide to Some Common Diatom Species from South Africa
Water Research Commission
2007
[36]
간행물
"All new faces of diatoms: potential source of nanomaterials and beyond"
[37]
논문
Structure and properties of oil bodies in diatoms
https://doi.org/10.1[...]
2017-07-17
[38]
논문
Bio-manufacturing technology based on diatom micro- and nanostructure
[39]
웹사이트
Diatoms
http://www.ucl.ac.uk[...]
2016-02-13
[40]
서적
The Molecular Life of Diatoms
https://books.google[...]
[41]
논문
Chytrid fungi distribution and co-occurrence with diatoms correlate with sea ice melt in the Arctic Ocean
[42]
논문
Comparative Sequence Analysis of Diatom Silicon Transporters: Toward a Mechanistic Model of Silicon Transport
2006
[43]
논문
Polycationic Peptides from Diatom Biosilica That Direct Silica Nanosphere Formation.
1999-11
[44]
서적
Handbook of Biomineralization: Biological Aspects and Structure Formation
Wiley-VCH Verlag GmbH
2007
[45]
간행물
"Two letters from a Gentleman in the Country, relating to Mr. Leeuwenhoek's Letter in Transaction, no. 283."
[46]
간행물
"[https://www.jstor.org/stable/23357402 Ueber den feinbau der kieselmembran der diatomeen]"
https://www.jstor.or[...]
[47]
논문
Understanding Diatom Cell Wall Silicification—Moving Forward
Frontiers Media SA
2018-04-11
[48]
논문
A gene family of silicon transporters
Springer Science and Business Media LLC
[49]
논문
Polycationic Peptides from Diatom Biosilica That Direct Silica Nanosphere Formation
American Association for the Advancement of Science (AAAS)
1999-11-05
[50]
논문
Species-specific polyamines from diatoms control silica morphology
2000-12-05
[51]
논문
Extensive and Intimate Association of the Cytoskeleton with Forming Silica in Diatoms: Control over Patterning on the Meso- and Micro-Scale
Public Library of Science (PLoS)
2010-12-10
[52]
논문
Characterization and Localization of Insoluble Organic Matrices Associated with Diatom Cell Walls: Insight into Their Roles during Cell Wall Formation
Public Library of Science (PLoS)
2013-04-23
[53]
논문
Nanopatterned protein microrings from a diatom that direct silica morphogenesis
2011-02-07
[54]
논문
Silicanin-1 is a conserved diatom membrane protein involved in silica biomineralization
Springer Science and Business Media LLC
2017-07-24
[55]
논문
Characterization of a New Protein Family Associated With the Silica Deposition Vesicle Membrane Enables Genetic Manipulation of Diatom Silica
Springer Science and Business Media LLC
2017-10-18
[56]
서적
Sinking properties of some phytoplankton shapes and the relation of form resistance to morphological diversity of plankton — an experimental study
http://real.mtak.hu/[...]
Springer Netherlands
2019-10-04
[57]
논문
Diel changes in sedimentation characteristics of Ditylum brightwelli: Changes in cellular lipid and effects of respiratory inhibitors and ion-transport modifiers1
1977-05-01
[58]
논문
Diatom gliding is the result of an actin-myosin motility system
1999-09-01
[59]
웹사이트
raphid diatoms
http://tolweb.org/ra[...]
2010-02
[60]
서적
The Biology of Diatoms
University of California Press
2013-11-14
[61]
간행물
"The diatom sex clock and its evolutionary significance"
[62]
간행물
"Experimental studies on sexual reproduction in diatoms"
London
[63]
간행물
"Sexuality"
Oxford: Blackwell Scientific Publications.
[64]
논문
Size reduction, reproductive strategy and the life cycle of a centric diatom
The Royal Society
1992-05-29
[65]
논문
Sexual reproduction in the marine centric diatom Ditylum brightwellii (Bacillariophyta)
Informa UK Limited
[66]
간행물
Light is a key factor in triggering sexual reproduction in the pennate diatom Haslea ostrearia.
[67]
간행물
The dynamics of sexual phase in the marine diatom Pseudo-nitzschia multistriata (Bacillariophyceae).
[68]
간행물
Identification of the meiotic toolkit in diatoms and exploration of meiosis-specific SPO11 and RAD51 homologs in the sexual species Pseudo-nitzschia multistriata and Seminavis robusta.
[69]
논문
Accelerated dissolution of diatom silica by marine bacterial assemblages
[70]
논문
The Structure of Microbial Community and Degradation of Diatoms in the Deep Near-Bottom Layer of Lake Baikal
[71]
논문
Silicate as regulating nutrient in phytoplankton competition
[72]
논문
Role of diatoms in regulating the ocean's silicon cycle
[73]
논문
Insights into global diatom distribution and diversity in the world's ocean
2016-02-29
[74]
논문
Plankton Evolution
[75]
서적
A taxonomic guide to some common marine phytoplankton
https://books.google[...]
Biopress
2013-11-13
[76]
논문
In situ growth rates of marine phytoplankton: Approaches to measurement, community and species growth rates
[77]
웹사이트
Didymo, Aliens Among Us
http://alienspecies.[...]
[78]
웹사이트
DEP Reports Didymo Discovered in the West Branch Farmington River
http://www.ct.gov/de[...]
2015-04-27
[79]
웹사이트
Didymo Stakeholder Update – 31 October 2008
http://www.biosecuri[...]
MAF Biosecurity New Zealand
2013-12-01
[80]
논문
Silicate regulation of new production in the equatorial Pacific upwelling
[81]
논문
Role of sinking in diatom life-history cycles: Ecological, evolutionary and geological significance
[82]
논문
The transport and function of silicon in plants
[83]
논문
A proton buffering role for silica in diatoms
[84]
문서
Methods in Plant Histology
University of Chicago Press
1901
[85]
논문
The Silica Balance in the World Ocean: A Reestimate
[86]
서적
Scientists on Gaia
https://archive.org/[...]
MIT Press
2013-11-14
[87]
논문
Secular Distribution of Biogenic Silica through the Phanerozoic: Comparison of Silica-Replaced Fossils and Bedded Cherts at the Series Level
[88]
논문
Paleozoic and Mesozoic silica-rich seawater: Evidence from hematitic chert (jasper) deposits
[89]
논문
Radiolarian palaeoecology and radiolarites: Is the present the key to the past?
[90]
논문
Decline in Mesozoic reef-building sponges explained by silicon limitation
[91]
논문
Silica, diatoms, and Cenozoic radiolarian evolution
[92]
논문
Carbon Dioxide Concentration Mechanisms in Natural Populations of Marine Diatoms: Insights From Tara Oceans
Frontiers Media SA
2021-04-30
[93]
논문
Global Carbon Budget 2020
Copernicus GmbH
2020-12-11
[94]
논문
Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions
Wiley
2015-10-11
[95]
논문
Evolution and metabolic significance of the urea cycle in photosynthetic diatoms
http://www.locus.ufv[...]
2021-05-20
[96]
논문
The Genome of the Diatom Thalassiosira Pseudonana : Ecology, Evolution, and Metabolism
https://digital.libr[...]
American Association for the Advancement of Science (AAAS)
[97]
논문
Evolution and metabolic significance of the urea cycle in photosynthetic diatoms
http://repositorio.u[...]
Springer Science and Business Media LLC
[98]
논문
Inactivation of P haeodactylum tricornutum urease gene using transcription activator-like effector nuclease-based targeted mutagenesis
Wiley
2014-10-10
[99]
논문
Coupled Effects of Light and Nitrogen Source on the Urea Cycle and Nitrogen Metabolism over a Diel Cycle in the Marine Diatom Thalassiosira pseudonana
Elsevier BV
[100]
논문
Utilisation of seaweed carbon by three surface-associated heterotrophic protists, Stereomyxa ramosa, Nitzschia alba and Labyrinthula sp.
2000
[101]
논문
Culture and Nutrition of Some Apochlorotic Diatoms of the Genus Nitzschia
1967
[102]
논문
Diatoms respire nitrate to survive dark and anoxic conditions
[103]
논문
Nitrate respiration and diel migration patterns of diatoms are linked in sediments underneath a microbial mat
[104]
논문
How many species of algae are there?
[105]
서적
Identifying Marine Diatoms and Dinoflagellates
Academic Press
1996-01-25
[106]
서적
The Diatoms: Biology & Morphology of the Genera
https://books.google[...]
Cambridge University Press
2013-11-13
[107]
서적
Freshwater Algae: Their microscopic world explained
Biopress Limited
[108]
논문
The species concept in diatoms: Evidence for morphologically distinct, sympatric gamodemes in four epipelic species
[109]
논문
Catalogue of the Diatom Genera
[110]
문서
The [[World Register of Marine Species]] lists 1,356 diatom genus names from all habitats as at July 2020, of which 1,248 are "accepted".
[111]
문서
Queries to the World Register of Marine Species, July 2020, return 299 "fossil only" genus names, of which 285 are "accepted".
[112]
논문
The limits of nuclear-encoded SSU rDNA for resolving the diatom phylogeny
[113]
논문
A preliminary multigene phylogeny of the diatoms (Bacillariophyta): challenges for future research
[114]
논문
Signal, uncertainty, and conflict in phylogenomic data for a diverse lineage of microbial eukaryotes (Diatoms, Bacillariophyta)
[115]
논문
Review of the phylogenetic reconstruction of the diatoms using molecular tools with an analysis of a seven gene data set using multiple outgroups and morphological data for a total evidence approach
http://plymsea.ac.uk[...]
2020-07-20
[116]
논문
Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes
2018-09-26
[117]
논문
All New Faces of Diatoms: Potential Source of Nanomaterials and Beyond
Frontiers Media SA
2017-07-05
[118]
논문
Evolution of the Diatoms (Bacillariophyta)
[119]
논문
Diagenetic origin of quartz silt in mudstones and implications for silica cycling
[120]
논문
Is the origin of the diatoms related to the end-Permian mass extinction?
[121]
논문
The evolution of silicification in diatoms: Inescapable sinking and sinking as escape?
[122]
논문
The Evolution of Modern Eukaryotic Phytoplankton
[123]
논문
Impact of Grassland Radiation on the Nonmarine Silica Cycle and Miocene Diatomite
[124]
논문
Cenozoic Planktonic Marine Diatom Diversity and Correlation to Climate Change
[125]
문서
"Climate Change 2007: Synthesis Report"
[126]
논문
Methods and applications of Cenozoic marine diatom biostratigraphy
[127]
웹사이트
What Are "Proxy" Data?
https://www.ncdc.noa[...]
2020-10-20
[128]
논문
Photic zone changes in the north-west Pacific Ocean from MIS 4–5e
Copernicus GmbH
2015-01-06
[129]
논문
Polar ocean stratification in a cold climate
https://gfzpublic.gf[...]
Springer Science and Business Media LLC
2021-12-15
[130]
논문
North Pacific seasonality and the glaciation of North America 2.7 million years ago
https://gfzpublic.gf[...]
Springer Science and Business Media LLC
2021-12-15
[131]
논문
Diatom δ18O evidence for the development of the modern halocline system in the subarctic northwest Pacific at the onset of major Northern Hemisphere glaciation
http://eprints.notti[...]
American Geophysical Union (AGU)
2006-02-24
[132]
논문
Surface-water freshening: A cause for the onset of North Pacific stratification from 2.75 Ma onward?
Elsevier BV
[133]
논문
Salinity changes in the North West Pacific Ocean during the late Pliocene/early Quaternary from 2.73Ma to 2.52Ma
http://nora.nerc.ac.[...]
Elsevier BV
2021-12-15
[134]
논문
Mid Holocene origin of the sea-surface salinity low in the subarctic North Pacific
Elsevier BV
[135]
논문
A pervasive link between Antarctic ice core and subarctic Pacific sediment records over the past 800kyrs
Elsevier BV
[136]
논문
Consistent relationship between global climate and surface nitrate utilization in the western subarctic Pacific throughout the last 500 ka
https://archimer.ifr[...]
American Geophysical Union (AGU)
2021-12-15
[137]
논문
Glacial/interglacial changes in nutrient supply and stratification in the western subarctic North Pacific since the penultimate glacial maximum
Elsevier BV
[138]
논문
Controls on deglacial changes in biogenic fluxes in the North Pacific Ocean
Elsevier BV
[139]
논문
Cretaceous record of diatom evolution, radiation, and expansion
[140]
서적
Proceedings of the Tenth International Diatom Symposium
Koeltz Scientific Books
[141]
서적
Eocene-Oligocene climatic and biotic evolution
Princeton University Press
[142]
논문
Appearance and extinction of planktonic diatoms during the past 18 m.y. in the Pacific and Southern oceans
[143]
논문
Genome Properties of the Diatom Phaeodactylum tricornutum
[144]
논문
The Diatom EST Database
[145]
논문
Comparative Genomics of the Pennate Diatom Phaeodactylum tricornutum
[146]
논문
Update of the Diatom EST Database: A new tool for digital transcriptomics
[147]
논문
The Genome of the Diatom Thalassiosira Pseudonana: Ecology, Evolution, and Metabolism
[148]
논문
The Phaeodactylum genome reveals the evolutionary history of diatom genomes
[149]
논문
A Very High Fraction of Unique Intron Positions in the Intron-Rich Diatom Thalassiosira pseudonana Indicates Widespread Intron Gain
[150]
논문
Potential impact of stress activated retrotransposons on genome evolution in a marine diatom
[151]
논문
Genomic Footprints of a Cryptic Plastid Endosymbiosis in Diatoms
http://epic.awi.de/2[...]
2019-01-13
[152]
논문
Genome editing in diatoms: achievements and goals
http://nbn-resolving[...]
2021-05-20
[153]
논문
Transformation of Nonselectablporter Genes in Marine Diatomse Re
2015-05-01
[154]
논문
Designer diatom episomes delivered by bacterial conjugation
2015-04-21
[155]
논문
An Expanded Plasmid-Based Genetic Toolbox Enables Cas9 Genome Editing and Stable Maintenance of Synthetic Pathways in Phaeodactylum tricornutum
2018-02-16
[156]
논문
A CRISPR/Cas9 system adapted for gene editing in marine algae
2016-07-01
[157]
논문
All New Faces of Diatoms: Potential Source of Nanomaterials and Beyond
[158]
논문
New optimized method for low-temperature hydrothermal production of porous ceramics using diatomaceous earth
https://www.research[...]
2020-04-08
[159]
논문
Physical-chemical and mineralogical-petrographic examinations of diatomite from deposit near village of Rožden, Republic of Macedonia
http://js.ugd.edu.mk[...]
2020-04-08
[160]
논문
Effect Of Thermal Treatment Of Trepel At Temperature Range 800-1200˚C
2019-12-31
[161]
논문
Nature's Nanotechnologists: Unveiling the Secrets of Diatoms
[162]
논문
Star Trek replicators and diatom nanotechnology
[163]
간행물
Diatoms could triple solar cell efficiency
http://www.eetimes.c[...]
2009-04-13
[164]
논문
Milking Diatoms for Sustainable Energy: Biochemical Engineering versus Gasoline-Secreting Diatom Solar Panels
[165]
논문
Qualitative Diatom Analysis as a Tool to Diagnose Drowning
[166]
논문
Exploration of marine phytoplankton: from their historical appreciation to the omics era
[167]
논문
Unmasking "The Eldest Son of The Father of Protozoology": Charles King
2019-08-01
[168]
서적
On the Origin of Species by Means of Natural Selection: Or the Preservation of Favoured Races in the Struggle for Life
[169]
문서
秋田県の珪藻2-豊川の珪藻秋田県の珪藻2-豊川の珪藻
http://diatomology.o[...]
[170]
문서
The Revised Classification of Eukaryotes
http://www.paru.cas.[...]
[171]
뉴스
世界最速増殖の植物プランクトン、高松の干潟に 読売新聞オンライン 10月3日
https://web.archive.[...]
[172]
뉴스
高速増殖の植物プランクトン確認 CO2の効率吸収に活用も 47NEWS 2012年7月17日
https://web.archive.[...]
[173]
문서
The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists
null
[174]
문서
The Diatoms. Biology and Morphology of the Genera
Cambridge University Press
[175]
AlgaeBase
Bacillariophyta
[176]
논문
Higher Level Classification of All Living Organisms
[177]
논문
Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes
https://hal.archives[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com