미크로랍토르
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
미크로랍토르는 중국 랴오닝성 지우포탕 지층에서 발견된 깃털 공룡으로, 2000년에 명명되었다. 이 공룡은 앞다리와 뒷다리에 모두 날개가 있어 활공 또는 동력 비행을 했을 것으로 추정되며, 멜라노솜 분석 결과 깃털에 무지개빛이 있었을 가능성이 제기되었다. 미크로랍토르는 다양한 동물을 먹이로 삼는 기회주의적 포식자였으며, 비행 능력과 식성에 대한 연구가 활발히 진행되고 있다.
더 읽어볼만한 페이지
- 동물의 비행 - 유시충
유시충은 곤충학에서 날개 있는 사회성 곤충을, 식물학에서는 씨앗의 날개 모양 구조를 의미한다. - 동물의 비행 - 혼인 비행
혼인 비행은 성숙한 개미 군체에서 처녀 여왕과 수컷 개미가 짝짓기를 위해 집단적으로 비행하는 현상이며, 근친 교배를 피하고 짝짓기 후 수컷은 죽고 암컷은 새로운 군체를 건설한다. - 깃털공룡 - 시노사우롭테릭스
시노사우롭테릭스는 초기 백악기 압트절에 살았던 작은 이족 보행 수각류 공룡으로, 깃털 화석이 발견된 최초의 공룡이며, 꼬리의 띠 색깔과 콤프소그나투스와 유사한 구조, 긴 꼬리가 특징이다. - 깃털공룡 - 벨로키랍토르
벨로키랍토르는 백악기 후기 동아시아에 서식한 드로마에오사우루스과의 공룡으로, 날렵한 몸놀림과 낫 모양의 발톱을 이용해 사냥했으며, 깃털 공룡이었고, 프로토케라톱스와 싸우는 화석을 통해 생태 연구에 중요한 자료가 되지만, 영화 쥬라기 공원의 묘사는 실제와 차이가 있다. - 2000년 기재된 화석 분류군 - 제노스밀루스
제노스밀루스는 초기 플라이스토세 시대 북미에 살았던 멸종된 고양이과 동물로, 튼튼한 체격과 독특한 송곳니 형태를 지니며 사회적 동물이었을 가능성이 제기된다. - 2000년 기재된 화석 분류군 - 레페노마무스
레페노마무스는 백악기 전기에 살았던 세첨치아목 트라이코노돈트 포유류로, 랴오닝성 이셴 층에서 발견되었으며, 특히 R. robustus 화석에서 어린 프시타코사우루스 골격이 발견되어 중생대 포유류가 육식성이었음을 보여준다.
| 미크로랍토르 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Microraptor |
| 명명자 | Xu 외, 2000 |
| 속 | 미크로랍토르속 |
| 종 | Microraptor zhaoianus (모식종) Microraptor gui Microraptor hanqingi |
| 이명 | Cryptovolans (Czerkas 외, 2002) Tetrapterornis (Miller, 2004, 무효명) |
| 시대 | 전기 백악기, |
| 분류 | |
| 상목 | 공룡상목 |
| 목 | 용반목 |
| 목 계급 없음 | 진용반류 |
| 아목 | 수각아목 |
| 하목 | 테타누라류 |
| 하목 계급 없음 | 코엘루로사우루스류 |
| 소목 계급 없음 | 데이노니코사우루스류 |
| 과 | †드로마에오사우루스과 |
| 아과 | 미크로랍토르아과 |
2. 역사
2000년 중국 랴오닝성의 지우포탕 지층에서 화석이 발견되어 미크로랍토르라는 이름이 붙었다.[4]
1999년 내셔널 지오그래픽지에 깃털을 가진 새로운 종의 공룡인 아르카이오랍토르 랴오닝겐시스(Archaeoraptor liaoningensis)로 기재되었던 첫 표본은, 이후 합성 화석임이 밝혀졌고, 2000년에 쉬싱에 의해 '''미크로랍토르 자오이아누스'''(Microraptor zhaoianus)로 다시 기재되었다.[49][5] 종소명 ''zhaoianus''는 중국의 고생물학자 자오 시진에서 유래한다.[50]
2002년 마크 노렐 등은 BPM 1 3-13 표본을 통해 미크로랍토르가 다리에도 긴 비행 깃털을 가진 최초의 공룡이라고 설명했다.[9] 그러나 스티븐 체르카스는 미크로랍토르가 손과 팔에만 깃털이 있다고 묘사하며 노렐의 해석을 비판했다.[49]
2003년 쉬와 동료들이 발표한 '''미크로랍토르 구이'''(Microraptor gui)는 앞다리 날개와 완전히 분리된 "뒷날개"를 보여주었고, 노렐의 해석이 옳았음을 증명했다.[52] 앞다리뿐만 아니라 뒷다리에도 발달한 비행용 깃털을 가지고 있다는 점이 현생 조류와 가장 큰 차이점이다. 깃털의 형태가 현생 조류의 날개깃과 마찬가지로 좌우 비대칭을 이루고 있다는 점에서, 이 동물이 실제로 비행했을 가능성이 높다고 여겨진다.[52]
2010년 이후, 중국 랴오닝성 지우포탕 지층에서 수백 점의 ''미크로랍토르'' 표본이 발견되어, 이 지역에서 가장 풍부한 비조류 공룡 화석 중 하나가 되었다.[13]
2. 1. 명명 논란
'미크로랍토르'의 초기 명명은 첫 번째 표본이 여러 종의 화석이 섞인 키메라(chimera) 표본이었기 때문에 논란이 있었다. 이 표본은 얀오르니스, 미크로랍토르, 그리고 아직 기술되지 않은 세 번째 종의 화석을 조합하여 만들어졌으며, 중국에서 미국으로 밀수된 것이었다.[4]1999년, 내셔널 지오그래픽지는 이 표본을 '아르카이오랍토르 랴오닝겐시스(Archaeoraptor liaoningensis)'라는 새로운 종으로 기재하였다.[49] 하지만, 이후 이 표본이 합성 화석임이 밝혀졌다.[49]
2000년, 쉬싱은 이 표본의 꼬리 부분을 '미크로랍토르 자오이아누스(Microraptor zhaoianus)'로 다시 기재하였다.[5] 한편, 스미소니언 협회의 스토르스 L. 올슨은 꼬리를 제외한 부분을 '아르카이오랍토르 랴오닝겐시스'(''Archaeoraptor liaoningensis'')로 명명하며, 고생물학 기록에서 이 이름을 제거하려 시도했다.[4]
두 이름이 동일한 개체를 기준 표본으로 지정했기 때문에, ''미크로랍토르 자오이아누스''는 ''아르카이오랍토르 랴오닝겐시스''의 후배 동의어가 될 상황이었다. 그러나 올슨이 새로운 분류군을 확립하기 위한 공식적인 요구 사항을 충족했는지에 대한 의문이 제기되었다.[6]
결과적으로, 학계에서는 표본의 키메라 특성과 쉬싱의 이름(''미크로랍토르'')이 더 자주 사용된다는 점을 고려하여, "아르카이오랍토르"는 ''노멘 바눔''(부적절하게 설명되었기 때문에), ''미크로랍토르''는 ''노멘 프로테쿰''(더 많은 출판물에서 사용되었고 적절하게 설명되었기 때문에)으로 간주한다.[8]
종소명 ''zhaoianus''는 중국의 고생물학자 자오 시진에서 유래하며, 학명 전체의 의미는 "자오 씨의 작은 도둑"이다.[50]
2. 2. 추가 표본 발견
2002년 마크 노렐 등은 BPM 1 3-13 표본을 발표했지만, 이름을 붙이지는 않았다.[9] 그해 말, 스티븐 체르카스 등은 이 표본에 ''크립토볼란스 파울리''(Cryptovolans pauli)라는 이름을 붙이고, 추가로 두 개의 표본을 이 종으로 분류했다. '크립토볼란스'라는 속명은 그리스어 ''kryptos''(숨겨진)와 라틴어 ''volans''(날아가는)에서 유래했다.[15] 종명 ''pauli''는 고생물학자 그레고리 S. 폴의 이름을 딴 것이다.[15]''C. pauli''의 모식 표본은 지우포탕 지층에서 발견되었으며, 현재 중국 랴오닝성 베이퍄오 고생물 박물관에 보관되어 있다. 표본 번호는 LPM 0200(모식 표본), LPM 0201, 그리고 부모식 표본 LPM 0159이다. 이들은 비교적 완전하지만 부분적으로 손상된 압착 화석 형태로 보존되어 있다.[15]
체르카스 등(2002)은 일차 깃털(새의 특징), 융합된 흉골, 28~30개의 척추로 구성된 꼬리, 짧은 세 번째 손가락 등을 근거로 ''크립토볼란스''를 진단했다.[15] 그러나 이 특징들 중 일부는 팔이 아닌 다리에 부착된 깃털을 포함하며, ''미크로랍토르''에서도 나타난다.[10] BPM 1 3-13은 초기에 묘사된 ''미크로랍토르'' 표본보다 꼬리가 길고 꼬리 척추가 24~26개였다.[9]
이후 연구들에서 ''크립토볼란스''를 구별하는 특징들이 ''미크로랍토르'' 표본들에서도 다양하게 나타난다는 것이 밝혀졌다. 2004년 필 센터 등의 연구에서는 이러한 특징들이 단일 종 ''미크로랍토르'' 내에서의 개체 변이를 나타낸다고 보았고, ''크립토볼란스 파울리''를 ''미크로랍토르 자오이아누스''의 주니어 동의어(junior synonym)로 제안했다.[2] 앨런 페두치아와 톰 홀츠 등도 이 의견을 지지했다.[11][12]
2003년 쉬싱 등은 뒷날개와 비례적 차이를 가진 '미크로랍토르 구이(Microraptor gui)'를 발표했으나,[52] 이 역시 '미크로랍토르 자오이아누스'의 동종이명으로 보는 견해도 있다.[51]
2010년 이후, 중국 랴오닝성 지우포탕 지층에서 수백 점의 ''미크로랍토르'' 표본이 발견되어, 이 지역에서 가장 풍부한 비조류 공룡 화석 중 하나가 되었다.[13] 2010년에는 중국 박물관들에 300개 이상의 미기재된 ''미크로랍토르'' 표본이 보고되었지만, 상당수는 개인 수집가에 의해 변형되거나 합성된 것이었다.[37]
2. 3. 연구 및 논쟁
노렐 등(2002)은 BPM 1 3-13 표본을 통해 미크로랍토르가 다리에도 긴 비행 깃털을 가진 최초의 공룡이라고 설명했다.[49] 그러나 체르카스(2002)는 저서 《깃털 공룡과 비행의 기원》 표지에 그림을 싣고, 미크로랍토르가 손과 팔에만 깃털이 있다고 묘사하며 노렐의 해석을 비판했다. 그는 노렐이 "자신이 믿고 싶어하는 것을 보기 때문"에 뒷다리의 날개깃털을 잘못 해석했다고 주장했다. 또한 노렐이 드로마에오사우루스가 새라는 결론을 내리지 못한 것에 대해 "선입견의 맹목적인 영향"에 굴복했다고 비난했다.[49]쉬와 동료들(2003)이 발표한 추가적인 《미크로랍토르》 표본은 앞다리 날개와 완전히 분리된 "뒷날개"를 보여주었고, 노렐의 해석이 옳았음을 증명했다. 쉬는 이 표본들을 《미크로랍토르》 표본과 뒷날개 및 비례적 차이가 있는 새로운 종으로 간주하여 《미크로랍토르 구이》라고 명명했지만, 센터는 이러한 분류에 의문을 제기하며 대부분의 차이점이 크기, 즉 나이 차이에 해당한다고 보았다.[49]
체르카스는 융합된 흉골과 비대칭 깃털을 근거로 《시조새》보다 《미크로랍토르》가 더 발전된 현대 조류의 특징을 가졌다고 주장하며, 《미크로랍토르》가 더 잘 비행할 수 있었을 것이라고 추정했다. 그는 드로마에오사우루스과가 실제로 기저 조류 그룹일 수 있으며, 《데이노니쿠스》와 같은 더 크고 나중의 종은 2차적으로 비행 능력을 상실했을 것이라는 가설을 제시했다.[49] 그러나 현재 학계에서는 드로마에오사우루스과가 일부 공기역학적 능력을 가진 조상으로부터 진화했는지 결론 내릴 증거가 충분하지 않다는 것이 중론이다. 쉬 등(2003)은 기저 드로마에오사우루스가 작고, 수목성이며, 활강할 수 있었을 것이라고 제안했지만, 터너 등(2007)은 조상 드로마에오사우루스가 활강하거나 비행할 수 없었지만, 몸집이 작았다(약 65cm 길이, 질량 600~700g)는 좋은 증거가 있다고 제안했다.[49]
1999년 내셔널 지오그래픽지에 깃털을 가진 새로운 종의 공룡인 아르카이오랍토르 랴오닝겐시스로 기재되었던 첫 표본은, 이후 합성 화석임이 밝혀졌고, 2000년에 '''미크로랍토르 자오이아누스'''로 다시 기재되었다.[49]
조류를 제외한 공룡으로서는 매우 소형으로, 그레고리 폴은 그 전체 길이를 0.7미터로 하였다.[51] 종소명 ''zhaoianus''는 중국의 고생물학자 자오 시진에서 유래한다.[50]
2003년에 기재된 '''미크로랍토르 구이'''(顧氏小盜龍|구스 샤오다오룽|고씨소도룡중국어)의 화석에는 발달한 깃털이 보존되어 있었다. 앞다리뿐만 아니라 뒷다리에도 발달한 비행용 깃털을 가지고 있다는 점이 현생 조류와 가장 큰 차이점이다. 깃털의 형태가 현생 조류의 날개깃과 마찬가지로 좌우 비대칭을 이루고 있다는 점에서, 이 동물이 실제로 비행했을 가능성이 높다고 여겨진다.[52]
3. 특징
''미크로랍토르''(Microraptor)는 가장 작은 비조류 공룡 중 하나로, ''M. gui''의 모식 표본은 길이 77cm, 날개 길이 88cm~94cm, 무게 0.5kg~1.4kg으로 측정되었다.[36][17][34] 더 큰 표본들은 길이 최소 80cm, 날개 길이 99cm 이상, 무게 1.25kg~1.88kg이었을 것이다.[34] 미크로랍토르는 극도로 작은 크기 외에도 깃털과 날개의 흔적이 발견된 최초의 비조류 공룡 중 하나였다. 초기 조류와 깃털 달린 공룡 중에서도 특이하게, ''미크로랍토르''는 앞다리와 뒷다리 모두에 긴 비행 깃털을 가진 몇 안 되는 조류 조상 중 하나이다. 몸은 두꺼운 깃털로 덮여 있었고, 꼬리 끝에는 다이아몬드 모양의 부채꼴 깃털이 있어 비행 중 안정성을 높이는 역할을 했을 것으로 추정된다.[18]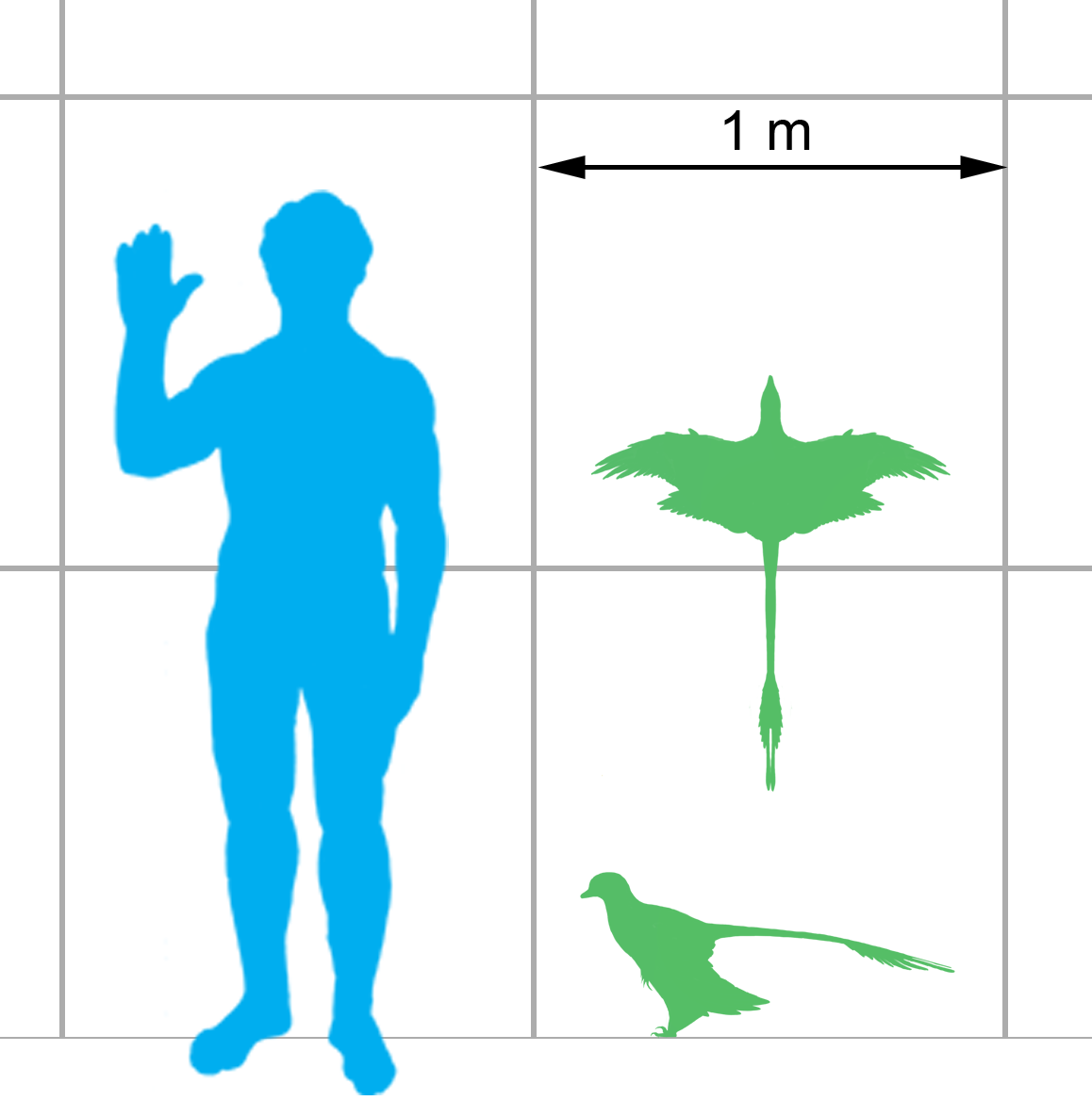
2012년 리고앙구오 외 연구진은 새로운 표본 BMNHC PH881을 토대로 ''미크로랍토르''의 깃털 색상을 주사 전자 현미경 기술을 사용하여 화석화된 멜라노솜(색소 세포)을 분석하고 현대 조류의 멜라노솜 배열과 비교했다. ''미크로랍토르''의 멜라노솜은 현대 조류에서 검고 광택이 나는 색상과 일치하는 방식으로 배열되어 있었다. 이러한 막대 모양의 좁은 멜라노솜은 현대의 찌르레기와 매우 유사하게 겹겹이 쌓여 배열되어, ''미크로랍토르''의 깃털에서 무지개빛을 나타냈다.[19]
미크로랍토르는 톱니가 없거나 부분적으로만 톱니가 있는 이빨, 잘록한 '허리', 그리고 유난히 긴 위팔뼈 등 원시 조류 및 원시 트로오돈과와 공유하는 해부학적 특징을 가지고 있다. 특히 기저 트로오돈과인 ''시노베나토르''와 유사성이 높다.
4. 분류
미크로랍토르는 드로마이오사우루스과(Dromaeosauridae) 미크로랍토리아(Microraptoria)에 속한다. 미크로랍토리아에는 톈위랍토르(Tianyuraptor), 헤스페로니쿠스(Hesperonychus), 시노르니토사우루스(Sinornithosaurus), 그라실리랍토르(Graciliraptor), 창위랍토르(Changyuraptor) 등이 있다. 2024년 연구에서는 미크로랍토리아와 진드라코랍토르류를 포함하는 새로운 분기군인 세라랍토리아(Serraraptoria)가 제안되었다.[22]
2012년 필 센터 등이 실시한 분석에 따른 분기도는 다음과 같다.[21]
미크로랍토르의 첫 표본은 1999년 내셔널 지오그래픽지에 깃털을 가진 새로운 종의 공룡인 "아르카이오랍토르 랴오닝겐시스"로 기재되었다. 복원 표본이 표지를 장식하며 단기간 유명해졌지만, 이후 이 표본은 두 동물 화석을 조합한 "합성 화석"으로 밝혀졌다. 2000년 원래 표본의 하반신은 드로마이오사우루스과 신속 신종 '''미크로랍토르 자오이아누스'''로 다시 기재되었다. 종소명 ''zhaoianus''는 중국 고생물학자 자오 시진에서 유래하며, 학명 전체는 "자오 씨의 작은 도둑"을 의미한다.[50]
2003년 기재된 '''미크로랍토르 구이''' (顧氏小盜龍|Gùshì xiǎodàolóng중국어, 고씨소도룡) 화석에는 발달된 깃털이 보존되어 있었다. 미크로랍토르 구이와 현생 조류의 가장 큰 차이점은 앞다리뿐 아니라 뒷다리에도 발달한 비행용 깃털이 있다는 것이다. 뒷다리 깃털의 발달 방식은 불확실하지만, 깃털 형태가 현생 조류 날개깃처럼 좌우 비대칭이어서 실제로 비행했을 가능성이 높다고 여겨진다.[52] 날개 펼치는 방법에 대해서는 앞다리 날개를 수평으로 펼치고 뒷다리 깃털을 비스듬히 펼치는 설과, 앞다리 날개와 평행하게 뒷다리를 펼치는 설이 있다.[53]
미크로랍토르 자오이아누스는 조류를 제외한 공룡 중 매우 소형이다.[49] 그레고리 폴은 전체 길이를 0.7미터로 추정했고[51], '공룡 박람회 2006' 공식 도록은 성체 개체 길이를 약 40cm로 제시했다.[49] 다만, 같은 전시회에서 63cm에 달하는 대형 표본도 전시되었다.[49]
미크로랍토르아과에 속하는 캉귤라프토르와 비교하면 두개골은 작고 삼각형이며,[51] 두껍고 톱니가 잘 보이지 않는다.[51] 목은 다소 짧고, 흉골은 유합되어 1장의 판상 구조를 이룬다.[51] 뒷다리 발톱은 강하게 굽어져 수목 생활을 했을 것으로 추정되며,[51] 이빨 형태로 보아 소형 먹이를 포식했을 것으로 보인다.[51] 화석은 중국 북동부 주포탕 층에서 발견되며, 호수가 있는 습윤한 숲 환경에서 살았을 것으로 추정된다.[51] 겨울은 춥고 눈이 많이 내렸다고 한다.[51]
미크로랍토르 구이는 미크로랍토르 자오이아누스의 주니어 시노님(junior synonym)으로 간주되기도 한다.[51]
5. 고생물학
미크로랍토르는 앞다리와 뒷다리에 모두 비행 깃털을 가진 '네 날개' 구조를 가지고 있었으며, 이는 오늘날 탠덤익기 항공기의 비행 표면 배열과 유사하다. 팔과 손에는 10cm에서 20cm 길이의 긴 깃털이 있었고, 다리와 발에는 11cm에서 15cm 길이의 깃털이 있었다. 뒷다리의 긴 깃털은 현대 조류처럼 진정한 비행 깃털이었으며, 팔, 다리, 꼬리 깃털에 비대칭적인 깃대가 있었다. 현대 조류의 날개처럼 주깃(손에 고정)과 부깃(팔에 고정)을 모두 가지고 있었고, 뒷다리에도 비행 깃털이 발 윗부분 뼈와 위·아래 다리에 고정되어 있었다.[23]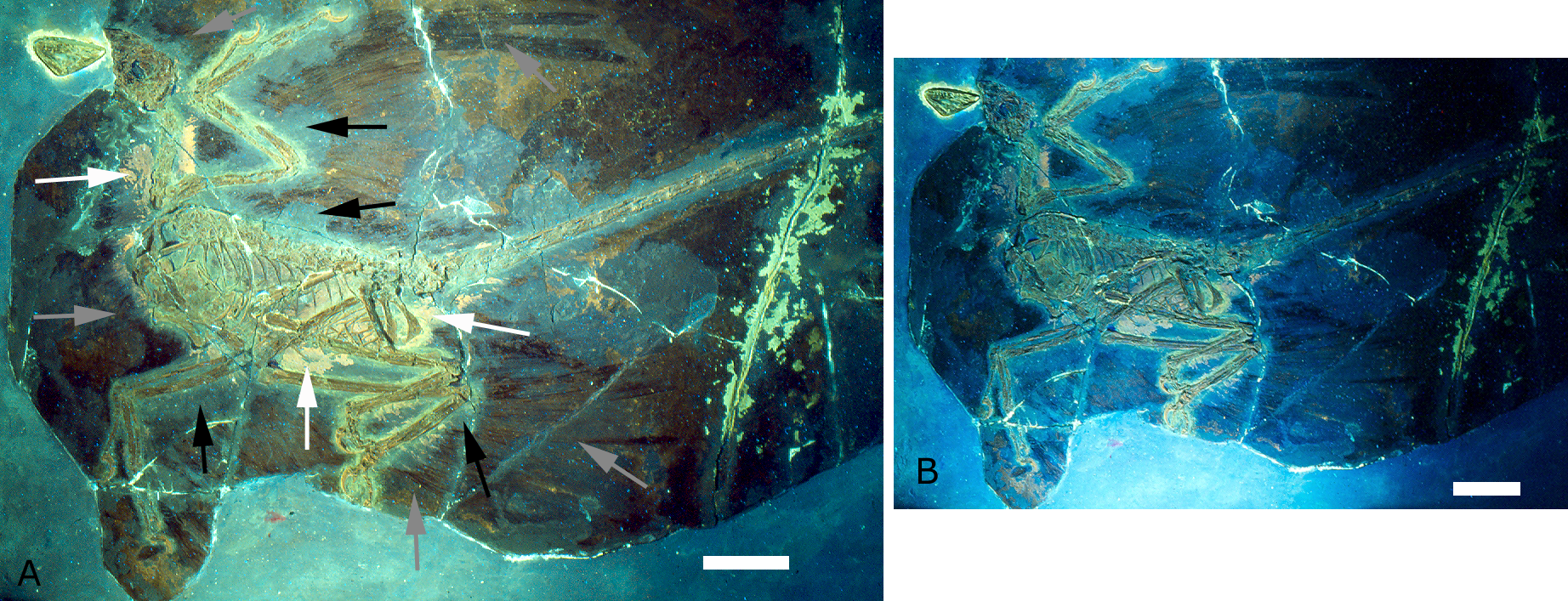
미크로랍토르에서 발견된 독특한 날개 배열은 현대 조류의 비행 진화가 4개의 날개 단계를 거쳤는지, 아니면 미크로랍토르와 같은 4개의 날개를 가진 활공 생물이 후손을 남기지 못한 진화적 측면 가지였는지에 대한 의문을 제기했다. 1915년 자연사학자 윌리엄 비비는 조류의 비행 진화가 4개의 날개(또는 테트라프테릭스) 단계를 거쳤을 수 있다고 주장했다.[42]
2003년에 기재된 미크로랍토르 구이(顧氏小盜龍|구스샤오다오룽중국어)의 화석에는 발달한 깃털이 보존되어 있었다. 미크로랍토르 구이와 현재 조류의 가장 큰 차이점은 앞다리뿐만 아니라 뒷다리에도 발달한 비행용 깃털을 가지고 있다는 것이다. 뒷다리 깃털이 어떻게 발달했는지는 확실하지 않지만, 깃털의 형태가 현생 조류의 날개깃과 마찬가지로 좌우 비대칭을 이루고 있다는 점에서, 이 동물이 실제로 비행했을 가능성이 높다고 여겨진다.[52]
미크로랍토르는 뒷날개가 발목까지 뻗어 있고, 앞날개 깃털 또한 길어서 지상에서 이동하거나 달리는 데 어려움이 있었을 것으로 추측된다.
5. 1. 비행 능력
미크로랍토르는 앞다리와 뒷다리에 모두 비행 깃털을 가진 '네 날개' 구조를 가지고 있었으며, 이는 오늘날 탠덤익기 항공기의 비행 표면 배열과 유사하다. 팔과 손에는 10cm에서 20cm 길이의 긴 깃털이 있었고, 다리와 발에는 11cm에서 15cm 길이의 깃털이 있었다. 뒷다리의 긴 깃털은 현대 조류처럼 진정한 비행 깃털이었으며, 팔, 다리, 꼬리 깃털에 비대칭적인 깃대가 있었다. 현대 조류의 날개처럼 주깃(손에 고정)과 부깃(팔에 고정)을 모두 가지고 있었고, 뒷다리에도 비행 깃털이 발 윗부분 뼈와 위·아래 다리에 고정되어 있었다.[23]원래 미크로랍토르는 활공 비행을 하는 동물로, 주로 나무에서 살았을 것으로 추정되었는데, 뒷날개가 땅 위를 걷는 능력을 방해했을 것이기 때문이다.[24] 일부 고생물학자들은 깃털 공룡들이 활공이나 진정한 비행의 전조로서 나무에서 낙하산을 이용하여 땅 위의 먹이를 공격하거나 매복했을 가능성을 제기했다.[25] 2007년 연구에서 미크로랍토르의 결합된 날개 표면이 낙하산으로 안전하게 땅에 착륙하기에는 너무 좁아서 높은 곳에서는 부상을 입을 수 있다는 것이 밝혀졌지만, 짧은 거리를 낙하산 활강했을 가능성은 열어두었다.[36][25] 풍동 실험 결과 낮은 고도 사이를 활공할 때 높은 항력을 감수하면서 높은 양력 계수를 유지하는 것이 미크로랍토르에게 가장 효율적인 전략이었을 가능성이 입증되었다.[26] 하지만 미크로랍토르가 수목성 활공기였다는 생각은 이 동물이 정기적으로 나무를 오르거나 심지어 나무에서 살았다는 것에 의존하는데, 해부학 연구에 따르면 이 동물의 사지 비율은 등반가보다는 현대 지상 조류에 가깝고, 골격은 나무를 오르는 데 특화된 동물에게서 기대되는 적응의 흔적을 보이지 않는다.[27][28]
고생물학자 스티븐 체르카스는 ''미크로랍토르''(''Cryptovolans pauli'')가 동력 비행을 했을 수 있으며, 시조새보다 더 나은 비행 능력을 가졌을 수 있다고 주장했다. 그는 미크로랍토르의 융합된 흉골, 비대칭 깃털, 어깨띠의 특징이 단순한 활공이 아니라 자체 동력으로 비행할 수 있음을 나타낸다고 언급했다.[29] 오늘날 대부분의 과학자들은 미크로랍토르가 비행 동물에게서 기대되는 해부학적 특징을 가지고 있었지만, 조류에 비해 덜 발달된 형태의 비행이었을 것이라는 점에 동의한다. 예를 들어, 일부 연구에서는 어깨 관절이 완전한 날갯짓 비행 스트로크를 허용하기에는 너무 원시적이었다고 제안한다. 수각류 공룡의 조상 해부학에서 어깨 소켓은 아래쪽과 약간 뒤쪽을 향하고 있어서 동물이 팔을 수직으로 올리는 것이 불가능했는데, 이는 조류의 날갯짓 비행 스트로크에 필수적인 조건이다. 수각류 해부학 연구에서는 어깨 소켓이 척추에 가까운 높고 위쪽으로 향하는 조류와 같은 위치로 이동한 것은 엔안티오르니테스와 같은 비교적 진보된 조류가 나타난 이후라는 것을 시사했다.[29] 하지만, 다른 과학자들은 미크로랍토르를 포함한 일부 파라베스 수각류의 어깨띠가 어깨 관절이 등 윗부분에만 위치할 수 있도록 굴곡져 있어 날개의 거의 수직적인 상승 스트로크가 가능했을 것이라고 주장했다. 이 잠재적으로 진보된 어깨 해부학은 손목을 어깨에 연결하는 피막과 알룰라와 결합하여 미크로랍토르가 진정한 동력 비행을 할 수 있었음을 나타낼 수 있다.[30]
다른 연구에서는 미크로랍토르의 날개가 완전한 수직 비행 스트로크 없이도 동력 비행에 필요한 양력을 생성할 수 있을 만큼 충분히 컸음을 입증했다. 파라베스의 초기 비행 능력을 연구한 2016년 연구에서는 미크로랍토르가 날개 보조 경사 달리기뿐만 아니라 날개 보조 도약 및 지상 기반 이륙도 할 수 있었다는 것을 보여주었다.[28]
스티븐 체르카스, 그레고리 S. 폴 등은 미크로랍토르가 비행할 수 있으면서도 드로마이오사우루스과라는 사실은 드로마이오사우루스과 (나중에 더 큰 종인 데이노니쿠스 포함)가 2차적으로 비행 능력을 상실했음을 시사한다고 주장했다. 쉬와 동료들의 연구 또한 드로마이오사우루스과의 조상이 작고 수목성이며 활공 비행을 할 수 있었을 것이라고 시사했지만, 활공에 적합하지 않은 짧은 앞다리를 가진 더 원시적인 드로마이오사우루스과가 발견되면서 이러한 견해에 의문이 제기되었다.[24][31] 하지만, 다른 파라베스의 비행 능력에 대한 연구는 그들 대부분이 제한된 비행 스트로크와 비교적 작은 날개 때문에 동력 비행에 충분한 양력을 얻지 못했을 것이라는 것을 보여주었다. 이러한 연구는 미크로랍토르가 조류의 조상과 독립적으로 비행과 관련 특징(융합된 흉골, 알룰라 등)을 진화시켰을 것이라고 결론지었다.[28][32][33][34] 2024년, 키아와 오코너는 중생대 조류와 미크로랍토르가 현대 비행 조류와 일치하는 깃털 형태를 가지고 있는 반면, 안키오르니티데와 카우디프테릭스는 2차적으로 비행 능력을 상실했다고 분석했다.[35]
미크로랍토르에서 발견된 독특한 날개 배열은 현대 조류의 비행 진화가 4개의 날개 단계를 거쳤는지, 아니면 미크로랍토르와 같은 4개의 날개를 가진 활공 생물이 후손을 남기지 못한 진화적 측면 가지였는지에 대한 의문을 제기했다. 1915년 자연사학자 윌리엄 비비는 조류의 비행 진화가 4개의 날개(또는 테트라프테릭스) 단계를 거쳤을 수 있다고 주장했다.[42] 채터지(Chatterjee)와 템플린(Templin)은 이러한 가능성에 대해 강한 입장을 취하지 않고, 전통적인 해석과 테트라프테릭스 단계 모두 동등하게 가능하다고 언급했다. 그러나 다양한 깃털 공룡, 시조새 및 맹금류와 같은 일부 현대 조류에서 비정상적으로 긴 다리 깃털의 존재를 바탕으로, 그리고 발에 긴 1차 깃털을 가진 공룡(예: 페도페나)의 추가 발견을 통해, 저자들은 형태학과 계통 발생학 모두에서 현재의 증거가 조류 비행이 어느 시점에서 공유된 사지 지배에서 앞다리 지배로 전환되었음을 시사하며, 모든 현대 조류가 4개의 날개를 가진 조상 또는 적어도 현대 구성과 비교하여 비정상적으로 긴 다리 깃털을 가진 조상으로부터 진화했을 수 있다고 주장했다.[36]
2003년에 기재된 미크로랍토르 구이(顧氏小盜龍|구스샤오다오룽중국어)의 화석에는 발달한 깃털이 보존되어 있었다. 미크로랍토르 구이와 현재 조류의 가장 큰 차이점은 앞다리뿐만 아니라 뒷다리에도 발달한 비행용 깃털을 가지고 있다는 것이다. 뒷다리 깃털이 어떻게 발달했는지는 확실하지 않지만, 깃털의 형태가 현생 조류의 날개깃과 마찬가지로 좌우 비대칭을 이루고 있다는 점에서, 이 동물이 실제로 비행했을 가능성이 높다고 여겨진다.[52] 날개를 펼치는 방법에 대해서는 앞다리 날개를 수평으로 펼치고 뒷다리 깃털을 비스듬히 펼치는 설과, 앞다리 날개와 평행하게 뒷다리를 펼치는 설이 있다.[53]
5. 2. 뒷날개 자세
상카르 차터지는 2005년에 ''미크로랍토르''가 활공하거나 날기 위해서는 앞날개와 뒷날개가 서로 다른 수준에 있어야 하며(마치 복엽기처럼), 겹쳐져서는 안 된다(마치 잠자리처럼)고 주장했고, 후자의 자세는 해부학적으로 불가능했을 것이라고 말했다. 차터지는 이 복엽기 모델을 사용하여 활공 가능한 방법을 계산할 수 있었고, ''미크로랍토르''가 대부분 ''푸고이드'' 스타일의 활공을 사용했을 가능성이 높다고 결론 내렸다. 즉, 높은 곳에서 뛰어내리면 동물은 깊은 U자형 곡선으로 아래로 급강하한 다음 다시 날아올라 다른 나무에 착지했을 것이다. 복엽기 날개 구조에 직접 사용되지 않은 깃털, 예를 들어 경골과 꼬리의 깃털은 항력을 제어하고 비행 경로, 탄도 등을 변경하는 데 사용되었을 수 있다. 뒷날개의 방향 또한 동물이 활공 비행을 제어하는 데 도움이 되었을 것이다. 차터지는 또한 컴퓨터 알고리즘을 사용하여 동물의 비행 능력을 테스트하여 ''미크로랍토르''가 수동적인 활공 외에 진정한 동력 비행이 가능한지 여부를 테스트했다. 그 결과 데이터는 ''미크로랍토르''가 수평 동력 비행을 유지하는 데 필요한 조건을 갖추고 있음을 보여주었고, 따라서 동물이 활공 대신 비행했을 가능성도 이론적으로 존재한다.[36]일부 고생물학자들은 복엽기 가설에 의문을 제기하며 다른 구성을 제안해 왔다. 2010년 알렉산더(Alexander) 등이 수행한 연구에서는 활공 테스트를 수행하기 위해 사용된 가벼운 3차원 물리 모델의 제작을 설명했다. 이들은 모델에 여러 뒷다리 구성을 사용하여 복엽기 모델이 부자연스럽지는 않지만 구조적으로 결함이 있으며 안정적인 활공을 위해서는 머리가 무거운 무게 분포가 필요했는데, 이는 가능성이 낮다고 판단했다. 이 연구는 측면으로 벌어진 뒷날개 구조가 ''미크로랍토르''에게 가장 생물학적 및 공기역학적으로 일관된 구성임을 나타냈다.[37] 그러나 브로엄(Brougham)과 브루사테(Brusatte)의 추가 분석에서는 알렉산더의 모델 재구성이 ''미크로랍토르''에 대한 모든 가용 데이터와 일치하지 않으며, 이 연구가 ''미크로랍토르''의 가능한 비행 패턴을 결정하기에 충분하지 않다고 결론지었다. 브로엄과 브루사테는 알렉산더와 그의 팀이 사용한 모델의 해부학을 비판하며, 엉덩이 해부학이 다른 드로마에오사우루스류와 일치하지 않는다고 지적했다. 대부분의 드로마에오사우루스과에서는 엉덩이 뼈의 특징으로 인해 다리가 수평으로 벌어지는 것을 방지한다. 대신 다리는 몸 아래 수직 위치에 고정된다. 알렉산더의 팀은 평평하게 짓눌린 ''미크로랍토르'' 표본을 사용하여 모델을 만들었는데, 브로엄과 브루사테는 이것이 실제 해부학을 반영하지 못한다고 주장했다.[38] 2010년 후반에 알렉산더의 팀은 이러한 비판에 대응하며, 3차원으로 보존된 완전한 엉덩이 뼈로 알려진 관련 드로마에오사우루스 ''Hesperonychus'' 역시 엉덩이 소켓이 부분적으로 위를 향하고 있어 다른 드로마에오사우루스류보다 다리가 더 많이 벌어질 수 있다고 지적했다.[39] 그러나 하트먼(Hartman)과 동료들은 2019년 계통 발생학적 분석을 기반으로 ''Hesperonychus''가 드로마에오사우루스류가 아니라 현대 조류와 유사한 아비알레에 속하며 ''Balaur bondoc''에 가깝다고 제안했다.[40]
5. 3. 지상 보행
미크로랍토르는 뒷날개가 발목까지 뻗어 있고, 앞날개 깃털 또한 길어서 지상에서 이동하거나 달리는 데 어려움이 있었을 것으로 추측된다. 2010년 코윈 설리번(Corwin Sullivan) 등의 연구에 따르면, 날개를 최대한 접더라도 팔을 중립적인 위치에 두거나 먹이를 공격할 때처럼 앞으로 뻗으면 깃털이 땅에 끌렸을 것이라고 한다. 미크로랍토르가 날개 깃털 손상을 피하려면 날개를 들어 올리거나 위팔을 완전히 뒤로 젖혀야만 했을 것이다. 따라서 발톱 달린 앞다리로 먹이를 잡거나 물체를 조작하는 것은 불가능했을 것으로 보인다.[41]5. 4. 식성
2010년 연구자들은 ''미크로랍토르 자오이아누스''(M. zhaoianus)의 모식 표본을 추가로 준비했고, 그 결과 보존된 것으로 추정되는 내장 내용물을 발견했다. 2022년 데이비드 혼(David Hone)과 동료들이 이 내용물에 대한 연구를 발표했는데, 여기에는 포유류의 유해가 포함되어 있었다. 유해는 주로 완전하고 연결된 오른쪽 발(모든 발목뼈, 중족골, 대부분의 발가락뼈 포함)과 추가적인 긴 뼈의 줄기, 그리고 잠재적으로 다른 조각들이었다. 발 골격은 ''에오마이아(Eomaia)''와 ''시노델피스(Sinodelphys)''의 발과 유사하며, 80mm의 주둥이에서 항문까지의 길이와 13g에서 43g의 질량을 가진 동물에 해당한다. 발의 발톱은 ''에오마이아''나 ''시노델피스''보다 덜 구부러져 있는데, 이는 이 포유류가 나무를 오를 수는 있지만 두 속보다는 덜 효과적이었고, 따라서 수목형이라기보다는 잠재적으로 활주형이었을 가능성이 있음을 나타낸다.[43][44]포유류가 ''미크로랍토르''에게 잡아먹혔는지, 아니면 단순히 시체를 청소한 것인지는 불분명하다. 하지만 섭취된 다른 명확한 신체 부위가 없다는 것은 근육량이 적은 발이 시체 섭취의 후기 단계에서, 아마도 시체 청소를 통해 먹혔을 수 있음을 시사한다. 이 발견은 수생 포유류를 확실히 섭취한 수각류의 드문 예시이며, 유일한 다른 예시는 콤프소그나투스과의 ''시노사우롭테릭스(Sinosauropteryx)'' 표본이다.[43][44]
2011년 12월 6일 ''국립 과학원 회보''(Proceedings of the National Academy of Sciences)에 실린 논문에서, 징마이 오코너(Jingmai O'Connor)와 공동 저자들은 ''미크로랍토르 구이''(Microraptor gui) 표본을 묘사했다. 이 표본의 복부에는 나무에서 사는 에난티오르니테스류 새의 뼈, 특히 부분적인 날개와 발이 포함되어 있었다. 뼈의 위치는 새가 통째로 머리부터 삼켜졌음을 의미하며, 저자들은 이를 통해 ''미크로랍토르''가 시체 청소를 하는 대신 나무에서 새를 잡아서 먹었다는 것을 암시한다고 해석했다.[45]
2013년 연구자들은 다른 ''M. gui'' 표본의 복강에서 물고기 비늘을 발견했다고 발표했다.[46] 저자들은 ''M. gui''가 수목 환경에서만 사냥했다는 이전의 제안에 반박하며, 물고기도 잘 사냥하는 동물이라고 제안했다. 또한 이 표본이 물고기를 먹는 식단에 적응했을 가능성이 있다고 주장하며, 턱의 처음 세 개의 이가 앞쪽 위쪽으로 기울어져 있는 점을 지적했는데, 이는 어식성과 자주 연관되는 특징이다.[46] 그들은 ''미크로랍토르''가 기회주의적 포식자이며, 수목 서식지와 수생 서식지 모두에서 가장 흔한 먹이를 사냥했다고 결론 내렸다.[46]
이 두 연구 모두 각 내장 내용을 포식의 예시로 간주했다. 그러나 혼(Hone)과 동료들(2022)은 이러한 해석의 신뢰성에 의문을 제기하며, 둘 다 시체 청소로 설명될 수 있다고 썼다. 또한, 그들은 다양한 척추동물 내장 내용물(예: 물고기, 포유류, 도마뱀, 새)을 일반적인 사냥 전략의 증거로 인용하고, 두 서식지 모두에서 사냥을 위한 전문화가 필요하지 않다는 점을 들어 ''미크로랍토르''가 수목이나 수생 환경, 또는 둘 다에 특화된 사냥꾼이라는 주장에 반박했다.[44]
2019년, 새로운 속의 경설류 도마뱀(''인드라사우루스(Indrasaurus)'')가 ''미크로랍토르''의 위에서 발견된 표본에서 묘사되었다. ''미크로랍토르''는 사냥감을 머리부터 삼킨 것으로 보이는데, 이는 현대의 육식성 조류와 도마뱀의 일반적인 행동이다. ''인드라사우루스'' 뼈에는 뚜렷한 구멍과 흉터가 없었는데, 이는 ''미크로랍토르''가 도마뱀을 먹은 직후, 상당한 소화가 일어나기 전에 죽었음을 나타낸다.[47]
동족 파라베스류인 ''안키오르니스(Anchiornis)''와 달리, ''미크로랍토르''는 위 내용물을 보존하고 있는 네 개의 ''미크로랍토르'' 표본이 존재함에도 불구하고, 알약 형태로 발견된 적이 없다. 이는 ''미크로랍토르''가 소화되지 않는 털, 깃털, 뼈 조각을 알약 대신 배설물로 배출했음을 시사한다.[47]
6. 대중문화
BBC 다큐멘터리 《공룡 vs 포유류》 후편에서 공룡에서 조류로의 진화를 설명하는 역할을 맡았다.[1] 《플래닛 다이노소어》에서도 조류 진화를 설명하며, 포식자로서는 활공성 도마뱀을 쫓고 피식자로서는 시노르니토사우루스에게 쫓기는 모습으로 등장한다.[3] 《프레히스토릭 파크》 3회~최종회에서는 화산 지대 숲에서 티타노사우루스류(이시사우루스)에 달라붙어 생활하는 모습이 그려졌다.[2]
참조
[1]
논문
A new phylogeny of Dromaeosauridae
https://core.ac.uk/r[...]
2004
[2]
논문
Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda)
[3]
논문
Rarity of molt evidence in early pennaraptoran dinosaurs suggests annual molt evolved later among Neornithes
[4]
간행물
Countdown to Piltdown at ''National Geographic'': the rise and fall of ''Archaeoraptor''
Backbone
2000
[5]
간행물
The smallest known non-avian theropod dinosaur
http://research.amnh[...]
Nature
2000
[6]
웹사이트
Archaeoraptor still a nomen nudum
http://dml.cmnh.org/[...]
2001-01-04
[7]
웹사이트
So why hasn't Tyrannosaurus been renamed Manospondylus?
http://www.miketaylo[...]
2002-08-27
[8]
웹사이트
Archaeoraptor v Microraptor
http://dml.cmnh.org/[...]
2001-01-01
[9]
간행물
'Modern' feathers on a non-avian dinosaur
Nature
2002-03-07
[10]
논문
Four-winged dinosaurs from China
http://doc.rero.ch/r[...]
[11]
문서
Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
http://www.geol.umd.[...]
2011
[12]
논문
Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence
[13]
논문
Non-Avian dinosaur fossils from the Lower Cretaceous Jehol Group of western Liaoning, China
[14]
논문
'Modern' feathers on a non-avian dinosaur
[15]
서적
Feathered Dinosaurs and the Origin of Flight
The Dinosaur Museum
2002-08-01
[16]
논문
Four-winged dinosaurs from China
http://doc.rero.ch/r[...]
[17]
논문
A new tiny dromaeosaurid dinosaur from the Lower Cretaceous Jehol Group of western Liaoning and niche differentiation among the Jehol dromaeosaurids
http://www.ivpp.cas.[...]
[18]
간행물
New Specimens of ''Microraptor zhaoianus'' (Theropoda: Dromaeosauridae) from Northeastern China
American Museum Novitates
2002
[19]
논문
Reconstruction of ''Microraptor'' and the Evolution of Iridescent Plumage
2012
[20]
뉴스
Feathers Worth a 2nd Look Found on a Tiny Dinosaur
https://www.nytimes.[...]
2012-04-22
[21]
논문
New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail
[22]
논문
The smallest known specimen of ''Microraptor'' (Dinosauria: Dromaeosauridae) from the Jiufotang Formation in northeastern China
[23]
논문
The extent of the preserved feathers on the four-winged dinosaur ''Microraptor gui'' under ultraviolet light
[24]
간행물
Four-winged dinosaurs from China
Nature
2003-01-23
[25]
서적
Prehistoric Life
Dorling Kindersley
[26]
논문
Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight
[27]
논문
Assessing Arboreal Adaptations of Bird Antecedents: Testing the Ecological Setting of the Origin of the Avian Flight Stroke
2011-08-09
[28]
논문
The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents
[29]
논문
Scapular orientation in theropods and basal birds, and the origin of flapping flight
[30]
논문
Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, ''Anchiornis'' and Scansoriopterygidae
[31]
논문
A basal dromaeosaurid and size evolution preceding avian flight
[32]
논문
A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight
2019-07-10
[33]
논문
Potential for Powered Flight Neared by Most Close Avialan Relatives, but Few Crossed Its Thresholds
2020
[34]
논문
High flyer or high fashion? A comparison of flight potential among small-bodied paravians
http://digitallibrar[...]
2020
[35]
논문
Functional constraints on the number and shape of flight feathers
2024-02-20
[36]
논문
Biplane wing planform and flight performance of the feathered dinosaur ''Microraptor gui''
http://www.pnas.org/[...]
[37]
논문
Model tests of gliding with different hindwing configurations in the four-winged dromaeosaurid ''Microraptor gui''
[38]
논문
Distorted ''Microraptor'' specimen is not ideal for understanding the origin of avian flight
[39]
논문
Reply to Brougham and Brusatte: Overall anatomy confirms posture and flight model offers insight into the evolution of bird flight
[40]
논문
A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight
2019-07-10
[41]
논문
The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs
[42]
논문
Tetrapteryx stage in the ancestry of birds
[43]
간행물
"THE WINGED NON-AVIAN DINOSAUR MICRORAPTOR FED ON MAMMALS: IMPLICATIONS FOR THE JEHOL BIOTA ECOSYSTEM"
[44]
논문
Generalist diet of ''Microraptor zhaoianus'' included mammals
https://zenodo.org/r[...]
[45]
논문
Additional specimen of ''Microraptor'' provides unique evidence of dinosaurs preying on birds
[46]
논문
Piscivory in the feathered dinosaur ''Microraptor''
[47]
논문
Microraptor with Ingested Lizard Suggests Non-specialized Digestive Function
2019-07-11
[48]
논문
Nocturnality in dinosaurs inferred from scleral ring and orbit morphology
[49]
서적
世界の巨大恐竜博2006 生命と環境─進化のふしぎ
"[[日本経済新聞社]]、[[日本放送協会|NHK]]、[[NHKプロモーション]]、[[日経ナショナルジオグラフィック社]]"
[50]
서적
語源が分かる恐竜学名辞典
"[[北隆館]]"
2017-01-20
[51]
서적
グレゴリー・ポール恐竜事典 原著第2版
"[[共立出版]]"
2020-08-31
[52]
서적
地球を支配した恐竜と巨大生物たち
"[[日経サイエンス社]]"
[53]
서적
鳥類学者 無謀にも恐竜を語る
新潮社
2018-07-10
[54]
웹사이트
玉虫色の輝き、ミクロラプトルの羽毛
http://natgeo.nikkei[...]
日経ナショナルジオグラフィック社
2023-11-30
[55]
서적
공룡대백과
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com