2025. 9. 21. 오후 9:04:00
[암석 속 과학 이야기]수억년 데고 눌리고…지구의 장대한 역사, 변성암 한 조각으로 ‘압축’
출처: 경향 신문 ( 한국 / 한국어 )
트라이아스기는 지질 시대의 한 시기로, 1834년 독일 지질학자 프리드리히 폰 알베르티가 명명했으며, '삼위일체'를 의미하는 그리스어에서 유래되었다. 이 시기는 초기, 중기, 후기로 구분되며, 2억 5100만 년 전부터 1억 9900만 년 전까지 지속되었다. 트라이아스기에는 판게아 초대륙이 존재했고, 기후는 덥고 건조했으며, 다양한 해양 및 육상 생물이 번성했다. 초기에는 페름기-트라이아스기 대멸종 이후 생태계가 회복되었고, 파충류의 다양성이 증가했으며, 최초의 공룡과 포유류가 나타났다. 트라이아스기 말에는 대멸종이 발생하여 해양 생태계에 큰 피해를 입혔으며, 공룡이 번성하는 계기가 되었다. 한국에서는 영덕 심성암체와 트라이아스기 지층이 발견되며, 일본 미나미산리쿠정 사라카이 마을에는 트라이아스기 후기의 조개 화석 산지가 있다.
| 트라이아스기 | |
|---|---|
| 개요 | |
 | |
| 시기 | 중생대의 첫 번째 시기 |
| 기간 | 약 2억 5190만 년 전부터 2억 140만 년 전까지 |
| 이전 | 페름기 |
| 이후 | 쥐라기 |
| 지질 | |
| 주요 사건 | 판게아 형성 트라이아스기-쥐라기 멸종 사건 |
| 기후 | |
| 특징 | 전반적으로 덥고 건조한 기후 |
| 생물 | |
| 주요 생물 | 공룡의 등장 해양 파충류의 번성 암모나이트의 다양화 |
| 산소 농도 | 16% |
| 이산화탄소 농도 | 1750ppm |
| 평균 온도 | 17°C |
| 하위 구분 | |
| 시기 구분 | 전기 트라이아스기 중기 트라이아스기 후기 트라이아스기 |
| 세부 시대 | 인두안절 (Induan) 올레네크절 (Olenekian) 아니시안절 (Anisian) 라디니안절 (Ladinian) 카르니안절 (Carnian) 노리안절 (Norian) 레티안절 (Rhaetian) |
트라이아스기라는 명칭은 1834년 독일의 지질학자 프리드리히 폰 알베르티가 남부 독일에서 발견된 세 개의 뚜렷한 암석층(하부 분트사암(다채로운 사암), 중부 조개암(조개 함유 석회암), 상부 케우퍼(색깔 있는 점토))을 따라 명명한 것이다.[8]
트라이아스기는 초기, 중기, 후기로 나뉘며, 각 시기는 다시 세부적인 절(age)로 구분된다.
| 기/절 | 동물 화석 단계 | 시간 범위 |
|---|---|---|
| 후기 트라이아스기 | 라이티안절 | 2억 300만 년 전 ~ 1억 9900만 년 전 |
| 노리안절 | 2억 1600만 년 전 ~ 2억 300만 년 전 | |
| 카르니안절 | 2억 2800만 년 전 ~ 2억 1600만 년 전 | |
| 중기 트라이아스기 | 라디니안절 | 2억 3700만 년 전 ~ 2억 2800만 년 전 |
| 아니시안절 | 2억 4500만 년 전 ~ 2억 3700만 년 전 | |
| 초기 트라이아스기 | 올레네키안절 | 2억 4900만 년 전 ~ 2억 4500만 년 전 |
| 인두안절 | 2억 5100만 년 전 ~ 2억 4900만 년 전 |
| 기/절 | 동물 화석 단계 | 시간 범위 |
|---|---|---|
| 초기 트라이아스기 | 올레네키안절 | 2억 4900만 년 전 ~ 2억 4500만 년 전 |
| 인두안절 | 2억 5100만 년 전 ~ 2억 4900만 년 전 |
| 기/절 | 동물 화석 단계 | 시간 범위 |
|---|---|---|
| 중기 트라이아스기 | 라디니안절 | 2억 3700만 년 전 ~ 2억 2800만 년 전 |
| 아니시안절 | 2억 4500만 년 전 ~ 2억 3700만 년 전 |
트라이아스기라는 명칭은 남독일에서 발견된 이 시기의 지층에서 유래했다. 페름기(이첩기)의 상층에, 상위로부터 코이퍼 사암(Keuper, 英語版, 상첩통, 붉은색 사암), 무셸칼크(Muschelkalk, 英語版, 각회통, 흰색 석회암), 분터 사암(Bunter, 英語版 또는 Buntsandstein, 英語版, 반사통, 갈색 사암)과 같이 퇴적 조건이 다른 3개의 층이 겹쳐 있었던 것에서 기인한다. 독일의 지질학자 프리드리히 폰 알베르티(英語版)가 1834년에 명명했다.[73][74]
유럽에서 분터는 얕은 웅덩이에 퇴적된 색채가 풍부한 퇴적물을 함유하는 계열, 무셸칼크는 조개류 화석을 동반하는 석회암 계열이며, 코이퍼는 척박한 건조를 나타내는 암염과 석고 층을 동반하는 대륙의 퇴적물 계열로 알려져 왔다. 그러나 오늘날에는 네 번째 계열로 레에티크가 포함되어 트라이아스기 최신의 지층에 위치하고 있다.[75]
실제로는 독일 주변의 해성층은 트라이아스기 중기에 속하는 연대의 것으로 한정되어 있기 때문에, 트라이아스기 전체를 통틀어 편년하기 위해서는 알프스 산맥, 히말라야 산맥, 그리고 북아메리카 대륙 북부에서의 해양 동물의 화석이 풍부한 지층도 병용하며, 이것들을 표준으로 국제적인 시기 구분이 설정되어 있다.[76]
트라이아스기 동안, 지구의 거의 모든 육지는 하나의 거대한 초대륙인 판게아(Pangaea)로 집중되어 있었다.[9] 판게아의 서쪽 가장자리는 판탈라사라는 거대한 대양과 접해 있었고, 동쪽 가장자리는 테티스 해와 고테티스 해라는 두 개의 광대한 해양 분지로 침식되었다.[10][11] 판게아는 트라이아스기 동안, 특히 그 후기에 광범위한 단층과 열곡(rift) 분지에 의해 분열되기 시작했다.[13]
페름기-트라이아스기 대멸종 이후 생태계가 회복되면서 트라이아스기에는 다양한 생물군이 번성했다. 해양에서는 새로운 유형의 산호가 나타나 데본기 또는 현대의 거대한 산호초 시스템에 비해 규모가 작은 산호초 패치를 형성했다. 관벌레(Serpulidae)는 중기 트라이아스기에 나타났고, 마이크로콘키드가 풍부했다. 암모나이트는 페름기 대멸종에서 살아남은 단일 계통에서 분화하며 회복했고, 이매패류는 중기 트라이아스기 동안 급격히 분화하기 시작하여 바다에서 매우 풍부해졌다.[46] 벨렘나이트와 유사한 생물도 다수 나타났다.[75]
3. 고지리 및 기후
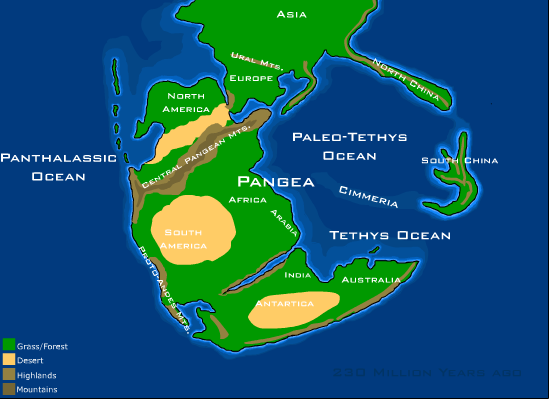
트라이아스기의 기후는 일반적으로 덥고 건조했으며, 전형적인 퇴적물은 적색층 사암과 증발암이다. 극지방은 온대 기후를 보였으며, 숲과 파충류를 포함한 척추동물에게 적합한 환경을 제공했다.[20] 판게아의 거대한 크기는 전 세계적인 해양의 완화 효과를 제한했으며, 강한 대조는 강렬한 적도 횡단 몬순을 유발했다.[20]
초기 트라이아스기는 매우 더웠으며, 중기에는 기온이 다소 하락했다가 후기에는 다시 온난화되는 경향을 보였다.[24][23]
4. 생물
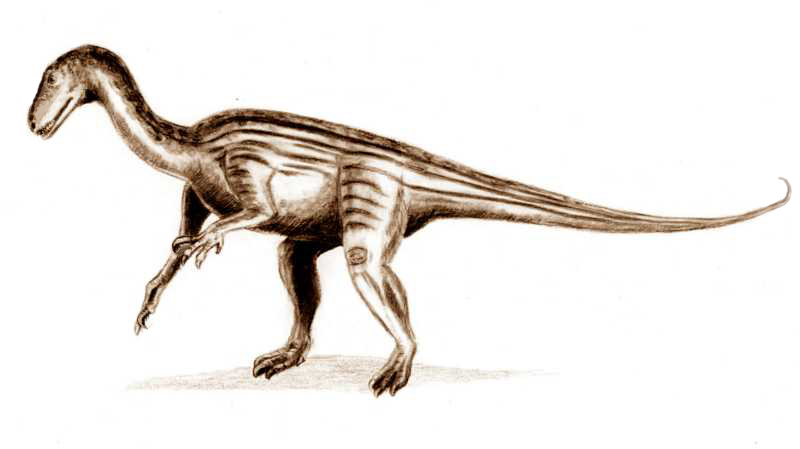
육상에서는 파충류의 다양성이 크게 증가했다. 특히 궁상목 파충류는 페름기-트라이아스기 대멸종에 대응하여 적응 방산으로 다양성이 폭발적으로 증가했다.[57] 플라케리아스, 칸네메에리아, 포스토수쿠스 등이 대표적이다. 또한 플라테오사우루스, 코엘로피시스, 헤레라사우루스 등 초기 공룡도 출현했다.

하늘에는 에우디모르포돈, 페테이노사우루스, 프레온닥틸루스와 같은 초기 익룡이 나타났다.
곤충 중에서는 침이 발달하지 않은 원시 벌목, 티타노프테라목(Titanoptera), 대벌레목, 노린재목 등이 있었다.
육상 식물에서는 양치식물과 겉씨식물이 번성했고[73], 소나무와 삼나무의 원조가 되는 침엽수가 나타났다.[77] 종자식물이면서 독립된 정자를 만드는 은행나무류나 소철류, 베네티티스류도 많았다. 습지에는 양치식물의 고란초과의 유연종인 고대 석송이 풍부하게 남아 고사리와 속새도 조밀하게 분포했다.[75]
4. 1. 육상 파충류
궁상목 파충류는 페름기-트라이아스기 대멸종에 대응하여 적응 방산으로 다양성이 폭발적으로 증가했다.[57] 초기 트라이아스기에는 여러 주요 궁상목 그룹이 나타났다. 목이 길고 도마뱀과 비슷한 초기 궁상목은 원룡류로 알려졌으며, 이는 진정한 분지군보다는 측계통군일 가능성이 높다. 타니스트로페우스과는 목의 크기를 극도로 늘린 원룡류의 한 종류로, 가장 큰 속인 ''Tanystropheus''는 몸통보다 목이 더 길었다. 원룡류의 일종인 샤로비프테리기다이과는 활공을 위해 길어진 뒷다리를 사용했다. 린코사우루스류와 알로코토사우루스류와 같은 다른 궁상목은 대부분 몸집이 튼튼한 초식동물로, 특수한 턱 구조를 가지고 있었다.
통통한 몸매의 초식동물인 린코사우루스류는 짧은 기간 동안 번성했으며, 약 2억 2천만 년 전에 멸종되었다. 이들은 트라이아스기 중기에 매우 흔했으며, 카르니안 시대의 많은 생태계에서 주요 대형 초식동물이었다. 이들은 앞턱의 부리와 여러 줄의 이가 있는 위턱을 따라 있는 판으로 식물을 잘라 먹었다. 알로코토사우루스류는 이구아나와 같은 파충류로, ''Trilophosaurus''(세 개의 왕관 모양 이를 가진 후기 트라이아스기 파충류), ''Teraterpeton''(부리 모양의 긴 주둥이를 가짐) 및 ''Shringasaurus''(3m의 몸길이에 달하는 뿔이 있는 초식동물)가 있다.
궁상목의 한 그룹인 궁상목은 날카로운 이빨과 직립한 다리 자세를 특징으로 하는 활동적인 포식 생활 방식으로 구분되었다. 궁상목은 트라이아스기에 다양했으며, 모든 형태와 크기의 다양한 육상 및 반수생 포식자가 포함되었다. 머리가 크고 튼튼한 에리트로수쿠스과는 초기 트라이아스기에서 지배적인 육식 동물 중 하나였다. 식물룡류는 후기 트라이아스기에 번성한 특히 흔한 그룹이었다. 이 긴 주둥이를 가진 반수생 포식자는 살아있는 악어와 유사하며 비슷한 생활 방식을 가졌을 가능성이 높으며, 물가 주변에서 물고기와 작은 파충류를 사냥했다. 그러나 이러한 유사성은 표면적인 것에 불과하며 수렴 진화의 대표적인 사례이다.
진정한 궁룡류는 초기 트라이아스기에 나타나 조류 발목 골격류(새의 조상)와 가짜 악어류(악어의 조상)의 두 갈래로 나뉘었다. 조류 발목 골격류는 생태계의 작은 구성 요소였지만, 결국 후기 트라이아스기에 최초의 익룡과 공룡을 낳았다. 초기 긴 꼬리를 가진 익룡이 노리아기에 나타나 빠르게 전 세계로 퍼졌다. 트라이아스기 공룡은 카르니안기에 진화했으며 초기 용각류와 수각류를 포함한다. 대부분의 트라이아스기 공룡은 작은 포식자였으며, 1m에서 2m 길이의 ''Coelophysis''와 같은 몇몇 종만이 흔했다. 트라이아스기 용각류는 주로 세계의 더 시원한 지역에 서식했다.[58]
대형 포식자 ''스모크'' 역시 궁룡류였을 가능성이 높지만, 이것이 원시 공룡인지 가짜 악어류인지 확실하지 않다.
가짜 악어류는 트라이아스기에 훨씬 더 생태적으로 우세했으며, 대형 초식 동물(예: 에토사우루스류), 대형 육식 동물 ("라오이수키아류"), 최초의 악어상류 ("스페노수키아류")를 포함했다. 에토사우루스류는 후기 트라이아스기의 마지막 3천만 년 동안 흔했던 갑옷을 두른 파충류로, 트라이아스기-쥐라기 대멸종 때 멸종되었다. 대부분의 에토사우루스류는 초식성이었으며 낮은 곳에서 자라는 식물을 먹었지만, 일부는 고기를 먹었을 수도 있다. "라오이수키아류"(공식적으로는 파라크로코딜로모르파류로 알려짐)는 대부분의 트라이아스기 육상 생태계의 핵심 포식자였다. 거대한 사족 보행 사냥꾼, 날렵한 이족 보행 잡식 동물, 등을 따라 깊은 돛을 가진 둔한 짐승을 포함하여 25종 이상의 종이 발견되었다. 이들은 나중에 수각류가 채운 대형 포식자 틈새를 차지했을 가능성이 높다. "라오이수키아류"는 쥐라기까지 살아남은 유일한 가짜 악어류인 작고 가볍게 지어진 악어상류의 조상이 되었다.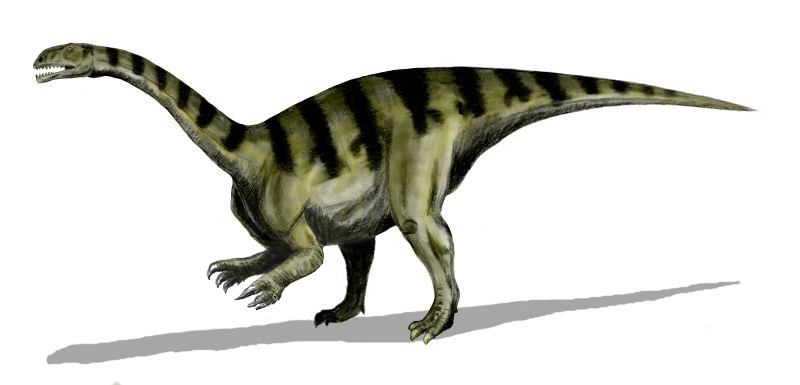
4. 2. 해양 파충류
트라이아스기에는 다양한 종류의 해양 파충류가 존재했다. 특히 테티스해 지역에서는 중기 트라이아스기에 파키플레우로사우루스와 노토사우루스 같은 사우롭테리지아가 흔했다. 또한 플라코돈트, 최초의 초식성 해양 파충류인 아토포덴타투스, 그리고 최초의 플레시오사우루스가 있었다. 도마뱀과 유사한 탈라토사우리아 (아스케프토사우루스)와 어룡이 초기 트라이아스기의 바다에 나타나 곧 다양해졌다. 중기 트라이아스기에 이르러 일부 어룡은 매우 큰 몸집을 갖게 되었다.[60]
이 시기에는 육상에서만 볼 수 있었던 파충류 중 일부가 바다로 진출했다.[81] 어룡, 헤엄치기에 특화된 지느러미 모양의 다리를 가진 플라코돈 등의 장경룡류, 탈라토사우루스류, 이판목 등이 이에 해당한다.[75][81]
4. 3. 기타 동물
수궁류는 트라이아스기에도 생존했으며, 키노돈트는 중요한 초식 동물 및 육식 동물이었다. 키노그나투스(Cynognathus)는 곤드와나의 올레네크기와 아니시안 시대의 특징적인 최상위 포식자였다.[61] 칸네마이에리아형 이궁류와 곰포돈트 키노돈트는 이 시기 대부분 동안 중요한 초식 동물이었다. 테로케팔루스는 대형 포식자(''모스코리누스'')와 초식 형태(바우리과)를 모두 포함했으며, 이들은 이 시기 중반에 멸종했다.[61] 에크테니니드 키노돈트는 후기 트라이아스기에 대형의 주행성 포식자 역할을 했다. 카르니아기 (후기 트라이아스기의 초기) 동안, 일부 진보된 키노돈트에서 최초의 포유류가 등장했다.[61]
Temnospondyl 양서류는 페름기-트라이아스기 대멸종에서 살아남은 그룹 중 하나였다.[55] 트라이아스기 생존자들은 수생 또는 반수생이었으며, 초기 트라이아스기에 모두 멸종된 ''Tupilakosaurus'', ''Thabanchuia'', Branchiosauridae 및 ''Micropholis''와 백악기까지 생존한 성공적인 Stereospondyli에 의해 대표되었다. ''Mastodonsaurus''와 같은 가장 큰 트라이아스기 스테레오스폰딜은 길이가 4m 에서 6m에 달했다.[55][56]
어류 중에서는 조기어류가 트라이아스기 초기에 다양화를 겪었다.[50] 실러캔스는 데본기 이후 초기 트라이아스기에 가장 높은 다양성을 보였다.[49] 사우릭티스와 비르게리아와 같은 육식성 조기어류는 길이가 1.2m 이상으로 자랐으며, 초기 트라이아스기에 나타나 이 기간 전체에 걸쳐 널리 퍼지고 성공했다.[51]
곤충 중에서는 완전변태곤충이 다양화되었다.[47] 중생대 트라이아스기 중기에 수생 곤충이 빠르게 다양해졌으며, 이 시기는 대부분의 현대 곤충 종을 포함하는 완전변태곤충의 중요한 다양화 시기를 나타낸다.[47] 침이 발달하지 않은 원시 벌목, 티타노프테라목(Titanoptera), 대벌레목, 노린재목이 있었다.
4. 4. 식물

육상 식물에서는 양치식물과 겉씨식물이 번성했다.[73] 소철류, 은행나무류, 볼치아목 등 다양한 겉씨식물이 나타났다. 습지에는 고대 석송이 풍부하게 남아 고사리와 속새도 조밀하게 분포했다.[75]
초기 트라이아스기에는 석탄 퇴적물이 거의 발견되지 않는데, 이를 "석탄 간극"이라고 부르며, 페름기-트라이아스기 대멸종의 영향으로 추정된다.[38]
영덕군의 영덕 심성암체는 트라이아스기의 화강암이다. 경상남도, 평안남도, 함경남도 등에서 트라이아스기 지층이 발견된다.
일본 기타카미 산지 남부 태평양 연안에 있는 미야기현 미나미산리쿠정 사라카이 마을에는 트라이아스기 후기 노리아절의 조개 화석 산지가 있으며, "사라카이 동물군" 또는 "사라카이 화석군"이라고 불린다.[73]
트라이아스기는 대멸종으로 끝났는데, 이는 특히 바다에서 심각했다. 이빨화석이 사라졌고, 어룡과 사경룡을 제외한 모든 해양 파충류도 사라졌다. 완족동물과 연체동물(예: 복족류)과 같은 무척추동물도 심각한 영향을 받았다. 해양에서는 해양 과의 22%와 해양 속의 약 절반이 사라졌을 가능성이 있다.[64]
트라이아스기 말 대멸종 사건이 모든 육상 생태계에서 똑같이 파괴적이지는 않았지만, 몇몇 중요한 크루로타르시 (이전에 테코돈트로 함께 분류된 대형 아르코사우루스 파충류)의 분기와 대부분의 대형 미로이빨 양서류, 소형 파충류 그룹, 그리고 대부분의 시냅시드가 사라졌다. 몇몇 초기 원시 공룡도 멸종했지만, 더 적응력이 뛰어난 공룡은 살아남아 쥐라기로 진화했다. 중생대 세계를 지배하게 된 살아남은 식물에는 현대의 구과 식물과 시카데오이드가 포함되었다.
후기 트라이아스기 멸종의 원인은 불확실하다. 이는 초대륙 판게아가 약 2억 200만 년에서 1억 9,100만 년 전(40Ar/39Ar 연대)에 분리되기 시작하면서 발생한 거대한 화산 폭발과 함께 일어났으며,[64] 중앙 대서양 마그마 지역(CAMP)을 형성했다.[65] 이는 지구가 처음 식고 안정된 이후 알려진 가장 큰 내륙 화산 사건 중 하나이다. 멸종 사건의 다른 가능성이지만 덜 가능성이 높은 원인으로는 지구 냉각 또는 심지어 운석 충돌이 있으며, 이로 인해 캐나다퀘벡의 마니쿠아간 저수지가 포함된 충돌구가 지목되었다. 그러나 마니쿠아간 충돌 용융물은 214±1 Mya로 연대가 측정되었다. 트라이아스기-쥐라기 경계의 시기도 최근에 Mya로 더욱 정확하게 고정되었다. 두 연대 모두 방사성 연대 측정의 더 정확한 형태, 특히 충돌 시 생성된 지르콘에서 우라늄이 납으로 붕괴되는 것을 사용하여 정확성을 높이고 있다. 따라서 증거는 마니쿠아간 충돌이 트라이아스기 말보다 약 10±2 Ma 앞섰음을 시사한다. 따라서 관찰된 대멸종의 직접적인 원인이 될 수 없었다.[66]
후기 트라이아스기 멸종의 수는 논쟁의 대상이다. 일부 연구에서는 트라이아스기 말에 최소 두 번의 멸종 기간이 있었고, 1,200만에서 1,700만 년의 간격이 있었다고 제시한다. 그러나 이에 반대하는 것은 북미 동물군에 대한 최근 연구이다. 페트리파이드 포레스트 국립공원 (Petrified Forest National Park)은 애리조나 북동부에 늦은 카르니안-초기 노리안 육상 퇴적물의 독특한 시퀀스가 있다. 2002년 분석에서는 고환경에 유의미한 변화가 없다는 것을 발견했다.[67] 그곳에서 가장 흔한 화석인 식물공룡은 속 수준에서만 변화를 겪었으며, 종의 수는 동일하게 유지되었다. 다음으로 가장 흔한 사지동물인 일부 에토사우루스와 초기 공룡은 변동 없이 통과했다. 그러나 식물공룡과 에토사우루스 모두 트라이아스기 말 대멸종 사건으로 완전히 멸종된 아르코사우루스 파충류 그룹에 속했다.
몇몇 초식 아르코사우로모르프 그룹이 멸종된 말-카르니안 멸종이 있었을 가능성이 있다. 대형 초식 수궁류인 카네메이에리아 디키노돈트와 트라베르소돈 키노돈트는 판게아 북쪽 절반(라시아)에서 크게 감소했다.
트라이아스기 내, 그리고 그 끝에서의 이러한 멸종은 공룡이 점유되지 않은 많은 생태적 지위로 확장될 수 있게 했다. 공룡은 점차 우세해지고 풍부해지고 다양해졌으며, 이후 1억 5천만 년 동안 그 상태를 유지했다. 진정한 "공룡 시대"는 트라이아스기가 아닌 그 뒤의 쥐라기 및 백악기 시대이다.
트라이아스기 말에 다시 약간 소규모의 대량 절멸이 있었다. 해양에서는 암모나이트의 많은 종이 사라졌고, 어룡 등의 해양 파충류도 타격을 입었다. 육상에서는 키노돈류, 디키노돈류의 대부분의 종을 비롯한 대량의 단궁류 (포유류형 파충류)가 멸종했다.[81] 트라이아스기 말기를 살아남은 공룡 등 육생 척추동물은 번식 방식 (알 등)과 생활 방식에서 건조에 특히 강한 타입으로 여겨진다.[81] 또한, 파충류도 단궁류와 마찬가지로 대형 동물을 중심으로 많은 종이 멸종했다. 아직 비교적 소형이었던 공룡은 트라이아스기 말기에는 용각류와 같은 대형 종도 출현하여, 그 후 급속도로 발전해 간다. 멸종의 원인으로는, 직경 3.3 - 7.8km 정도의 운석 낙하[87] 혹은 중앙 대서양 마그마 분포역 (Central Atlantic Magmatic Province)에서의 화산 활동과의 관련성이 지적되고 있다.[88][89][90] 이러한 환경 변화를 겪는 가운데, 수궁류는 생태계의 조연으로 변모했고, 한때 패권을 자랑했던 크루로타르시류는 사라져 갔다. 그리고 민첩하고 호흡 효율이 좋은 공룡이 생태계의 주역을 맡게 된다.[91] 또한 공룡의 선구로 등장한 실레사우루스류 또한 자손에게 자리를 내주었다.
[1]
논문
The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary
https://stratigraphy[...]
2020-12-08
[2]
논문
The Global Stratotype Sections and Point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains, Northern Calcareous Alps, Tyrol, Austria)
2013-09-01
[3]
사전
Triassic
[4]
웹사이트
International Stratigraphic Chart
https://stratigraphy[...]
International Commission on Stratigraphy
2023-07-19
[5]
논문
Recovery from the most profound mass extinction of all time
2008
[6]
논문
Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs
http://palaeo.gly.br[...]
2012-01-14
[7]
뉴스
'Lethally Hot' Earth Was Devoid of Life – Could It Happen Again?
http://news.national[...]
2012-10-19
[8]
서적
Beitrag zu einer Monographie des bunten Sandsteins, Muschelkalks und Keupers, und die Verbindung dieser Gebilde zu einer Formation
https://archive.org/[...]
J. G. Cotta
1834
[9]
논문
Carbon (δ13C) isotope variations indicate climate shifts and reflect plant habitats in the Middle Triassic (Anisian, Pelsonian) succession at Kühwiesenkopf / Monte Prà della Vacca (Dolomites, Northeast Italy)
https://www.scienced[...]
2022-12-01
[10]
논문
Disentangling climate signal from tectonic forcing: The Triassic Aghdarband Basin (Turan Domain, Iran)
https://www.scienced[...]
2023-01-10
[11]
서적
Earth History and Palaeogeography
Cambridge University Press
2022-05-16
[12]
서적
A Guide to the Sydney basin
Geological Survey of NSW
1980
[13]
웹사이트
Lecture 10 – Triassic: Newark, Chinle
http://rainbow.ldeo.[...]
[14]
서적
Encyclopedia of Dinosaurs
Academic Press
[15]
논문
Denudation surfaces and tectonics in the southernmost part of the Baltic Shield
[16]
논문
Deep weathering, neotectonics and strandflat formation in Nordland, northern Norway
2013
[17]
논문
Episodic burial and exhumation of the southern Baltic Shield: Epeirogenic uplifts during and after break-up of Pangaea
[18]
논문
The inheritance of a Mesozoic landscape in western Scandinavia
[19]
논문
Triassic eustatic variations reexamined
https://www.geosocie[...]
Geological Society of America
2018-12-01
[20]
문서
[21]
논문
The Pangaea Megamonsoon records: Evidence from the Triassic Mungaroo Formation, Northwest Shelf of Australia
https://www.scienced[...]
2023-01-09
[22]
논문
Triassic climates – State of the art and perspectives
[23]
논문
Phanerozoic paleotemperatures: The earth's changing climate during the last 540 million years
https://www.scienced[...]
2023-09-22
[24]
논문
Long-term cycles of Triassic climate change: a new δ18O record from conodont apatite
https://linkinghub.e[...]
2023-09-22
[25]
논문
Background Earth system state amplified Carnian (Late Triassic) environmental changes
https://linkinghub.e[...]
2023-09-22
[26]
논문
Discovery of a major negative 13C spike in the Carnian (Late Triassic) linked to the eruption of Wrangellia flood basalts
https://pubs.geoscie[...]
2023-09-22
[27]
논문
Palynological record of the Carnian Pluvial Episode from the northwestern Sichuan Basin, SW China
https://linkinghub.e[...]
2023-09-22
[28]
논문
A Middle–Late Triassic (Ladinian–Rhaetian) carbon and oxygen isotope record from the Tethyan Ocean
https://www.scienced[...]
2023-11-24
[29]
논문
Impact of 10-Myr scale monsoon dynamics on Mesozoic climate and ecosystems
2020-07-23
[30]
논문
Deep CO2 in the end-Triassic Central Atlantic Magmatic Province
2020-04-07
[31]
논문
Proliferation of Isoëtalean Lycophytes During the Permo-Triassic Biotic Crises: A Proxy for the State of the Terrestrial Biosphere
2021-03-02
[32]
논문
Bennettitalean Leaves From the Permian of Equatorial Pangea—The Early Radiation of an Iconic Mesozoic Gymnosperm Group
[33]
웹사이트
Caught between two mass extinctions: The rise and fall of Dicroidium
https://depositsmag.[...]
2023-09-23
[34]
논문
An overview of fossil Ginkgoales
https://linkinghub.e[...]
2009-03-01
[35]
논문
Conifer Diversity in the Middle Triassic: New Data from the Fossillagerstätte Kühwiesenkopf/Monte Prà della Vacca (Pelsonian, Anisian) in the Dolomites (Northeastern Italy)
https://www.journals[...]
2021-07-01
[36]
논문
A new Cheirolepidiaceae (Coniferales) from the Early Jurassic of Patagonia (Argentina): Reconciling the records of impression and permineralized fossils
https://bsapubs.onli[...]
2017-02-00
[37]
논문
Events near the time of the Permian-Triassic boundary
[38]
논문
Global coal gap between Permian-Triassic extinction and Middle Triassic recovery of peat-forming plants
[39]
논문
A unique geochemical record at the Permian/Triassic boundary
https://www.nature.c[...]
2023-11-24
[40]
논문
Carbon isotopic evidence for terminal-Permian methane outbursts and their role in extinctions of animals, plants, coral reefs, and peat swamps
https://cpb-us-e1.wp[...]
2020-12-14
[41]
웹사이트
How snowball Earth gave rise to complex life – Cosmos Magazine
https://cosmosmagazi[...]
2017-08-16
[42]
웹사이트
December: Phytoplankton News University of Bristol
https://www.bristol.[...]
[43]
웹사이트
The rise of algae in Cryogenian oceans and the emergence of animals – ResearchGate
https://www.research[...]
[44]
논문
Carbonate platform crisis in the Carnian (Late Triassic) of Hanwang (Sichuan Basin, South China): Insights from conodonts and stable isotope data
https://linkinghub.e[...]
2023-09-22
[45]
논문
Calcareous tubeworms of the Phanerozoic
http://www.kirj.ee/p[...]
2012-09-16
[46]
논문
Middle Triassic (Anisian) diversified bivalves: depositional environments and bivalve assemblages in the Leidapo Member of the Qingyan Formation, southern China
https://www.scienced[...]
2023-03-31
[47]
논문
Middle-Late Triassic insect radiation revealed by diverse fossils and isotopic ages from China
2018-09-07
[48]
논문
Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution
https://hal.science/[...]
2016-02-00
[49]
논문
New coelacanth material from the Middle Triassic of eastern Switzerland, and comments on the taxic diversity of actinistans
[50]
논문
A hiatus obscures the early evolution of Modern lineages of bony fishes
2021-01-00
[51]
논문
Marine Early Triassic Actinopterygii from Elko County (Nevada, USA): implications for the Smithian equatorial vertebrate eclipse
[52]
논문
Ceratodus tunuensis, sp. nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of central East Greenland
[53]
논문
Hybodont sharks from the Jurassic of Jaisalmer, western India
2021-08-04
[54]
서적
Fishes and the Break-up of Pangaea
https://doi.org/10.1[...]
Geological Society of London
[55]
서적
The Ecology and Behavior of Amphibians
https://books.google[...]
University of Chicago Press
[56]
서적
Vertebrate Palaeontology
https://books.google[...]
John Wiley & Sons
2009
[57]
서적
Evolution: The Story of Life
The Natural History Museum
[58]
논문
Climatic controls on the ecological ascendancy of dinosaurs
2023-01-09
[59]
논문
Early Triassic Marine Biotic Recovery: The Predators' Perspective
https://doi.org/10.1[...]
[60]
논문
New Finds of Triassic Marine Reptiles from Eastern Russia: Ammonoid Age Control and Possible Evidence for Ichthyopterygian Affinities
https://link.springe[...]
2024-08-13
[61]
논문
Selective Factors Associated with the Origin of Fur and Feathers
https://www.research[...]
2023-11-24
[62]
논문
A Mesozoic fossil lagerstätte from 250.8 million years ago shows a modern-type marine ecosystem
https://u-bourgogne.[...]
[63]
논문
Unexpected Early Triassic marine ecosystem and the rise of the Modern evolutionary fauna
[64]
논문
[65]
논문
Extensive 200-million-year-old continental flood basalts of the Central Atlantic Magmatic Province
[66]
논문
[67]
웹사이트
No Significant Nonmarine Carnian-Norian (Late Triassic) Extinction Event: Evidence From Petrified Forest National Park
http://gsa.confex.co[...]
2003-12-12
[68]
문서
基底年代の数値では、この表と本文中の記述では、異なる出典によるため違う場合もある。
[69]
문서
百万年前
[70]
뉴스
「始生代」の新名称、日本地質学会が2018年7月に改訂
2018-07-00
[71]
웹사이트
International Chronostratigraphic Chart(国際年代層序表)
http://www.geosociet[...]
日本地質学会
2020-04-17
[72]
문서
掘りたて恐竜展 展覧会図録
重慶自然博物館
[73]
논문
速水(2004)
[74]
논문
浜田・益富(1966)
[75]
논문
クルテン(1983)
[76]
웹사이트
INTERNATIONAL CHRONOSTRATIGRAPHIC CHART(国際年代層序表)
http://www.geosociet[...]
日本地質学会
2021-03-10
[77]
논문
斎藤(1979)
[78]
논문
フォーティ(2003)
[79]
문서
オーストリア共和国チロル州のセント・カシン
[80]
문서
ユリ型をしたウミユリ
[81]
문서
三畳紀の世界
掘りたて恐竜展 展覧会図録
[82]
서적
哺乳類型爬虫類 三畳紀の項
[83]
논문
Tetrapod association and palaeoenvironment of the Los Colorados Formation (Argentina): a significant sample from Western Gondwana at the end of the Triassic
2004
[84]
논문
The evolution of locomotor stamina in tetrapods: circumventing a mechanical constraint
1987
[85]
서적
生物の上陸
1982
[86]
서적
生物の上陸
1982
[87]
웹사이트
岐阜と大分から巨大隕石落下の証拠:最大で直径約8kmと推定
http://www.kumamoto-[...]
海洋研究開発機構
[88]
논문
Terrestrial and marine extinction at the Triassic-Jurassic boundary. synchoronized with major carbon cycle perturbation: A link to initiation of massive volcanism
2002
[89]
논문
Fossil plants and global warming at the Triassic-Jurassic boundary.
1999
[90]
논문
Macroecological responses of terrestrial vegetation to climatic and atmospheric change across the Triassic/Jurassic boundary in East Greenland.
2007
[91]
논문
Models for the rise of the dinosaurs
2014
[92]
웹인용
스콧 R. 쇼. <곤충 연대기>(원제
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com