배쪽 피개부
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
배쪽 피개부(VTA)는 중뇌에 위치하며, 흑색질과 인접해 있는 뇌 영역으로, 도파민성 뉴런이 밀집되어 있어 보상 회로, 동기 부여, 인지 기능, 약물 의존 등 다양한 기능에 중요한 역할을 한다. VTA는 전전두피질, 측좌핵, 편도체 등 여러 뇌 영역과 연결되어 있으며, 글루탐산, GABA, 콜린성 신경 전달 물질의 입력을 받는다. 주요 출력으로는 중변연계 경로와 중피질 경로가 있으며, 이들을 통해 전전두피질과 측좌핵으로 정보를 전달한다. VTA는 여러 하위 구역으로 나뉘며, 각 구역은 서로 다른 신경 전달 물질을 사용하고, 다양한 뇌 영역으로 투사한다. 또한, 약물 중독과 같은 질환과도 관련이 있으며, 갭 접합을 통한 빠른 신호 전달을 가능하게 하는 GABA성 뉴런 네트워크를 가지고 있다.
더 읽어볼만한 페이지
- 도파민 - 도파민성
도파민성은 도파민 전구체, 수용체 리간드, 수송체 및 대사 조절제 등을 포함하여 도파민 작용에 영향을 주는 약물과 물질, 그리고 그 효과를 포괄하는 용어이다. - 도파민 - 도파민작동성 세포군
- 중간뇌 - 흑색질
흑색질은 기저핵의 일부로 중뇌에서 가장 큰 핵이며, 안구 운동, 운동 계획, 보상 회로 등 다양한 뇌 기능에 관여하고, 특히 치밀부의 도파민성 뉴런은 운동 조절에 중요한 역할을 수행한다. - 중간뇌 - 아래둔덕
아래둔덕은 중뇌에 위치하여 청각 정보를 처리하고 음원 위치 결정 및 반사 작용에 중요한 역할을 하며, 다양한 뇌 영역과 연결되어 소리 분석 등의 기능을 수행한다. - 생리학 - 항상성
항상성은 생명체가 생존을 위해 체온, 혈당, 혈액 pH, 수분 및 이온 균형 등 내부 환경의 안정성을 유지하려는 경향으로, 음성 피드백 시스템, 신경계, 내분비계 등을 통해 조절된다. - 생리학 - 스트레스 요인
스트레스 요인은 개인이 환경적 요구에 적절히 대응하지 못할 때 심리적, 신체적 반응을 일으키는 자극으로, 지속시간, 강도, 예측 가능성, 통제 가능성에 따라 영향이 달라지며, 인지된 스트레스 척도 등으로 측정 가능하다.
| 배쪽 피개부 | |
|---|---|
| 일반 정보 | |
| 라틴어 이름 | area tegmentalis ventralis |
| 약칭 | VTA |
| 속하는 부분 | 중뇌 |
| 위치 | |
| 뇌 | 중뇌 바닥 |
| 구성 요소 | |
| 신경 세포 그룹 | 중뇌 피개(被蓋) |
| 혈액 공급 | |
| 동맥 | 해당 정보 없음 |
| 정맥 | 해당 정보 없음 |
| 기능 | |
| 역할 | 보상, 동기 부여, 인지 기능 |
| 관련 정보 | |
| 신경 전달 물질 | 도파민 |
2. 구조
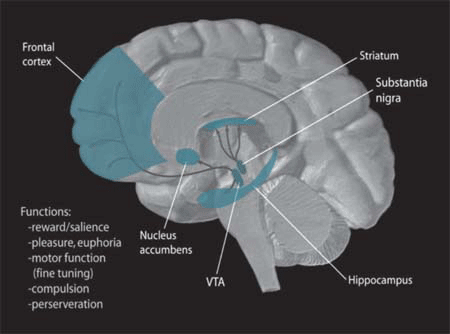
신경생물학자들은 인간 및 다른 영장류의 뇌에서 배쪽 피개부(VTA)를 흑색질 및 주변 핵과 구분하는 데 어려움을 겪어왔다. 원래 배쪽 피개부는 '핵'으로 지정되었지만, 해당 지역의 다양한 세포구조적 특징과 인접 지역과의 명확한 경계가 없다는 점 때문에 '영역'이라는 용어가 더 적절하게 사용되었다.[3] VTA는 A10으로 지정된다.[4]
배쪽 피개 영역은 중뇌에 위치하며, 유두체와 후시상하부는 간뇌에 포함되어 배쪽 피개 영역에서 머리쪽으로 뻗어 있다. 적색핵은 외측에, 눈돌림 신경 섬유는 배쪽 피개 영역의 배쪽 안쪽에 위치한다.[5] 교와 후뇌는 배쪽 피개 영역의 꼬리쪽에, 흑색질은 배쪽 피개 영역의 외측에 위치한다.[6]
2. 1. 하위 구분
VTA는 흑색질 주변 핵(PN), 팔다리 색소 영역(PBP), 반사 다발 주위 영역(PFR), 두부 중앙 피개 핵(RMTg)의 네 가지 주요 구역으로 나뉜다. PN과 PBP는 도파민성 세포가 풍부한 반면, PFR과 RMTg는 이러한 뉴런의 밀도가 낮다. PFR과 RMTg에는 크기가 작고 옅게 염색되는 티로신 수산화 효소(TH) 양성 세포체의 밀도가 낮고, RMTg는 대부분 GABA성 세포로 구성된다. 반면에 PN과 PBP는 중간에서 큰 크기의 TH 양성 세포체로 구성되며, 이 세포체는 적당하게 염색된다.[3]2. 2. 입력
VTA는 전전두피질, 다리교뇌 피개핵(PPTg), 외측배쪽 피개핵, 시상하핵, 선조체 종말상 핵, 상구, 중뇌수도 회백질, 외측 고삐, 등쪽 솔기 핵, 외측 시상하부, 시각전영역으로부터 글루탐산성 입력을 받는다.[8][7] 이러한 글루탐산성 입력은 VTA 세포 발화를 조절하며, 글루탐산성 뉴런이 활성화되면 VTA 도파민 뉴런의 발화율과 발화가 증가한다. 배쪽 피개 영역의 꼬리(tVTA, RMTg)는 GABA성 입력을 VTA에 투사하여 VTA 도파민 경로의 "마스터 브레이크" 역할을 한다.[9][10]VTA는 측좌핵, 복측 창백, 등쪽 솔기 핵, 외측 시상하부, 중뇌수도 회백질, 선조체 종말상 핵, 로스트로메디얼 피개핵(RMTg)으로부터 GABA성 입력을 받는다.[7] 외측 고삐는 RMTg GABA성 뉴런을 흥분시켜 VTA의 도파민성 뉴런에 억제 효과를 발휘하며, 이는 보상 예측 오류에 중요하다.[11]
다리교뇌 피개핵(PPTg)과 외측배쪽 피개핵으로부터의 콜린성 입력은 VTA 뉴런의 방전 특성을 강화하여 보상 회로에 조절적인 영향을 미친다.[13]
2. 3. 출력
VTA의 주요 출력 경로는 전전두피질로 가는 중변연계 경로와 측좌핵으로 가는 중피질 경로이다.[14][15] 중내측 고삐 경로는 도파민 외에 글루탐산과 GABA를 방출하는 VTA 뉴런으로 구성된다.[16][17] VTA는 편도체, 내후피질, 대상회, 해마, 측좌핵, 후각망울, 전전두피질 등으로 투사된다.[14]2. 4. 발달
포유류의 뇌 발달 과정에서 흑색질과 배쪽 피개부(VTA) 뉴런은 처음에는 등외측 및 내측 선조체 모두에 연결된다. 그러나 출생 시기가 되면 흑색질의 도파민성 뉴런은 배외측 선조체에만, 배쪽 피개부의 도파민성 뉴런은 내측 선조체에만 연결된다. 이러한 연결의 정교화는 불필요한 곁가지(측부 가지)를 제거함으로써 이루어진다.[1]3. 기능
VTA의 도파민 뉴런은 보상 회로, 동기 부여, 인지, 약물 의존 등 다양한 기능을 수행하며, 여러 정신 질환과 관련되어 있다.[19][20] VTA는 편도체로부터 감정 출력을 처리하고, 회피 및 공포 조건화에도 역할을 한다. VTA 뉴런은 새로운 자극, 예상치 못한 보상, 보상 예측 감각 신호에 반응하며, 보상 기대 오차를 인코딩한다.
헬렌 피셔의 연구에 따르면, 격렬한 사랑과 관련된 감정 상태는 VTA의 활동과 상관관계가 있으며, 이는 거부당한 파트너의 강박적인 행동을 설명하는 데 도움이 될 수 있다.[21]
VTA의 도파민 신경은 긍정적 및 부정적 강화 학습, 의사 결정, 작업 기억[37], 보상의 현저성(가치의 절대값)[38], 자극의 현저성, 혐오 자극[39][40]에서 중요한 역할을 담당한다.
3. 1. 갭 접합의 존재
VTA는 갭 접합으로 연결된 γ-아미노부티르산(GABA)성 신경세포 네트워크를 가지고 있어, 전기 전도를 통해 빠른 신호 전달이 가능하다.[22][23]3. 2. 신경 구성
배쪽 피개부(VTA)는 흑색질과 마찬가지로 멜라닌 색소를 함유한 도파민성 뉴런으로 구성되어 있다.[24] 최근 연구에 따르면 도파민성 뉴런은 VTA 내 전체 뉴런의 50-60%를 차지한다.[25] 이는 VTA 내 뉴런의 77%가 도파민성이라는 이전의 증거와는 차이가 있다.[26]기능적으로 구별되는 뇌 구조인 가쪽배쪽피개핵(RMTg)에는 상당수의 GABA성 뉴런이 존재한다.[9][10] 이 GABA성 뉴런은 전전두피질, 측좌핵, 청반 등 뇌 전체에 투사되는 도파민성 뉴런의 발화를 조절한다. VTA는 소수의 흥분성 글루탐산성 뉴런도 포함한다.
VTA의 도파민 신경은 다양한 동기 부여 행동에 기여하며, 다른 행동은 고유한 신경 회로를 가진 다른 도파민 신경에 의해 매개된다. VTA의 출력 신경 세포는 다른 뇌 영역뿐만 아니라 GABA 신경 및 글루탐산 신경의 정보도 통합하여 미세 회로를 형성한다. VTA의 GABA 신경 및 글루탐산 신경 일부는 도파민 신경과 동일한 뇌 영역에서 입출력을 받으며, VTA의 신경 세포 일부는 도파민과 함께 글루탐산 또는 GABA를 함께 방출한다.
3. 3. 변연계 루프
변연계 회로는 기저핵의 직접 경로 운동 회로와 매우 유사하다. 두 시스템 모두 피질에서 선조체(측좌핵)로의 주요 흥분성 입력이 있으며, 중뇌는 신경조절성 도파민 뉴런을 선조체로 투사하고, 선조체는 창백핵으로의 핵간 연결을 만들고, 창백핵은 시상으로의 출력을 가지며, 시상은 피질로 투사하여 회로를 완성한다. 변연계 회로는 피질 입력의 출처와 성질, 입력을 처리하는 선조체와 창백핵의 분할, 중뇌에서 도파민성 뉴런의 출처, 그리고 창백핵 출력의 시상 표적에 의해 운동 회로와 구별된다.변연계 회로는 인지 및 정동 기능을 제어하고 운동 회로는 운동을 제어한다.
3. 4. CA3 루프
CA3-VTA 연결은 외측 중격을 매개로 하여 보상 관련 맥락을 연결하는 데 중요한 역할을 한다. 2011년 연구진은 가성 광견병 바이러스(PRV)를 경시냅스 추적자로 사용하여 VTA에 주입했다. VTA에 단측 주입한 결과, 주입 후 48시간부터 CA3에서 양측 PRV 표지가 나타났다. VTA PRV 주입 전에 꼬리쪽 등쪽 외측 중격(cd-LS)을 손상시키면 CA3에서 PRV로 표지된 뉴런이 유의미하게 감소했다. CA3의 세타파 자극은 VTA의 도파민 세포 발화율을 증가시키고, VTA의 GABA 뉴런 발화율을 감소시켰다. VTA 뉴런의 정체성은 기록 뉴런의 신경생물틴 표지, 그리고 티로신 수산화 효소(TH)에 대한 조직학적 염색으로 확인되었다. GABA 효능제를 통해 CA3를 일시적으로 비활성화하면 정맥 내 코카인에 대한 지렛대 누르기의 맥락 유발 재발을 방지했다.[27]CA3의 글루탐산성 세포 활성화는 cd-LS의 GABA성 세포 활성화를 유발하고, 이는 VTA의 GABA 연접 뉴런을 억제하여 도파민 세포를 긴장성 억제로부터 해제하고, 도파민 세포의 발화율을 증가시키는 기능적 회로 루프가 제안되었다.[27]
3. 5. 보상 시스템
인간 뇌의 도파민 보상 회로는 복측 중뇌에서 핵 수염 복합체로 이어지는 두 개의 투사 시스템으로 구성된다. 첫째, 후내측 VTA와 중심 선상 라페 세포는 선택적으로 내측 선조체로 투사되며, 여기에는 내측 후각 결절과 내측 NAC 껍질이 포함된다. 둘째, 외측 VTA는 주로 외측 선조체로 투사되며, 여기에는 NAC 핵, 내측 NAC 껍질 및 외측 후각 결절이 포함된다. 이러한 경로는 각각 중-내측 선조체 및 중-외측 선조체 도파민 시스템이라고 불린다. 내측 투사 시스템은 정동 및 동기에 의해 특징지어지는 각성의 조절에 중요하며, 외측 투사 시스템은 목표 지향적 행동에 관여한다. 외측 부분과 달리 내측 부분은 보상 자극이 아닌 유해 자극에 의해 활성화된다.[28][29] 따라서 NAC 껍질과 후부 VTA가 보상 시스템과 관련된 주요 영역이다.VTA의 도파민 신경은 긍정적 및 부정적 강화 학습, 의사 결정, 작업 기억[37], 보상의 현저성(가치의 절대값)[38], 자극의 현저성, 혐오 자극[39][40]에서 중요한 역할을 담당한다.
3. 6. 질환
흑색질과 배쪽 피개부(VTA)의 도파민성 경로가 파괴되면 조현병, 파킨슨병, 주의력 결핍 과잉 행동 장애(ADHD)와 같은 여러 질환이 발생한다.[1] 현재 이러한 질환과 관련된 뉴런 간의 미묘한 차이점을 밝히고, 특정 도파민 투사를 선택적으로 치료하는 방법을 찾기 위한 연구가 진행 중이다.[1]3. 7. 약물 중독
측좌핵과 배쪽 피개 영역(VTA)은 중독성 약물이 작용하는 주요 부위이다. 코카인, 알코올, 아편유사제, 니코틴, 칸나비노이드, 암페타민 및 그 유사체와 같은 약물은 측좌핵에서 도파민의 작용을 연장시키거나, VTA에서 뉴런의 활성화를 자극하여 도파민이 강화 신호를 처리하는 신경 조절 영향을 변화시킨다.[30][31]이러한 약물의 남용은 도파민 방출을 자극하여 보상 및 정신 운동 효과를 유발한다. 반복적인 도파민 자극은 중변연계 도파민 시스템의 기능적 변화를 일으켜 강박적인 약물 복용 행동으로 이어진다. 중독된 개인의 VTA에서는 티로신 수산화 효소 활성이 증가하고, 흥분성 입력에 반응하는 뉴런의 능력도 증가하는데, 이는 전사 인자 CREB 활성 증가와 글루탐산의 AMPA 수용체의 중요한 서브유닛인 GluR1의 상향 조절 때문이다.
약물 금단 현상은 보상 기능 결핍으로 인해 발생하며, 약물이나 관련 자극에 노출되면 약물 탐구 행동이 다시 나타날 수 있다.[32][33][34]
4. 비교 해부학 및 진화
신경생물학자들은 인간 및 다른 영장류의 뇌에서 흑색질(SN) 및 주변 핵과 배쪽 피개부(VTA)를 구별하는데 종종 어려움을 겪었다. 원래 배쪽 피개부는 '핵'으로 지정되었지만, 시간이 지남에 따라 해당 지역의 다양한 세포구조적 특징과 인접 지역과의 명확한 경계가 없다는 점 때문에 '영역'이라는 용어가 더 적절하게 사용되었다.[3] 배쪽 피개부(VTA)에 대한 선택적인 변연계 관련 구심성 신경 섬유 때문에 VTA의 세포는 주변 세포와 구별하기 위해 A10으로 지정된다.[4]
1964년 이후의 모든 연구는 설치류에서 인간에 이르기까지 모든 포유류의 배쪽 피개부(VTA) 사이의 인상적인 일반적인 유사성을 강조해왔다. 이러한 연구들은 쥐, 토끼, 개, 고양이, 주머니쥐, 영장류, 인간에 초점을 맞췄다. A10 세포의 등쪽 범위 변화와 같은 약간의 차이점이 관찰되었다. 구체적으로 말하면, A10 세포의 등쪽 피크는 다른 포유류에 비해 영장류에서 더 넓게 나타난다. 또한, 배쪽 피개부의 도파민성 세포 수는 계통 발생적 진행에 따라 증가한다. 예를 들어, 쥐의 배쪽 피개부에는 약 25,000개의 뉴런이 있는 반면, 33세 남성의 배쪽 피개부에는 약 450,000개의 세포체가 존재한다.[35]
5. 중요 포인트 (일본어 문서 번역 및 요약)
VTA의 도파민 신경은 다양한 동기 부여 행동에 기여하며, 각 행동은 고유한 신경 회로를 가진 다른 도파민 신경에 의해 매개된다. VTA의 출력 신경 세포는 다른 뇌 영역의 정보뿐만 아니라 GABA 신경 및 글루탐산 신경의 정보도 통합하여 미세 회로를 형성한다. VTA의 GABA 신경 및 글루탐산 신경 일부는 도파민 신경과 동일한 뇌 영역에서 입출력을 받는다. VTA 신경 세포 일부는 도파민과 함께 글루탐산 또는 GABA를 함께 방출한다. 광유전학 연구는 강화 학습 및 동기 부여와 관련하여 고유한 기능을 가진 VTA 신경 세포의 발화 패턴 및 결합 형식을 밝혀냈다.[37][38][39][40][41][42][43][44]
참조
[1]
논문
Afferent projections to the ventral tegmental area of Tsai and interfascicular nucleus: a horseradish peroxidase study in the rat
1979-09
[2]
논문
Brain activation during human male ejaculation
2003-10
[3]
논문
Functional neuroanatomical review of the ventral tegmental area
https://dare.uva.nl/[...]
2019
[4]
논문
Dopamine neuron systems in the brain: an update
2007-05
[5]
논문
Intramesencephalic course of the oculomotor nerve fibers: microanatomy and possible clinical significance
[6]
논문
The anatomy of the human medial forebrain bundle: Ventral tegmental area connections to reward-associated subcortical and frontal lobe regions
2018-01-01
[7]
논문
Ventral tegmental area: cellular heterogeneity, connectivity and behaviour
2017-02-01
[8]
논문
Dynamic regulation of midbrain dopamine neuron activity: intrinsic, synaptic, and plasticity mechanisms
2011-12
[9]
논문
A new control center for dopaminergic systems: pulling the VTA by the tail
2012-11
[10]
논문
Braking dopamine systems: a new GABA master structure for mesolimbic and nigrostriatal functions
2012-10
[11]
논문
Neural Circuitry of Reward Prediction Error
2017-07
[12]
논문
Subpallidal outputs to the nucleus accumbens and the ventral tegmental area: anatomical and electrophysiological studies
http://www.sciencedi[...]
2020-10-04
[13]
논문
Segregated cholinergic transmission modulates dopamine neurons integrated in distinct functional circuits
2016
[14]
서적
Molecular Neuropharmacology: A Foundation for Clinical Neuroscience
McGraw-Hill Medical
[15]
논문
Magnesium in drug dependences
2008-03
[16]
논문
A Unique Population of Ventral Tegmental Area Neurons Inhibits the Lateral Habenula to Promote Reward
2013-11-20
[17]
논문
Single rodent mesohabenular axons release glutamate and GABA
2014-11
[18]
논문
Functional Connectome Analysis of Dopamine Neuron Glutamatergic Connections in Forebrain Regions
2015-12
[19]
논문
A VTA to basal amygdala dopamine projection contributes to signal salient somatosensory events during fear learning
2020-04
[20]
논문
Dopamine is necessary for cue-dependent fear conditioning
2009-09
[21]
논문
Sex and pairing status explain variations in the activation of nonapeptide receptors in song and motivation regions
2016-10
[22]
서적
Biology In Focus
2019
[23]
논문
Connexin-36 gap junctions mediate electrical coupling between ventral tegmental area GABA neurons
2006-07
[24]
웹사이트
Info
http://cogprints.org[...]
cogprints.org
2019-12-24
[25]
논문
The ventral tegmental area revisited: is there an electrophysiological marker for dopaminergic neurons?
2006-12
[26]
논문
Two types of neurone in the rat ventral tegmental area and their synaptic inputs
1992-05
[27]
논문
Linking context with reward: a functional circuit from hippocampal CA3 to ventral tegmental area
2011-07
[28]
논문
Dopamine reward circuitry: two projection systems from the ventral midbrain to the nucleus accumbens-olfactory tubercle complex
2007-11
[29]
논문
Phasic excitation of dopamine neurons in ventral VTA by noxious stimuli
2009-03
[30]
논문
Addiction and the brain
https://academic.oup[...]
1999
[31]
논문
A Neurobehavioral Approach to Addiction: Implications for the Opioid Epidemic and the Psychology of Addiction
2019
[32]
논문
Relapse to drug seeking following prolonged abstinence: the role of environmental stimuli
2008
[33]
논문
Chronic Stress, Drug Use, and Vulnerability to Addiction
2008
[34]
논문
Neurocircuitry of Addiction
2010
[35]
논문
Ventral tegmental (A10) system: neurobiology. 1. Anatomy and connectivity
http://cogprints.org[...]
1987-05
[36]
논문
Ventral tegmental area: cellular heterogeneity, connectivity and behaviour
2017
[37]
논문
Reward-motivated learning: mesolimbic activation precedes memory formation
2006
[38]
논문
The debate over dopamine's role in reward: the case incentive salience
2007
[39]
논문
Phasic excitation of dopamine neuron in the ventral VTA by noxious stimuli
2009
[40]
논문
Dopamine in Motivational control: Rewarding, Aversive and Alerting
2010
[41]
논문
A Unique Population of Ventral Tegmental Area Neurons Inhibits the Lateral Habenula to Promote Reward
2013
[42]
논문
Contemporary approaches to neural circuit manipulation and mapping: focus on reward and addiction
2014
[43]
논문
GABA Neurons of the VTA Drive Conditioned Place Aversion
2012
[44]
논문
VTA glutamatergic inputs to nucleus accumbens drive aversion by acting on GABAergic interneurons
2016
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com