영장류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
영장류는 쇄골, 손발의 엄지, 발가락, 뇌 발달 등의 특징을 가진 포유류의 한 목(目)으로, 나무 위 생활에 적응하며 다양한 특징을 갖는다. 굽은코원숭이아목(여우원숭이, 로리스 등)과 곧은코원숭이아목(안경원숭이, 원숭이, 유인원 등)으로 나뉘며, 뇌의 발달, 시각, 색각, 사회적 행동 등에서 특징을 보인다. 이동 방식, 성적 이형성, 인지 능력, 의사 소통 능력 등에서 다양성을 보이며, 생태적으로 열대 지역에 주로 서식한다. 인간과의 관계에서는 질병 전파, 법적 지위, 애완동물, 과학 연구, 보존 등의 측면에서 상호 작용하며, 멸종 위기에 놓인 종이 많다.
더 읽어볼만한 페이지
- 영장류 - 유인원
유인원은 꼬리 없는 영장류로, 인간을 포함하여 긴팔원숭이과와 사람과로 나뉘며, 높은 지능과 복잡한 사회 구조를 공유하지만 서식지 파괴와 사냥으로 멸종 위기에 처해 있다. - 영장류 - 원숭이
원숭이는 신세계원숭이와 구세계원숭이를 포함하는 영장류의 한 종류로, 크기와 몸무게, 서식지가 다양하며 인간과 유사한 특징을 지니고 있어 다양한 문화권에서 상징적인 의미를 지니고 있고 여러 목적으로 활용되지만 윤리적 논쟁 또한 존재한다. - 칼 폰 린네가 명명한 분류군 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다. - 칼 폰 린네가 명명한 분류군 - 잉어
잉어는 동아시아에 널리 분포하는 민물고기로, 하천 중류 이하의 진흙 바닥을 선호하며, 길조의 상징으로 여겨지며, 다양한 요리 재료로 활용되며, 기생충 감염 위험에 주의해야 한다.
| 영장류 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| |
| |
| |
| |
| |
| |
| |
| 학명 | Primates |
| 명명자 | Linnaeus, 1758 |
| 하위 분류군 계급 | 아목 |
| 하위 분류군 | 곡비원아목 직비원아목 †Altiatlasius |
| |
| 동의어 | Plesiadapiformes (cladistically 크라운 영장류 포함) |
| 무게 | 30g |
| 최대 무게 | 200kg |
| 생물학적 분류 | |
| 상목 | Euarchontoglires |
| 일반 정보 | |
| 와싱턴 협약 | 와싱턴 협약 부속서 II |
| 화석 범위 | 초기 팔레오세부터 현재 (65.9~0 백만 년 전, 가장 오래된 기록은 85백만 년 전) |
2. 어원
영어 ''primates''라는 이름은 고대 프랑스어 또는 프랑스어 primat프랑스어에서 유래되었으며, 이는 라틴어 primat-la의 명사형 용법, 즉 primusla('최고, 으뜸')에서 파생되었다.[3] 이 이름은 칼 린네가 이들을 동물의 "최고" 분류군이라고 생각했기 때문에 붙여졌다.[4]
영장류의 진화는 크게 세 단계로 나눌 수 있다. 첫째, 곡비원류와 직비원류의 분기, 둘째, 직비원류 내 안경원숭이하목과 원숭이하목의 분기, 셋째, 원숭이하목 내 광비원류(신세계원숭이)와 협비원류(구세계원숭이와 유인원)의 분기이다.
비교적 최근까지 영장류의 서로 다른 그룹 간의 관계가 명확하게 이해되지 않았기 때문에 일반적으로 사용되는 용어는 다소 혼란스러웠다. 예를 들어, 'ape'(유인원)은 'monkey'(원숭이)의 대체 용어 또는 꼬리가 없고 비교적 인간과 유사한 영장류를 지칭하는 데 사용되었다.[5][6]
3. 진화
영장류는 포유강 진수하강에 속하는 영장상목의 일원으로, 날원숭이목 및 나무두더쥐목과 함께 영장동물 계통군을 형성한다. 이 계통군은 설치동물 계통군(설치류와 토끼목)과 합쳐져 진(참)영장류상목을 이룬다.[270] 분자생물학적 연구에 따르면, 날원숭이목의 두 종은 나무두더쥐목보다 영장류와 더 밀접한 관계가 있다.[270]
영장류의 기원은 약 6,500만 년 전 백악기 말로 거슬러 올라가지만,[275] 화석 기록 상 가장 오래된 영장류는 플레시아다피스(약 5,500~5,800만년 전)이다.[276][277] 분자시계 연구는 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정한다.[278][279][280]

영장류는 다른 동물군에 비해 연구가 잘 되고 있음에도 불구하고, 최근에도 새로운 종들이 발견되고 있다. 2001년 콜린 그로브스의 《영장류 분류(''Primate Taxonomy'')》는 약 350종을 수록하고 있으며,[274] 《세계의 포유류 종, ''Mammal Species of the World''》(MSW3) 제3판에는 376종이 수록되어 있다.[264] 그러나 MSW3는 2003년판에 알려진 일부 종들을 누락하고 있으며, 기록된 종의 수는 424종, 아종을 포함하면 658종에 육박한다.
3. 1. 진화 초기
영장류의 계통은 약 6,500만 년 전 백악기 말 무렵에 시작된 것으로 추정된다.[251] 가장 오래된 영장류 화석은 백악기 말기 북아메리카 서부에서 발견된 플레시아다피스류(가짜 영장류)이다.
신생대 초기 팔레오세에는 북미에서 푸르가토리우스(''Purgatorius'')가 나타났으며,[30][26] 고신세 후기에는 아프리카에서 알티아틀라시우스(''Altiatlasius'')가,[27] 고신세-에오세 전환기에는 북반구 대륙에서 칸티우스(''Cantius''), 돈루셀리아(''Donrussellia''), 알타니우스(''Altanius''), 플레시아다피스(''Plesiadapis'') 및 테일하르디나(''Teilhardina'') 등이 나타났다.[28][29][30] 분자 시계 연구에서는 영장류 계통의 기원이 약 8,500만 년 전 백악기 중기까지 거슬러 올라간다고 추정한다.[31][32][33]
분지학적 관점에서 영장목은 단계통군이다. 곡비원아목("젖은 코" 영장류)은 약 6,300만 년 전 원시 영장류 계통에서 분리되었으며,[34] 여우원숭이 관련 5개 과와 로리스과, 갈라고과로 구성된 2개 과를 포함한다.[1][36] 에오세 동안 북반구 대륙은 아다피스류와 오모미스류가 지배했다.[42][37] 아다피스류는 곡비원류의 일종으로 간주되지만, 최근 연구에서는 다위니우스 마실라에(''Darwinius masillae'')가 이 그룹에 속한다는 것이 밝혀졌다.[38] 오모미스류는 안경원숭이, 원숭이, 유인원과 밀접한 관련이 있었으며, 약 3,000만 년 전에 멸종했다.[37] 반면 아다피스류는 약 1,000만 년 전까지 생존했다.[39]
유전학 연구에 따르면 마다가스카르의 여우원숭이는 약 7,500만 년 전에 로리스류와 분기되었다.[35] 5,000만 년에서 8,000만 년 사이에 뗏목 이동을 통해 아프리카에서 마다가스카르로 이동한 것으로 추정된다.[35][40][41]
직비원아목("마른 코" 영장류)은 안경원숭이하목과 원숭이하목으로 구성된다. 안경원숭이하목은 약 5,800만 년 전에 나타났으며,[43][44] 가장 오래된 직비원류 화석은 중국에서 발견된 아르키케부스(''Archicebus'')이다.[45] 원숭이하목은 약 4,000만 년 전에 등장했으며,[37] 광비원소목(신세계원숭이)과 협비원소목(구세계원숭이, 사람 등 유인원)으로 나뉜다.[1] 신세계원숭이는 에오세 동안 섬 건너뛰기 또는 뗏목 이동을 통해 아프리카에서 남아메리카로 이동했을 것으로 추정된다.[42]
유인원과 원숭이는 중신세부터 아프리카에서 유럽과 아시아로 퍼져나갔다.[51] 최초의 호미닌 화석은 북아프리카에서 발견되었으며, 500만~800만 년 전으로 거슬러 올라간다.[37] 구세계원숭이는 약 180만 년 전에 유럽에서 사라졌다.[52] 분자 및 화석 연구에 따르면 현생 인류는 10만~20만 년 전 아프리카에서 기원했다.[53]
3. 2. 곡비원아목과 직비원아목의 분기
영장류는 진수하강에 속하는 영장상목 계통군의 일부이다. 분자생물학적 연구에 따르면, 날원숭이목의 두 종은 나무두더쥐목보다 영장류와 더 밀접한 관계가 있다.[270] 이들 세 목은 영장동물 계통군을 형성하며, 이는 설치동물 계통군과 합쳐져 영장상목 계통군을 이룬다.
영장류 계통 분류는 약 6500만 년 전으로 거슬러 올라가지만, 화석 기록으로 남아 있는 가장 오래된 영장류는 백악기 후기의 플레시아다피스(약 5,500~5,800만년 전)이다.[276][277] 분자시계 연구는 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정한다.[278][279][280]
곡비원아목은 약 6300만 년 전에[281] 원시 영장류 계통에서 분리된 것으로 추정된다.[282] 에오세 동안, 북반구 대륙에는 아디피드와 오모미드가 살고 있었다.[289][284] 오모미드는 약 3000만 년 전에 사라졌고,[284] 아다피드는 약 1,000만년 전까지 살아남았다.[286]
직비원아목은 안경원숭이와 원숭이하목으로 나뉜다.[264] 안경원숭이는 약 5800만 년 전에 가장 원시적인 부류를 대표한다.[290][291]
원숭이하목은 약 4000만 년 전에 등장했으며,[284] 신세계원숭이를 포함하는 광비원소목과 구세계원숭이와 사람 등의 유인원을 포함하는 협비원소목으로 나뉜다.[264] 에오시미드는 아시아에서 발달했지만 수백만 년 전에 멸종되었다.[292]
신세계원숭이의 기원은 명확하지 않지만, 분자적 서열 연구에서는 3500만 년 전에 협비원류와 분화되었음을 시사한다.[293] 진원류는 아프리카에서 기원하여 일부가 남아메리카로 이동했을 것으로 추정된다.[289] 대륙 이동설에 따르면, 1 kg의 작은 영장류가 뗏목을 타고 대서양을 건너 남아메리카에 도착했을 가능성이 있다.[289][294]
유인원과 원숭이들은 마이오세가 시작될 무렵에 아프리카에서 유럽과 아시아로 퍼졌다.[295] 가장 오래된 인류 화석은 500-800만년 전으로 거슬러 올라가며, 북아프리카에서 발견되었다.[284] 구세계원숭이들은 약 180만년 전에 유럽에서 사라졌다.[296] 현세 인류는 10-20만년 전 아프리카에 기원을 두고 있다고 본다.[297]
3. 2. 1. 여우원숭이의 마다가스카르 이주
발생학적 연구에 의하면, 마다가스카르의 여우원숭이는 얼추 7500만 년 전에 로리스하목에서 갈라졌다.[282] 이들 연구는 염색체와 분자적 증거뿐만 아니라 여우원숭이가 다른 곡비원류 영장류보다 서로 밀접한 관련이 있음을 보여준다.[282][287] 그러나 마다가스카르는 1억 6천만 년 전에 아프리카 대륙에서, 9천만 년 전에 인도에서 분리되었다.[288] 여우원숭이들이 다른 곡비원류 영장류보다 서로 더 밀접한 관계에 있기 때문에, 일단의 매우 적은 규모의 선조 개체군이 5천만 년 전과 8천만 년 전 사이에 일종의 단일한 뗏목 사건을 통해 마다가스카르로 왔기 때문으로 생각된다.[282][287][288] 아프리카와 인도로부터의 여러 차례의 이주와 같은, 기타 여러 이주 조건들을 시험해봤으나 어느 것도 유전적이며 분자학적인 증거에 의해 뒷받침되지 않았다.[289]
최근까지 아이아이는 곡비원류로 분류하기 어려웠다.[264] 아이아이과를 여우원숭이하목 영장류(여우원숭이와 로리스로 분리되는 것보다 더 최근에 여우원숭이 계열에서 분리되는 것을 의미)의 일종으로 분류하거나 다른 모든 곡비원류의 자매군의 일종으로 분류하는 이론이 제안되었다. 2008년에, 아이아이과는 여우원숭이하목이 되어야 하고, 뗏목을 타고 섬으로 건너 온 같은 선조 여우원숭이 개체군의 자손임이 확인되었으며, 아이아이과는 결과적으로 자신의 이름을 딴 아이아이하목에서 나와서, 여우원숭이하목에 속하게 되었다.[282]
3. 3. 직비원아목의 진화
영장목은 영장상목의 일원으로, 포유강의 진수하강에 속한다. 최근의 분자생물학적 연구에 따르면, 날원숭이류는 나무두더쥐류보다 영장류와 더 밀접한 관련이 있다.[270] 한때 나무두더쥐류는 영장류로 간주되기도 했다.[271] 이들 세 목은 영장동물 계통군을 형성하며, 설치동물 계통군(설치류와 토끼류)과 합쳐져 영장상목 계통군을 이룬다. 일부 과학자들은 날원숭이류를 영장류의 아목으로 간주하여 진영장아목(Euprimates)으로 부르기도 한다.[272]
영장류 계통 분류는 최소 6500만 년 전까지 거슬러 올라가지만,[275] 화석 기록으로 남아 있는 가장 오래된 영장류는 백악기 후기의 플레시아다피스(약 5,500~5,800만년 전)이다.[276][277] 분자시계 연구는 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정한다.[278][279][280]
분지학적 추정에 따르면, 영장목은 단계통군이다. 곡비원아목(구부러진 코 또는 젖는 코를 지닌 영장류)은 일반적으로 약 6300만 년 전에[281] 원시 영장류 계통에서 분리된 것으로 생각된다.[282] 곡비원류는 여우원숭이 관련 5개 과와 로리스과, 갈라고과를 포함하여 7개의 과로 이루어져 있다.[264][283] 에오세 동안, 북반구 대륙의 대부분은 아다피드와 오모미드가 우위를 점하며 살고 있었다.[289][284] 아다피드는 곡비원류의 일종으로 간주되지만, 현대의 여우원숭이들처럼 촘촘한 치아를 갖고 있지는 않다. 최근의 연구는 이 집단을 다위니우스 마실라에(''Darwinius masillae'')라는 이름으로 제안하고 있다.[285] 오모미드는 안경원숭이, 원숭이, 유인원과 밀접한 관련이 있으며 약 3000만 년 전에 사라졌고,[284] 아다피드는 약 1,000만년 전까지 살아남았다.[286]
발생학적 연구에 따르면, 마다가스카르의 여우원숭이는 대략 7500만 년 전에 로리스하목에서 갈라졌다.[282] 여우원숭이가 다른 곡비원류 영장류와 서로 밀접한 관련이 있다는 사실은 염색체와 분자적 증거를 통해 알 수 있다.[282][287] 마다가스카르는 1억 6천만년 전에 아프리카 대륙에서, 9천만 년 전에 인도에서 분리되었다.[288] 매우 적은 규모의 선조 개체군이 5천만 년 전과 8천만 년 전 사이에 일종의 뗏목 사건을 통해 마다가스카르로 왔기 때문으로 추정된다.[282][287][288]
아이아이는 최근까지 곡비원류로 분류하기 어려웠으나,[264] 2008년에 아이아이과는 여우원숭이하목에 속하며, 뗏목을 타고 섬으로 건너 온 같은 선조 여우원숭이 개체군의 자손임이 확인되었다.[282]
직비원아목(단순한 코 또는 "마른 코"를 갖고 있는 영장류)은 안경원숭이과(안경원숭이하목에 속하는)의 원원류 안경원숭이(약 5800만 년 전에 가장 원시적인 부류를 대표[290][291])와 원숭이하목으로 이루어져 있다.
가장 오래된 인류 화석은 500-800만년 전으로 거슬러 올라가며, 북아프리카에서 발견되었다.[284] 구세계원숭이들은 약 180만년 전에 유럽에서 사라졌다.[296] 분자학적 연구와 화석 연구는 현세 인류가 10-20만년 전 아프리카에 기원을 두고 있다고 본다.[297]
영장류는 다른 동물군에 비해 연구가 잘 되고 있음에도 불구하고, 최근에도 몇몇 새로운 종들이 발견되고 있다. 2001년 콜린 그로브스의 《영장류 분류(''Primate Taxonomy'')》는 약 350종을 수록하고 있으며,[274] 《세계의 포유류 종, ''Mammal Species of the World''》(MSW3) 제3판에는 376종이 수록되어 있다.[264] 그러나 MSW3는 2003년판에 알려진 일부 종들을 누락하고 있으며, 기록된 종의 수는 424종, 아종을 포함하면 658종에 육박한다.
3. 3. 1. 진원하목
원숭이하목은 약 4000만 년 전에 등장했으며,[284] 광비원소목(신세계원숭이)과 협비원소목(구세계원숭이 및 사람 등의 유인원)의 두 분류군으로 나뉜다.[264] 에오시미드는 아시아에서 발달했지만 수백만 년 전에 멸종된 세 번째 분류군이다.[292]
신세계원숭이의 기원은 여우원숭이처럼 명확하지 않다. 분자 서열 연구는 광비원류와 협비원류의 분화 시기를 3300만~7000만 년 전으로 추정하는 반면, 미토콘드리아 염기 서열 연구는 3500만 년 전으로 추정한다.[293] 진원류는 아프리카에서 기원하여 일부가 이동한 것으로 추정되며, 발견되는 표본들도 이를 뒷받침한다.[289] 진원류 영장류는 에오세 동안 대서양의 융기된 능선과 낮은 해수면을 이용하여 섬들을 징검다리 삼아 남아메리카로 이동했을 것으로 추정된다.[289] 이는 뗏목을 이용한 대륙간 이주로 설명할 수 있다. 대륙 이동설에 따르면 당시 대서양은 오늘날처럼 넓지 않았으며,[289] 연구 결과는 1kg의 작은 영장류가 식물 뗏목에서 13일 동안 생존할 수 있음을 보여준다.[294] 해류와 풍속을 고려하면 대륙 간 항해에 충분한 시간이었다.
유인원과 원숭이들은 마이오세가 시작될 무렵 아프리카에서 유럽과 아시아로 퍼졌으며,[295] 곧이어 로리스와 안경원숭이도 같은 여정을 떠났다.
협비원류와 광비원류가 분기된 후, 사람상과와 꼬리감는원숭이상과는 2,800만 년에서 2,400만 년 전에 분기되었다.[260][261] 이 시기에 요산 산화 효소 활성이 소실된 것으로 추정되는데,[262] 이는 요산이 항산화 물질로서 비타민 C를 부분적으로 대체할 수 있기 때문이다.[263]
4. 분류
칼 폰 린네는 《자연의 체계》 제10판에서 영장목을 사람속(''Homo''), 시미아(''Simia''), 여우원숭이속(''Lemur''), 박쥐속(''Vespertilio'')으로 분류했다.[12] '영장류'의 '영(霊)'은 넋이나 귀신이라는 의미보다는 뛰어나고 신비한 힘을 가졌다는 의미로, 사람을 포함한 원숭이 무리를 동물 진화의 최종 형태로 인식하여 붙여진 이름이다. 영어 이름 Primate영어 역시 대주교나 최고위를 의미하는 단어이다.
이전에는 뇌가 작고 후각이 발달했으며 코가 긴 여우원숭이류, 로리스류, 안경원숭이류를 원원아목(Prosimii)으로, 뇌가 크고 시각이 발달했으며 코가 짧은 분류군을 진원아목(Anthropoidea)으로 묶었다.[239] 그러나 연구 결과, 안경원숭이가 원원류의 다른 그룹보다 진원류에 더 가깝다는 것이 밝혀졌다. 이에 따라 현재는 여우원숭이류, 로리스류를 묶어 '''굽은코원숭이류(굽은코원숭이아목, 굽은코류, 굽은코아목)''', 안경원숭이류를 포함한 그 외 영장류를 '''곧은코원숭이류(곧은코원숭이아목, 곧은코류, 곧은코아목)'''라고 부르며, 정식 분류 체계에서는 "원원류"라는 명칭은 사용하지 않는다.[250]
1984년 앤더슨과 존스가 굽은코원숭이류와 곧은코원숭이류의 분류를 도입하기 전,[15] 영장류는 원원류와 진원류의 두 상과로 나뉘었다.[303]
4. 1. 현존하는 영장류 분류
영장목은 영장상목의 계통 분류의 일부로, 포유강의 진수하강에 속한다. 최근의 분자생물학적 연구에 따르면, 날원숭이류의 두 종은 나무두더쥐류보다는 영장류와 더 밀접한 관계가 있다.[270] 이 세 목은 영장동물 계통군을 형성하며, 이는 설치동물 계통군(설치류와 토끼류)과 합쳐져 영장상목 계통군을 이룬다.영장류 계통 분류는 최소 6500만 년 전까지 거슬러 올라가지만,[275] 화석 기록으로 남아 있는 가장 오래된 영장류는 백악기 후기의 플레시아다피스(약 5,500~5,800만년 전)이다.[276][277] 분자시계 연구를 포함한 다른 연구에서는 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정한다.[278][279][280]
분지학적 추정에 따르면, 영장목은 단계통군이다. 곡비원아목은 약 6300만 년 전에[281] 원시 영장류 계통에서 분리된 것으로 추정된다.[282] 7개의 곡비원류 과는 5개의 여우원숭이 관련 과와 나머지 로리스과, 갈라고과로 구성된다.[264][283] 에오세 동안, 북반구 대륙의 대부분은 아디피드와 오모미드가 우세하게 살고 있었다.[289][284]
발생학적 연구에 따르면, 마다가스카르의 여우원숭이는 약 7500만 년 전에 로리스하목에서 갈라졌다.[282] 그러나 마다가스카르는 1억 6천만년 전에 아프리카 대륙에서, 9천만 년 전에 인도에서 분리되었다.[288] 여우원숭이들이 다른 곡비원류 영장류보다 서로 더 밀접한 관계에 있기 때문에, 소규모 선조 개체군이 5천만 년 전과 8천만 년 전 사이에 뗏목 사건을 통해 마다가스카르로 왔을 것으로 추정된다.[282][287][288]
직비원아목은 안경원숭이과(안경원숭이하목에 속하는 단계통군)와 원숭이하목으로 구성된다.[264] 안경원숭이는 약 5800만 년 전에 가장 원시적인 부류를 대표한다.[290][291]
원숭이하목은 약 4000만 년 전에 등장했으며,[284] 신세계원숭이를 포함하는 광비원소목과 구세계원숭이와 사람 등의 유인원을 포함하는 협비원소목으로 나뉜다.[264]
마이오세가 시작될 무렵, 유인원과 원숭이들은 아프리카에서 유럽과 아시아로 퍼졌다.[295] 가장 오래된 인류 화석은 500-800만년 전으로 거슬러 올라가며, 북아프리카에서 발견되었다.[284] 일부 분자학적 연구와 화석 연구는 현세 인류가 10-20만년 전 아프리카에 기원을 두고 있다고 본다.[297]
2001년 콜린 그로브스의 《영장류 분류(''Primate Taxonomy'')》에는 약 350종이 수록되었으며,[274] 《세계의 포유류 종, ''Mammal Species of the World''》(MSW3)의 제3판에는 376종이 수록되었다.[264] 기록된 종의 수는 계속 증가하여 424종, 아종을 포함하면 658종에 이른다.
다음은 영장류의 목록이다.[264][283][298]
| 목 | 아목 | 하목 | 과 | 종 수 |
|---|---|---|---|---|
| 영장목 | 곡비원아목 | 여우원숭이하목 | 난쟁이여우원숭이과 | 40 |
| 아이아이과 | 1 | |||
| 여우원숭이과 | 22 | |||
| 족제비여우원숭이과 | 27 | |||
| 인드리과 | 19 | |||
| 로리스하목 | 로리스과 | 14 | ||
| 갈라고과 | 22 | |||
| 직비원아목 | 안경원숭이하목 | 안경원숭이과 | 13 | |
| 원숭이하목 | 광비원류: 신세계원숭이 | 비단원숭이과 | 49 | |
| 꼬리감는원숭이과 | 14 | |||
| 올빼미원숭이과 | 11 | |||
| 사키원숭이과 | 57 | |||
| 거미원숭이과 | 29 | |||
| 협비원류 | 긴꼬리원숭이상과 | 151 | ||
| 사람상과 | 긴팔원숭이과 | 18 | ||
| 사람과 | 8 |

영장목은 1758년 린네의 《자연의 체계》 제10판[299]에서 사람속(''Homo''), 시미아(''Simia''), 여우원숭이속(''Lemur''), 박쥐속(''Vespertilio'')으로 명명되었다.
1984년 앤더슨과 존스가 곡비원류와 직비원류라는 분류를 도입하기 전,[302] 영장류는 원원류(Prosimii)와 진원류(Anthropoidea)의 두 상과로 나뉘었다.[303]
"영장류"의 '영(霊)'은 넋이나 귀신이라는 의미보다는, 뛰어나고 신비한 힘을 가졌다는 의미이다. 사람이나 사람을 포함한 원숭이 무리를 동물 진화의 최종 형태로 인식하여 붙여진 이름이다. 영어 이름 Primate영어 역시 대주교나 최고위를 의미하는 단어이다.
이전에는 뇌가 작고 후각이 발달했으며 코가 긴 여우원숭이류, 로리스류, 안경원숭이류를 원원아목으로, 뇌가 크고 시각이 발달했으며 코가 짧은 분류군을 진원아목으로 묶었다.[239] 현재는 여우원숭이류, 로리스류를 '''굽은코원숭이류'''(굽은코원숭이아목, 굽은코류, 굽은코아목), 안경원숭이류를 포함한 그 외 영장류를 '''곧은코원숭이류'''(곧은코원숭이아목, 곧은코류, 곧은코아목)라고 부르며, 정식 분류 체계에서는 "원원류"라는 명칭은 사용하지 않는다.[250]
아래의 분류는 일본 몽키 센터 영장류 한국어 명칭 편찬 워킹 그룹 (2018)을 따른다.[240] 단, 하목의 한국어 명칭은 가와다 등 (2018)을 따른다.[241]
- 직비원아목
- * 안경원숭이하목
- ** 안경원숭이과
- * 진원하목
- ** 협비소목
긴꼬리원숭이상과
* 긴꼬리원숭이과
사람상과
* 사람과
* 긴팔원숭이과
- ** 광비소목
꼬리감는원숭이상과
* 거미원숭이과
* 꼬리감는원숭이과 (밤원숭이과를 분할하는 설도 있음[238])
사키상과
* 사키과
- 굽은코원아목
- * 여우원숭이하목
- ** 여우원숭이상과
난쟁이여우원숭이과
인드리과
여우원숭이과
갈기여우원숭이과
- ** 아이아이상과
아이아이과
- * 로리스하목
- ** 갈라고과
- ** 로리스과
4. 2. 계통 분류
다음은 영장상목의 계통 분류이다.[267][268][269]{| class="wikitable"
|-
! 영장상목
|-
|
{| class="wikitable"
|-
! 설치동물
|
|-
! 영장동물
|
{| class="wikitable"
|-
| 나무두더지목
|-
! 영장형류
|
{| class="wikitable"
|-
| 날원숭이목
|-
|
|}
|}
|}
|}
영장목은 영장상목 계통군의 일부로, 포유강의 진수하강에 속한다. 최근의 분자생물학적 연구에 따르면, 날원숭이류의 두 종은 나무두더쥐류보다 영장류에 더 가깝다.[270] 한때 나무두더쥐류를 영장류로 간주하기도 했다.[271] 이들 세 목은 영장동물 계통군을 형성하며, 설치동물 계통군(설치류와 토끼류)과 합쳐 영장상목 계통군을 이룬다. 영장동물과 영장상목 둘 다 상목으로 분류하기도 한다. 일부 과학자들은 날원숭이류를 영장류의 아목으로 간주하여 진영장아목(眞靈長亞目, Euprimates)으로 부르기도 한다.[272]
다음은 영장류의 계통 분류이다.[273][274][274]
{| class="wikitable"
|-
! 영장형류
|-
|
{| class="wikitable"
|-
| 날원숭이목 
|-
! 영장목
|
{| class="wikitable"
|-
! 직비원아목
|
{| class="wikitable"
|-
! 원숭이하목
|
{| class="wikitable"
|-
! 협비원소목
|
{| class="wikitable"
|-
! 사람상과
|
{| class="wikitable"
|-
! 사람과
|
{| class="wikitable"
|-
! 사람아과
|
{| class="wikitable"
|-
! 사람족
|


|-
| 고릴라족 
|}
|-
| 오랑우탄아과 
|}
|-
| 긴팔원숭이과 
|}
|-
| 구세계원숭이 (긴꼬리원숭이상과)
|}
|-
| 신세계원숭이 (광비원소목) 
|}
|-
! 안경원숭이하목
|

|}
|-
! 곡비원아목
|
{| class="wikitable"
|-
! 여우원숭이하목
|
여우원숭이상과  |
로리스상과  |
|}
|}
|}
|}
영장류 계통 분류는 최소 6500만 년 전까지 거슬러 올라가는 것으로 추정되지만,[275] 화석 기록으로 남아 있는 가장 오래된 영장류는 백악기 후기의 플레시아다피스(약 5,500~5,800만년 전)이다.[276][277] 분자시계 연구를 포함한 다른 연구를 통해, 영장류 분화의 기원을 백악기 중기 약 8,500만년 전 경으로 추정하고 있다.[278][279][280]
근대 분지학적인 추정에 의하면, 영장목은 단계통군이다. 곡비원아목은 일반적으로 약 6300만 년 전에[281] 원시 영장류 계통에서 분리된 것으로 생각하고 있다.[282] 7개의 곡비원류 과는 5개의 여우원숭이 관련 과와 나머지 로리스과, 갈라고과로 이루어져 있다.[264][283] 이전의 분류 체계는 족제비여우원숭이과를 여우원숭이과에, 갈라고과를 로리스과에 포함시켜 분류하여, 현재의 분류 체계인 5+2개 과 분류 대신에 3+2개 과로 분류를 나누었다.[264] 에오세 동안, 북반구 대륙의 대부분은 2개의 그룹 아디피드와 오모미드가 우위를 점하며 살고 있었다.[289][284] 전자는 곡비원류의 일종으로 간주하지만, 현대의 여우원숭이들처럼 촘촘한 치아를 갖고 있지는 않다. 최근의 연구는 이 집단을 다위니우스 마실라에(''Darwinius masillae'')라는 이름으로 제안하고 있다.[285] 후자는 안경원숭이와 원숭이 그리고 유인원과 밀접한 관련이 있다. 이 두 그룹이 현존하는 영장류와 정확히 어떻게 관련이 있는지는 명확치 않다. 오모미드는 약 3000만 년 전에 사라졌고,[284] 반면에 아다피드는 약 1,000만년 전까지 살아남았다.[286]
발생학적 연구에 의하면, 마다가스카르의 여우원숭이는 얼추 7500만 년 전에 로리스하목에서 갈라졌다.[282] 이들 연구는 염색체와 분자적 증거뿐만 아니라 여우원숭이가 다른 곡비원류 영장류보다 서로 밀접한 관련이 있음을 보여준다.[282][287] 그러나 마다가스카르는 1억 6천만년 전에 아프리카 대륙에서, 9천만 년 전에 인도에서 분리되었다.[288] 여우원숭이들이 다른 곡비원류 영장류보다 서로 더 밀접한 관계에 있기 때문에, 일단의 매우 적은 규모의 선조 개체군이 5천만 년 전과 8천만 년 전 사이에 일종의 단일한 뗏목 사건을 통해 마다가스카르로 왔기 때문으로 생각된다.[282][287][288] 아프리카와 인도로부터의 여러 차례의 이주와 같은, 기타 여러 이주 조건들을 시험해봤으나 어느 것도 유전적이며 분자학적인 증거에 의해 뒷받침되지 않았다.[289]
최근까지 아이아이는 곡비원류로 분류하기 어려웠다.[264] 아이아이과를 여우원숭이하목 영장류(여우원숭이와 로리스로 분리되는 것보다 더 최근에 여우원숭이 계열에서 분리되는 것을 의미)의 일종으로 분류하거나 다른 모든 곡비원류의 자매군의 일종으로 분류하는 이론이 제안되었다. 2008년에, 아이아이과는 여우원숭이하목이 되어야 하고, 뗏목을 타고 섬으로 건너 온 같은 선조 여우원숭이 개체군의 자손임이 확인되었으며, 아이아이과는 결과적으로 자신의 이름을 딴 아이아이하목에서 나와서, 여우원숭이하목에 속하게 되었다.[282]
직비원아목은 2개의 자매 분류군으로 이루어져 있다.[264] 안경원숭이과(자신의 이름을 딴 안경원숭이하목에 속하는 단계통군)에 속하는 원원류 안경원숭이는 약 5800만 년 전에 가장 원시적인 부류를 대표한다.[290][291]
원숭이하목은 약 4000만 년 전에 등장했으며,[284] 2개의 분류군을 포함하고 있다: 남아메리카에서 발달했고 신세계원숭이를 포함하고 있는 광비원소목과 아프리카에서 발달하였고 구세계원숭이와 사람 등의 유인원을 포함하고 있는 협비원소목.[264] 세 번째 분류군, 에오시미드는 아시아에서 발달했지만 수백만 년 전에 멸종되었다.[292]
여우원숭이들처럼, 신세계원숭이도 기원이 명확하지 않다. 연쇄 상의 서열에 기반을 둔 분자적 서열 연구에서는 광비원류와 협비원류가 서로 분화된 때의 범위가 3300~7000만 년 전으로 넓은 변이를 보이는 것으로 추정되는 반면에 미토콘도리아 염기 서열에 기반을 둔 연구에서는 좀 더 일치하는 3500만 년 전임을 시사하고 있다.[293] 진원류는 아프리카에 단일한 기원을 두고 있고 일부가 이동했음을 자명하게 받아들이고 있으며, 표본도 잇따라 발견되었다.[289] 진원류 영장류는 에오세 동안에, 대서양의 융기된 능선과 해수면이 낮은 바다가 이들의 이동을 쉽게 했을 것이며, 섬들을 징검다리로 하여 대서양을 건너 남아메리카에 도착했을 것으로 추정된다.[289] 한번 더, 일종의 뗏목 사건을 통해 이와 같은 대륙간 이주를 설명할 수 있을 것이다. 대륙 이동설에 의하면 처음 만들어진 대서양은 절대로 오늘날과 같이 넓을 수 없으며,[289] 연구 결과는 1 kg의 작은 영장류가 식물로 된 뗏목에서 어떻게 13일을 생존했는지를 보여주고 있다.[294] 해류와 풍속을 감안할 때, 대륙간을 항해할 충분한 시간이 제공되었다.
유인원과 원숭이들은 마이오세가 시작될 무렵에 아프리카에서 유럽과 아시아로 퍼졌다.[295] 곧 이어, 로리스와 안경원숭이도 같은 여정을 떠났다. 가장 오래된 인류 화석은 500-800만년 전으로 거슬러 올라가며, 북아프리카에서 발견되었다.[284] 구세계원숭이들은 약 180만년 전에 유럽에서 사라졌다.[296] 일부 분자학적 연구와 화석 연구는 일반적으로 현세 인류가 10-20만년 전 아프리카에 기원을 두고 있다고 본다.[297]
영장류는 다른 동물군에 비해 연구가 잘 되고 있음에도 불구하고, 최근에도 몇몇 새로운 종들이 발견되고 있다. 2001년에 콜린 그로브스가 쓴 《영장류 분류(''Primate Taxonomy'')》는 약 350종을 수록하고 있으며,[274] 점점 증가하여 《세계의 포유류 종, ''Mammal Species of the World''》(MSW3)의 제3판에는 376종이 수록되어 있다.[264] 그러나, MSW3는 현재 최종 2003년판에 알려진 일부 종들을 누락하고 있으며, 기록된 종의 수는 더 증가하여 424종을 기록하고 있고, 아종을 포함하면 658종에 육박하고 있다. 베마라하양털여우원숭이 (''Avahi cleesei'', 영국 배우이자 여우원숭이광인 존 클리세이의 이름을 땄다.)와 황금궁전티티원숭이를 포함하여 여러 새로 발견된 종들이 MSW3 목록에 누락되어 있다.
"영장류"라는 단어에서 '영(霊)'은 혼이나 유령이라는 한자 자체의 의미보다는, 뛰어난 것, 신비한 힘을 가지고 있다는 의미가 강하다. 즉, 이는 사람이나, 사람을 포함한 원숭이의 무리를 동물의 진화의 최종 형태라고 인식하여 붙여진 이름이다. 영어 이름인 Primate영어 역시 대주교나 최고위를 의미하는 단어이며, 역시 비슷한 관점에서 붙여진 이름이다.
이전에는 주로 뇌가 작고 후각이 발달했으며 코가 긴 여우원숭이류, 로리스류, 안경원숭이류를 원원아목으로, 그 외 주로 뇌가 크고 시각이 발달했으며 코가 짧은 분류군을 진원아목으로 묶었다.[239] 연구가 진행됨에 따라, 안경원숭이가 이른바 원원류의 다른 그룹보다 진원류에 더 가깝다는 것이 밝혀졌다. 이로 인해 현재는 여우원숭이류, 로리스류를 묶어 "'''굽은코원숭이류'''(굽은코원숭이아목, 굽은코류, 굽은코아목)", 안경원숭이류를 포함한 그 외 영장류를 "'''곧은코원숭이류'''(곧은코원숭이아목, 곧은코류, 곧은코아목)"라고 부르며, 정식 분류 체계에서는 "원원류"라는 명칭은 사용하지 않는다.[250]
이하의 분류, 한국어 명칭은, 일본 몽키 센터 영장류 한국어 명칭 편찬 워킹 그룹 (2018)을 따른다.[240] 단, 대부분의 한국어 명칭은 공통되지만, 하목의 한국어 명칭에 관해서는 가와다 등 (2018)을 따랐다.[241]
{| class="wikitable"
|-
! 직비원아목
|-
|
{| class="wikitable"
|-
! 안경원숭이하목
|
| 안경원숭이과 |
|-
! 진원하목
|
{| class="wikitable"
|-
! 협비소목
|
{| class="wikitable"
|-
! 긴꼬리원숭이상과
|
| 긴꼬리원숭이과 |
|-
! 사람상과
|
| 사람과 |
| 긴팔원숭이과 |
|}
|-
! 광비소목
|
{| class="wikitable"
|-
! 꼬리감는원숭이상과
|
|-
! 사키상과
|
| 사키과 |
|}
|}
|}
|-
! 굽은코원아목
|-
|
{| class="wikitable"
|-
! 여우원숭이하목
|
{| class="wikitable"
|-
! 여우원숭이상과
|
|-
! 아이아이상과
|
|}
|-
! 로리스하목
|
| 갈라고과 |
| 로리스과 |
|}
|}
|}
5. 해부학, 생리학 및 형태학
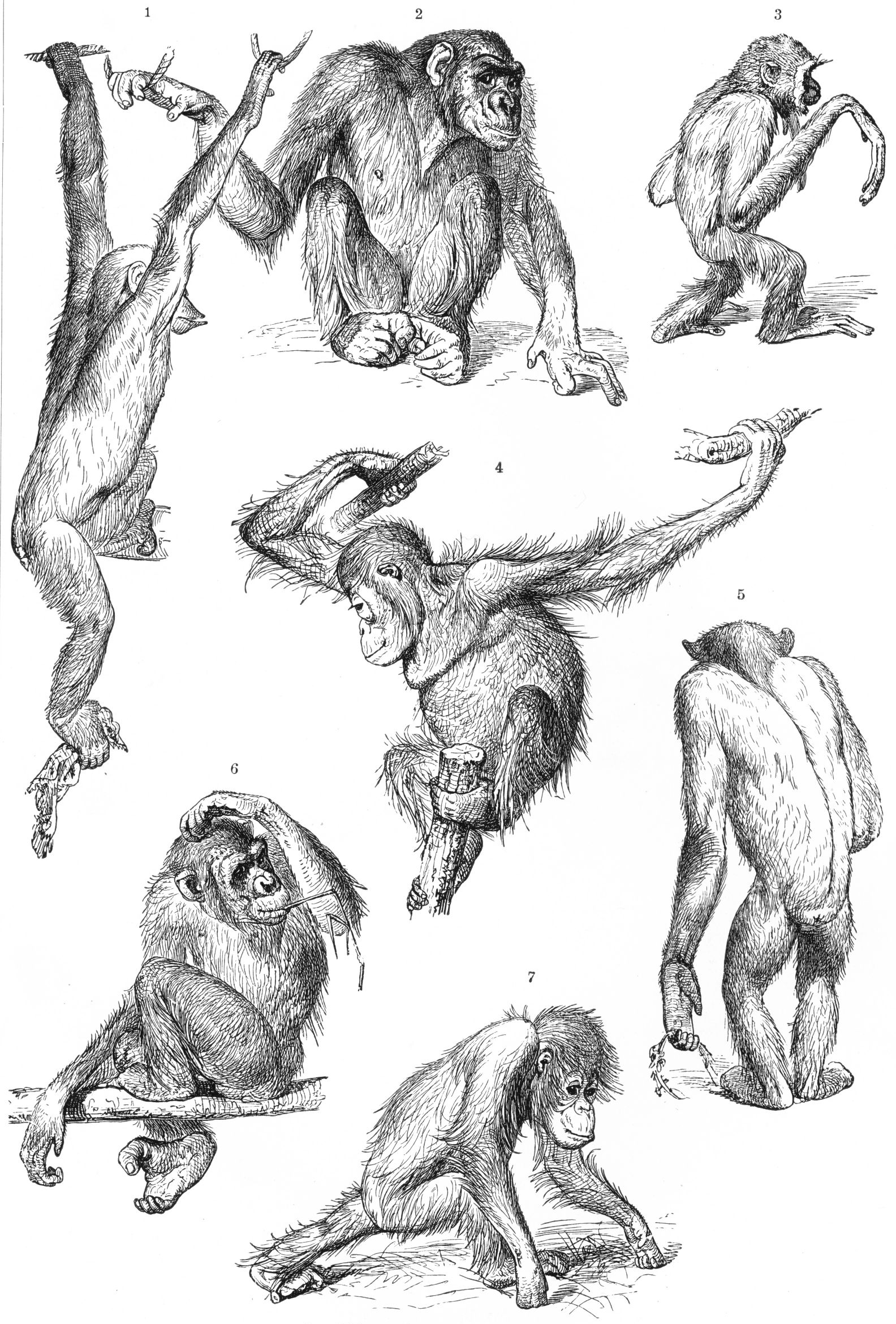
영장류는 둥글고 큰 두개골을 가지고 있으며, 이는 특히 사람을 포함한 원숭이아목에서 두드러진다.[72] 뇌용적(두개골 내부 용적)은 뇌 크기를 반영하며, 사람은 가장 큰 비인간 영장류보다 뇌 크기가 3배 더 크다.[65] 영장류 진화의 주요 방향은 두뇌, 특히 감각 인지, 운동 명령 생성, 공간 추론, 의식적 사고, 그리고 사람의 언어와 관련 있는 신피질의 발달이다.[224] 다른 포유류가 후각에 크게 의존하는 반면, 영장류의 수상 생활은 촉각과 시각이 지배적인 감각 시스템으로 이끌었으며, 뇌의 후각 영역 축소 및 복잡한 사회적 행동으로 이어졌다.[66]
영장류는 두개골 앞쪽에 정면을 향한 눈을 가지고 있어 양안시를 통해 정확한 거리 인식이 가능하다.[72] 눈 구멍 위의 눈썹뼈는 씹는 동안 스트레스를 받는 얼굴의 약한 뼈를 강화한다. 곡비원류는 눈을 보호하기 위해 눈 구멍 주위에 후안와봉(postorbital bar)을 가지고 있는 반면, 상위 영장류인 직비원류는 완전히 닫힌 구멍으로 진화했다.[69]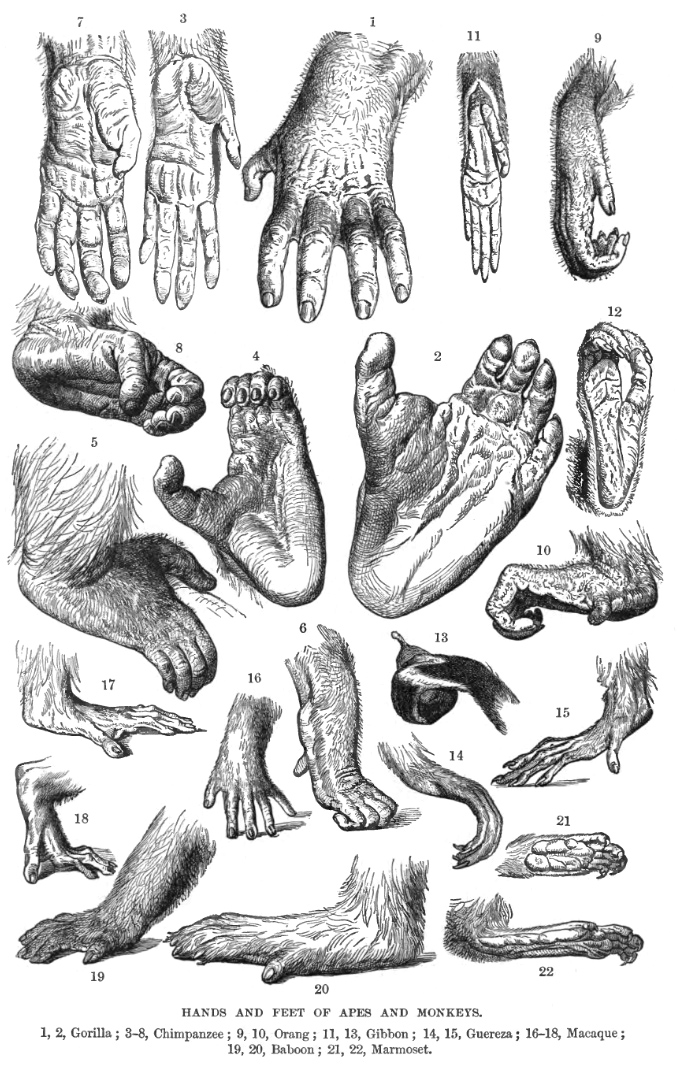
영장류는 일반적으로 각 사지에 다섯 개의 손가락을 가지고 있으며, 각 손가락과 발가락 끝에 손톱이 있다. 손과 발의 밑면에는 민감한 패드가 손가락 끝에 있다. 대부분은 엄지손가락이 서로 마주보는 특징을 가지고 있는데, 이는 영장류의 특징이지만, 사람에게 가장 잘 발달되어 있으며, 주머니쥐와 코알라 등도 엄지손가락을 가지고 있다.[72] 원원류는 각 발의 두 번째 발가락에 털을 다듬는 데 사용되는 화장실 발톱을 가지고 있다.[72]
영장류의 쇄골은 견갑골의 중요한 요소로, 견관절의 넓은 이동성을 허용한다.[70] 유인원은 구세계원숭이에 비해 더 움직이는 어깨 관절과 팔을 가지고 있다.[6] 뭔가를 쥘 수 있는 꼬리는 신세계 거미원숭이과와 카푸친원숭이에서 발견된다.[73][74] 수컷 영장류는 아래로 처진 음경과 음낭으로 내려온 고환을 가지고 있다.[75][76]
영장류는 주둥이가 감소하는 방향으로 진화적 경향을 보인다.[70] 신세계원숭이와 구세계원숭이는 코의 구조와 치열 배열에 의해 구별된다.[66] 신세계 원숭이의 콧구멍은 옆을 향하고, 구세계 원숭이의 콧구멍은 아래를 향한다.[66] 영장류의 치아 패턴은 다양하며, 일부는 대부분의 절치를 잃었지만, 모두 적어도 하나의 하악 절치를 유지한다.[66] 대부분의 협비원류에서 하악 절치는 치아 빗을 형성하며, 이는 털을 다듬거나 먹이를 찾는 데 사용된다.[66][76] 구세계 원숭이는 신세계 원숭이의 12개에 비해 8개의 소구치를 가지고 있다. 구세계 종은 치관의 수에 따라 유인원과 원숭이로 나뉜다. 원숭이는 4개, 유인원은 5개를 가지고 있다.[66]
5. 1. 특징
영장목은 수상(樹上) 생활에 적응하면서 다양한 특징을 가지게 되었다.[310] 주요 특징은 다음과 같다.- 흉곽 안에 쇄골이 있다.[310]
- 어깨 관절은 모든 방향으로 큰 각도를 움직일 수 있다.[310]
- 엄지손가락과 나머지 손가락이 서로 마주 보게 할 수 있으며, 앞뒤 좌우 손과 발에 각각 5개씩의 손발가락을 갖고 있다.[310]
- 대부분의 종이 손톱과 발톱이 있고, 현존하는 모든 종들은 엄지 손발가락의 손발톱이 평평하다.[308]
- 손가락 끝단에 민감한 촉각을 지닌 피부가 있다.[310]
- 눈구멍이 뼈로 둘러싸여 있다.[309]
- 주둥이가 짧고 얼굴이 평평한 편이며, 시각에 비해 후각은 떨어진다(대부분의 직비원류가 그렇고, 곡비원류는 덜한 편이다).[309]
- 입체시가 가능하고 높은 시각과 색각을 갖춘 복잡한 시각 인식 체계를 갖고 있다.[310]
- 뇌는 잘 발달된 후엽소뇌와 새발톱고랑(조거구)를 갖고 있다.[309]
- 몸 크기에 비해 상대적으로 큰 뇌를 갖고 있으며, 특히 유인원은 큰 뇌를 갖고 있다.[310]
- 대뇌 피질이 더 크게 분화되어 있다.[310]
- 원시 포유류보다 치아 개수가 상대적으로 적다.[310]
- 치아가 3종류로 나뉜다.[309]
- 맹장이 잘 발달되어 있다.[309]
- 2개의 유선(乳腺)이 있다.[310]
- 보통 한 번에 한 마리의 새끼를 밴다.[310]
- 수컷 생식기와 고환이 흔들거린다.[309]
- 임신 기간과 발생 기간이 길다.[310]
- 몸통을 똑바로 유지하려고 하며, 이것이 두 발 보행을 하게 만든다.[310]
모든 영장류가 이러한 특징을 모두 가지고 있는 것은 아니며, 일부 특징은 다른 포유류에도 존재한다.[309] 예를 들어 거미원숭이는 엄지손가락이 매우 작고, 목도리여우원숭이는 6개의 유선을 가지고 있다.[309]
영장류는 높은 수준의 사회적 행동을 보이며, 복잡한 위계 체계를 갖는 경우가 많다.[311] 신세계원숭이는 일부일처제를 보이는 경우가 많고, 수컷이 새끼를 돌보기도 한다.[312]
양안시는 거리를 정확하게 지각하는 데 도움을 주며, 이는 나무 위에서 생활하는 데 유리하다.[310] 곡비원류는 눈을 보호하기 위해 안와 둘레에 후안와봉을 가지고 있으며, 직비원류는 완전히 발달된 안와를 가지고 있다.[313]
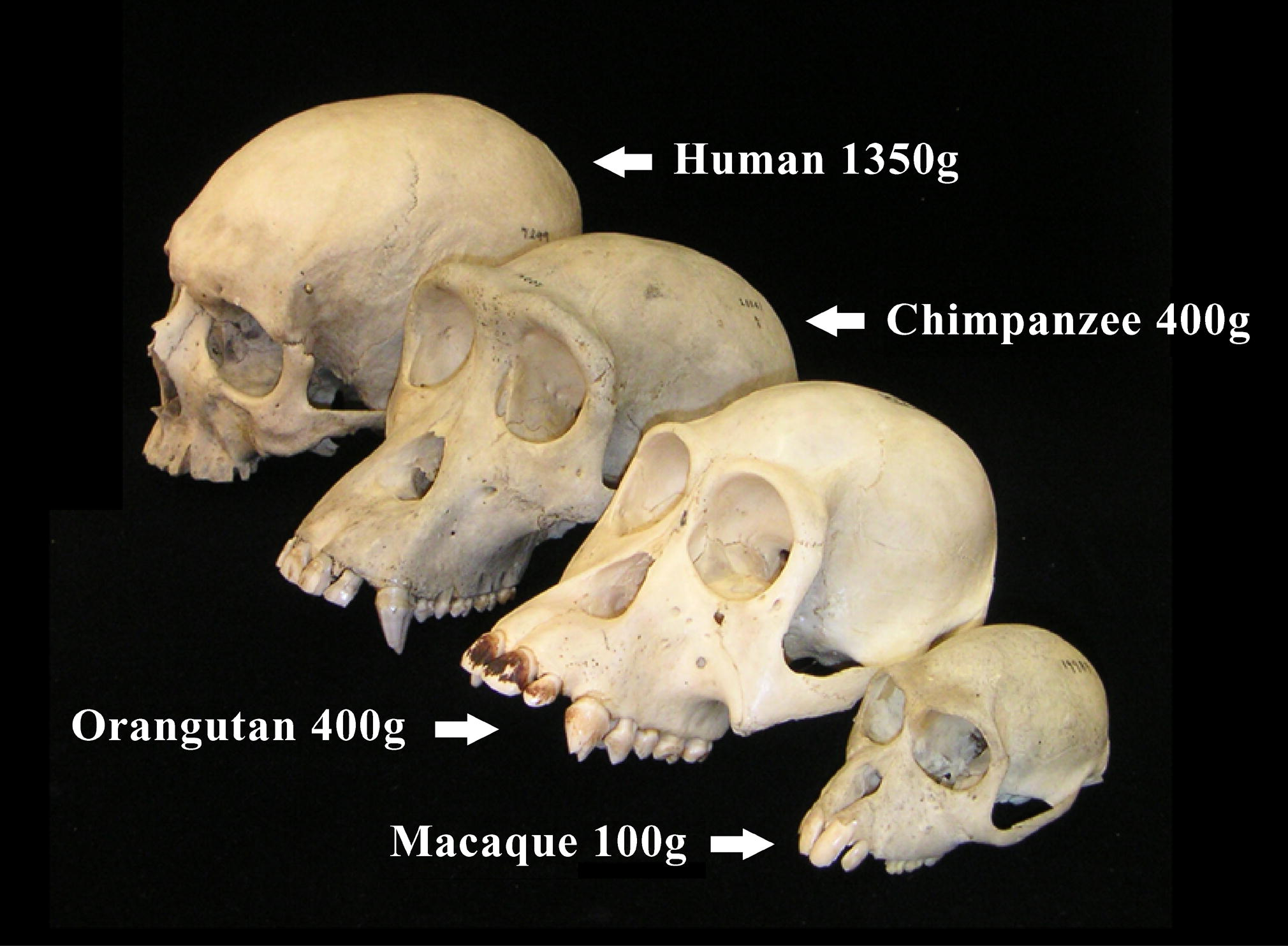
영장류 두개골은 둥글고 큰 덮개를 가지고 있으며, 특히 사람을 포함한 인류에게 두드러진다.[310] 뇌용적(두개골 내 부피)은 뇌 크기를 반영하며, 사람이 다른 영장류보다 세 배나 크다.[314] 영장류 진화의 주요 방향은 두뇌, 특히 신피질의 발달에 있으며, 이는 감각 인지, 운동 명령 생성, 공간 추론, 의식적 사고, 그리고 사람의 언어와 관련이 있다.[315]
영장류는 각 손발에 5개의 손발가락을 가지고 있으며, 손발톱이 있다. 손발 바닥 면에는 민감한 피부가 있다. 대부분 마주보게 할 수 있는 손가락을 가지고 있으나, 주머니쥐와 같은 다른 동물들도 이 특징을 가지고 있다.[310] 원원류는 각 발의 두 번째 발가락에 털을 손질할 때 사용하는 "몸단장용-발톱"을 가지고 있다.[310]
영장류의 쇄골은 팔이음뼈의 중요한 요소로, 어깨 관절의 운동성을 넓게 만든다.[311] 뭔가를 쥘 수 있는 꼬리를 지닌 영장류는 신세계원숭이인 거미원숭이과 원숭이뿐이다.
영장류는 진화하면서 주둥이가 줄어드는 경향을 보인다.[311] 신세계원숭이와 구세계원숭이는 코 구조와 치열에서 차이를 보인다.[316]
영장류의 색각 진화는 진수하강 포유류 중에서 독특하다. 야행성 포유류 조상은 망막에서 3개의 추체 중 하나를 상실했지만, 협비원류는 적록 옵신 유전자가 복제되어 3색성 색각을 가지게 되었다.[309][319] 광비원류는 일부만이 3색성 색각을 가지며, 이는 X 염색체의 대립 유전자와 관련이 있다.[309][320] 고함원숭이는 3색성 색각을 통해 잎을 구별하는 데 유리하다.[323]

진원류에서는 성별 간 차이인 성적 이형성이 나타나며, 몸무게, 송곳니 크기, 털과 피부 색깔 등에서 차이를 보인다.[324][325][326][327][328]

영장류는 팔로 매달려 건너다니기, 두발보행, 뛰어오르기 등 다양한 이동 방식을 사용한다. 사람은 현존하는 유일한 두발보행 영장류이다.
영장류는 진보된 인지 능력을 가지고 있으며, 일부는 도구를 사용하고 지능적인 사냥 전략을 구사한다.[346][347][348] 또한 사회적 지능, 기호 사용 능력, 언어의 일부 양상을 이해하는 능력을 보여준다.[349][350][351][352][353][354]
여우원숭이, 로리스, 안경원숭이, 신세계원숭이는 후각 신호에 의존하는 반면, 구세계원숭이와 유인원은 시각이 주 감각 기관으로 진화했다.[315] 영장류는 발성, 몸짓, 얼굴 표현을 통해 의사소통을 한다.[357]
영장류는 다른 포유류보다 발달이 늦고, 새끼는 어미의 보살핌을 받는다.[296] 또한 비슷한 크기의 다른 포유류보다 수명이 길다.[296]


영장류는 과일, 잎, 곤충 등 다양한 먹이를 섭취하며, 일부는 특이한 음식을 먹기 위해 특화된 기관을 가지고 있다.[296] 겔라다는 풀을 주식으로 하는 유일한 영장류이며, 안경원숭이는 육식만 하는 유일한 영장류이다.[360][361]

영장류는 대부분 열대 우림에 서식하며, 일부는 온대 기후 지역에도 서식한다. 사람은 추운 지방에서도 살고 있다.
5. 2. 성적 이형성
같은 종에서 다른 성별의 개체 사이의 차이인 성적 이형성(동종이형)은 진원류에서 흔히 나타나며, 신세계 종들보다는 구세계 종들(유인원과 일부 원숭이) 사이에서 그 차이가 더 큰 것으로 보인다.[324][325][326][327][328]
최근 연구에서는 영장류에서 나타나는 이형성의 변이와 성적 이형성의 근본 원인을 조사하기 위해 DNA 비교 방법을 사용하고 있다. 영장류는 흔히 몸무게와 송곳니의 크기, 털과 피부의 색깔에서 성적 이형성을 보인다. 이러한 성적 이형성은 짝짓기 방식,[329] 크기,[329] 서식 환경과 먹이를 포함한 다양한 요인에 의해 영향을 받거나 영향을 끼친다.[330]
비교 분석을 통해 영장류의 성 결정, 자연 선택, 짝짓기 방식 사이의 관계를 더 완벽하게 이해할 수 있게 되었다. 연구에 따르면 성적 이형성은 암수의 특징 변화를 유발한다.[331] 일반적인 성장 과정에서 나타나는 비례적인 신장, 즉 발생적 크기 조정은 성적 이형성과 성장 형태 사이의 관계에 대한 통찰력을 제공한다.[332] 화석 기록의 일부 증거는 성적 이형성의 수렴 진화를 시사하며, 일부 멸종된 인류는 현존하는 영장류보다 성적 이형성이 더 큰 것으로 보인다.[331]
5. 3. 이동
영장류 종은 팔로 매달리며 건너다니기, 두발 보행, 뛰어오르기, 나무 위와 지상에서의 네발 보행, 기어오르기, 앞다리의 지관절(指關節)의 등을 땅에 대고 걷기를 통해 이동하거나 이 방법들을 조합하여 이동한다.[333] 몇몇 원원류는 주로 수직으로 매달리거나 뛰어 오르기를 한다. 이들에는 부시베이비의 다수와 모든 인드리(예를 들어, 시파카, 아바히 그리고 인드리), 족제비여우원숭이, 그리고 모든 안경원숭이가 포함된다.[333]나머지 진원류는 나무 위에서 네발을 사용해 움직이거나 나무를 타고 오른다. 일부는 또한 지상에서 네발로 걷거나 일부는 뛰어다닌다. 대부분의 원숭이는 나무 위에서 생활하거나 지상에서 네발로 걷거나 기어오르기 둘 다를 한다. 긴팔원숭이, 무리키 그리고 거미원숭이들 모두 "팔로 매달려 건너다니기"를 광범위하게 사용한다.[296] 양털원숭이들 또한 종종 두 팔로 매달려 건너다닌다.[322] 오랑우탄은 일종의 "사지형 기어오르기"라고 불리는 비슷한 방식으로 이동하는데, 이 방식은 나무들 속에서 자신의 무거운 몸을 이동시킬 때, 자신의 팔과 다리를 사용하는 것이다.[296]
침팬지와 고릴라는 앞다리의 지관절(指關節)의 등을 땅에 대고 걸으며,[296] 짧은 거리는 두발로 걸을 수 있다. 오스트랄로피테신과 초기 인류와 같은 수많은 종들이 완전히 두발로 이동했음이 밝혀져 있지만, 사람만이 이러한 특징을 갖는 현존하는 유일한 종이다.
5. 4. 색각
영장류는 수상(樹上) 생활에 적응하면서 다양한 특징을 갖게 되었는데, 그 중 하나가 발달된 시각 인식 체계이다.[310] 특히 색각은 영장류의 생존과 번식에 중요한 역할을 담당한다.영장류의 먼 척추동물 조상은 3색성 색각을 보유하고 있었지만, 야행성이었던 포유류 조상은 중생대 기간 동안 망막에서 3개의 추체 중 하나를 상실하여 2색성 색각 또는 단색성 색각을 갖게 되었다.[309] 그러나 협비원류(구세계원숭이와 유인원)는 약 3~4천만 년 전, 적록 옵신 유전자가 유전자 복제되면서 3색성 색각을 다시 획득하게 되었다.[309][319]
반면, 광비원류(신세계원숭이)는 일부만이 3색성 색각을 가지고 있다.[320] 암컷은 X 염색체의 같은 자리에 위치하는 옵신 유전자(적록)의 2개의 대립 유전자가 이형이어야 3색성 색각을 가질 수 있다.[309] 수컷은 2색성 색각만 가능하지만, 암컷은 2색성 또는 3색성 색각이 모두 가능하다. 곡비원류는 색채 식별 능력이 없지만, 광비원류와 비슷한 범위의 색각을 가지고 있다는 연구 결과도 있다.[309]
고함원숭이(광비원류의 일종)는 유전자 복제를 통해 전형적인 3색성 색각을 획득했다.[321] 이들은 잎을 주식으로 하는데, 어리고 영양가 높은 잎을 구별하는 데 3색성 색각이 유리하게 작용한다.[323]
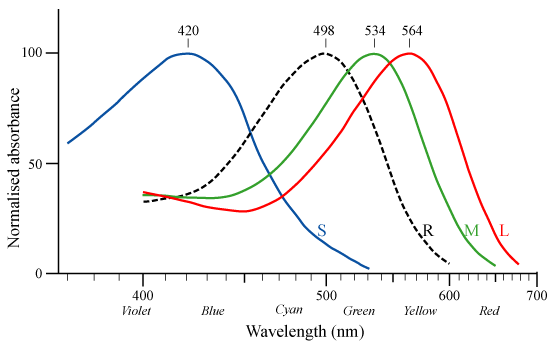
척추동물의 색각은 망막 안에 어떤 타입의 원추 세포를 가지고 있는지에 따라 결정된다. 어류, 양서류, 파충류, 조류는 4가지 타입의 원추 세포(4색형 색각)를 가진 것이 많다. 따라서 이러한 생물은 장파장역에서 단파장역인 근자외선까지 인식할 수 있다고 생각된다. 한편 대부분의 포유류는 원추 세포를 2가지 타입(2색형 색각)밖에 갖지 않는다. 파충류의 조상에서 갈라져 나온 포유류의 조상은 처음에는 4가지 타입의 모든 원추 세포를 가지고 있었을 것으로 생각되지만, 2억 2,500만 년 전에는 최초의 진정한 포유류라고 불리는 아데로바실레우스가 출현했다. 이러한 초기의 포유류는 (공룡 등의 파충류와의 경쟁을 피했기 때문에) 주로 야행성이었기 때문에, 색각은 생존에 필수적이지 않았다. 그 결과, 4가지 타입 중 2가지 타입의 원추 세포를 잃고, 파랑을 중심으로 감지하는 S원추체와 빨강을 중심으로 감지하는 L원추체 2원추체만을 보유하게 되었다. 이것은 적색과 녹색을 충분히 구별할 수 없는 소위 "적록색맹" 상태이다. 이 색각이 포유류의 자손에게 유전적으로 계승되게 되었다.[256]
인간을 포함한 구세계 영장류(협비원류)의 조상은 약 3,000만 년 전, X 염색체에 L원추체에서 변이된 녹색을 중심으로 감지하는 새로운 타입의 원추체(M원추체) 시물질의 유전자가 출현하여, 이형 접합체의 2개의 X 염색체를 가진 암컷만이 3색형 색각을 갖게 되었으며, 더 나아가 이형 접합체의 암컷에서 상동 재조합에 의한 유전자 중복의 변이를 일으켜 동일한 X 염색체상에 2가지 타입의 원추체 시물질 유전자가 보존되게 되어 X 염색체를 1개밖에 갖지 않는 수컷도 3색형 색각을 갖게 되었다. 이것에 의해, 제3의 원추 세포가 "재생"되었다. 3색형 색각은 비타민 C가 풍부한 색깔이 선명한 과실 등의 발견에 유리했을 것으로 생각된다.[254][256]
협비원류의 마카크류에 색맹이 인간보다 매우 적다는 것을 고려하면, 인간의 조상이 수렵 생활을 하게 되면서 3색형 색각의 우위성이 낮아지고, 2색형 색각의 도태압이 낮아졌다고 생각된다.[256] 색맹의 출현 빈도는 협비원류의 게잡이원숭이에서 0.4%, 침팬지에서 1.7%이다.[254] 광비원류에서도 밤원숭이는 1색형 색각이며, 고함원숭이는 협비원류와 마찬가지로 3색형 색각을 재획득하고 있다.[254][257] 한편, 고함원숭이는 획일적인 3색형 색각이 아니라, 고도한 색각 다형성이라는 지적도 있다.[258] 이러한 밤원숭이, 고함원숭이를 제외한 나머지 신세계 원숭이(광비하목)는 이형 접합체의 X 염색체를 2개 가진 암컷만이 3색형 색각을 가지며, 수컷은 모두 색맹이다. 이것은 협비하목과 같은 X 염색체상에서의 상동 재조합에 의한 유전자 중복의 변이를 일으키지 않았기 때문이다.[254] 인간은 위와 같은 초기 포유류와 영장목 협비하목의 조상의 X 염색체의 유전자 변이를 계승하고 있기 때문에, L원추체만을 보존한 X 염색체와 관련된 적록색맹이 연관 열성 유전을 한다. 남성에서는 X 염색체의 적록색맹 유전자를 계승하고 있으면 색맹이 발현되고, 여성에서는 2개의 X 염색체 모두 적록색맹 유전자를 계승하고 있는 경우에 적록색맹이 발현된다.[259]
6. 생태
영장류는 수상(樹上) 생활 환경에 적응하면서 다양한 특징을 갖게 되었다.[310] 주요 특징은 다음과 같다.
- 흉곽 안에 쇄골이 있다.[310]
- 어깨 관절은 모든 방향으로 크게 움직일 수 있다.[310]
- 엄지손가락과 나머지 손가락이 서로 마주 보게 할 수 있으며, 손과 발에는 각각 5개씩의 손발가락이 있다.[310]
- 대부분의 종은 손톱과 발톱이 있다.[308]
- 현존하는 모든 종은 엄지손발가락의 손발톱이 평평하다.[308]
- 손가락 끝에는 민감한 촉각을 지닌 피부가 있다.[310]
- 눈구멍이 뼈로 둘러싸여 있다.[309]
- 주둥이가 짧고 얼굴이 평평한 편이며, 시각에 비해 후각은 떨어진다.(대부분의 직비원류가 그렇고, 곡비원류는 덜한 편이다).[309]
- 입체시가 가능하고 높은 시각과 색각을 갖춘 복잡한 시각 인식 체계를 갖고 있다.[310]
- 뇌는 잘 발달된 후엽소뇌와 새발톱고랑(조거구)를 갖고 있다.[309]
- 몸 크기에 비해 상대적으로 큰 뇌를 갖고 있으며, 유인원은 특히 뇌가 크다.[310]
- 대뇌 피질이 더 크게 분화되어 있다.[310]
- 원시 포유류보다 치아 개수가 상대적으로 적다.[310]
- 치아가 3종류로 나뉜다.[309]
- 맹장이 잘 발달되어 있다.[309]
- 2개의 유선(乳腺)이 있다.[310]
- 보통 한 번에 한 마리의 새끼를 밴다.[310]
- 수컷 생식기와 고환이 흔들거린다.[309]
- 임신 기간과 발생 기간이 길다.[310]
- 몸통을 똑바로 유지하려고 하며, 이것이 두 발 보행을 하게 만들었다.[310]
영장류 두개골은 둥글고 큰 덮개를 가지고 있으며, 특히 진원류에게 두드러진 특징이다. 두개골 덮개는 큰 뇌를 보호하는 역할을 한다.[310] 뇌용적(두개골 내의 부피)은 뇌 크기를 반영하며, 사람이 나머지 영장류들보다 세 배나 크다.[314] 영장류 진화의 주요 방향은 두뇌의 정교화에 있으며, 특히 감각 인지와 운동 명령의 생성, 공간 추론, 의식적인 사유, 그리고 사람의 언어와 관련이 있는 신피질(대뇌 피질의 일부분)에 있다.[315]
영장류는 원래 각 손발에 5개의 손발가락(五指)을 가지고 있으며, 각 손발가락의 끝에는 각질로 된 손발톱이 있다. 손발의 바닥 면은 손발의 끝에 민감한 피부를 지니고 있다.
영장류의 쇄골은 팔이음뼈의 두드러진 요소로써 유지되어 왔으며, 이는 어깨 관절의 운동성을 폭넓게 만든다.[311]
영장류는 진화적으로 주둥이가 줄어드는 경향을 보여주고 있다.[311]
영장류에 있어서 색각의 진화는 다수의 진수하강 포유류 중에서 독특하다. 영장류의 먼 척추동물 조상이 3색성 색각을 보유한 반면에, 야행성이자 온혈 동물인 포유류 조상은 중생대 기간 동안에 망막에서 3개의 추체 중의 하나를 상실했다.
같은 종에서 다른 성의 개체 사이의 차이인, 성적 이형성(동종이형)이 진원류에서 흔히 나타나며, 신세계원숭이보다는 구세계원숭이(유인원과 일부 원숭이) 사이에서 그 차이가 더 큰 것으로 보인다.
영장류 종은 팔로 매달리며 건너다니기, 두 발 보행, 뛰어오르기, 나무 위와 지상 에서의 네발보행, 기어오르기, 앞다리의 지관절(指關節)의 등을 땅에 대고 걷기를 통해 이동하거나 이 방법들을 조합하여 이동한다.
영장류는 진일보한 인지 능력을 지니고 있다. 일부는 도구를 만들어, 먹이를 구하고 사회적 표현을 하는 데 사용한다.[346][347]
영장류는 다른 포유류들보다 발달이 늦다.[296]
영장류는 여러 가지의 음식물을 섭취한다.
영장류의 포식자에는 다양한 종의 식육목, 맹금류, 파충류, 그리고 다른 영장류 등이 있다.
영장류의 공통 조상은 야행성이었을 것으로 생각되지만[248], 주행성이었을 것이라는 설도 있다[247]。
6. 1. 사회 체계
영장류의 사회 체계는 매우 다양하며, 크게 암컷 이동 시스템, 수컷 이동 시스템, 일부일처 종, 고립 종의 네 가지 범주로 나눌 수 있다.| 사회 체계 | 설명 | 해당 종 |
|---|---|---|
| 암컷 이동 시스템 | 암컷이 태어난 집단을 떠나고, 수컷은 남아있는 시스템 | 침팬지, 네안데르탈인, 오스트랄로피테쿠스, 파란트로푸스 로부스투스, 거미원숭이, 무리키 |
| 수컷 이동 시스템 | 수컷이 청소년기에 이주하고 암컷은 태어난 집단에 머무는 시스템. 일부다처제 및 다수 수컷 사회가 여기에 속함 | 고리꼬리원숭이, 카푸친원숭이, 구세계 원숭이 |
| 일부일처 종 | 수컷과 암컷이 짝을 이루고, 새끼와 함께 생활하는 시스템. 부모는 영토 방어와 양육을 분담 | 긴팔원숭이 |
| 고립 종 | 수컷이 여러 암컷의 활동 범위를 포함하는 영토를 방어하는 시스템 | 느린로리스, 오랑우탄 |
이 외에도 울음원숭이, 고릴라, 콜로부스원숭이, 비단원숭이 등은 수컷과 암컷 모두 태어난 집단을 떠나는 시스템을 가지고 있다.
영장류의 사회 시스템은 근친교배를 피하기 위한 적응으로 볼 수 있다. 연구에 따르면, 근친교배된 새끼의 유아 사망률은 그렇지 않은 경우보다 높다.[103][104]
일부 영장류는 분열융합 사회를 이루기도 한다. 제인 구달은 침팬지에게서 이러한 사회 구조를 발견했으며, 개코원숭이, 거미원숭이, 보노보, 겔라다원숭이 등도 분열융합 사회를 이룬다. 인간 역시 수렵 채집 사회에서 분열융합 사회를 형성한다.
영장류의 사회 집단 내에서는 협력과 경쟁이 균형을 이룬다. 사회적 그루밍, 음식 공유, 집단 방어 등은 협력적 행동의 예시이며, 공격적인 행동은 경쟁을 나타낸다. 2023년에는 보노보 집단이 서로 협력할 수 있다는 증거가 보고되기도 했다.[110][111]
6. 2. 이종간 군집
여러 영장류 종은 야생에서 서로 어울리는 것으로 알려져 있다. 이러한 연합 중 일부는 광범위하게 연구되었다. 아프리카의 타이 숲에서는 다이애나원숭이, 캠벨모나원숭이, 작은코원숭이, 서부붉은콜로부스, 킹콜로부스(서부 검은흰콜로부스), 매부리망가베이를 포함한 여러 종이 포식자 방어 행동을 조율하며, 이들은 포식자 경고 소리를 조율한다.[112] 이 원숭이들의 포식자 중에는 침팬지가 있다.[113]붉은꼬리원숭이는 서부붉은콜로부스, 푸른원숭이, 울프모나원숭이, 망토게레자, 검은크레스트망가베이, 앨런늪원숭이를 포함한 여러 종과 어울린다.[106] 이 종 중 일부는 침팬지의 먹이가 된다.[114]
남아메리카에서는 다람쥐원숭이가 카푸친원숭이와 어울린다.[115] 이는 포식자 방어 이점보다 다람쥐원숭이에게 식량 획득의 이점이 더 클 수 있다.[115]
6. 3. 인지 능력과 의사 소통
영장류는 진일보한 인지 능력을 지니고 있다. 일부는 도구를 만들어 먹이를 구하고 사회적 표현을 하는 데 사용한다.[346][347] 일부는 협력과 힘, 지위 등이 필요한 지능적인 사냥 전략을 구사한다.[348] 이들은 자신의 지위를 인지하고, 꾸미거나 속일 수 있는 능력이 있다.[349] 자신들과 같은 종인지 다른 종인지도 인식할 수 있으며,[350][351] 기호를 사용하는 법을 배우고, 일부 문법 규칙과 수와 순서의 개념을 포함한 인간 언어의 양상을 이해할 수 있다.[352][353][354] 영장류 인지에 대한 연구는 문제 해결, 기억, 사회적 상호작용, 일종의 마음 이론, 그리고 수리 및 공간 지각, 추상적 개념 등을 탐구한다.[355]여우원숭이, 로리스, 안경원숭이, 신세계원숭이들은 사회적, 생식 행동의 여러 국면에서 후각 신호에 의존한다.[315] 분화된 분비선인 페로몬을 자신의 영역을 표시하기 위해 사용하며, 서골비기관을 통해 탐지한다. 이 과정은 이들 영장류의 의사소통 행동 중에서 가장 큰 부분이다.[315] 구세계원숭이와 유인원들은 대부분 이 능력이 흔적으로만 남아 있고, 삼색형 색각의 시력이 주 감각 기관으로 진화했다.[356] 영장류는 또한 심리 상태를 전달하기 위해 발성, 몸짓, 얼굴 표현을 사용한다.[357]
6. 4. 생활사
영장류는 다른 포유류보다 발달 속도가 느리다.[52] 사람을 제외한 모든 영장류의 새끼들은 어미가 모유 수유를 하며, 돌보고 데리고 다닌다.[296][52] 일부 종에서는, 그 집단 내의 수컷들이 새끼들을 보호하며, 데리고 다니는데, 특히 그 수컷들은 그들의 아버지로 보인다.[296][52] 형제자매와 아주머니와 같은 새끼들의 기타 친척들 또한 이들을 보살필 수 있다.[296][52] 대부분의 영장류 어미들은 새끼들의 모유 수유 기간 동안에는 배란이 중단된다. 새끼들이 젖을 떼면 그 어미는 다시 생식을 할 수 있다.[296][52] 이는 젖을 계속 먹으려는 새끼와 젖떼기 갈등을 일으키는 원인이 되기도 한다.[296][52]영장류는 비슷한 크기의 다른 포유류에 비해, 젖을 떼고 성적으로 성숙해지는 기간이 길다.[296][52] 성적으로 성숙해지기 이전 기간 동안에, 영장류는 포식과 굶주림에 더 민감하다. 이 시기 동안에 이들은 먹이를 구하고 맹수를 피하는 방법을 경험을 통해 얻는다.[296][52] 또한 이들은 종종 놀이를 통해 군집 생활 및 싸움 기술을 배운다.[296][52]
영장류, 특히 암컷들은, 비슷한 크기의 다른 포유류보다 수명이 더 길다.[296][52]
6. 5. 먹이
영장류는 다양한 음식을 먹는다. 사람을 포함한 현대 영장류의 많은 특징들은 초기 조상들이 열대 우림의 나무 위에서 먹이를 구하던 습성에서 유래했다고 알려져 있다.[141] 대부분의 영장류는 에너지를 얻기 위해 탄수화물과 지질이 포함된 과일을 먹는다.[52] 곡비원류는 다른 포유류처럼 비타민 C를 만들 수 있지만, 직비원류는 이 능력을 잃어 식단을 통해 비타민 C를 섭취해야 한다.[142]많은 영장류는 과일, 잎, 수지(樹脂) 또는 곤충 등과 같은 특정 음식을 먹기 위해 특별한 신체 구조를 가지고 있다.[52] 예를 들어, 고함원숭이, 흑백콜로부스, 족제비여우원숭이는 소화하기 어려운 잎에서 영양분을 흡수하기 위해 소화관이 길다.[52] 수지를 먹는 마모셋은 나무껍질을 벗겨 수지를 꺼낼 수 있도록 튼튼한 앞니를 가지고 있으며, 손톱 대신 발톱이 있어 나무에 매달릴 수 있다.[52] 아이아이는 설치류와 같은 이빨과 길고 얇은 가운데 손가락을 가지고 있어 딱따구리와 비슷한 역할을 한다. 나무를 두드려 곤충 애벌레를 찾은 다음, 나무에 구멍을 내고 가운데 손가락을 넣어 애벌레를 꺼낸다.[143] 일부 종은 추가적인 특징을 가지고 있다. 예를 들어, 회색뺨망가베이는 다른 원숭이가 열 수 없는 단단한 과일과 씨앗을 열 수 있도록 이빨의 법랑질이 두껍다.[52] 겔라다는 주로 풀을 먹는 유일한 영장류 종이다.[144]

안경원숭이는 유일하게 육식만 하는 영장류로, 곤충, 갑각류, 작은 척추동물, 뱀(독사 포함)을 먹는다.[145] 카푸친원숭이는 과일, 잎, 꽃, 꽃봉오리, 꿀, 씨앗 등 다양한 식물과 곤충, 새 알, 새, 도마뱀, 다람쥐, 박쥐와 같은 작은 척추동물도 먹는다.[92]
침팬지는 다양한 음식을 먹는 잡식성 동물이다. 과일을 가장 좋아하며, 과일이 없을 때에도 찾아서 먹는다. 잎, 잎눈, 씨앗, 꽃, 줄기, 나무껍질, 수지도 먹는다. 곤충과 고기는 식단의 약 2%를 차지한다.[146][147] 서부붉은콜로부스와 같은 다른 영장류를 잡아먹기도 한다.[113] 보노보는 잡식성으로 과일을 주로 먹지만, 잎, 아노말루어, 하늘다람쥐, 듀이커와 같은 작은 척추동물, 무척추동물도 먹는다.[148][149] 때로는 하위 영장류를 섭취하기도 한다.[150][151]

약 10,000년 전 농업이 발달하기 전까지, 호모 사피엔스는 수렵 채집 방식으로 식량을 얻었다. 과일, 곡물, 덩이줄기, 버섯, 곤충 애벌레, 수생 연체동물과 같은 고정된 식량원과 사냥해야 하는 야생 동물을 함께 섭취했다.[152] 인류는 호모 에렉투스 시대부터 음식을 준비하고 요리하기 위해 불을 사용한 것으로 추정된다.[153] 약 1만 년 전에 농업을 발전시키면서[154] 식단이 크게 변화했다. 이러한 식단의 변화는 인간의 생물학을 변화시켰을 수 있다. 낙농업의 확산은 새롭고 풍부한 식량을 제공하여 일부 성인에게 유당을 소화하는 능력이 진화하게 되었다.[155][156]
7. 분포
영장류는 주로 아프리카, 아시아, 아메리카 대륙의 열대 위도에서 서식한다. 열대 지역 외에 사는 종으로는 일본 혼슈와 홋카이도의 일본원숭이, 북아프리카의 마카카원숭이, 중국의 여러 랑구르 등이 있다.[186] 동아시아에는 온대 지역까지 분포하는 종이 있으며, 특히 일본원숭이는 가장 높은 위도에 분포하는 원숭이로 알려져 있다. 곡비원아목 및 안경원숭이류는 아시아·아프리카의 열대 지역, 광비원류는 중남미의 열대, 유인원을 포함하는 협비원류는 아시아·아프리카의 열대 지역에서 온대 지역 일부에 걸쳐 분포하고 있다. 유럽에는 거의 서식하지 않으며, 지브롤터 해협 건너 바바리원숭이 한 종만이 서식한다. 또한, 북아메리카 대륙 북부 (앵글로아메리카 상당 지역)에도 분포하지 않는다.
영장류는 열대 및 아열대 습윤 활엽수림에 서식하는 경향이 있지만, 온대림, 사바나, 사막, 산, 해안 지역에서도 발견된다.[186] 열대 지역의 영장류 종 수는 강수량과 열대 우림 면적과 상관 관계가 있는 것으로 나타났다.[187] 열대 우림에서 과식자의 25%~40%를 차지하는 영장류는 많은 나무 종의 씨앗을 퍼뜨림으로써 중요한 생태학적 역할을 한다.[188]
영장류의 서식지는 다양한 고도에 걸쳐 있다. 검은코원숭이는 헝돤산맥에서 4700m 고도에서 발견되었고,[189] 산악고릴라는 비룽가산맥을 넘어 4200m에서 발견될 수 있으며,[190] 겔라다는 에티오피아 고원에서 5000m까지의 고도에서 발견되었다.[191] 코주부원숭이, 드브라자원숭이, 앨런늪원숭이 등 일부 종은 수생 환경과 상호 작용하며 수영하거나 잠수하기도 한다.[192] 인도원숭이와 회색랑구르와 같은 일부 영장류는 인간이 개조한 환경을 이용할 수 있으며 심지어 도시에 살기도 한다.[106][193]
8. 인간과의 상호 작용
영장류 잡종은 사육 상태뿐만 아니라 야생에서도 발생한다.[304][305][306] 두 종의 활동 범위가 겹치는 잡종 지역에서 잡종이 생겨나며, 동물원과 같이 인간에 의해 만들어지거나 약탈과 같은 환경적 압박 때문에 발생하기도 한다.[305] 다른 속에 속하는 영장류 간의 잡종(속간 교잡)도 야생에서 발견되는데, 이는 겔라다속과 망토개코원숭이 사이에서만 발생한다.[307]
8. 1. 질병 전파
사람과 비인간 영장류(NHP) 사이의 밀접한 상호작용은 인수공통감염병의 전파 경로를 만들 수 있다. 헤르페스바이러스과(특히 B 바이러스), 폭스바이러스과, 홍역, 에볼라, 광견병, 마버그 바이러스, 바이러스성 간염과 같은 바이러스는 사람에게 전파될 수 있다. 어떤 경우에는 이 바이러스가 사람과 비인간 영장류 모두에게 잠재적으로 치명적인 질병을 일으킨다.[194]8. 2. 법적, 사회적 상태
영장류 잡종은 사육 상태에서 종종 생기지만,[304] 야생에서 생기기도 한다.[305][306] 잡종은 2개 종의 활동 범위가 잡종 지역을 형성하여 겹칠 때 생긴다. 잡종은 동물들이 동물원 등에 있을 때 인간에 의해 만들어지거나 약탈과 같은 환경적 압박때문에 만들어질 수 있다.[305] 속간 교잡 즉, 다른 속 사이의 잡종 또한 야생에서 발견되기도 한다. 비록 그들이 몇 백만 년 동안 구별되는 속에 속해 있지만, 이종 교잡은 단지 겔라다속과 망토개코원숭이 사이에서만 발생한다.[307]
오직 인간만이 법적으로 인격으로 인정받으며 국제 연합 세계 인권 선언에 의해 보호받는다.[195] 반면, 비인간 영장류(NHP)의 법적 지위는 많은 논쟁의 대상이며, 유인원 프로젝트(GAP)와 같은 단체는 최소한 일부에게 법적 권리를 부여하기 위해 캠페인을 벌이고 있다.[196] 2008년 6월, 스페인은 의회 초당파 환경위원회가 GAP의 권고안, 즉 침팬지, 오랑우탄 및 고릴라를 동물 실험에 사용하지 말 것을 촉구하면서, 세계 최초로 일부 NHP의 권리를 인정한 국가가 되었다.[197][198]
많은 종의 NHP가 인간에 의해 애완동물로 길러진다. 다른 영장류를 구하기 위한 연합(AESOP)은 약 15,000마리의 NHP가 미국에서 이국적인 애완동물로 살고 있다고 추산한다.[199] 중국 중산층의 증가로 최근 몇 년 동안 이국적인 애완동물로서의 NHP에 대한 수요가 증가했다.[200] 1975년 미국에서 애완동물 거래를 위한 NHP 수입이 금지되었지만, 여전히 미국-멕시코 국경을 따라 밀수가 발생하고 있으며, 원숭이는 3000USD에서 유인원은 30000USD까지 가격이 형성되어 있다.[201]
영장류는 실험실에서 모델 생물로 사용되며 우주 임무에도 사용되었다.[202] 그들은 장애가 있는 인간을 위한 보조 동물 역할을 한다. 카푸친원숭이는 사지마비 환자를 돕도록 훈련될 수 있다. 이들의 지능, 기억력 및 손재주는 그들을 이상적인 조력자로 만든다.[203]
NHP는 전 세계의 동물원에 보관된다. 역사적으로 동물원은 주로 오락의 한 형태였지만, 최근에는 보존, 교육 및 연구에 초점을 맞추고 있다. GAP는 모든 NHP를 동물원에서 풀어줘야 한다고 주장하지 않는데, 이는 주로 사육된 영장류가 야생으로 풀려났을 때 생존하는 데 필요한 지식과 경험이 부족하기 때문이다.[204]
8. 3. 애완동물
많은 종의 비인간 영장류(NHP)가 인간에 의해 애완동물로 길러진다. 다른 영장류를 구하기 위한 연합(AESOP)은 약 15,000마리의 NHP가 미국에서 이국적인 애완동물로 살고 있다고 추산한다.[199] 1975년 미국에서 애완동물 거래를 위한 NHP 수입이 금지되었지만, 여전히 미국-멕시코 국경을 따라 밀수가 발생하고 있으며, 원숭이는 3000USD에서 유인원은 30000USD까지 가격이 형성되어 있다.[201] 중국 중산층의 증가로 최근 몇 년 동안 이국적인 애완동물로서의 NHP에 대한 수요가 증가했다.[200]8. 4. 과학 연구
영장류 잡종은 사육 상태에서 종종 생기지만,[304] 야생에서 생기기도 한다.[305][306] 잡종은 2개 종의 활동 범위가 잡종 지역을 형성하여 겹칠 때 생긴다. 잡종은 동물들이 동물원 등에 있을 때 인간에 의해 만들어지거나 약탈과 같은 환경적 압박 때문에 만들어질 수 있다.[305] 속간 교잡, 즉 다른 속 사이의 잡종 또한 야생에서 발견되기도 한다. 비록 그들이 몇 백만 년 동안 구별되는 속에 속해 있지만, 이종 교잡은 단지 겔라다속과 망토개코원숭이 사이에서만 발생한다.[307]
수천 마리의 비인간 영장류가 인간과의 심리적, 생리적 유사성 때문에 전 세계에서 연구에 사용된다.[206][205] 특히 비인간 영장류의 뇌와 눈은 다른 어떤 동물보다 인간 해부학과 더 가깝다. 비인간 영장류는 일반적으로 전임상 시험, 신경과학, 안과학 연구 및 독성 연구에 사용된다. 붉은털원숭이는 다른 마카크, 아프리카 녹색 원숭이, 침팬지, 개코원숭이, 다람쥐원숭이 및 비단원숭이와 마찬가지로 야생에서 포획되거나 목적에 맞게 사육된다.[206][207]
2005년, 유인원 프로젝트(GAP)는 미국에서 사육 중인 3,100마리의 비인간 영장류 중 1,280마리가 실험에 사용되었다고 보고했다.[196] 2004년, 유럽 연합은 이러한 실험에 약 10,000마리의 비인간 영장류를 사용했고, 2005년 영국에서는 3,115마리의 비인간 영장류를 대상으로 4,652건의 실험이 진행되었다.[208] 많은 국가의 정부는 사육되는 비인간 영장류에 대한 엄격한 관리 요구 사항을 가지고 있다. 미국에서는 연방 지침에 따라 비인간 영장류의 사육, 먹이 주기, 환경 개선 및 번식 측면을 광범위하게 규제한다.[209] 유럽 동물 실험 종식 연합과 같은 유럽 단체는 유럽 연합의 동물 실험 법률 검토의 일환으로 실험에 모든 비인간 영장류 사용 금지를 모색하고 있다.[210]
제2차 세계 대전 이후, 이마니시 긴지 등이 미야자키현의 고시마와 다카사키야마에서 야생 일본원숭이 무리에 먹이를 주는 데 성공한 이후, 일본의 영장류 연구는 비약적인 발전을 이루었다. 이마니시 등의 일본원숭이의 문화적 행동에 대한 연구는 세계적으로 주목받았으며, 이후 영장류 연구의 방향성에 중대한 지침을 제시했다.
그 후에도 일본원숭이에 그치지 않고, 이타니 준이치로 등 많은 일본인들이 고릴라, 침팬지, 보노보 등의 유인원을 비롯하여, 동남아시아부터 인도에 걸친 꼬리원숭이, 남아메리카의 신세계원숭이 등 거의 모든 원숭이를 망라한 현장 연구를 수행하며, 선도적인 연구를 지속하고 있다.
일본의 원숭이학은 생태학적 연구뿐만 아니라 사회학, 생리학, 유전학, 형태학, 운동학 등 다방면에 걸쳐, 주로 교토 대학과 오사카 대학에서 오늘날에도 활발한 연구가 이루어지고 있다.
8. 5. 보존
국제 자연 보전 연맹(IUCN)은 영장류의 3분의 1 이상을 심각한 멸종 위기 또는 취약종으로 분류한다. 약 60%의 영장류 종이 멸종 위협에 처해 있으며, 세부적으로 마다가스카르 종의 87%, 아시아 종의 73%, 아프리카 종의 37%, 남미 및 중앙 아메리카 종의 36%가 이에 해당한다.[211] 또한, 전체 영장류 종의 75%가 개체 수 감소를 겪고 있다.[211] 모든 영장류 종은 부록 II에 등재되어 무역이 규제되며, 그중 50종 및 아종은 부록 I에 등재되어 무역으로부터 완전한 보호를 받는다.[212][213]



영장류 종의 주요 위협 요인으로는 삼림 벌채, 숲 파편화, 원숭이 몰이(영장류의 작물 약탈로 인해 발생),[214] 그리고 약용, 애완용, 식용 목적의 영장류 사냥 등이 있다. 특히 대규모 열대 우림 개간은 영장류를 가장 심각하게 위협하는 요인으로 꼽힌다.[215][216][217] 전체 영장류 종의 90% 이상이 열대 우림에 서식하고 있다.[216][218] 산림 손실의 주된 원인은 농업을 위한 개간이지만, 상업적 벌목, 자급 목적의 목재 수확, 광업, 댐 건설 등도 열대 우림 파괴에 영향을 미친다.[218] 일례로, 인도네시아에서는 팜유 생산 확대를 위해 광대한 저지대 숲이 개간되었으며, 위성 이미지 분석 결과 1998년과 1999년 사이에 르우제르 생태계에서만 연간 1,000마리의 수마트라 오랑우탄이 사라진 것으로 나타났다.[219]
몸집이 큰(5kg 이상) 영장류는 밀렵꾼에게 더 큰 이익을 가져다주기 때문에 멸종 위험이 더 크다.[218] 이들은 성숙이 늦고 출생 간격이 길어, 밀렵이나 애완동물 거래로 인해 개체 수가 줄어들면 회복 속도가 더디다.[220] 아프리카 일부 도시의 조사에 따르면, 도시 지역에서 소비되는 단백질의 절반이 부시미트 거래에서 비롯된다.[221] 게논이나 드릴과 같은 멸종 위기 영장류는 지속 가능한 수준을 훨씬 넘어서는 강도로 사냥되고 있다.[221] 이는 큰 몸집, 운반의 용이성, 그리고 개체당 높은 수익성 때문이다.[221] 농업이 삼림 서식지를 잠식하면서, 영장류는 농작물을 먹이로 삼아 농부들에게 막대한 경제적 손실을 입히기도 한다.[222] 이러한 영장류의 작물 약탈은 지역 주민들에게 영장류에 대한 부정적 인식을 심어주어, 보존 노력을 더욱 어렵게 만든다.[223]
마다가스카르에서는 1,500년 전 인간이 정착한 이후, 사냥과 서식지 파괴로 인해 최소 8개 종류와 15개의 대형 영장류 종이 멸종했다.[224] 멸종된 영장류에는 은색 고릴라보다 더 큰 여우원숭이인 ''아르케오인드리''와 팔레오프로피테쿠스과, 고대여우원숭이과 등이 포함된다.[224]
아시아의 경우, 힌두교, 불교, 이슬람교에서는 영장류 고기 섭취를 금지하지만, 여전히 식용을 목적으로 한 사냥이 이루어지고 있다.[218] 일부 소규모 전통 종교에서는 영장류 고기 섭취를 허용하기도 한다.[225][226] 애완동물 거래와 전통 의학 또한 불법 사냥 수요를 부추기는 요인이다.[200][227][228] 붉은털원숭이는 1960년대에 과도한 포획으로 개체 수가 위협받은 후 보호를 받았으며, 이 보호 프로그램은 매우 성공적이어서 현재는 서식지 전역에서 해충으로 여겨질 정도이다.[217]
중앙아메리카와 남아메리카에서는 숲 파편화와 사냥이 영장류에게 가장 큰 두 가지 문제이다. 현재 중앙아메리카에는 넓은 숲이 드물게 남아있다.[215][229] 이는 숲의 가장자리 면적을 넓혀 농지 침입, 습도 감소, 식생 변화와 같은 가장자리 효과에 취약하게 만든다.[230][231] 이동 제한은 근친 교배를 증가시키고, 이는 병목 현상으로 이어져 유전적 다양성 감소와 같은 부정적인 결과를 초래할 수 있다.[232][233]
현재 21종의 영장류가 심각한 멸종 위기에 처해 있으며, 그중 7종(실키 시파카, 들라쿠르랑구르, 흰머리랑구르, 회색 다리 두크, 통킹 스너브코 원숭이, 크로스리버 고릴라, 수마트라 오랑우탄)은 2000년 이후 IUCN의 "세계에서 가장 멸종 위기에 처한 영장류 25종" 목록에 계속 포함되어 있다.[234] 미스 왈드론붉은콜로부스는 1993년부터 1999년까지 아종의 흔적을 찾을 수 없어 최근 멸종된 것으로 발표되었다.[235] 이후 몇몇 사냥꾼이 이들을 발견하고 사살했지만, 여전히 이 아종의 생존 전망은 매우 어둡다.[236]
8. 5. 1. 한국의 영장류 연구
제2차 세계 대전 이후, 이마니시 긴지 등이 미야자키현의 고시마와 다카사키야마에서 야생 일본원숭이 무리에 먹이를 주는 데 성공한 이후, 일본의 영장류 연구는 비약적인 발전을 이루었다. 이마니시 등의 일본원숭이의 문화적 행동에 대한 연구는 세계적으로 주목받았으며, 이후 영장류 연구의 방향성에 중대한 지침을 제시했다.그 후에도 일본원숭이에 그치지 않고, 이타니 준이치로 등 많은 일본인들이 고릴라, 침팬지, 보노보 등의 유인원을 비롯하여, 동남아시아부터 인도에 걸친 꼬리원숭이, 남아메리카의 신세계원숭이 등 거의 모든 원숭이를 망라한 현장 연구를 수행하며, 선도적인 연구를 지속하고 있다.
일본의 원숭이학은 생태학적 연구뿐만 아니라 사회학, 생리학, 유전학, 형태학, 운동학 등 다방면에 걸쳐, 주로 교토 대학과 오사카 대학에서 오늘날에도 활발한 연구가 이루어지고 있다.
참조
[1]
MSW3 Groves
[2]
논문
The evolutionary radiation of plesiadapiforms
2017
[3]
백과사전
Primate
http://www.merriam-w[...]
Merriam-Webster
2008-07-21
[4]
서적
The Book of Popular Science
https://books.google[...]
[5]
EB1911
[6]
뉴스
Why don't humans have tails? Scientists find answers in an unlikely place
https://www.cnn.com/[...]
CNN
2024-03-23
[7]
Citation
The Natural History of the Gorilla
Weidenfeld & Nicolson
[8]
Citation
Classification by molecules: What's in it for field botanists?
http://www.watsonia.[...]
2010-02-07
[9]
Citation
Contemporary Debates in Philosophy of Biology
[10]
서적
The Human Lineage
https://books.google[...]
John Wiley & Sons
[11]
서적
South American Primates: Comparative Perspectives in the Study of Behavior, Ecology, and Conservation
Springer
[12]
서적
Sistema naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus differentiis, synonimis locis. Tomus I
Impensis direct. Laurentii Salvii, Holmia
[13]
서적
Sistema naturae sive regna tria Naturae systematice proposita per classes, ordines, genera, & species
apud Theodorum Haak, Lugduni Batavorum
[14]
서적
Annales Françaises et Etrangères d'Anatomie et de Physiologie Appliquées à la Médicine et à l'Histoire Naturelle, 3
[15]
서적
Orders and Families of Recent Mammals of the World
John Wiley and Sons
[16]
서적
Classification of Mammals: Above the species level
Columbia University Press
[17]
서적
Primate Behavioral Ecology
Pearson Allyn and Bacon
[18]
논문
Molecular and Genomic Data Identify the Closest Living Relative of Primates
2007-11-02
[19]
서적
A Complete Guide to Monkeys, Apes and Other Primates
https://archive.org/[...]
Viking Press
[20]
서적
Classification of Mammals Above the Species Level
Columbia University Press
[21]
논문
New perspectives on anthropoid origins
[22]
논문
Macroevolutionary Dynamics and Historical Biogeography of Primate Diversification Inferred from a Species Supermatrix
[23]
논문
Genomic data reject the hypothesis of a prosimian primate clade
2011-09
[24]
논문
Primate phylogenetic relationships and divergence dates inferred from complete mitochondrial genomes
2014-06
[25]
논문
A Mitogenomic Phylogeny of Living Primates
2013-07-16
[26]
논문
Earliest Palaeocene purgatoriids and the initial radiation of stem primates
[27]
논문
New perspectives on anthropoid origins
[28]
논문
Deep Time and the Search for Anthropoid Origins
http://www.paleontol[...]
[29]
논문
Estimating the phylogeny and divergence times of primates using a supermatrix approach
2009-10-27
[30]
논문
The placental mammal ancestor and the post–K-Pg radiation of placentals
2013-02-08
[31]
논문
Molecular Clock Calibrations and Metazoan Divergence Dates
https://archive.org/[...]
1999-09
[32]
웹사이트
Scientists Push Back Primate Origins From 65 Million To 85 Million Years Ago
https://www.scienced[...]
2008-10-24
[33]
논문
Using the fossil record to estimate the age of the last common ancestor of extant primates
https://archive.org/[...]
2002-04-18
[34]
논문
Molecular Remodeling of Members of the Relaxin Family During Primate Evolution
[35]
논문
Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs
[36]
논문
Lemur Diversity in Madagascar
https://web.archive.[...]
2019-09-24
[37]
서적
Primates in Perspective
Oxford University Press
[38]
논문
"Darwinius masillae is a strepsirrhine—a reply to Franzen et al. (2009)"
https://web.archive.[...]
2015-09-04
[39]
서적
Primate Evolution and Human Origins
Benjamin/Cummings
[40]
서적
Mammals of Madagascar, A Complete Guide
A&C Black Publishers
[41]
서적
Lemurs of Madagascar
Conservation International
[42]
웹사이트
Primate Evolution
https://web.archive.[...]
University of Edinburgh
2000-10-20
[43]
간행물
Evolutionary Biology of Tarsiers
https://web.archive.[...]
2008-08-22
[44]
논문
Rapid electrostatic evolution at the binding site for cytochrome c on cytochrome c oxidase in anthropoid primates
2005-05-03
[45]
웹사이트
Early Primate Weighed Less Than an Ounce
https://web.archive.[...]
ScienceNow
2013-06-07
[46]
논문
Evidence for an Asian origin of stem anthropoid s
2012
[47]
논문
Late Middle Eocene primate from Myanmar and the initial anthropoid colonization of Africa
[48]
논문
Anthropoid primates from the Oligocene of Pakistan (Bugti Hills): Data on early anthropoid evolution and biogeography
2005-06-14
[49]
논문
Timing the Origin of New World Monkeys
[50]
논문
The origin of platyrrhines: An evaluation of the Antarctic scenario and the floating island model
https://archive.org/[...]
[51]
논문
Middle Miocene Dispersals of Apes
[52]
서적
Primate Behavioral Ecology
Allyn & Bacon
[53]
서적
Vertebrate Life
https://archive.org/[...]
Pearson
[54]
서적
Primate Taxonomy
Smithsonian Institution Press
[55]
웹사이트
Primate diversity by region
http://www.primate-s[...]
International Union for the Conservation of Nature
2021-03-01
[56]
논문
Songs of hybrid gibbons (Hylobates lar × H. muelleri)
[57]
논문
Naturally occurring primate hybrid
[58]
논문
Sociological study of a wild group of hybrid baboons between Papio anubis and P. hamadryas in the Awash Valley, Ethiopia
1979-01
[59]
논문
Intergeneric Hybrid Baboons
[60]
논문
Cloning of Macaque Monkeys by Somatic Cell Nuclear Transfer
2018-01-24
[61]
논문
These monkey twins are the first primate clones made by the method that developed Dolly
https://www.science.[...]
2018-01-24
[62]
논문
First monkeys cloned with technique that made Dolly the sheep - Chinese scientists create cloned primates that could revolutionize studies of human disease.
2018-01-24
[63]
뉴스
First monkey clones created in Chinese laboratory
https://www.bbc.com/[...]
BBC News
2018-01-24
[64]
뉴스
Scientists Successfully Clone Monkeys; Are Humans Up Next?
https://www.nytimes.[...]
The New York Times
2018-01-24
[65]
서적
An Introduction to Human Evolutionary Anatomy
https://archive.org/[...]
Academic Press
[66]
웹사이트
"Primates" (On-line)
http://animaldiversi[...]
Animal Diversity Web
2008-06-03
[67]
논문
Visual Acuity and the Evolution of Signals
https://www.cell.com[...]
2018-07-29
[68]
간행물
The Evolution of High Visual Acuity in the Anthropoidea
https://doi.org/10.1[...]
Springer US
2023-07-30
[69]
서적
Humankind Emerging
Allyn & Bacon
[70]
웹사이트
Archonta: Primates
http://www.palaeos.c[...]
Palaeos
2006-01-08
[71]
서적
Wheeler's Dental Anatomy, Physiology, and Occlusion
https://archive.org/[...]
W.B. Saunders
[72]
서적
Vertebrate Life
https://archive.org/[...]
Pearson
[73]
논문
The ecological role of the prehensile tail in white-faced capuchins (Cebus capucinus)
https://archive.org/[...]
1999-11
[74]
논문
Tail growth tracks the ontogeny of prehensile tail use in capuchin monkeys (Cebus albifrons and C. apella)
https://archive.org/[...]
2011-11
[75]
서적
Primate Anatomy: An Introduction
https://books.google[...]
Academic Press
2010-07-27
[76]
간행물
Primates
The Brown Reference Group plc
[77]
논문
Mammals in Which Females are Larger Than Males
[78]
논문
Seasonality, Fasting Endurance, and Body Size in Mammals
1985-07
[79]
논문
Sex-differences in the canines of the gibbon (''Hylobates lar'')
[80]
논문
The functional adaptations of primate molar teeth
[81]
서적
Sexual selection and the descent of man
Aldine Transaction
[82]
논문
The quantitative assessment of phylogenetic constraints in comparative analyses: Sexual dimorphism in body weight among primates
https://archive.org/[...]
1985-11
[83]
논문
Correlates of sexual dimorphism in primates: Ecological and size variables
[84]
논문
Sexual dimorphism in primate evolution
[85]
논문
Sexual dimorphism and facial growth in papionine monkeys
[86]
서적
Primate Ecology and Social Structure Volume 1: Lorises, Lemurs and Tarsiers
https://archive.org/[...]
Pearson Custom Publishing & Prentice Hall
[87]
서적
Selected Readings in the Anthropology of Religion: Theoretical and Methodological Essays
https://archive.org/[...]
Greenwood Publishing Group
[88]
논문
Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (''Setonix brachyurus'') and quenda (''Isoodon obesulus'')
[89]
논문
Photosensitive and photostable pigments in the retinae of Old World monkeys
http://jeb.biologist[...]
2008-06-16
[90]
논문
Evolution and selection of trichromatic vision in primates
[91]
논문
Evolution and function of routine trichromatic vision in primates
https://archive.org/[...]
[92]
서적
Primate Ecology and Social Structure, Volume 2: New World Monkeys
Pearson Custom Publishing & Prentice Hall
[93]
서적
Current Problems in Sociobiology
Cambridge University Press
[94]
논문
Genetic correlates of social behavior in wild chimpanzees: evidence from mitochondrial DNA
1997-09
[95]
뉴스
Neanderthal family found cannibalised in cave in Spain
https://www.bbc.co.u[...]
2010-12-21
[96]
뉴스
Ancient cave women 'left childhood homes'
https://www.bbc.co.u[...]
2011-06-02
[97]
논문
Strontium isotope evidence for landscape use by early hominins
2011-06-01
[98]
서적
Primates in Perspective
Oxford University Press
[99]
서적
Primates in Perspective
Oxford University Press
[100]
학위논문
Behavior and ecology of wild slow lorises (''Nycticebus coucang''): social organization, infant care system, and diet
http://deposit.ddb.d[...]
Bayreuth University
[101]
서적
Primates in Perspective
Oxford University Press
[102]
서적
Great Ape Societies
https://archive.org/[...]
Cambridge Univ Press
[103]
논문
Inbreeding depression in non-human primates: a historical review of methods used and empirical data
https://hal.archives[...]
2007-12
[104]
논문
Effect of inbreeding on infant mortality in captive primates
http://si-pddr.si.ed[...]
[105]
논문
Noninvasive paternity assignment in Gombe chimpanzees
https://archive.org/[...]
2001-05
[106]
서적
The Pictorial Guide to the Living Primates
https://archive.org/[...]
Pogonias Press
[107]
간행물
Fission–fusion populations
2009-08
[108]
서적
Vertebrate Life
https://archive.org/[...]
Pearson
[109]
문서
Primate Societies
University of Chicago Press
[110]
뉴스
Scientists Find First Evidence That Groups of Apes Cooperate - Some bonobos are challenging the notion that humans are the only primates capable of group-to-group alliances.
https://www.nytimes.[...]
2023-11-16
[111]
간행물
Cooperation across social borders in bonobos
https://www.science.[...]
2023-11-16
[112]
서적
Monkeys of Tai Forest, An African Primate Community
Cambridge University Press
[113]
서적
Monkeys of Tai Forest, An African Primate Community
Cambridge University Press
[114]
서적
Chimpanzee and Red Colobus : the ecology of predator and prey
https://archive.org/[...]
Harvard University Press
[115]
서적
On the Move : how and why animals travel in groups
University of Chicago Press
[116]
간행물
Relaxed selective pressure on an essential component of pheromone transduction in primate evolution
http://www.pnas.org/[...]
2008-07-23
[117]
서적
Encyclopedia of Language and Linguistics
Elsevier
[118]
간행물
The facial expression musculature in primates and its evolutionary significance
https://www.research[...]
[119]
간행물
Ape gestures and language evolution
[120]
간행물
Diurnal Distribution of Loud Calls in Sympatric Wild Indris (Indri indri) and Ruffed Lemurs (Varecia variegata): Implications for Call Functions
http://gibbons.de/ma[...]
[121]
간행물
Primate communication in the pure ultrasound
[122]
간행물
Roars of Black Howler Monkeys (Alouatta caraya): Evidence for a Function in Inter-Group Spacing
[123]
웹사이트
Black howler monkey
https://nationalzoo.[...]
Smithsonian's National Zoo & Conservation Biology Institute
2016-04-25
[124]
간행물
The vocal organ of the Howling monkey (Alouatta palliata)
[125]
간행물
Vervet Monkey Alarm Calls: Semantic communication in a Free-Ranging Primate
https://www.research[...]
[126]
간행물
Call combinations and compositional processing in wild chimpanzees
2023
[127]
간행물
Suffixation influences receivers' behaviour in non-human primates
2015
[128]
간행물
Morphologically structured vocalizations in female Diana monkeys
2016
[129]
간행물
Call Combinations in Great Apes and the Evolution of Syntax
2020
[130]
간행물
Brief report: Occurrence of anti-resonance in the song of the siamang (Hylobates syndactylus)
[131]
간행물
Monkey vocal tracts are speech-ready
[132]
간행물
Evidence of a Vocalic Proto-System in the Baboon (Papio papio) Suggests Pre-Hominin Speech Precursors
[133]
간행물
Orangutan information broadcast via consonant-like and vowel-like calls breaches mathematical models of linguistic evolution
[134]
간행물
Vocal responsiveness in male wild chimpanzees: implications for the evolution of language
2000-08
[135]
간행물
Male infanticide leads to social monogamy in primates
[136]
간행물
Social and genetic aspects of paternity in wild long-tailed macaques (Macaca fascicularis)
[137]
간행물
Nests, Tree Holes, and the Evolution of Primate Life Histories
[138]
간행물
Park or ride? Evolution of infant carrying in primates.
Kluwer Academic Publishing
[139]
웹사이트
Humans And Primates Burn 50 Percent Fewer Calories Each Day Than Other Mammals
http://www.ibtimes.c[...]
IBT Media Inc.
2014-01-14
[140]
간행물
Menopause in nonhuman primates?
[141]
간행물
Diet and Primate Evolution
http://nature.berkel[...]
[142]
논문
Vitamin C biosynthesis in prosimians: Evidence for the anthropoid affinity of Tarsius
http://www3.intersci[...]
2010-03-16
[143]
논문
Independent digit control in foraging by the aye-aye (''Daubentonia madagascariensis'')
[144]
웹사이트
"''Theropithecus gelada''"
http://animaldiversi[...]
2008-08-08
[145]
서적
Tarsiers Past, Present and Future
Rutgers University Press
[146]
서적
The Chimpanzees of Gombe: Patterns of Behavior
https://archive.org/[...]
Belknap Press of Harvard University Press
[147]
웹사이트
WHAT DO CHIMPS EAT?
http://www.allaboutw[...]
2013-04-22
[148]
논문
Observations on the meat-eating behavior of wild bonobos (''Pan paniscus'') at Wamba, Republic of Zaire
[149]
서적
Bonobo Husbandry Manual
American Association of Zoos and Aquariums
[150]
논문
Evidence for the consumption of arboreal, diurnal primates by bonobos (''Pan paniscus'')
[151]
논문
Primate hunting by bonobos at LuiKotale, Salonga National Park
2008-10-14
[152]
논문
Origins and evolution of the Western diet: health implications for the 21st century
2005-02
[153]
논문
Human eating behaviour in an evolutionary ecological context
2002-11
[154]
웹사이트
Earliest agriculture in the Americas
http://www.archaeolo[...]
[155]
논문
The gourmet ape: evolution and human food preferences
2009-09
[156]
논문
Phylogenetic analysis of the evolution of lactose digestion in adults
https://archive.org/[...]
1997-10
[157]
서적
The Evolution of Primate Societies
University of Chicago Press
[158]
논문
Tool Use and Tool Making in Wild Chimpanzees
[159]
논문
Why some capuchin monkeys (''Cebus apella'') use probing tools (and others do not)
https://archive.org/[...]
[160]
논문
Capuchin cognitive ecology: cooperation based on projected returns
[161]
논문
Recognizing facial cues: Individual discrimination by chimpanzees (''Pan troglodytes'') and rhesus monkeys (''Macaca mulatta'')
https://archive.org/[...]
[162]
논문
Neocortex size predicts deception rate in primates
[163]
논문
Visual kin recognition in chimpanzees
[164]
논문
Discrimination of macaques by macaques: The case of sulawesi species
[165]
논문
Object permanence in orangutans (''Pongo pygmaeus''), chimpanzees (''Pan troglodytes''), and children (''Homo sapiens'')
https://archive.org/[...]
[166]
논문
Use of experimenter-given cues during object-choice tasks by chimpanzees (''Pan troglodytes''), an orangutan (''Pongo pygmaeus''), and human infants (''Homo sapiens'')
https://archive.org/[...]
1998-06
[167]
논문
Rhesus monkeys use geometric and nongeometric information during a reorientation task
http://cogprints.org[...]
[168]
서적
Primate Cognition
Oxford University Press US
[169]
논문
Do some taxa have better domain-general cognition than others? A metaanalysis of nonhuman primate studies
[170]
논문
The evolution of primate general and cultural intelligence
http://lalandlab.st-[...]
2011-07-04
[171]
웹사이트
Toolmaking
https://www.janegood[...]
The Jane Goodall Institute
2013-08-01
[172]
웹사이트
Bonobos
http://www.clemetzoo[...]
ApeTag
2013-08-03
[173]
논문
" A comparison of bonobo and chimpanzee tool use: evidence for a female bias in the ''Pan'' lineage"
http://www.emory.edu[...]
[174]
간행물
Orangutans use simple tools to catch fish
https://www.wired.co[...]
2013-08-05
[175]
논문
First observation of tool use in wild gorillas
[176]
논문
Use of a club by a wild white-faced capuchin (''Cebus capucinus'') to attack a venomous snake (''Bothrops asper'')
[177]
논문
Wild capuchin monkeys (''Cebus libidinosus'') use anvils and stone pounding tools
[178]
논문
The physical characteristics and usage patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand
[179]
논문
Defensive stoning by baboons
[180]
서적
Mind the Gap: Tracing the Origins of Human Universals
Springer
[181]
논문
Drinking tools of wild chimpanzees at Bossou
[182]
웹사이트
Sumatran orangutans
http://orangutanisla[...]
OrangutanIslands.com
2013-08-02
[183]
논문
Manufacture and use of tools in wild Sumatran orangutans
https://archive.org/[...]
[184]
웹사이트
Mandrill monkey makes 'pedicuring' tool
http://www.bbc.co.uk[...]
BBC
2013-08-11
[185]
웹사이트
Gorillas and Tools – Part I
http://www.rozhlas.c[...]
2013-08-04
[186]
서적
The Princeton Encyclopedia of Mammals
Princeton and Oxford University Press
[187]
논문
Geographic and climatic control of primate diversity
1995-08-15
[188]
서적
Primates in Perspective
Oxford University Press
[189]
논문
Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (''Rhinopithecus bieti'')
1994-04
[190]
서적
The Mountain Gorilla: Ecology and Behavior
https://archive.org/[...]
University Chicago Press
[191]
서적
Primate Societies
The University of Chicago Press
[192]
논문
Patterns of Water Use in Primates
[193]
서적
Primates in Perspective
Oxford University Press
[194]
논문
Zoonoses Acquired from Pet Primates
http://pin.primate.w[...]
2008-08-11
[195]
웹사이트
The Universal Declaration of Human Rights
https://www.un.org/O[...]
United Nations
2008-12-02
[196]
웹사이트
Declaration on Great Apes
http://www.greatapep[...]
Great Ape Project
2008-06-16
[197]
뉴스
Spanish parliament approves 'human rights' for apes
https://www.theguard[...]
2008-11-10
[198]
뉴스
Of great apes and men
https://www.theguard[...]
2008-11-10
[199]
간행물
The Perils of Keeping Monkeys as Pets
http://news.national[...]
2013-02-06
[200]
논문
Primate conservation in Vietnam: toward a holistic environmental narrative
https://archive.org/[...]
2004-06
[201]
웹사이트
IPPL News: The US Pet Monkey Trade
http://www.aesop-pro[...]
International Primate Protection League
2008-08-04
[202]
웹사이트
The Beginnings of Research in Space Biology at the Air Force Missile Development Center, 1946–1952
https://history.nasa[...]
NASA
2008-08-18
[203]
뉴스
Monkeys as Helpers To Quadriplegics At Home
https://query.nytime[...]
2008-10-08
[204]
서적
Encountering Gorillas: A Chronicle of Discovery, Exploitation, Understanding, and Survival
https://archive.org/[...]
Rowman and Littlefield
[205]
논문
Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees
2001-02
[206]
웹사이트
The supply and use of primates in the EU
http://www.ebra.org/[...]
European Biomedical Research Association
2008-08-18
[207]
논문
A Demographic Analysis of Primate Research in the United States
[208]
서적
Statistics of scientific procedures on living animals: Great Britain 2005
http://www.official-[...]
The Stationery Office
2008-06-16
[209]
웹사이트
Nonhuman Primates: Research Animals
http://awic.nal.usda[...]
United States Department of Agriculture
2008-07-14
[210]
웹사이트
Directive 86/609
http://www.eceae.org[...]
European Coalition to End Animal Experiments
2008-10-08
[211]
논문
Impending extinction crisis of the world's primates: Why primates matter
2017-01-01
[212]
서적
Born to be wild: Primates are not pets
http://www.ifaw.org/[...]
International Fund for Animal Welfare
2011-02-26
[213]
웹사이트
Appendices I, II and III
http://www.cites.org[...]
Convention on International Trade in Endangered Species of Wild Fauna and Flora
2012-04-02
[214]
서적
Mammals of Ghana, Sierra Leone, and the Gambia
Trendrine
[215]
논문
Primate conservation in the new millennium: the role of scientists
[216]
서적
Primate Societies
University of Chicago Press
[217]
논문
Status, conservation and management of primates in India
http://www.wii.gov.i[...]
2008-08-04
[218]
서적
Primate Conservation Biology
University of Chicago Press
[219]
논문
Dramatic decline in orangutan numbers in the Leuser Ecosystem, northern Sumatra
[220]
논문
Predicting extinction risk in declining species
[221]
논문
Impact of market hunting on mammal species in Equatorial Guinea
https://archive.org/[...]
[222]
논문
Crop-raiding by wild vertebrates: The farmer's perspective in an agricultural community in western Uganda
[223]
논문
Primate conservation and local communities: Ethical issues and debates
https://archive.org/[...]
[224]
백과사전
Primate
http://www.britannic[...]
Encyclopædia Britannica, Inc.
2008-07-21
[225]
논문
Primates in Northeast India: an overview of their distribution and conservation status
http://www.wii.gov.i[...]
2008-08-04
[226]
논문
Distribution and abundance of primates in rainforests of the Western Ghats, Karnataka, India and the conservation of ''Macaca silenus''
2004-10-01
[227]
논문
Conservation of the Javan gibbon ''Hylobates moloch'': population estimates, local extinction, and conservation priorities
http://rmbr.nus.edu.[...]
2008-08-04
[228]
논문
Abundance and distribution of sympatric gibbons in a threatened Sumatran rain forest
2004-04-01
[229]
논문
Non-flying mammals and landscape changes in the tropical forest region of Los Tuxtlas, Mexico
1994-09-01
[230]
서적
Primates in Fragments: Ecology and Conservation
Kluwer Academic/Plenum Publishers
[231]
논문
Species loss in fragments of tropical rain forest: a review of the evidence
https://archive.org/[...]
[232]
서적
Primates in Fragments: Ecology and Conservation
Kluwer Academic/Plenum Publishers
[233]
서적
Conservation Genetics: Case Histories from Nature
https://archive.org/[...]
Kluwer Academic Publishers
[234]
서적
Primates in Peril: The World's 25 Most Endangered Primates 2008–2010
http://www.primate-s[...]
IUCN/SSC Primate Specialist Group (PSG), International Primatological Society (IPS), and Conservation International (CI)
[235]
논문
Extinction of a West African Red Colobus Monkey
2000-10-01
[236]
논문
Update on the Search for Miss Waldron's Red Colobus Monkey
2005-06-01
[237]
문서
I, II and III (valid from 31 July 2021)
https://cites.org/si[...]
[238]
서적
Order Primates
http://www.departmen[...]
Johns Hopkins University Press
[239]
서적
霊長類
平凡社
[240]
웹사이트
日本モンキーセンター 霊長類和名リスト 2018年11月版
http://www.j-monkey.[...]
2018-12-16
[241]
논문
世界哺乳類標準和名目録
日本哺乳類学会
[242]
논문
哺乳類の日本語分類群名,特に目名の取扱いについて 文部省の“目安”にどう対応するか
日本哺乳類学会
[243]
논문
サルの分類名(その1:マカク)
日本霊長類学会
[244]
간행물
用語解説
朝日新聞社
[245]
논문
サルの分類名(その8:原猿)
日本霊長類学会
[246]
논문
サルの分類名(その6:マーモセット科)
https://doi.org/10.2[...]
日本霊長類学会
1988
[247]
논문
霊長類の共通祖先は夜行性だったか
https://hdl.handle.n[...]
京都大学学術出版会
2007
[248]
논문
昼も夜も動くキツネザル:周日行性の系統発生と至近メカニズム,および適応的意義をさぐる
https://doi.org/10.2[...]
日本霊長類学会
2017
[249]
논문
霊長類の起源と進化
https://doi.org/10.3[...]
バイオメカニズム学会
1997
[250]
서적
霊長類学への招待
京都大学学術出版会
2007
[251]
웹사이트
霊長類の進化とその系統樹
http://www.pri.kyoto[...]
[252]
논문
Vitamin C biosynthesis in prosimians: evidence for the anthropoid affinity of Tarsius
[253]
서적
系統樹をさかのぼって見えてくる進化の歴史
ベレ出版
2014-10-25
[254]
웹사이트
霊長類の色覚と進化
http://www.pri.kyoto[...]
2004-09-18
[255]
논문
Evolution and selection of trichromatic vision in primates
[256]
웹사이트
1.4 なぜ赤オプシン遺伝子と緑オプシン遺伝子が並んで配置しているのか
http://www.nig.ac.jp[...]
[257]
논문
新世界ザルRed-Green視物質遺伝子と色覚の進化
2000
[258]
논문
恒常的3色型色覚とされてきたホエザル属における種内L-Mオプシン多型の発見
https://doi.org/10.1[...]
2011
[259]
웹사이트
1.6 女性で赤緑色盲が少ない理由
http://www.nig.ac.jp[...]
[260]
뉴스
サルとヒトとの進化の分岐、定説より最近か ミシガン大
https://www.afpbb.co[...]
AFPBB News
2010-07-16
[261]
간행물
Nature2010年7月15日号
네이처
2010-07-15
[262]
논문
On the loss of uricolytic activity during primate evolution--I. Silencing of urate oxidase in a hominoid ancestor
[263]
간행물
Similar Functions of Uric Acid and Ascorbate in ManSimilar Functions of Uric Acid and Ascorbate in Man
http://www.drproctor[...]
[264]
문서
[265]
논문
DNA 레벨 상의 영장류 진화 및 사람아과 분류
[266]
논문
초행렬 접근을 이용한 영장류 계통발생 및 분기 싯점 추산(Estimating the phylogeny and divergence times of primates using a supermatrix approach)
[267]
논문
Molecular phylogenetics and the origins of placental mammals
http://www.ncbi.nlm.[...]
[268]
논문
Mammalian mitogenomic relationships and the root of the eutherian tree
http://www.pnas.org/[...]
[269]
논문
Evolutionary history of 7SL RNA-derived SINEs in Supraprimates
http://dx.doi.org/10[...]
[270]
논문
Molecular and Genomic Data Identify the Closest Living Relative of Primates
http://www.sciencema[...]
2008-08-17
[271]
서적
A Complete Guide to Monkeys, Apes and Other Primates
Viking Press
[272]
서적
Classification of Mammals Above the Species Level
https://archive.org/[...]
Columbia University Press
[273]
서적
The Human Lineage
https://books.google[...]
John Wiley & Sons
[274]
서적
Primate Taxonomy
Smithsonian Institution Press
[275]
논문
유인원 기원에 관한 새로운 관점(New perspectives on anthropoid origins)
[276]
웹인용
Nova – Meet Your Ancestors
http://www.pbs.org/w[...]
미국 공영 방송망(PBS)
2008-10-24
[277]
웹인용
플레시아다피스(Plesiadapis)
https://www.dmr.nd.g[...]
노스다코타 지질조사국(North Dakota Geological Survey)
2008-10-24
[278]
논문
Molecular Clock Calibrations and Metazoan Divergence Dates
1999-09
[279]
웹인용
Scientists Push Back Primate Origins From 65 Million To 85 Million Years Ago
http://www.scienceda[...]
Science Daily
2008-10-24
[280]
논문
Using the fossil record to estimate the age of the last common ancestor of extant primates
2002-04-18
[281]
저널
Molecular Remodeling of Members of the Relaxin Family During Primate Evolution
http://mbe.oxfordjou[...]
2008-08-22
[282]
저널
Development and Application of a Phylogenomic Toolkit: Resolving the Evolutionary History of Madagascar's Lemurs
https://web.archive.[...]
2008-08-22
[283]
저널
Lemur Diversity in Madagascar
2008-12
[284]
서적
Primates in Perspective
Oxford University Press
[285]
저널
Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology
http://www.plosone.o[...]
[286]
서적
Primate Evolution and Human Origins
Benjamin/Cummings
[287]
서적
Mammals of Madagascar, A Complete Guide
A&C Black Publishers
[288]
서적
Lemurs of Madagascar
https://archive.org/[...]
Conservation International
[289]
웹인용
Primate Evolution
http://homepage.mac.[...]
University of Edinburgh
2008-10-29
[290]
저널
Evolutionary Biology of Tarsiers
https://web.archive.[...]
2008-08-22
[291]
저널
Rapid electrostatic evolution at the binding site for cytochrome c on cytochrome c oxidase in anthropoid primates
http://www.pubmedcen[...]
2008-08-22
[292]
저널
Anthropoid primates from the Oligocene of Pakistan (Bugti Hills): Data on early anthropoid evolution and biogeography
http://www.pubmedcen[...]
2005-06-14
[293]
저널
Timing the Origin of New World Monkeys
http://mbe.oxfordjou[...]
[294]
저널
The origin of platyrrhines: An evaluation of the Antarctic scenario and the floating island model
[295]
저널
Middle Miocene Dispersals of Apes
[296]
서적
Primate Behavioral Ecology
Allyn & Bacon
[297]
서적
Vertebrate Life
Pearson
[298]
서적
South American Primates: Comparative Perspectives in the Study of Bahavior, Ecology, and Conservation
Springer
2009
[299]
서적
Sistema naturae per regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus differentiis, synonimis locis. Tomus I
Impensis direct. Laurentii Salvii, Holmia
[300]
서적
Sistema naturae sive regna tria Naturae systematice proposita per classes, ordines, genera, & species
apud Theodorum Haak, Lugduni Batavorum
[301]
서적
Annales Françaises et Etrangères d'Anatomie et de Physiologie Appliquées à la Médicine et à l'Histoire Naturelle, 3
[302]
서적
Classification of Mammals: Above the species level
Columbia University Press
[303]
서적
Primate Behavioral Ecology
[304]
저널
Songs of hybrid gibbons (''Hylobates lar'' × ''H. muelleri'')
[305]
저널
Naturally occurring primate hybrid
[306]
저널
Sociological study of a wild group of hybrid baboons between ''Papio anubis'' and ''P. hamadryas'' in the Awash Valley, Ethiopia
[307]
저널
Intergeneric Hybrid Baboons
[308]
저널
Nails and claws in primate evolution
[309]
서적
Primates
https://archive.org/[...]
The Brown Reference Group plc
[310]
서적
Vertebrate Life
Pearson
[311]
웹인용
Archonta: Primates
https://web.archive.[...]
Palaeos
2008-06-03
[312]
서적
Vertebrate Life
Pearson
[313]
서적
Humankind Emerging (8th edition)
Allyn & Bacon
[314]
서적
An Introduction to Human Evolutionary Anatomy
Academic Press
[315]
웹인용
영장류(브리태니커 백과사전 온라인)
http://www.britannic[...]
브리태니커 백과사전
2008-07-21
[316]
웹인용
'"Primates" (On-line)'
http://animaldiversi[...]
Animal Diversity Web
2008-06-03
[317]
서적
Wheeler's Dental Anatomy, Physiology, and Occlusion
W.B. Saunders
[318]
저널
Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (''Setonix brachyurus'') and quenda (''Isoodon obesulus'')
[319]
저널
Photosensitive and photostable pigments in the retinae of Old World monkeys
http://jeb.biologist[...]
2008-06-16
[320]
저널
Evolution and selection of trichromatic vision in primates
[321]
저널
Evolution and function of routine trichromatic vision in primates
[322]
서적
Primate Ecology and Social Structure, Volume 2: New World Monkeys
Pearson Custom Publishing & Prentice Hall
[323]
저널
Evolution and selection of trichromatic vision in primates
[324]
저널
Mammals in Which Females are Larger Than Males
[325]
저널
Seasonality, Fasting Endurance, and Body Size in Mammals
[326]
저널
Sex-differences in the canines of the gibbon (''Hylobates lar'')
[327]
저널
The functional adaptations of primate molar teeth
[328]
서적
Sexual selection and the descent of man
Aldine Transaction
[329]
저널
The quantitative assessment of phylogenetic constraints in comparative analyses: Sexual dimorphism in body weight among primates
[330]
저널
Correlates of sexual dimorphism in primates: Ecological and size variables
[331]
저널
Sexual dimorphism in primate evolution
[332]
저널
Sexual dimorphism and facial growth in papionine monkeys
[333]
서적
영장류 생태 및 사회 구조 1권: 로리스, 여우원숭이 및 안경원숭이
Pearson Custom Publishing & Prentice Hall
[334]
서적
Current Problems in Sociobiology
캠브리지 대학교 출판부
[335]
서적
Primates in Perspective
옥스포드 대학교 출판부
[336]
서적
Primates in Perspective
Oxford University Press
[337]
서적
Primates in Perspective
Oxford University Press
[338]
저널
Noninvasive paternity assignment in Gombe chimpanzees
[339]
서적
The Pictorial Guide to the Living Primates
https://archive.org/[...]
Pogonias Press
[340]
서적
Vertebrate Life
Pearson
[341]
문서
[342]
서적
Monkeys of Tai Forest, An African Primate Community
Cambridge University Press
[343]
서적
Monkeys of Tai Forest, An African Primate Community
Cambridge University Press
[344]
서적
Chimpanzee and Red Colobus : the ecology of predator and prey
https://archive.org/[...]
Harvard University Press
[345]
서적
On the Move : how and why animals travel in groups
University of Chicago Press
[346]
저널
Tool Use and Tool Making in Wild Chimpanzees
[347]
저널
Why some capuchin monkeys (''Cebus apella'') use probing tools (and others do not)
[348]
저널
Capuchin cognitive ecology: cooperation based on projected returns
[349]
저널
Recognizing facial cues: Individual discrimination by chimpanzees (''Pan troglodytes'') and rhesus monkeys (''Macaca mulatta'')
[350]
저널
Visual kin recognition in chimpanzees
[351]
논문
Discrimination of macaques by macaques: The case of sulawesi species
[352]
논문
Object permanence in orangutans (''Pongo pygmaeus''), chimpanzees (''Pan troglodytes''), and children (''Homo sapiens'')
[353]
논문
Use of experimenter-given cues during object-choice tasks by chimpanzees (''Pan troglodytes''), an orangutan (''Pongo pygmaeus''), and human infants (''Homo sapiens'')
[354]
논문
Rhesus monkeys use geometric and nongeometric information during a reorientation task
[355]
서적
Primate Cognition
https://archive.org/[...]
Oxford University Press US
[356]
논문
Relaxed selective pressure on an essential component of pheromone transduction in primate evolution
http://www.pnas.org/[...]
2008-07-23
[357]
서적
Encyclopedia of Language and Linguistics
Elsevier
2008-07-23
[358]
간행물
Diet and Primate Evolution
Scientific American
1993-08
[359]
논문
Independent digit control in foraging by the aye-aye (''Daubentonia madagascariensis'')
[360]
웹인용
"''Theropithecus gelada''"
http://animaldiversi[...]
2008-08-08
[361]
서적
Tarsiers Past, Present and Future
Rutgers University Press
[362]
서적
Monkeys of the Taï Forest : an African primate community
Cambridge University Press
[363]
서적
Chimpanzee and red colobus : the ecology of predator and prey
https://archive.org/[...]
Harvard University Press
[364]
논문
Geographic and climatic control of primate diversity
http://www.pnas.org/[...]
1995-08-15
[365]
서적
Primates in Perspective
Oxford University Press
[366]
논문
Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (''Rhinopithecus bieti'')
[367]
서적
The Mountain Gorilla: Ecology and Behavior
University Chicago Press
[368]
서적
Primates in Perspective
Oxford University Press
[369]
논문
Zoonoses Acquired from Pet Primates
http://pin.primate.w[...]
Veterinary Clinics of North America, Small Animal Practice
2008-08-11
[370]
웹인용
Declaration on Great Apes
http://www.greatapep[...]
Great Ape Project
2008-06-16
[371]
웹인용
Spanish parliament approves 'human rights' for apes
http://www.guardian.[...]
The Guardian
2008-06-26
[372]
웹인용
Of great apes and men
http://www.guardian.[...]
The Guardian
[373]
웹인용
The HSUS Applauds the U.S. Senate for Unanimously Passing the Captive Primate Safety Act
http://www.hsus.org/[...]
미국 휴메인 소사이어티
2006-07-06
[374]
논문
Primate conservation in Vietnam: toward a holistic environmental narrative
[375]
웹인용
IPPL News: The US Pet Monkey Trade
http://www.aesop-pro[...]
국제영장류보호연맹
2008-08-04
[376]
웹인용
The Beginnings of Research in Space Biology at the Air Force Missile Development Center, 1946-1952
http://history.nasa.[...]
미국 항공우주국
2008-08-18
[377]
웹인용
Monkeys as Helpers To Quadriplegics At Home
http://query.nytimes[...]
1987-06-17
[378]
논문
Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees
http://www.pubmedcen[...]
2008-10-08
[379]
웹인용
The supply and use of primates in the EU
http://www.ebra.org/[...]
European Biomedical Research Association
2008-08-18
[380]
논문
A Demographic Analysis of Primate Research in the United States
http://www.worldcong[...]
2008-10-08
[381]
서적
Statistics of scientific procedures on living animals: Great Britain 2005
http://www.official-[...]
The Stationery Office
2006-07
[382]
웹인용
Nonhuman Primates: Research Animals
http://awic.nal.usda[...]
United States Department of Agriculture
[383]
웹인용
Directive 86/609
http://www.eceae.org[...]
European Coalition to End Animal Experiments
2008-10-08
[384]
서적
Mammals of Ghana, Sierra Leone, and the Gambia
https://archive.org/[...]
Trendrine
[385]
논문
Primate conservation in the new millennium: the role of scientists
[386]
서적
Primate Societies
University of Chicago Press
[387]
저널
Status, conservation and management of primates in India
http://www.wii.gov.i[...]
2008-08-04
[388]
서적
Primate Conservation Biology
https://archive.org/[...]
University of Chicago Press
[389]
저널
Dramatic decline in orangutan numbers in the Leuser Ecosystem, northern Sumatra
[390]
저널
Predicting extinction risk in declining species
[391]
저널
Impact of market hunting on mammal species in Equatorial Guinea
[392]
저널
Crop-raiding by wild vertebrates: The farmer's perspective in an agricultural community in western Uganda
[393]
저널
Primate conservation and local communities: Ethical issues and debates
[394]
백과사전
Primate
http://www.britannic[...]
Encyclopædia Britannica, Inc.
2008-07-21
[395]
저널
Primates in Northeast India: an overview of their distribution and conservation status
http://www.wii.gov.i[...]
2008-08-04
[396]
저널
Distribution and abundance of primates in rainforests of the Western Ghats, Karnataka, India and the conservation of ''Macaca silenus''
[397]
저널
Conservation of the Javan gibbon ''Hylobates moloch'': population estimates, local extinction, and conservation priorities
http://rmbr.nus.edu.[...]
2008-08-04
[398]
저널
Abundance and distribution of sympatric gibbons in a threatened Sumatran rain forest
[399]
저널
Non-flying mammals and landscape changes in the tropical forest region of Los Tuxtlas, Mexico
[400]
서적
Primates in Fragments: Ecology and Conservation
Kluwer Academic/Plenum Publishers
[401]
저널
Species loss in fragments of tropical rain forest: a review of the evidence
[402]
서적
Primates in Fragments: Ecology and Conservation
https://archive.org/[...]
Kluwer Academic/Plenum Publishers
[403]
서적
Conservation Genetics: Case Histories from Nature
https://archive.org/[...]
Kluwer Academic Publishers
1996
[404]
저널
Primates in Peril: The World's 25 Most Endangered Primates, 2006 – 2008
http://www.primate-s[...]
2008-08-04
[405]
저널
Extinction of a West African Red Colobus Monkey
[406]
저널
Update on the Search for Miss Waldron’s Red Colobus Monkey
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com