바다나리
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
바다나리는 극피동물에 속하는 해양 무척추동물로, 고대 그리스어 '백합'에서 유래된 이름처럼 백합과 유사한 형태를 지닌다. 몸은 줄기, 컵 모양의 중앙 몸체, 팔로 구성되며, 입과 항문은 위쪽에 위치한다. 바다나리는 주로 플랑크톤을 걸러 먹는 여과 섭식자이며, 팔을 이용해 먹이를 포획한다. 화석 기록은 오르도비스기부터 나타나며, 다양한 멸종과 적응 과정을 거쳐 현재 약 650여 종이 존재한다. 바다나리는 줄기 유무, 팔의 형태 등에 따라 여러 목으로 분류되며, 일부 화석은 장신구나 문화적 상징으로 사용되기도 한다.
더 읽어볼만한 페이지
- 고생대의 무척추동물 - 필석
필석은 캄브리아기부터 석탄기 지층에서 화석으로 발견되는 고생대 초기의 멸종된 해양 동물 군체로, 반삭동물로 분류되며, 지질 시대 구분에 중요한 표준 화석으로 활용되어 오르도비스기와 실루리아기 지층 연구에 기여한다. - 고생대의 무척추동물 - 바다나리아문
바다나리아문은 전통적으로 줄기를 가진 극피동물 분류군으로 여겨졌으나, 현재는 바다나리강과 동일시되며, 파라크리노이데아와 스타일로포라가 제외될 경우 현존하는 바다나리류를 모두 포함하는 분류군이 된다. - 극피동물 - 별불가사리
별불가사리(Patiria pectinifera)는 동북아시아 해역에 분포하며 얕은 조간대에서 서식하고, 조류 등을 섭취하며, 난모세포 연구에 사용되는 모델 생물이다. - 극피동물 - 저서생물
저서생물은 물 밑바닥이나 표면에 서식하는 생물들을 통칭하며, 수생 생태계에서 중요한 역할을 하지만 여러 요인으로 위협받고 있다. - 화석 - 삼엽충
삼엽충은 고생대 캄브리아기부터 페름기까지 번성한 절지동물로, 약 5억 2천만 년 전 등장하여 2억 7천만 년 동안 생존하며 바다에 서식했다. - 화석 - 규화목
나무가 땅에 묻혀 광물질에 의해 화석화된 규화목은 일본과 미국 등지에서 발견되며, 특히 일본에서는 다양한 종류가 천연기념물로 지정되어 보호받고 있다.
| 바다나리 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Crinoidea |
| 명명자 | 밀러, 1821 |
| 화석 범위 | 오르도비스기에서 현생 |
| 주요 그룹 | |
| 주요 그룹 | †카메라타 펜타크리노이데아 관절강 (700 현생 종) †디스파리다 †포로크리노이데아 †플렉시빌리아 |
| 한국어 이름 | |
| 한국어 이름 | ~우미유리, ~우미시다 |
| 영어 이름 | sea lilies, feather-stars |
| 일본어 이름 | |
| 일본어 이름 | ウミユリ綱 |
| 일본어 이름 (로마자) | Umiryuri-kō |
| 하위 분류 | |
| 하위 분류 | 관절아강 Articulata (현생) 유리아강 Inadunata (절멸) 가곡아강 Flexibilia (절멸) 원정아강 Camerata (절멸) |
| 강 | |
| 강 | 바다나리강 |
| 강 명명자 | Miller, 1821 |
| 하위 링크 | 목 |
| 기타 | |
| 설명 | 바다나리는 극피동물문의 한 강이다. "바다 백합"이라고도 불린다. 바다나리는 700종 이상의 살아있는 종을 포함한다. 9,000m 깊이에서도 발견된다. 이들의 뼈는 사암에 보존되어 있다. 이들의 화석은 석회암에서 발견된다. |
| 출처 | Encyclopædia Britannica Online Etymology Dictionary |
| 추가 정보 | 2023년 8월 12일, 과학자들은 20개의 팔을 가진 새로운 바다 생물을 발견하고 딸기를 따서 이름을 지었다. Business Insider 크와잘레인 환초의 얕은 물 바다나리 동물군에 관한 연구 Pacific Science 일본 이즈-오가사와라 해구에서 발견된 유경 바다나리 밀집 서식지 Zoological Science |
2. 어원
"바다나리강"을 뜻하는 "Crinoidea"라는 이름은 고대 그리스어 κρίνον|크리논grc(''krínon'', 백합)에서 유래했으며, "~와 같은"이라는 의미를 지닌 접미사 '-oid'가 붙은 것이다.[11][12] 영어 일반명인 'crinoid' 역시 같은 어원을 가진다. 성체 형태로 줄기를 이용해 해저에 부착된 바다나리는 흔히 '바다 백합'이라고 불린다. 반면, 부착되지 않고 자유롭게 움직이는 형태는 '깃별나리'라고 하며, 이들은 대부분 코마툴리다목(Comatulida)에 속한다.
바다나리는 암수딴몸으로, 각 개체는 수컷 또는 암컷이다. 대부분의 종에서 생식소는 몸의 일부인 깃털(pinnule)에 위치하지만, 일부 종에서는 팔에 있기도 하다. 모든 깃털이 생식에 관여하는 것은 아니며, 몸 중앙의 갓(crown)에 가장 가까운 깃털들만 생식 기능을 가진다. 생식세포(정자와 알)는 생식체강(genital coelom)에 둘러싸인 생식관에서 만들어진다. 성숙하면 깃털이 파열되어 정자와 알을 주변 바닷물로 방출하며, 체외수정이 이루어진다.[15]
바다나리강(Crinoidea)은 1821년 밀러(Miller)에 의해 처음 정의된 이후 극피동물의 독립적인 분지로 인정받아 왔다.[33] 현생종이 속하는 분류군은 관절바다나리아강(Articulata)뿐이며, 대표적인 종으로는 가는발깃갯고사리, 일본깃갯고사리 등이 있다.[47] 현존하는 관절바다나리아강에는 약 540종이 포함되며[33], 현존하는 목으로는 깃털별목(Comatulida), 굽은바다나리목(Cyrtocrinida), 깊은바다나리목(Hyocrinida), 줄기바다나리목(Isocrinida)이 있다.[33]
3. 형태

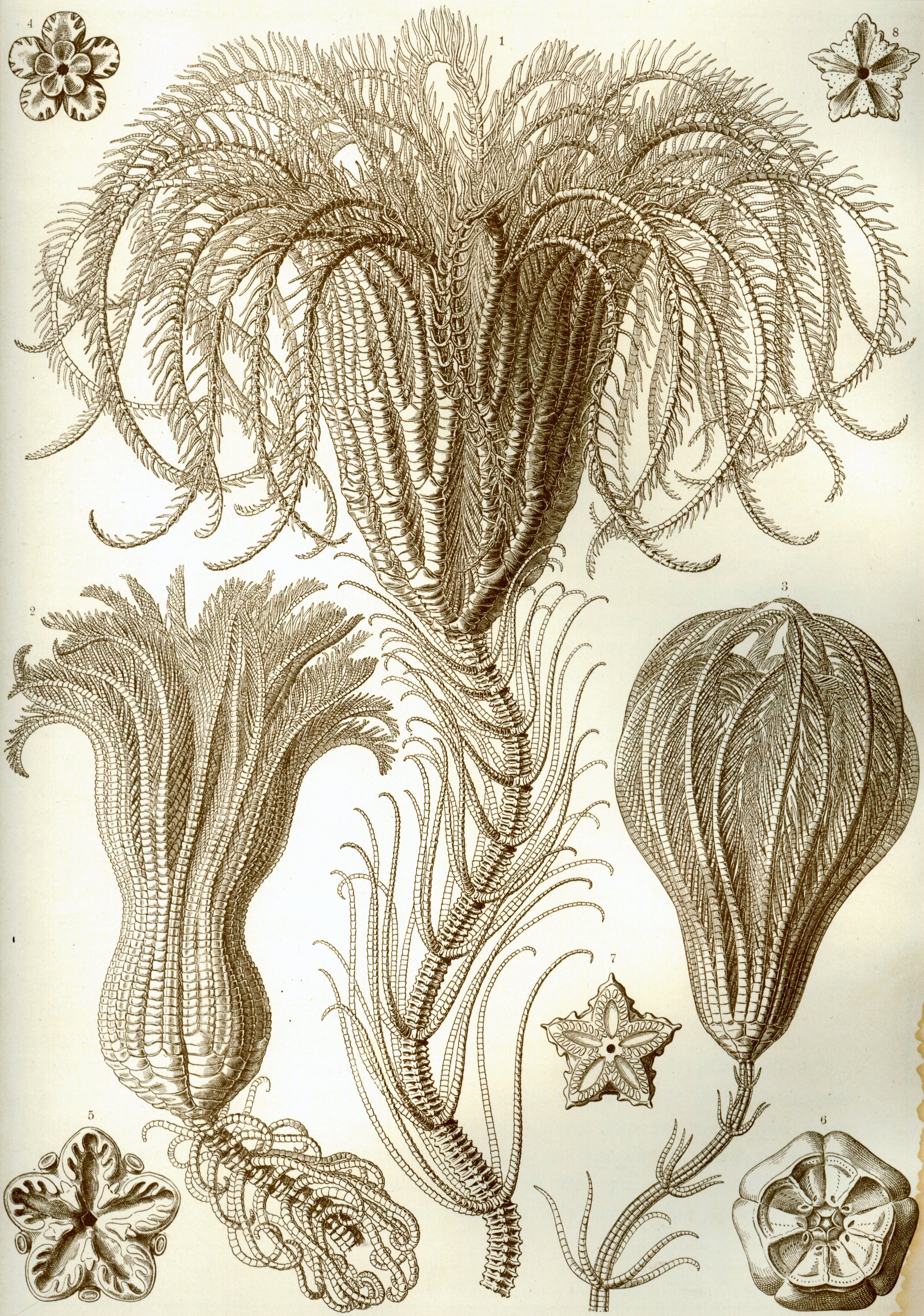
바다나리의 기본적인 몸 형태는 줄기(성체 깃털별에는 없음)와, 컵 모양의 중앙 몸체인 술잔(theca)과 보통 가지가 갈라지고 깃털 모양인 다섯 개의 팔 또는 광선으로 이루어진 갓(crown)으로 구성된다. 입과 항문은 모두 술잔의 위쪽에 위치하며, 이로 인해 등쪽(윗면)이 구강면이 된다. 이는 성게, 불가사리, 뱀거미불가사리와 같은 다른 극피동물 그룹과는 달리 입이 아랫면에 있는 것과 대조적이다.[13] 바다나리의 몸 대부분은 수많은 석회질 골편(ossicle)으로 이루어져 있으며, 연조직은 적은 비율을 차지한다. 이 골편들은 화석화가 잘 되며, 영국 클리더로 주변의 하부 석탄기 시대 석회암 층은 다양한 바다나리 화석으로 거의 전적으로 형성되어 있다.[14]
줄기는 다공성이 높은 골편 기둥으로 구성되며, 인대 조직으로 연결된다. 이는 편평한 부착기 또는 마디가 있는 뿌리 모양의 구조인 촉수(cirri)의 소용돌이를 통해 기질에 부착된다. 더 많은 촉수가 줄기 위쪽에 나타날 수 있다. 단단한 표면에 부착하는 바다나리의 경우, 촉수는 튼튼하고 굽어 있으며 새 발과 비슷하지만, 바다나리가 부드러운 퇴적물에 서식하는 경우 촉수는 가늘고 막대기 모양일 수 있다. 어린 깃털별은 줄기가 있지만, 나중에 없어지며, 많은 종들이 갓의 기저에 몇 개의 촉수를 유지한다. 대부분의 살아있는 바다나리는 자유 유영성이며, 퇴화된 줄기만 가지고 있다. 아직도 줄기를 유지하는 심해 종의 경우, 길이가 최대 1m에 이를 수 있지만(보통 훨씬 작다), 화석 종은 20m 길이의 줄기를 가진 것으로 알려져 있다.[15]
술잔(theca)은 오방사대칭(다섯 부분 대칭)이며, 다른 극피동물의 몸이나 원반과 상동적이다. 술잔의 기저는 컵 모양의 골편(뼈 판) 세트인 꽃받침(calyx)으로 형성되고, 윗면은 약하게 석회화된 막질 원반인 덮개(tegmen)으로 형성된다. 덮개는 관족이 돌출되는 깊은 홈을 포함하는 다섯 개의 "보행구역(ambulacral area)"과 그 사이에 있는 다섯 개의 "보행간 구역(interambulacral area)"으로 나뉜다. 입은 덮개의 중앙 또는 가장자리 근처에 있으며, 보행구역 홈은 팔의 기저에서 입으로 이어진다. 항문 또한 덮개에 위치하며, 종종 보행간 구역 안의 작은 원뿔 모양의 돌출부 위에 위치한다. 술잔은 비교적 작고 바다나리의 소화 기관을 포함한다.[15]
팔은 줄기와 유사한 일련의 관절 골편으로 지지된다. 원시적으로 바다나리는 다섯 개의 팔만 가지고 있었지만, 대부분의 현대 형태에서는 이것이 두 번째 골편(brachial II)에서 두 개로 나뉘어 총 열 개의 팔을 갖게 된다. 대부분의 살아있는 종, 특히 자유 유영성 깃털별에서 팔은 여러 번 더 갈라져 최대 200개의 가지를 생성한다. 관절이 있기 때문에 팔은 말릴 수 있다. 팔의 양쪽에는 번갈아 가며 "깃가지(pinnule)"라고 알려진 더 작은 관절 부속물이 늘어서 있어 깃털 모양을 나타낸다. 팔과 깃가지 모두 보행구역 홈의 가장자리를 따라 관족을 가지고 있다. 관족은 크기가 다른 세 개의 그룹으로 나타나며, 흡반이 없고 먹이 입자를 잡고 조작하는 데 사용된다. 홈에는 먹이를 팔을 따라 입으로 이동시키는 데 도움이 되는 섬모가 장착되어 있다.[15] 팔이 갈라지는 방식은 거미불가사리류와 차이가 있는데, 바다나리는 팔의 기부에서 잘 분지하고 끝은 별로 분지하지 않아 꽃잎처럼 보이는 반면, 거미불가사리류는 끝으로 갈수록 계속 가지를 쳐서 부채꼴로 펼쳐지는 경향이 있다.
4. 생태

바다나리는 수동적인 부유물 여과섭식자로, 깃털 모양의 팔을 이용하여 지나가는 바닷물에서 플랑크톤과 작은 부유물 입자를 걸러 먹는다. 팔은 부채꼴 모양으로 들어 올려져 해류와 수직으로 유지된다. 이동성 바다나리는 먹이 섭취 기회를 극대화하기 위해 바위, 산호초 또는 다른 돌출부에 붙어 산다. 먹이 입자는 가장 긴 일차 관족에 의해 포획되는데, 이 관족은 완전히 뻗어 깃 모양의 부속지에서 똑바로 서서 먹이를 잡는 그물 역할을 한다. 반면, 이차 및 삼차 관족은 주변의 물체를 다루는 데 관여한다.[15]
관족은 끈적끈적한 점액으로 덮여 있어 접촉하는 모든 입자를 붙잡는다. 먹이 입자가 잡히면 관족은 이를 순환구로 옮기고, 그곳의 섬모가 점액과 먹이 입자를 입 쪽으로 이동시킨다. 순환구 측면의 주름은 점액 흐름을 일정하게 유지하는 데 도움을 준다. 먹이를 잡는 표면의 총 길이는 매우 클 수 있다. 예를 들어, 24cm 길이의 팔을 가진 일본 해백합의 56개 팔은 깃 모양의 부속지를 포함하여 총 길이가 80m에 이른다. 일반적으로 플랑크톤이 비교적 적은 환경에 사는 바다나리는 먹이가 풍부한 환경에 사는 종보다 더 길고 가지가 많이 갈라진 팔을 가진다.[15]
입은 짧은 식도로 이어지며, 진정한 위는 없다. 식도는 장에 직접 연결되는데, 장은 꽃받침 내부를 한 바퀴 도는 단일 고리 모양이다. 장에는 종종 수많은 맹낭(막힌 주머니)이 포함되어 있으며, 그중 일부는 길거나 가지가 갈라질 수 있다. 장의 끝은 짧은 근육질의 직장으로 열리고, 직장은 항문으로 이어져 테그멘(입반) 가장자리의 작은 원뿔형 돌출부 밖으로 열린다. 배설물은 크고 점액으로 뭉쳐진 덩어리 형태로 만들어져 테그멘 위로 떨어진 후 바닥으로 가라앉는다.[15]
해삼류인 칼로키다리스 미칸스(Calocidaris micans)의 표본이 바다나리류인 엔독소크리누스 파래(Endoxocrinus parrae) 근처에서 발견되었는데, 해삼의 소화관 내에서 많은 양의 바다나리 줄기 부분이 발견되었다. 이 줄기 부분은 부드러운 조직과 함께 관절로 연결된 골편으로 구성되어 있었지만, 주변 퇴적물에서는 부드러운 조직이 없는 분리된 골편만 발견되었다. 이는 이 해삼류가 바다나리를 포식하며, 바다나리가 도망치는 과정에서 줄기의 일부를 스스로 잘라낸다는 것을 시사한다.[16]
다양한 바다나리 화석은 선사시대 포식자의 존재를 암시한다. 예를 들어, 쥐라기 라거슈타테인 졸른호펜에서 발견된 어류와 두족류의 코프롤라이트(화석화된 배설물)에서는 원양성 바다나리인 사코코마(Saccocoma)와 같은 다양한 바다나리의 골편이 발견되었다.[17] 또한, 데본기 후기 폴란드에서는 코코스테아이(Coccosteidae)과에 속하는 플라코더미(판피어류)의 이빨 자국과 일치하는 흔적이 있는 손상된 바다나리 줄기가 발견되었다.[18] 일부 데본기부터 석탄기까지의 바다나리 꽃받침(calyx)에서는 달팽이류인 플라티케라스(Platyceras)의 껍질이 밀접하게 붙어 있는 것이 발견되었다.[19] 일부는 달팽이가 항문 위에 위치하여 배설물 섭식을 하는 공생 관계를 시사하는 반면, 다른 것들은 달팽이가 직접 구멍 위에 위치하여 바다나리에게 해를 끼치는 관계였을 가능성을 시사한다.[20]
다른 극피동물처럼 바다나리도 수관계를 가지고 있으며, 이는 관족 내의 수압을 유지하는 역할을 한다. 하지만 다른 극피동물과 달리 마드레포라이트(천공판)를 통해 외부 해수와 연결되어 있지 않고, 수많은 작은 구멍을 통해 체강(몸통 속 공간)과만 연결된다. 주요 체액 저장소는 근육질 벽으로 둘러싸인 환상관이며, 이는 석회질 물질로 덮인 석관을 통해 체강과 연결된다. 체강은 중격에 의해 여러 개의 서로 연결된 공간으로 나뉜다. 체강은 원반 내부의 내장을 둘러싸고 있으며, 줄기와 팔 안에도 가지를 뻗고, 더 작은 가지는 깃털 모양의 돌기(pinnule)까지 이어진다. 관족을 뻗는 것은 환상관의 수축에 의해 이루어진다. 세 개의 가는 체강 가지가 각 팔에 들어가는데, 두 개는 입쪽에, 하나는 입 반대쪽에 위치하며, 깃털 모양의 돌기까지 이어진다. 섬모의 작용으로 이 관들 안에서 느린 유체 흐름(초당 1mm)이 발생하는데, 입쪽 가지에서는 바깥쪽으로, 입 반대쪽 가지에서는 안쪽으로 흐르며, 이것이 영양분과 노폐물의 주요 이동 수단이다. 심장과 별도의 순환계는 없지만, 원반 기저부에는 축기관이라고 알려진 큰 혈관이 있으며, 기능을 알 수 없는 가느다란 막힌 관을 포함하고 있으며, 줄기까지 뻗어 있다.[15]
이러한 다양한 체액으로 채워진 공간은 신체 내 영양분을 운반하는 것 외에도 호흡계와 배설계의 기능을 모두 수행한다. 산소는 주로 관족을 통해 흡수되는데, 관족은 신체에서 가장 얇은 벽을 가진 부분이며, 팔의 넓은 표면적에서도 추가적인 가스 교환이 일어난다. 노폐물은 식세포성 체강세포에 의해 수집되므로, 특수한 배설 기관은 없다.[15]
바다나리의 신경계는 세 부분으로 나뉘며, 그 사이에는 수많은 연결이 존재한다. 구강부 또는 가장 위쪽 부분은 다른 극피동물의 신경계와 상동하는 유일한 부분으로, 입을 둘러싼 중앙 신경환과 팔로 분지되는 방사 신경으로 구성되며 감각 기능을 담당한다. 그 아래에는 중간 신경환이 있으며, 팔과 깃털에 신경을 공급하는 방사 신경을 내보낸다. 이 신경들은 운동성이며, 관족의 근육을 제어한다. 신경계의 세 번째 부분은 밑면에 있으며, 팔, 깃털 및 씨러스(cirri)의 굴곡 및 움직임을 담당한다. 이것은 꽃받침(calyx)의 기저 근처에 있는 신경 조직 덩어리에 중심을 두고 있으며, 각 팔에 하나의 신경과 줄기에 여러 개의 신경을 제공한다.[15]
바다나리는 불가사리나 거미불가사리처럼 무성생식을 할 수는 없지만, 잃어버린 신체 부위를 재생할 수 있는 능력이 뛰어나다. 포식자에게 뜯겨 나가거나 좋지 않은 환경 조건으로 손상된 팔은 다시 자랄 수 있으며, 심지어 내장 기관도 몇 주 안에 재생될 수 있다.[15] 줄기의 가장 위쪽 마디와 기저판은 전체 갓(crown)을 재생할 수 있다.[22] 이때 줄기, 특히 위쪽 5cm에서 나온 영양분과 다른 성분들이 갓 재생에 사용된다.[22] 바다나리는 고생대부터 신체 부위를 재생할 수 있었으며,[22] 이러한 재생 능력은 포식성 어류의 공격에서 살아남는 데 중요했을 수 있다.[15]
바다나리강은 암수딴몸이며, 체외수정에 의한 유성생식을 한다. 수컷 개체가 정자를, 암컷 개체가 알을 방출하면, 이것들이 해류를 타고 운반되어 수정 후에 유생이 된다. 발생 과정에서, 바다나리목(Comatulida)에서는 3가지 형태의 유생이 알려져 있다. 유영성이고 먹이를 먹지 않는 도리올라리아 유생이 바닥에 부착하여, 다음으로 줄기가 있는 시스티지안 유생, 펜타크리노이드 유생으로 형태를 바꾼다. 펜타크리노이드 유생은 성숙하면 줄기를 떼어내고, 자유롭게 헤엄치는 성체가 된다. 종에 따라 차이는 있지만, 수정 후 약 10~16개월 만에 생식 가능한 성체가 된다. 일부 종은 유생을 일시적으로 보호하기도 한다. 다른 4목(유경 바다나리류)의 발생 과정은 잘 알려져 있지 않다.
해백합류(바다나리목)는 얕은 바다에도 분포하며, 팔을 이용한 유영이나 팔 아랫면의 씨러스(cirrus)를 이용한 포복 운동을 한다. 반면, 유경 바다나리류는 주로 수심 100m 이상 깊은 곳에서만 서식한다. 일본에서는 트리노아시가 가장 얕은 수역에서 발견되는 유경 바다나리이다. 유경 바다나리는 보통 식물의 뿌리처럼 생긴 구조나 원반 모양의 줄기 끝으로 바닥에 부착하지만, 줄기를 끌면서 팔을 이용하여 이동할 수도 있다. 두 종류 모두 팔과 깃털 모양의 가지(pinnule)를 펼쳐 물속의 유기물 입자를 잡아먹는다.
2005년에는 서인도 제도의 그랜드 바하마 섬 근해에서 해저를 고속으로 이동하는 유경 바다나리가 보고되었다. 유경 바다나리류가 이동한다는 사실은 오래전부터 알려져 있었지만, 그 속도는 기껏해야 시간당 0.6m 정도였다. 하지만 이 보고된 종은 현지 조사에서 시간당 140m의 속도를 기록했다고 한다.[41] 참고로, 화석 종 중에는 뿌리 부분에 부력을 얻기 위한 구조를 가지고 물에 떠다녔던 것으로 보이는 종류도 알려져 있다.
5. 생식
일부 종은 수정란을 보호하기도 한다. 예를 들어 ''안테돈속 (Antedon)''과 같은 속은 표피선의 분비물로 수정란을 팔에 붙여두며, 특히 남극의 차가운 바다에 사는 종들은 알을 팔이나 깃털의 특별한 주머니에서 부화시킨다.[15]
수정란이 부화하면 자유롭게 헤엄치는 비텔라리아 유생(vitellaria larvae)이 된다. 이 유생은 좌우대칭이고 원통형이며, 몸 주위에는 섬모 띠가 있고 위쪽에는 감각털 다발이 있다. 다른 현생 극피동물류에는 먹이를 먹는 유생과 먹이를 먹지 않는 유생이 모두 존재하지만, 현생 바다나리는 모두 먹이를 먹지 않는 유생(lecithotrophic larvae)을 가진다. 이는 페름기-트라이아스기 멸종 사건 이후 병목 현상을 겪으면서 살아남은 조상 계통이 먹이를 먹는 유생 단계를 잃어버렸기 때문으로 추정된다.[21]
유생은 며칠 동안 떠다니다가 바닥에 내려앉아 아랫면의 접착선을 이용해 고착한다. 이후 긴 변태 과정을 거쳐 줄기가 있는 어린 개체(펜타크리노이드 유생)로 변하며 방사대칭 형태가 된다.[15] 특히 줄기 없이 자유롭게 생활하는 바다나리목(Comatulida)의 배 발생 과정에서는, 유영하며 먹이를 먹지 않는 도리올라리아 유생이 기질에 부착한 뒤 줄기가 있는 시스티지안 유생, 그리고 펜타크리노이드 유생으로 변태하는 3가지 형태의 유생 단계가 알려져 있다. 이 바다나리목 개체들은 성체가 되면서 줄기에서 떨어져 나온다.[15] 종에 따라 차이가 있지만, 수정 후 약 10~16개월이 지나면 생식이 가능한 성체가 된다. 줄기가 있는 다른 바다나리류(유경 바다나리류)의 발생 과정은 상대적으로 덜 알려져 있다.
6. 진화



바다나리강(Crinoidea)은 성게, 해삼과 같은 극피동물의 한 무리이다. 이름처럼 식물과 비슷한 줄기 구조를 가지며, 그 기부로 기질에 부착하는 특징이 있다. 그러나 대부분의 현생종은 유생 시기에만 부착성 생활을 하고, 성체가 되면 자유롭게 헤엄치는 자유유영성으로 전환한다. 현생종은 약 650종이 알려져 있으며, 과거에는 훨씬 더 번성하여 다양한 화석 종을 남겼다. 고생대 중기~후기에 형성된 일부 석회암은 대부분 바다나리강 생물의 유해로 이루어져 있을 정도이다.
광물질 골격을 가진 극피동물은 초기 캄브리아기(약 5억 4천만 년 전) 화석 기록에 처음 등장하며, 이후 약 1억 년 동안 바다나리류와 블라스토이드류(역시 줄기가 있는 여과섭식자)가 해양 생태계의 주요 구성원이었다.[27] 당시 극피동물문에는 강 수준의 분류군이 20개나 존재했지만, 여러 차례의 대량 멸종 사건을 거치며 현재는 5개의 강만이 살아남았다. 바다나리류의 길고 다양한 지질학적 역사는 극피동물이 여과 섭식 생활 방식에 얼마나 성공적으로 적응했는지를 보여준다.[15]
가장 오래된 바다나리 무리는 약 4억 8천만 년 전 오르도비스기 초기에 출현한 것으로 여겨진다. (버제스 혈암의 수수께끼 같은 생물 ''Echmatocrinus''를 제외한다면). 바다나리강의 기원에 대해서는 두 가지 주요 가설이 존재한다. 전통적인 견해는 바다나리가 방사상동물(에오크리노이드와 그 후손인 완족류, 시스토이드) 내에서 진화했다고 보지만, 더 널리 받아들여지는 다른 가설은 바다나리가 에드리오아스테로이드로부터 초기 단계에 분리되었다고 주장한다.[26] 이 논쟁은 세 후보 조상 그룹 모두 방사 대칭, 석회질 판, 줄기 또는 직접 부착 방식 등 여러 특징을 공유하기 때문에 명확한 결론을 내리기 어렵다.[26] 바다나리강은 같은 바다나리아문(Crinozoa)에 속하는 원시 바다나리류(Eocystoid) 조상 생물군에서 진화한 것으로 생각된다. 다른 원시 바다나리류(Eocrinoid) 그룹도 있지만, 이들은 이름과 달리 바다나리강의 직접적인 조상이 아니며, 페름기까지 번성했던 우미츠보미류(Blastoid)의 조상으로 여겨진다.
바다나리류는 진화 역사 동안 두 차례의 급격한 적응 방산을 경험했다. 첫 번째는 오르도비스기(4억 8500만 년 전 ~ 4억 4400만 년 전)에 일어났고, 두 번째는 트라이아스기 초기(약 2억 3000만 년 전)에 발생했다.[28] 특히 트라이아스기 방산 시기에는 유연한 팔을 가진 형태들이 널리 퍼졌으며, 주로 포식 압력에 대한 반응으로 고착성 생활 방식보다 이동성을 가진 종들이 훨씬 더 흔해졌다.[29] 이 두 번째 방산은 중생대 해양 혁명보다 약간 앞서 일어났는데, 이는 주로 해삼과 같은 다른 극피동물의 저서성 포식 활동 증가에 의해 촉진되었을 가능성이 있다.[30]
그러나 페름기 말에 발생한 대량 멸종은 바다나리류에게 큰 타격을 주었다. 이 사건으로 모든 블라스토이드류와 대부분의 바다나리류가 멸종했다.[28] 페름기 말 멸종 이후, 바다나리류는 고생대에 누렸던 형태적 다양성과 생태계에서의 우점적 지위를 완전히 회복하지 못했으며, 고생대와는 다른 생존 전략을 채택하게 되었다.[28]
일부 바다나리 화석, 예를 들어 ''펜타크리니테스(Pentacrinites)'' 속은 바다에 떠다니는 나무(부표목)에 붙어서 살았던 것으로 보이며, 완벽한 군집 형태의 화석이 종종 발견된다. 이는 부표목이 물에 잠겨 가라앉으면서 붙어 있던 바다나리 군집 전체가 함께 화석화된 것으로 해석된다. ''펜타크리니테스''의 줄기는 때때로 수 미터에 달할 정도로 길었다. 반면, 이들의 현생 친척들은 주로 심해의 부드러운 해류 속에서 살며, 줄기 끝으로 바위에 붙어 생활한다.
2012년, 지질학자들은 약 3억 4천만 년 전(미시시피기)의 여러 바다나리 화석에서 복잡한 유기 분자를 분리하는 데 성공했다고 보고했다. 이 분자들은 "방향족 또는 다환방향족 퀴논과 유사한" 것으로 확인되었으며, 특정 개체 화석과 명확하게 연관된 가장 오래된 유기 분자로 기록되었다. 이는 골편의 미세한 구멍들이 석회화 과정에서 방해석으로 채워지면서 유기 분자가 보존될 수 있었기 때문으로 추정된다.[31]
바다나리 화석, 특히 분리된 줄기 마디(골편)는 매우 풍부하게 발견되며, 때로는 퇴적암의 주요 구성 요소가 되기도 한다. 이렇게 바다나리 유해로 주로 이루어진 암석을 엔크리나이트(encrinite)라고 부른다.
대부분의 현생 바다나리류는 성체가 되면 줄기가 없는 자유 유영성 형태를 띤다. 이러한 자유 유영성 바다나리는 현생종뿐만 아니라 화석으로도 발견되는데, 대표적인 예로는 ''Marsupitsa'', ''Saccocoma'', ''Uintacrinus'', ''Pterocoma'' 등이 있다. 이들 화석은 주로 독일 바바리아주의 졸른호펜(Solnhofen) 지역에 있는 쥐라기 석회암 지층이나, 미국 캔자스주 니오브라라(Niobrara)의 백악기 지층에서 출토된다. 반면, 줄기가 있는 고착성 바다나리 화석은 전 세계 여러 지역의 다양한 지질 시대 지층에서 발견된다.시대 주요 발견 지역 실루리아기 영국, 미국 동부 데본기 미국 (켄터키주, 미시간주, 일리노이주, 뉴욕주), 독일 분덴바흐(Bundenbach) 근교 (훈스뤽 슬레이트(Hunsrück Slates)) 석탄기 영국, 벨기에, 러시아 미시시피기 (석탄기 전기) 미국 (아이오와주, 인디애나주, 앨라배마주) 펜실베이니아기 (석탄기 후기) 미국 중부 페름기 티모르섬 트라이아스기 독일, 미국 미주리주
바다나리강의 진화 과정은 극피동물 내에서 여과 섭식 생활 방식이 어떻게 발전해왔는지를 보여주는 중요한 사례이다. 고생대에는 바다나리류 외에도 앞서 언급된 블라스토이드류 역시 줄기를 이용한 여과 섭식자들이었으며, 고생대 지층에서 매우 풍부하게 발견된다. 블라스토이드 화석은 종류와 개체 수 면에서 바다나리류를 능가할 정도로 많이 출토되어, 당시 해저 생태계에서 두 그룹 간의 경쟁이 있었을 것으로 추정된다. 그러나 블라스토이드류는 P-T 경계에서 완전히 멸종하여 현생 후손을 남기지 못했다. 따라서 바다나리강 생물들은 과거 여과 섭식 극피동물의 진화와 생태를 이해하는 데 중요한 단서를 제공하는 살아있는 증거라고 할 수 있다.
7. 분류
라이트 등(Wright et al., 2017)은 극피동물의 계통 분류, 지질학적 역사, 분류법에 대해 논의하며 새로운 계통 기반 및 등급 기반 분류를 제안하였다.[32][33][34][35][36] 이들의 등급 기반 분류(목 수준까지)는 일부 그룹이 ''위치 미상''(incertae sedis)으로 남아 있지만, 다음과 같은 계층 구조를 가진다.
목 †유스테노크리니드목(Eustenocrinida)
목 †마에닐리크리니드목(Maennilicrinida)
목 †사각바다나리목(Tetragonocrinida)
목 †칼세오크리니드목(Calceocrinida)
''위치 미상'' 그룹: '호모크리니다목(Homocrinida)', '미엘로닥틸라목(Myelodactylida)', '피소크리노이데아상과(Pisocrinoidea)'
상목 †다공바다나리아상목(Porocrinoidea)
* 목 †히보크리니드목(Hybocrinida)
* 목 †다공바다나리목(Porocrinida)
상목 †가요성바다나리아상목(Flexibilia)
* 목 †사게노크리니드목(Sagenocrinida)
* 목 †탁소크리니드목(Taxocrinida)
대목 진클라디다(Eucladida)
* †암펠로크리니드목(Ampelocrinida) (''위치 미상'')
* 상목 †술잔형바다나리아상목(Cyathoformes)
** ''위치 미상'' 그룹: '시아토크리니다목(Cyathocrinida)', '덴드로크리니다목(Dendrocrinida)', '포테리오크리니다목(Poteriocrinida)'
* 상목 관절바다나리아강(Articulata)
** 목 †엔크리니드목(Encrinida)
** 목 †홀로크리니드목(Holocrinida)
** 목 †밀러크리니드목(Millericrinida)
** 목 †로베아크리니드목(Roveacrinida)
** 목 †윈타크리니드목(Uintacrinida)
** 목 깃털별목(Comatulida)
** 목 굽은바다나리목(Cyrtocrinida)
** 목 깊은바다나리목(Hyocrinida)
** 목 줄기바다나리목(Isocrinida)











8. 문화
린디스파르네(Lindisfarne)에서 채석한 석회암에서 추출하거나 해안가에서 발견된 바다나리 화석 줄기 마디는 목걸이나 묵주로 만들어졌다. 중세 시대에는 이를 세인트 Cuthbert의 구슬이라고 불렀다.[37] 미국 중서부에서도 바다나리 줄기의 화석화된 마디를 때때로 인디언 구슬이라고 부른다.[38] 바다나리의 한 종류인 ''Eperisocrinus missouriensis''는 미주리주의 주 화석으로 지정되어 있다.[39] 또한, 영화 시리즈 ''에일리언''에 등장하는 외계 생명체는 바다나리에서 영감을 받아 디자인되었다.[40]
참조
[1]
웹사이트
Crinoidea – Digital atlas of Ancient life
https://www.digitala[...]
[2]
WoRMS
Crinoidea
2013-01-30
[3]
서적
New Zealand inventory of biodiversity: 1. Kingdom Animalia: Radiata, Lophotrochozoa, Deuterostomia
Canterbury University Press
[4]
뉴스
Scientists found a new sea creature with 20 'arms' and named it after a strawberry
https://www.business[...]
2023-08-12
[5]
학술지
The Shallow-Water Crinoid Fauna of Kwajalein Atoll, Marshall Islands: Ecological Observations, Interatoll Comparisons, and Zoogeographic Affinities
https://scholarspace[...]
[6]
학술지
Discovery of Dense Aggregations of Stalked Crinoids in Izu-Ogasawara Trench, Japan
[7]
서적
Reproduction and Development in Echinodermata and Prochordata
https://books.google[...]
[8]
학술지
Diagenesis of a Crinoidal Sediment
1962
[9]
학술지
Aspects of the depositional environment and palaeoecology of crinoidal limestones
1968-09
[10]
학술지
Storm-dominated deposition of the Lower Jurassic crinoidal limestones in the Krížna unit, Western Tatra Mountains, Poland
2005-04
[11]
서적
Webster's New Universal Unabridged Dictionary
[12]
백과사전
crinoid
http://www.etymonlin[...]
[13]
서적
Australian Echinoderms: Biology, Ecology and Evolution
https://books.google[...]
Csiro Publishing
[14]
서적
Fossil Crinoids
https://books.google[...]
Cambridge University Press
[15]
서적
Invertebrate Zoology: A Functional Evolutionary Approach
https://archive.org/[...]
Thomson-Brooks/Cole
[16]
학술지
Urchins in the meadow: Paleobiological and evolutionary implications of cidaroid predation on crinoids
[17]
서적
Fossil Crinoids
Cambridge University Press
[18]
학술지
Inferred placoderm bite marks on Devonian crinoids from Poland
[19]
학술지
Predators and predation in Paleozoic marine environments
https://web.archive.[...]
2014-04-06
[20]
학술지
Infestation of Middle Devonian (Givetian) camerate crinoids by platyceratid gastropods and its implications for the nature of their biotic interaction
https://deepblue.lib[...]
[21]
학술지
The active evolutionary lives of echinoderm larvae
[22]
학술지
Regeneration in sea lilies
https://doi.org/10.1[...]
1992-06
[23]
웹사이트
About Crinoids
https://www.fossiler[...]
FossilEra
2019-03-15
[24]
학술지
The locomotion of the comatulid ''Florometra serratissima'' (Echinodermata: Crinoidea) and its adaptive significance
[25]
학회
Crawling In Stalked Crinoids: In Situ Observations, Functional Morphology, and Implications for Paleozoic Taxa
https://web.archive.[...]
2005-10-06
[26]
학술지
Pelmatozoan arms from the mid-Cambrian of Australia: Bridging the gap between brachioles and brachials? Comment: There is no bridge
[27]
웹사이트
Echinodermata: Fossil Record
http://www.ucmp.berk[...]
Museum of Paleontology: University of California at Berkeley
2019-03-30
[28]
학술지
Morphological diversity in the evolutionary radiation of Paleozoic and post-Paleozoic crinoids
[29]
학술지
Crinoid Ecological Morphology
[30]
학술지
Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution
[31]
웹사이트
Ancient fossilized sea creatures yield oldest biomolecules isolated directly from a fossil
http://phys.org/news[...]
2013-02-19
[32]
학술지
Phylogenetic taxonomy and classification of the Crinoidea (Echinodermata)
[33]
학술지
Early phylogeny of crinoids within the pelmatozoan clade
[34]
학술지
Bayesian estimation of fossil phylogenies and the evolution of early to middle Paleozoic crinoids (Echinodermata)
[35]
학술지
Phylogeny and morphologic evolution of the Ordovician Camerata (Class Crinoidea, Phylum Echinodermata)
[36]
저널
Fixed, free, and fixed: the fickle phylogeny of extant Crinoidea (Echinodermata) and their Permian-Triassic origin
[37]
저널
The Legend of St Cuthbert's Beads: A Palaeontological and Geological Perspective
[38]
웹사이트
Identifying Unknown Fossils (by their shape)
http://www.uky.edu/K[...]
Kentucky Geological Survey / University of Kentucky
2009-06-21
[39]
웹사이트
Missouri's State Fossil
http://www.sos.mo.go[...]
Office of the Secretary of State, Missouri
2019-03-31
[40]
뉴스
The Fossils That Inspired 'Alien'
https://www.forbes.c[...]
2019-04-26
[41]
학회발표
Crawling In Stalked Crinoids: In Situ Observations, Functional Morphology, and Implications for Paleozoic Taxa
[42]
웹인용
Sea lily
http://www.britannic[...]
Encyclopædia Britannica
2011-03-14
[43]
서적
Webster's New Universal Unabridged Dictionary
[44]
웹인용
crinoid
http://www.etymonlin[...]
Online Etymology Dictionary
[45]
저널
The Shallow-Water Crinoid Fauna of Kwajalein Atoll, Marshall Islands: Ecological Observations, Interatoll Comparisons, and Zoogeographic Affinities
https://scholarspace[...]
[46]
저널
Discovery of Dense Aggregations of Stalked Crinoids in Izu-Ogasawara Trench, Japan
http://www.bioone.or[...]
[47]
간행물
한국동물분류학회
집현사
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com