상위성
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
상위성은 유전자 간의 상호 작용으로 인해 발생하는 현상으로, 유전학 연구에서 중요한 개념이다. 윌리엄 베이트슨 등에 의해 처음 사용되었으며, 유전자 상호 작용에 대한 이해가 발전하면서 그 중요성이 부각되었다. 상위성은 유전자의 효과가 다른 유전자의 존재 여부에 따라 달라지는 것을 의미하며, 유전자의 효과는 표현형의 크기나 단백질 활성으로 정량화될 수 있다. 상위성은 가산성, 크기 상위성, 부호 상위성 등 다양한 유형으로 분류되며, 유전체 내 유전자 간의 상호 작용, 유전자 내 코돈의 상호 작용, 단백질 내 아미노산 간의 상호 작용 등 다양한 분자적 원인에 의해 발생한다. 상위성은 적합도 지형과 진화 가능성에 영향을 미치며, 성의 진화와도 관련이 있다. 연구 방법으로는 회귀 분석, 이중 돌연변이 주기, 계산 예측 등이 사용되며, 생쥐의 털색 유전과 같이 다양한 예시를 통해 상위성의 개념을 이해할 수 있다.
더 읽어볼만한 페이지
| 상위성 | |
|---|---|
| 유전학 | |
| 유형 | 유전자 상호작용 |
| 정의 | |
| 정의 | 상위성은 한 유전자의 돌연변이 표현형이 다른 유전자의 돌연변이에 의존하는 현상이다. |
| 설명 | |
| 설명 | 상위성(上位性, Epistasis)은 한 유전자의 발현이 다른 유전자에 의해 가려지거나 변경되는 현상을 지칭한다. 즉, 한 유전자의 표현형이 다른 유전자의 존재에 의해 영향을 받는 경우를 말한다. 이는 유전자 간의 상호작용의 한 형태로, 독립적인 유전자들의 작용이 서로 얽혀 나타나는 결과이다. 상위 유전자(上 위 遺傳子, epistatic gene)는 다른 유전자의 표현을 가리는 유전자이며, 하위 유전자(下位遺傳子, hypostatic gene)는 상위 유전자에 의해 표현이 가려지는 유전자이다. 상위성은 유전적 배경, 즉 다른 유전자의 존재가 특정 유전자의 효과를 변경할 때 발생한다. 따라서 상위성은 유전적 배경의 돌연변이 효과를 고려한다. 상위성은 그 효과가 양적일 때, 즉 돌연변이가 동일한 정량적 특성의 값을 변경할 때 "양적"으로 간주된다. 이러한 의미에서 상위성은 형질을 변경하는 유전자 간의 상호 작용으로 정의된다. |
| 역사 | |
| 역사 | 이 용어는 윌리엄 베이트슨에 의해 처음 만들어졌으며, 그는 1907년에 그레고어 멘델의 독립적인 작용 법칙에서 벗어난 유전적 상호작용을 설명하기 위해 이 용어를 사용했다. 그는 그리스어 단어 'ἐπίστασις' (epistasis)에서 이 용어를 파생시켰는데, 이는 '멈추다'를 의미한다. 상위성의 개념은 처음에 한 유전자좌의 특정 대립유전자가 다른 유전자좌의 대립유전자의 표현을 가린다고 가정했다. 즉, 표현형이 하나의 유전자좌에 의해 결정되는 것으로 여겨졌다. 나중에 이 개념은 특정 유전자좌의 각 유전자가 표현형에 영향을 미치기 위해 서로 상호 작용하는 것까지 확장되었다. 로널드 피셔는 상위성이 적응도에 미치는 영향에 대한 이론적 연구를 수행했다. 수잔 알렌은 섬모 운동의 유전학에서 상위성 연구를 진행했다. 조너선 호지킨은 선충류에서 성 결정의 유전학에서 상위성 연구를 진행했다. |
| 예시 | |
| 예시 | 인간의 색소침착에서 대립유전자 H는 대립유전자 O에 대해 상위성을 가지며, 이는 H 대립유전자가 두 개 있는 경우(HH) 개인이 O 유전자좌에서 어떤 대립유전자를 가지고 있든 상관없이 색소가 없음을 의미한다. 또 다른 예는 식물 Brassica oleracea에서 발견된다. 이 식물에서 유전자 A와 B는 꽃 색깔을 조절한다. 유전자 A가 aa로 표현되면 꽃은 흰색이 된다. 그러나 유전자 A가 A로 표현되면 유전자 B가 꽃 색깔을 결정한다. 유전자 B가 BB 또는 Bb로 표현되면 꽃은 빨간색이 되고 유전자 B가 bb로 표현되면 꽃은 보라색이 된다. 여기서 유전자 A는 유전자 B에 대해 상위성을 가진다. 래브라도 리트리버의 모피 색깔은 또 다른 예시이다. 유전자 E는 모피에 색소가 있는지 여부를 결정한다. ee인 경우 모피는 노란색이 되지만 E_인 경우 두 번째 유전자 B가 색깔을 결정한다. BB와 Bb는 검은색을, bb는 갈색을 나타낸다. 따라서 유전자 e는 유전자 B에 대해 상위성을 가진다. 상위성의 또 다른 예는 닭의 볏 모양에서 나타난다. 닭은 완두 볏, 장미 볏, 호두 볏 또는 단일 볏을 가질 수 있다. 볏 모양은 두 개의 유전자 R과 P에 의해 결정된다. RRpp, Rrpp는 장미 볏을 생성하고, rrPP, rrPp는 완두 볏을 생성하고, RrPp는 호두 볏을 생성하고, rপ্প는 단일 볏을 생성한다. 여기서 유전자 R과 P는 서로에게 상위성을 가진다. |
2. 역사
집단유전학에서 다루는 상위성은 발현형질이 적응도에 미치는 영향을 중심으로 기술하며, 이는 분자생물학에서 유전자 연관의 과정을 다루는 것과는 관점의 차이가 있다. 집단유전학은 유전자 자리를 차지하는 여러 대립형질에 의해 발현된 형질이 세대에 따라 어떠한 변화를 가져오는지를 주로 관찰한다.[71]
상위성이라는 용어는 20세기 초 윌리엄 베이트슨과 그의 동료 플로렌스 더럼, 뮤리엘 윌데일 온슬로우가 처음 사용했다.[4] 유전학의 역사를 거치며 유전자 상호 작용에 대한 이해와 용어 사용은 변화해왔다. 초기 자연 선택 모델에서는 각 유전자가 다른 유전자들과는 별개로 독립적인 특성을 나타내며 적응도에 기여한다고 보았다. 일부 입문 과정에서는 여전히 이러한 방식으로 집단 유전학을 가르치기도 한다. 이 때문에 진화 유전학자들은 유전자 상호 작용을 예외적인 현상으로 여기는 경향이 있었다. 그러나 실제로는 한 대립형질의 발현은 다른 많은 대립형질들과 복잡하게 얽혀 영향을 받는다.
고전 유전학에서는 상위성을 다음과 같이 설명했다. 예를 들어, 유전자 A와 B에 각각 돌연변이가 생겨 고유한 표현형을 나타낼 때, 두 돌연변이가 함께 나타난 결과가 유전자 A 돌연변이만 있을 때와 동일한 표현형을 보인다면, 유전자 A는 유전자 B에 대해 상위성을 나타내고, 유전자 B는 열성이라고 보았다. 전체 탈모 유전자가 갈색 머리 유전자에 대해 상위성을 보이는 것이 한 예시다. 이런 의미에서 상위성은 동일한 유전자좌 내 대립유전자 간 상호작용인 유전적 우성과는 구별되는 개념이다.
유전학 연구가 발전하고 분자생물학이 등장하면서, 상위성은 양적 형질 유전자좌(QTL)나 다유전자 유전과 관련하여 연구되기 시작했다. 오늘날 유전자의 효과는 신장, 색소 침착, 성장률과 같은 표현형의 크기나, 단백질 결합 또는 효소 촉매 같은 생화학적 단백질 활성을 측정하여 정량적으로 분석하는 경우가 많다. 점점 더 정교해지는 계산 생물학 및 진화 생물학 모델들은 유전체 전체에 걸친 유전자 상호 작용의 효과와 이것이 진화에 미치는 영향을 밝히는 것을 목표로 한다.[5][6][7] 하지만 상호작용하는 유전자 쌍을 찾아내는 것은 계산적으로나 통계적으로 매우 어렵기 때문에, 연구자들은 우선순위를 정해 분석하는 방법을 모색하고 있다.[8][9]
한편, 집단유전학에서 다루는 상위성은 발현형질이 적응도에 미치는 영향을 중심으로 기술하며, 이는 분자생물학에서 다루는 유전자 연관과 같은 현상을 다른 관점에서 보는 것이다. 분자생물학이 유전자 연관의 '과정'에 초점을 맞춘다면, 집단유전학은 특정 유전자 자리를 차지하는 여러 대립형질에 의해 나타난 발현형질이 세대를 거치며 어떻게 변화하는지를 관찰한다.[71]
3. 분류
상위성에 대한 용어는 과학 분야에 따라 다를 수 있다. 유전학자들은 종종 야생형과 (주로 해로운) 돌연변이 대립 유전자를 기준으로 유전적 향상, 합성 치사, 유전적 억제 등의 개념을 사용한다. 반대로, 생화학자는 유익한 돌연변이에 더 초점을 맞추며 상호 부호 상위성, 보상 돌연변이와 같은 용어를 사용할 수 있다.[16] 이러한 차이는 생물학의 여러 분과에서 유전자 간 상호작용의 '독립성'을 다양하게 해석하기 때문에 발생하기도 한다.[17]
고전적으로 상위성은 두 좌위 간의 상호작용이 멘델의 유전법칙에서 예상되는 표현형의 분리비를 어떻게 변화시키는지를 기준으로 분류되었다. 예를 들어 다음과 같은 유형들이 있다.
과거에는 12:3:1의 분리비를 보이는 피복 유전자의 작용만을 엄밀한 의미의 상위성으로 보았으나, 현재는 유전자 간 상호작용으로 인해 멘델의 분리비를 변화시키는 모든 경우를 넓은 의미에서 상위성이라고 부른다.
3. 1. 가산성 (Additivity)
두 개의 돌연변이는 이중 돌연변이의 효과가 각각의 단일 돌연변이 효과를 합한 것과 같을 때 순수하게 가산적이라고 본다.[18][19] 이는 유전자들이 서로 상호작용하지 않을 때, 예를 들어 서로 다른 대사 경로를 통해 작용하는 경우에 발생할 수 있다. 간단히 말해, 가산적 형질은 유전학의 역사 초기에 연구되었지만, 비교적 드물게 나타나며 대부분의 유전자는 어느 정도 상호작용을 보인다.[18][19]
가산성은 여러 유전자가 동일한 효과를 내기 위해 병렬적으로 작용할 때 나타날 수 있다. 예를 들어, 생물이 인을 필요로 할 때, 환경에서 다양한 인산화된 성분을 분해하는 여러 효소가 있다면, 이 효소들은 생물이 사용할 수 있는 인의 양을 늘리기 위해 가산적으로 작용할 수 있다. 그러나 인이 더 이상 성장과 번식에 제한 요소가 아니게 되면, 인 대사를 개선하는 돌연변이는 효과가 거의 없거나 전혀 없게 된다 (음성 상호작용). 특정 유전자 내의 일부 돌연변이 집합 역시 가산적인 특성을 보이는 것으로 밝혀졌다.[28]
하지만 오늘날 엄격한 의미의 가산성은 일반적인 규칙이라기보다는 예외적인 경우로 여겨진다. 이는 대부분의 유전자가 수백 또는 수천 개의 다른 유전자와 복잡한 유전자 네트워크 내에서 상호작용하기 때문이다.[18][19]
3. 2. 크기 상위성 (Magnitude epistasis)
크기 상위성은 여러 돌연변이가 함께 존재할 때 나타나는 표현형 효과가 각 돌연변이가 개별적으로 나타내는 효과의 단순 합과 어떻게 다른지를 설명하며, 특히 적응도에 미치는 영향을 중심으로 기술된다.
이중 돌연변이가 두 개의 단일 돌연변이의 영향을 합한 것보다 더 적합한 표현형을 가질 때, 이를 양성 상위성(positive epistasis)이라고 한다.
반대로, 두 개의 돌연변이가 함께 존재할 때 각각의 효과를 합한 것보다 덜 적합한 표현형을 초래할 때, 이를 음성 상위성(negative epistasis)이라고 한다.[20][21]
돌연변이 효과의 방향성(유리함/해로움)과 관계없이, 두 돌연변이가 함께 있을 때의 전체 효과 크기가 각 단일 돌연변이 효과의 합보다 더 클 때(더 급격할 때) 상승 상위성(synergistic epistasis)이라고 한다. 반대로, 전체 효과 크기가 각 단일 돌연변이 효과의 합보다 작을 때 길항 상위성(antagonistic epistasis)이라고 한다.[15]
유전적 강화(genetic enhancement)라는 용어는 두 개의 해로운 돌연변이가 함께 존재할 때, 각각의 효과를 단순히 더한 것보다 훨씬 더 심각한 표현형을 나타내는 경우(즉, 강한 음성/상승 상위성)에 사용되기도 한다.
한편, 강한 양성 상위성(특히 여러 요소가 동시에 존재해야만 기능이 발현되는 경우)은 때때로 창조론자들에 의해 환원 불가능한 복잡성의 근거로 제시되기도 하지만, 과학계에서는 이러한 주장의 대부분의 예시가 타당하지 않다고 본다.
3. 3. 부호 상위성 (Sign epistasis)
'''부호 상위성'''(sign epistasis) 또는 '''부호적 상호작용'''[22]은 특정 돌연변이의 효과가 다른 돌연변이의 존재 여부에 따라 정반대로 나타나는 현상을 말한다. 예를 들어, 단독으로는 해로운 돌연변이가 특정 유익한 돌연변이와 함께 존재할 때 오히려 그 효과를 향상시키는 경우가 이에 해당한다.[17] 가령, 발달된 감각 기관이 없다면 크고 복잡한 뇌는 에너지만 소모하는 비효율적인 기관일 수 있다. 하지만 뛰어난 감각 기관이 있다면, 크고 복잡한 뇌는 감각 정보를 효과적으로 처리하여 생존에 더 유리하게 작용할 수 있다. 만약 적합도 풍경(fitness landscape)에 이러한 부호적 상호작용이 없다면, 그 풍경은 '매끄럽다'(smooth)고 표현한다.
가장 극단적인 형태의 부호 상위성은 '''상호 부호적 상호작용'''(reciprocal sign epistasis)[23]이다. 이는 각각 해로운 두 개의 돌연변이가 함께 존재할 때 오히려 유익한 효과를 나타내는 경우이다. 예를 들어, 세균이 독소만 생산하면 스스로에게 해로울 수 있고, 독소 배출 시스템만 가지고 있으면 에너지를 낭비하게 된다. 하지만 독소 생산 능력과 배출 시스템을 모두 갖추면, 주변의 경쟁 생물체를 제거하여 자신의 적합도를 높일 수 있다. 적합도 풍경에 부호적 상호작용은 존재하지만 상호 부호적 상호작용이 없다면, 그 풍경은 '반매끄럽다'(semi-smooth)고 한다.[24]
상호 부호적 상호작용은 '''유전적 억제'''(genetic suppression)로 이어지기도 한다. 이는 두 개의 해로운 돌연변이가 함께 있을 때, 각각 단독으로 있을 때보다 해로운 정도가 덜해지는 현상이다. 즉, 하나의 돌연변이가 다른 돌연변이의 해로운 효과를 보상하는 것이다. 박테리오파지 T4의 조립 과정에서 이러한 유전적 억제의 명확한 예를 찾아볼 수 있다. 각각 다른 형태 발생 단백질의 양을 비정상적으로 만드는 두 개의 해로운 돌연변이가 함께 존재할 때, 오히려 긍정적인 상호작용을 일으킬 수 있다.[25] 만약 한 돌연변이가 특정 구조 단백질의 양을 감소시켜 바이러스 입자 조립의 불균형을 초래하고 생존 가능한 자손 바이러스 생성을 방해한다면, 다른 단백질의 양을 조절하는 두 번째 (억제) 돌연변이가 단백질 구성 요소 간의 균형을 회복시켜 생존 가능한 자손 바이러스 생산을 가능하게 할 수 있다.
유전적 억제라는 용어는 두 돌연변이가 함께 있을 때 단일 돌연변이 상태의 중간 정도 표현형을 나타내는 부호적 상호작용에도 사용된다. 이 경우, 더 심각한 표현형을 나타내는 단일 돌연변이의 효과가 다른 돌연변이나 유전적 조건에 의해 억제되는 것이다. 예를 들어, 이배체 생물에서 기능이 감소된(부분적 기능 상실) 돌연변이의 표현형은, 동일한 생화학 경로에서 반대 작용을 하는 유전자의 복사본 하나를 제거함으로써 완화될 수 있다. 이때 두 번째 유전자는 기능 감소 돌연변이의 '우성 억제자'라고 불린다. '우성'이라고 하는 이유는 억제 유전자의 정상(야생형) 복사본이 하나만 존재해도(즉, 이형 접합 상태에서도) 억제 효과가 나타나기 때문이다. 대부분의 유전자는 하나의 복사본만으로도 정상 기능을 하므로(반수체 불충분성이 드물기 때문에), 이형 접합 상태의 억제 돌연변이 자체는 정상 표현형을 보인다. 따라서 두 돌연변이가 함께 있는 상태(억제된 상태)의 표현형은 단일 돌연변이 상태의 중간 정도가 된다.
비상호 부호적 상호작용(non-reciprocal sign epistasis)에서는 돌연변이의 적합도 효과가 상호 부호적 상호작용에서 나타나는 극단적인 반전보다는 중간 정도의 변화를 보인다.
두 개의 돌연변이가 각각 단독으로는 생존에 문제가 없지만, 함께 존재할 때 치명적인 결과를 초래하는 경우를 '''합성 치사성'''(synthetic lethality) 또는 '''비연관 비상보성'''(unlinked non-complementation)이라고 한다.[26]
3. 4. 반수체 생물에서의 상위성 (Haploid organisms)
반수체 유기체에서 유전자형이 (두 개의 유전자 좌위에서) ''ab'', ''Ab'', ''aB'' 또는 ''AB''인 경우, 개별적으로 (Ab와 aB) 또는 조합(AB)으로 돌연변이가 표현형의 크기에 영향을 미치는 다양한 형태의 상위성을 생각해 볼 수 있다.
| 상호 작용 유형 | ab | Ab | aB | AB | 설명 |
| 상위성 없음 (가산) | 0 | 1 | 1 | 2 | AB = Ab + aB + ab |
| 긍정적(상승적) 상위성 | 0 | 1 | 1 | 3 | AB > Ab + aB + ab |
| 부정적(길항적) 상위성 | 0 | 1 | 1 | 1 | AB < Ab + aB + ab |
| 부호 상위성 | 0 | 1 | -1 | 2 | AB는 Ab 또는 aB와 반대 부호를 가짐 |
| 상호 부호 상위성 | 0 | -1 | -1 | 2 | AB는 Ab 그리고 aB와 반대 부호를 가짐 |
3. 5. 이배체 생물에서의 상위성 (Diploid organisms)
이배체 생물에서의 상위성은 각 유전자의 두 복제본이 존재하기 때문에 더욱 복잡해진다. 상위성은 유전자좌 사이에서 발생할 수 있을 뿐만 아니라, 이형접합체에서 각 유전자좌의 두 복제본 사이에서도 상호작용이 일어날 수 있다. 2개의 유전자좌, 2개의 대립유전자 체계의 경우, 8가지 독립적인 유형의 유전자 상호작용이 있다.[27]
| 가산적 A 유전자좌 | 가산적 B 유전자좌 | 우성적 A 유전자좌 | 우성적 B 유전자좌 | ||||||||||||||||||||||||||||||||
| aa | aA | AA | aa | aA | AA | aa | aA | AA | aa | aA | AA | ||||||||||||||||||||||||
| bb | 1 | 0 | –1 | bb | 1 | 1 | 1 | bb | –1 | 1 | –1 | bb | –1 | –1 | –1 | ||||||||||||||||||||
| bB | 1 | 0 | –1 | bB | 0 | 0 | 0 | bB | –1 | 1 | –1 | bB | 1 | 1 | 1 | ||||||||||||||||||||
| BB | 1 | 0 | –1 | BB | –1 | –1 | –1 | BB | –1 | 1 | –1 | BB | –1 | –1 | –1 | ||||||||||||||||||||
| 가산적-가산적 상위성 | 가산적-우성적 상위성 | 우성적-가산적 상위성 | 우성적-우성적 상위성 | ||||||||||||||||||||||||||||||||
| aa | aA | AA | aa | aA | AA | aa | aA | AA | aa | aA | AA | ||||||||||||||||||||||||
| bb | 1 | 0 | –1 | bb | 1 | 0 | –1 | bb | 1 | –1 | 1 | bb | –1 | 1 | –1 | ||||||||||||||||||||
| bB | 0 | 0 | 0 | bB | –1 | 0 | 1 | bB | 0 | 0 | 0 | bB | 1 | –1 | 1 | ||||||||||||||||||||
| BB | –1 | 0 | 1 | BB | 1 | 0 | –1 | BB | –1 | 1 | –1 | BB | –1 | 1 | –1 | ||||||||||||||||||||
생쥐의 털색은 여러 유전자에 의해 결정되는 상위성의 예시이다. 자연 상태의 생쥐는 각 털에 무늬가 있어 전체적으로 회색을 띈다. 생쥐 털색에 관여하는 주요 유전자는 다음과 같다.
| 염색체 위치 | 유전자 | 역할 | 발현 특징 |
|---|---|---|---|
| A | AA | 색소 형성을 지시한다 | 우성 동형접합 유전자 |
| Aa | 색소 형성을 지시한다 | 우성 대립형질 | |
| aa | 색소 형성을 막는다 | 열성 동형접합 유전자 | |
| B | BB | 회색의 무늬를 만든다 | 우성 동형접합 유전자 |
| Bb | 회색의 무늬를 만든다 | 우성 대립 유전자 | |
| bb | 무늬형성을 억제하여 검은색이 되게 한다 | 열성 동형접합 유전자 |
유전형이 AABB인 회색 생쥐와 aabb인 흰색 생쥐를 교배하면, 첫 번째 자식 세대(F1)의 유전형은 모두 AaBb가 되어 회색 털을 갖게 된다. 이 F1 세대를 다시 교배하여 얻은 두 번째 자식 세대(F2)에서는 털색의 발현 비율이 다음 표와 같이 9(회색):3(검은색):4(흰색)으로 나타난다.
| X | AB | Ab | ab | aB |
|---|---|---|---|---|
| AB | AA BB | AA Bb | Aa Bb | Aa BB |
| Ab | AA Bb | AA bb | Aa bb | Aa Bb |
| ab | Aa Bb | Aa bb | aa bb | aa Bb |
| aB | Aa BB | Aa Bb | aa Bb | aa BB |
이 결과는 색소 형성을 억제하는 열성 유전형질 aa가 털 무늬를 결정하는 B-b 대립유전자보다 상위에서 작용함을 보여준다. 즉, A-a 대립형질에 의해 조절되는 효소가 먼저 발현되어 색소 형성 여부를 결정한 후, B-b 대립형질에 관련된 효소가 털색 발현에 관여하기 때문이다.[72] 이러한 상위성 때문에 해당 대립형질들은 멘델의 유전법칙 중 독립의 법칙을 따르지 않는다.
단순한 예로, 두 좌위 간의 상호 작용 (상위성)을 생각해 보자. 하나의 좌위에 A와 a의 대립 유전자가 있고, 다른 좌위에 B와 b의 대립 유전자가 있다고 가정한다. A는 a에 대해 우성이며, B는 b에 대해 우성이라고 가정한다. AAbb와 aaBB를 교배한 잡종 1세대는 AaBb가 되며, 2세대는 AB:Ab:aB:ab = 9:3:3:1의 비율로 나타난다 (단순화를 위해 동일한 좌위에서 가장 우성인 유전자만 표시). 상위성이 있으면 관찰되는 분리비가 다음과 같이 변화한다.
- '''피복 유전자''' (dominant epistasis): A의 형질이 B의 형질을 덮어 가릴 때, AB와 Ab의 형질은 구분할 수 없으며, 분리비는 (AB+Ab):aB:ab = 12:3:1이 된다. 호박의 과피색 유전이 이에 해당한다.
- '''조건 유전자''' (recessive epistasis): A가 있을 때만 B가 발현되는 경우 (A가 없으면 B가 발현되지 않음), aB와 ab의 형질은 구분할 수 없으며, 분리비는 AB:Ab:(aB+ab) = 9:3:4가 된다. 양파의 색 유전이 이에 해당한다.
- '''보충 유전자''' (complementary genes): A와 a, B와 b 사이에 우열 관계가 없고, A와 B가 함께 존재할 때만 다른 형질이 나타나는 경우, 분리비는 AB:(Ab+aB+ab) = 9:7이 된다. 스위트피의 꽃 색깔이 이에 해당한다.
- '''억제 유전자''' (inhibitory gene): 겉으로 드러나는 형질은 B와 b뿐이며, A가 B의 발현을 억제하는 경우, 분리비는 (AB+Ab+ab):aB = 13:3이 된다. 누에나방의 고치 색깔이 이에 해당한다.
- '''동의 유전자''' (duplicate genes): A와 B가 동일한 형질을 발현하는 경우, 분리비는 (AB+Ab+aB):ab = 15:1이 된다. 황새냉이의 열매 모양이 이에 해당한다.
과거에는 12:3:1의 분리비를 보이는 피복 유전자의 작용만을 엄밀한 의미의 상위성으로 보았으나, 현재는 유전자 간 상호작용으로 인해 멘델의 분리비를 변화시키는 모든 경우를 넓은 의미에서 상위성이라고 부른다.
4. 유전적 및 분자적 원인
유전자 상호 작용에 대한 이해는 유전학의 역사를 통해 상당히 변화해 왔으며, 용어의 사용도 마찬가지이다. 상위성이라는 용어는 윌리엄 베이트슨과 그의 협력자 플로렌스 더럼과 뮤리엘 윌데일 온슬로우에 의해 처음 사용되었다.[4] 20세기 초에 고안된 자연 선택의 초기 모델에서는 각 유전자가 다른 유전자들의 평균적인 배경 속에서 독립적으로 작용하여 적응도에 기여하는 것으로 간주되었다. 일부 입문 과정에서는 여전히 이러한 방식으로 집단 유전학을 가르치기도 한다. 이러한 역사적 배경 때문에 진화 유전학자들은 유전자 상호 작용을 예외적인 현상으로 여기는 경향이 있었으나, 실제로는 어떤 하나의 대립유전자의 발현은 다른 많은 대립유전자들에 복잡하게 의존하는 것이 일반적이다.
고전 유전학에서는 유전자 A와 B가 각각 돌연변이되어 고유한 표현형을 나타낼 때, 두 돌연변이가 함께 존재함에도 유전자 A 돌연변이와 동일한 표현형을 보인다면 유전자 A가 유전자 B에 대해 상위성(epistasis)을 나타낸다고 보았다. 예를 들어, 전체 탈모 유전자는 갈색 머리 유전자에 대해 상위성을 나타낸다. 이러한 의미에서 상위성은 동일한 유전자좌 내 대립유전자 간의 상호 작용인 유전적 우성과는 구별된다. 유전학 연구가 발전하고 분자 생물학이 등장하면서, 상위성은 양적 형질 유전자좌(QTL) 및 다유전자 유전과 관련하여 더욱 깊이 연구되기 시작했다.
현대에 이르러 유전자의 효과는 표현형의 크기(예: 신장, 색소 침착, 성장률)나 생화학적 분석(예: 결합 또는 효소 촉매 활성)을 통해 정량적으로 측정된다. 점점 더 정교해지는 계산 생물학 및 진화 생물학 모델들은 유전체 전체에서 나타나는 상위성의 효과와 이것이 진화에 미치는 영향을 설명하고자 한다.[5][6][7] 하지만 유전자 상호 작용 쌍을 식별하는 것은 계산 및 통계적으로 여전히 어려운 과제이며, 일부 연구에서는 이러한 상호 작용 쌍의 우선순위를 정하는 방법을 개발하고 있다.[8][9]
유전자 간의 상호작용은 단순히 효과가 더해지는 가산적인 경우 외에도 다양한 방식으로 나타난다. 두 돌연변이가 함께 있을 때 예상보다 더 적합하거나 덜 적합한 표현형을 보이는 양성 상위성과 음성 상위성, 효과의 변화 폭이 예상보다 크거나 작은 상승 상위성과 길항 상위성, 한 돌연변이가 다른 돌연변이의 효과 방향을 바꾸는 부호적 상호작용, 해로운 돌연변이들이 함께 있을 때 오히려 유익해지는 상호 부호적 상호작용, 그리고 두 돌연변이가 각각은 생존 가능하지만 함께 존재하면 치명적인 합성 치사성 등 다양한 형태의 상위성이 존재한다.[10][11][13][20][21][12][15][22][17][23][26] 이러한 다양한 상호작용은 유전자의 기능과 진화 과정을 이해하는 데 중요한 열쇠가 된다.
4. 1. 유전자 간 상위성 (Epistasis between genes)
집단유전학에서 다루는 상위성은 발현형질이 적응도에 미치는 영향을 관찰한 결과를 중심으로 기술한 것으로, 분자생물학에서 다루는 유전자 연관과 동일한 현상이다. 그러나 분자생물학이 유전자 연관의 과정을 다루는 것에 비해 집단유전학은 유전자 자리를 차지하는 여러 대립형질에 의해 발현된 발현형질이 세대에 따라 어떠한 변화를 가져오는지를 관찰한다.[71]
생쥐의 털색은 여러 유전자에 의해 결정된다. 자연의 생쥐는 각 털에 무늬가 있어 전체적으로 보아 회색을 띈다. 생쥐의 털색에 관여하는 유전자는 다음의 표와 같다.
| 염색체 위치 | 유전자 | 역할 | 발현 특징 |
|---|---|---|---|
| A | AA | 색소 형성을 지시한다 | 우성 동형접합 유전자 |
| Aa | 색소 형성을 지시한다 | 우성 대립유전자 | |
| aa | 색소 형성을 막는다 | 열성 동형접합 유전자 | |
| B | BB | 회색의 무늬를 만든다 | 우성 동형접합 유전자 |
| Bb | 회색의 무늬를 만든다 | 우성 대립 유전자 | |
| bb | 무늬형성을 억제하여 검은색이 되게 한다 | 열성 동형접합 유전자 |
이제 유전형이 AABB인 회색 생쥐와 aabb인 흰색 생쥐를 교배하면 첫 번째 자식 세대 F1의 유전형은 AaBb로 회색 생쥐가 된다. F1을 다시 교배한 자식 세대 F2의 털색 발현 비율은 다음의 표와 같이 9(회색):3(검은색):4(흰색) 가 된다.
| X | AB | Ab | ab | aB |
|---|---|---|---|---|
| AB | AA BB | AA Bb | Aa Bb | Aa BB |
| Ab | AA Bb | AA bb | Aa bb | Aa Bb |
| ab | Aa Bb | Aa bb | aa bb | aa Bb |
| aB | Aa BB | Aa Bb | aa Bb | aa BB |
색소 형성을 억제하는 열성유전형질 aa가 B-b 대립유전자보다 상위에서 작용하는 것을 확인할 수 있다. 이는 A-a 대립형질에 의한 효소가 먼저 발현된 후 B-b의 털색 발현에 관여하는 효소가 발현하기 때문이다.[72] 이러한 이유로 상위성을 보이는 대립형질에서는 멘델의 유전법칙 가운데 독립의 법칙이 지켜지지 않는다.
이는 여러 유전자가 동일한 효과를 달성하기 위해 병렬적으로 작용할 때 발생할 수 있다. 예를 들어, 유기체가 인을 필요로 할 때, 환경에서 다양한 인산화된 성분을 분해하는 여러 효소가 유기체에 사용 가능한 인의 양을 증가시키기 위해 가산적으로 작용할 수 있다. 그러나 인이 더 이상 성장과 번식의 제한 요소가 아닌 시점이 불가피하게 발생하며, 따라서 인 대사 개선은 작거나 또는 전혀 효과가 없다(음성 상호작용). 유전자 내의 일부 돌연변이 집합도 특별히 가산적인 것으로 밝혀졌다.[28] 이제 엄격한 가산성은 규칙이라기보다는 예외로 간주된다. 이는 대부분의 유전자가 수백 또는 수천 개의 다른 유전자와 상호 작용하기 때문이다.[18][19]
생물체의 유전체 내에서 상위성은 유전체 내 유전자 간의 상호 작용으로 인해 발생한다. 이러한 상호 작용은 유전자가 예를 들어 다성분 단백질(예: 리보솜)의 별도 구성 요소이거나, 서로의 활성을 억제하거나, 한 유전자가 암호화한 단백질이 다른 유전자를 수정하는 경우(예: 인산화) 직접적으로 발생할 수 있다. 또는 유전자가 대사 경로 또는 네트워크, 발달 경로, 신호 전달 경로 또는 전사 인자 네트워크의 구성 요소를 암호화하는 경우 간접적으로 발생할 수 있다. 예를 들어, 페니실린을 합성하는 효소를 암호화하는 유전자는 대사 경로에서 필요한 전구체를 합성하는 효소가 없으면 곰팡이에게 아무 쓸모가 없다.
단순한 예로, 두 좌위 간의 상호 작용 (상위성)을 생각해 보자. 하나의 좌위에 A와 a의 대립 유전자가 있고, 다른 좌위에 B와 b의 대립 유전자가 있다고 가정한다. A는 a에 대해 우성이며, B는 b에 대해 우성이라고 가정한다. AAbb와 aaBB를 교배한 잡종 1세대는 AaBb가 되며, 2세대는 AB:Ab:aB:ab = 9:3:3:1이 된다 (단순화를 위해 동일한 좌위에서 가장 우성인 유전자만 표시). 상위성이 있으면 관찰되는 분리비가 변화한다.
;피복 유전자
: A의 형질이 B의 형질을 덮어 가릴 때, AB와 Ab의 형질은 구분할 수 없으며, (AB+Ab):aB:ab = 12:3:1이 된다. 이를 '''우성 상위'''(dominant epistasis)라고 한다. 이에 해당하는 예로 호박의 과피색 유전이 있다.
;조건 유전자
: A가 있을 때만 B가 발현되는 (A가 없을 때에는 B가 발현되지 않는) 경우, aB와 ab의 형질은 구분할 수 없으며, AB:Ab:(aB+ab) = 9:3:4가 된다. 이를 '''열성 상위'''(recessive epistasis)라고 한다. 양파의 색 유전이 이에 해당한다.
;보충 유전자
: A와 a, B와 b에는 우열이 없고, A와 B가 조합되었을 때만 다른 형질이 나타나는 경우, AB:(Ab+aB+ab) = 9:7이 된다. 스위트피의 꽃 색깔이 이에 해당한다.
;억제 유전자
: 겉으로 나타나는 형질은 B와 b뿐이며, A가 B의 발현을 억제하는 유전자인 경우, (AB+Ab+ab):aB = 13:3이 된다. 누에나방의 고치 색깔이 이에 해당한다.
;동의 유전자
: A와 B가 하나의 동일한 형질을 발현하는 경우, (AB+Ab+aB):ab = 15:1이 된다. 황새냉이의 열매 모양이 이에 해당한다.
엄밀한 의미에서 12:3:1이 되는 피복 유전자의 작용만이 상위성이라고 불렸지만, 현재는 분리비를 바꾸는 상호 작용은 모두 상위성이라고 불린다.
4. 2. 유전자 내 상위성 (Epistasis within genes)
두 개의 서로 다른 유전자에서 발생한 돌연변이가 상호작용하여 그 효과가 단순히 더해지지 않는 것처럼, 하나의 유전자 내 다른 위치(코돈)에서 발생한 돌연변이들도 서로 영향을 주고받으며 효과가 가산되지 않을 수 있다. 유전학에서는 이러한 현상을 유전자 내 억제(Intragenic suppression)라고 부르기도 하는데, 이는 한 유전자 내에서 발생한 해로운 돌연변이의 효과가 같은 유전자 내의 다른 위치에 발생한 두 번째 돌연변이에 의해 상쇄되거나 완화되는 경우를 말한다.
세균에 감염하는 바이러스인 세균 파지 T4의 rIIB 유전자에 변형이 생긴 돌연변이체들을 분석한 결과, 특정 돌연변이 쌍들은 서로의 효과를 억제할 수 있다는 사실이 밝혀졌다. 즉, 두 개의 돌연변이를 함께 가진 이중 돌연변이체가 각각의 단일 돌연변이체보다 오히려 야생형에 가까운 표현형을 나타내는 것이다.[29] 연구자들은 유전자 재조합 데이터를 이용하여 돌연변이들의 상대적인 위치를 파악했고, 이러한 정보들을 종합하여 1961년 유전 암호가 세 개의 염기서열로 이루어진다는 특성(3중항 특성)과 다른 주요 특징들을 처음으로 논리적으로 추론해낼 수 있었다.[1]
유전자 내 억제는 해당 유전자가 만드는 단백질 내부의 아미노산들이 서로 상호작용할 때도 발생할 수 있다. 단백질이 접혀서 특정 구조를 형성하고 기능을 수행하는 과정은 매우 복잡하기 때문에, 돌연변이의 효과가 단순히 더해지는 경우는 드물다.
단백질은 내부에 분산된 여러 협력적인 상호작용(소수성, 극성, 공유 결합 등) 네트워크를 통해 안정적인 3차 구조를 유지한다.[30] 어떤 돌연변이가 발생하여 다른 아미노산 잔기의 주변 환경을 변화시키면(직접 접촉하거나 단백질 구조의 변화를 유도하여) 상위성 상호 작용이 일어난다.[31] 예를 들어, 단백질의 안정성을 높이는 이황화 결합을 형성하는 두 개의 시스테인 아미노산을 생각해보자. 만약 돌연변이로 시스테인 하나만 존재한다면, 정확한 위치에 두 번째 시스테인이 있어 결합을 형성하지 않는 한 단백질 안정성에 거의 영향을 미치지 못한다. 하지만 두 개의 시스테인이 모두 존재하여 이황화 결합을 형성하면 단백질의 안정성은 크게 향상된다.[32] 이는 두 개의 시스테인 돌연변이를 가진 경우가 각각의 단일 시스테인 돌연변이보다 훨씬 더 높은 안정성을 보이는 긍정적 상위성(positive epistasis)으로 관찰된다.
반대로, 단백질은 해로운 돌연변이가 도입되었을 때 종종 돌연변이 강건성(mutational robustness)을 나타낸다. 이는 단백질을 안정화시키는 상호작용이 일부 파괴되더라도 특정 안정성 임계점에 도달하기 전까지는 기능을 유지하는 현상이다. 그러나 이 임계점을 넘어 추가적인 불안정화 돌연변이가 발생하면 단백질은 더 이상 제대로 접힐 수 없게 되어 기능에 매우 큰 손상을 입는다. 이는 각각의 돌연변이만으로는 거의 영향이 없지만, 함께 존재할 때 큰 해로운 영향을 미치는 부정적 상위성(negative epistasis)으로 나타난다.[33][41]
효소의 경우, 단백질 구조는 몇 개의 핵심적인 아미노산들을 정확한 기하학적 위치에 배열하여 화학 반응을 수행하는 활성 부위를 형성한다.[34] 활성 부위의 기능은 여러 구성 요소의 협력을 필요로 하는 경우가 많기 때문에, 이들 중 하나라도 돌연변이로 인해 제 기능을 못하게 되면 효소 활성이 크게 저하된다. 따라서 이미 비활성화된 효소에 두 번째 구성 요소의 돌연변이가 추가로 발생하더라도 그 영향은 비교적 작다. 예를 들어, 많은 효소에서 발견되는 촉매 삼합체(catalytic triad)의 구성 아미노산 중 하나를 제거하면 효소 활성이 생존 불가능할 정도로 낮아지는 경우가 많다.[35][36][37]
4. 2. 1. 이형접합체 상위성 (Heterozygotic epistasis)
배수성 생물은 각 유전자의 두 개의 복사본을 가지고 있다. 이들이 서로 다를 경우(이형접합성), 대립유전자의 두 가지 다른 복사본이 서로 상호작용하여 상위성을 유발할 수 있다. 이것은 때때로 '''대립유전자 상보''' 또는 '''유전자간 상보'''라고 불린다.이러한 상호작용은 여러 메커니즘을 통해 발생할 수 있다. 예를 들어, 한 대립유전자에서 나온 인핸서가 다른 대립유전자의 프로모터로부터 전사를 활성화시키는 트랜스벡션 현상이 있다. 또한, 기능하지 않는 두 개의 RNA 분자가 트랜스-스플라이싱 과정을 통해 결합하여 하나의 기능성 RNA를 생성할 수도 있다.
단백질 수준에서도 유사한 상호작용이 나타난다. 이합체로 기능하는 단백질의 경우, 각기 다른 대립유전자에서 만들어진 단백질 단위체가 결합하여 이종이합체를 형성할 수 있다. 이 이종이합체는 하나의 대립유전자에서만 만들어진 단백질로 구성된 동종이합체와는 다른 특성을 보일 수 있다.
박테리오파지 T4 돌연변이 연구는 이러한 대립유전자 상보의 좋은 예를 보여준다.[1] 동일한 유전자 내 다른 위치에 결함이 있는 두 종류의 돌연변이 파지를 함께 감염시키면, 각각의 돌연변이 파지만으로는 생존 가능한 자손을 만들 수 없지만, 두 돌연변이가 상보적으로 작용하여 생존 가능한 자손 파지를 만들어낼 수 있다. 이러한 유전자 내 상보 현상은 박테리오파지의 구조 단백질을 암호화하는 여러 유전자에서 확인되었으며[38], 이는 해당 단백질들이 이합체나 더 복잡한 다량체 형태로 기능한다는 것을 시사한다.[39]
5. 진화적 결과
(내용 없음 - 해당 섹션의 내용은 하위 섹션에서 상세히 다루므로 중복을 피하기 위해 생략함)
5. 1. 적합도 지형과 진화 가능성 (Fitness landscapes and evolvability)
집단유전학에서 다루는 상위성은 발현형질이 적응도에 미치는 영향을 관찰한 결과를 중심으로 기술한 것으로, 분자생물학에서 다루는 유전자 연관과 동일한 현상이다. 그러나 분자생물학이 유전자 연관의 과정을 다루는 것에 비해 집단유전학은 유전자 자리를 차지하는 여러 대립형질에 의해 발현된 발현형질이 세대에 따라 어떠한 변화를 가져오는지를 관찰한다.[71]
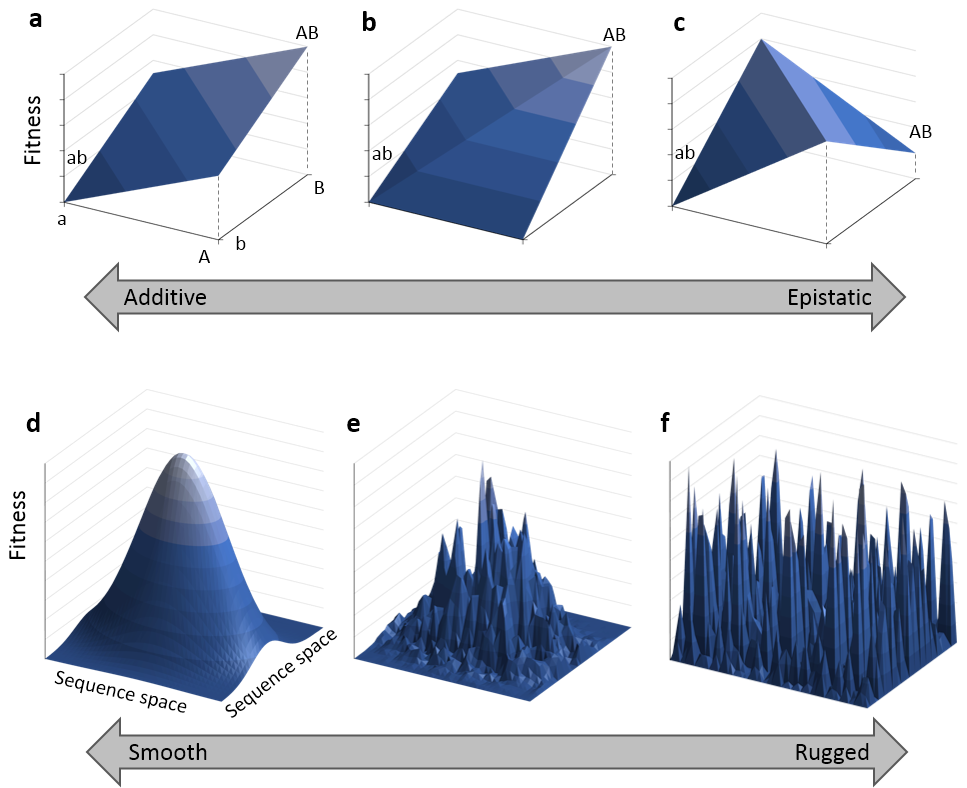
진화 유전학에서 상호작용의 부호는 일반적으로 상호작용의 크기보다 더 중요하게 여겨진다. 상호작용의 크기(양성 및 음성)는 유익한 돌연변이의 수에 영향을 미치지만, 상호작용의 부호는 돌연변이 조합이 유익한지 해로운지를 결정하기 때문이다.[10]
피트니스 랜드스케이프(적합도 지형)는 모든 유전자형을 2차원 공간에 배열하고 각 유전자형의 적합도를 표면의 높이로 표현한 것이다. 이는 한 유전자형에서 다음 유전자형으로, 즉 더 적합한 유전자형으로 이동하는 과정으로서 진화를 이해하기 위한 시각적 비유로 자주 사용된다.[18]
모든 돌연변이가 가산적이라면, 즉 각 돌연변이의 효과가 다른 돌연변이의 존재 여부와 관계없이 일정하다면, 돌연변이는 어떤 순서로든 획득될 수 있으며 지속적으로 적합도를 높이는 경로를 제공한다. 이 경우 적합도 지형은 완벽하게 부드럽고, 단 하나의 봉우리(전역 최댓값)만 존재하며, 모든 염기 서열은 유익한 돌연변이를 축적하며 '어떤 순서로든' 이 봉우리를 향해 진화할 수 있다. 반대로, 돌연변이가 상호작용을 통해 서로 영향을 주고받으면, 특정 돌연변이의 효과가 다른 돌연변이의 유전적 배경에 따라 달라지면서 적합도 지형은 거칠어진다.[40] 가장 극단적인 경우, 상호작용이 매우 복잡해지면 적합성은 유전자 서열과 거의 상관관계가 없어지고 지형의 모습은 무작위적이 된다. 이를 거친 적합도 지형이라고 하며, 이는 유기체의 진화적 최적화에 심각한 영향을 미친다.
거친 지형에서는 특정 돌연변이가 한 조합에서는 해롭지만 다른 조합에서는 유익할 수 있다. 이 때문에 가장 적합한 유전자형에 도달하기 위해서는 돌연변이를 '특정한 순서'로 축적해야만 한다. 이는 유기체가 '잘못된' 순서로 돌연변이를 획득하여 지역 최댓값에 갇힐 가능성을 높인다.[41][42] 예를 들어, 5개의 특정 돌연변이를 가진 TEM1 β-락타마제 변종은 3세대 항생제인 세포탁심을 분해할 수 있다.[43] 그러나 이 5개의 돌연변이를 얻는 120가지 가능한 경로 중 단 7%만이 실제로 진화 과정에서 선택될 수 있다. 나머지 경로는 중간 단계의 돌연변이 조합이 오히려 효소 활성을 감소시키는 '피트니스 계곡'(fitness valley)을 통과해야 하기 때문이다.
하지만 환경 변화, 즉 적합도 지형의 변화는 유기체가 지역 최댓값에서 벗어날 기회를 제공하기도 한다.[41] 앞선 β-락타마제 예시에서, 변화하는 항생제 환경에서의 선택은 진화 경로상에서 다른 돌연변이들과 긍정적으로 상호작용하는 "관문 돌연변이"(gateway mutation)의 등장을 유도했다. 이 관문 돌연변이는 개별적으로는 유익하지만 함께 있을 때 부정적 상호작용을 일으키던 다른 돌연변이들의 효과를 완화시켜, 결과적으로 피트니스 계곡을 건너 더 높은 적합도를 가진 변종으로 진화할 수 있게 했다. 이처럼 복잡한 환경이나 선택 압력은 단순한 양성 선택 모델에서 예측되는 지역 최댓값 문제를 우회할 수 있게 한다.
높은 수준의 상호작용은 일반적으로 진화의 제약 요인으로 간주된다. 상호작용이 많은 형질은 개선되기가 어려워 진화 가능성(evolvability)이 낮은 것으로 여겨진다. 이는 특정 유전적 배경에서는 유익한 돌연변이가 매우 드물기 때문이다. 부드러운 지형이 없다면 진화가 적합도 봉우리에 도달하기 어려워진다. 매우 거친 지형에서는 피트니스 계곡이 특정 유전자형으로의 접근을 차단하며, 접근을 허용하는 능선이 존재하더라도 드물거나 지나치게 길 수 있다.[44] 더욱이, 적응 과정 자체가 단백질을 적합도 지형의 더 불안정하거나 거친 영역으로 이동시킬 수도 있다.[45] 이러한 "적합도 영역"의 변화는 진화를 늦추는 요인이 될 수 있으며, 적응된 형질 간의 상충 관계(trade-off)를 나타낼 수 있다.
거친 적합도 지형이 적응 진화를 방해하는 것처럼 보이지만, 오히려 이것이 진화 가능성 자체의 진화를 이끄는 잠재적인 힘이 될 수 있다는 주장도 제기되었다. 마이클 콘래드는 1972년, 다른 유전자 자리에서 발생하여 적합도 지형을 부드럽게 만드는 돌연변이가 유익한 돌연변이의 생성을 촉진하고, 이들과 함께 히치하이킹하며 퍼져나갈 수 있다고 제안하며 진화 가능성의 진화 메커니즘을 처음 제시했다.[46][47] 루퍼트 리들은 1975년에 상호 부호 상호작용(reciprocal sign epistasis)을 보이는 유전자들이, 단일 돌연변이로는 나타나기 어려운 표현형을 달성하는 새로운 경로를 제공할 수 있다고 주장했다.[48][49]
거친 상호작용 적합도 지형은 또한 진화의 궤적에 영향을 미친다. 돌연변이가 많은 다른 돌연변이들과 상호작용할 때, 새로 축적되는 각 돌연변이는 이후에 나타날 수 있는 유익한 돌연변이의 종류를 극적으로 변화시킨다. 따라서 진화가 따르는 경로는 초기에 어떤 돌연변이가 고정되었는지에 크게 의존하게 된다. 결과적으로, 동일한 시작점에서 진화가 반복되더라도 부드럽고 가산적인 지형에서처럼 단일 전역 최댓값으로 수렴하기보다는, 서로 다른 지역 최댓값으로 분기하는 경향을 보인다.[50][51]
5. 2. 성의 진화 (Evolution of sex)
음의 상위성은 성(sex)과 밀접하게 연관되어 있다고 여겨진다. 이 아이디어는 무성 생식 집단과 유성 생식 집단의 디지털 시뮬레이션을 사용하여 실험적으로 검증되었다. 시간이 지남에 따라 유성 생식 집단은 더욱 음의 상위성, 즉 두 상호작용하는 대립 유전자에 의한 적합도 저하를 향해 이동하는 경향을 보였다. 음의 상위성은 상호작용하는 해로운 돌연변이를 가진 개체가 집단에서 효율적으로 제거되도록 하는 메커니즘으로 이해된다. 이를 통해 해당 대립 유전자가 집단에서 제거되어 전체적으로 더 적합한 집단을 만든다고 설명한다. 이 가설은 알렉세이 콘드라쇼프에 의해 제안되었으며, 때때로 결정론적 돌연변이 가설[52]이라고 불린다. 인공 유전자 네트워크를 사용한 테스트도 이루어졌다.[20]그러나 이 가설에 대한 증거가 항상 명확한 것은 아니며, 콘드라쇼프가 제안한 모델이 실제 관찰과는 다른 돌연변이 매개변수를 가정한다는 비판도 제기되었다.[53] 또한, 인공 유전자 네트워크를 사용한 테스트에서는 음의 상위성이 더 조밀하게 연결된 네트워크에서만 나타나는 경향을 보였으나,[20] 경험적 증거에 따르면 자연의 유전자 네트워크는 희소하게 연결되어 있다.[54] 이론적으로는 견고성을 위한 선택이 더 희소하게 연결되고 최소한으로 복잡한 네트워크를 선호하는 것으로 나타났다.[54]
6. 연구 방법 및 모델 시스템
집단유전학에서는 상위성을 특정 형질이 생물의 적응도에 미치는 영향을 중심으로 기술한다. 이는 분자생물학에서 다루는 유전자 연관과 동일한 현상이지만, 접근 방식에 차이가 있다. 분자생물학이 유전자 연관의 과정을 주로 다루는 반면, 집단유전학은 특정 유전자 자리를 차지하는 여러 대립형질에 의해 나타나는 형질이 세대에 걸쳐 어떻게 변화하는지를 관찰한다.[71]
6. 1. 회귀 분석 (Regression analysis)
수량 유전학은 유전자 상호작용으로 인해 발생하는 유전 분산에 주목한다. 특정 유전자 빈도에서 두 개의 유전자좌가 상호작용할 때, 회귀 분석을 이용하여 이를 8개의 독립적인 유전 효과로 나누어 분석할 수 있다. 이 회귀 분석에서는 관찰된 두 유전자좌의 유전 효과를 종속 변수로, "순수한" 유전 효과를 독립 변수로 설정한다. 회귀 분석은 가중치를 적용하기 때문에, 분산 성분이 어떻게 나뉘는지는 유전자 빈도에 따라 달라진다. 이러한 분석 방법을 통해, 세 개 이상의 유전자좌나 세포질과 핵 간의 상호작용으로 분석 대상을 확장할 수도 있다.[55]6. 2. 이중 돌연변이 주기 (Double mutant cycles)
유전자 내에서 상위성을 분석하기 위해 부위 특이적 돌연변이 유발 기법을 사용하여 다양한 유전자를 만들고, 이 유전자들이 만들어내는 단백질 산물의 특성(예: 안정성 또는 촉매 활성)을 분석할 수 있다. 이 과정을 이중 돌연변이 주기(double mutant cycle)라고 부르기도 한다. 이 방법은 원래의 정상적인 단백질(야생형), 돌연변이가 하나씩 있는 단백질 두 종류(단일 돌연변이체), 그리고 두 돌연변이를 모두 가진 단백질(이중 돌연변이체)을 만들어 각각 분석하는 것을 포함한다. 상위성은 두 돌연변이가 함께 있을 때 나타나는 효과와 각각의 돌연변이가 개별적으로 나타내는 효과의 합 사이의 차이로 측정된다.[56] 이는 상호 작용의 자유 에너지로 표현될 수 있다.같은 방법을 사용하여 더 많은 수의 돌연변이 사이의 상호 작용을 조사할 수도 있지만, 이를 위해서는 가능한 모든 조합을 만들고 분석해야 하는 어려움이 있다. 예를 들어, 5개의 다른 돌연변이가 있다면 총 120가지의 조합이 가능하며, 이 조합들 중 일부 또는 전부에서 상위성이 나타날 수 있다.
6. 3. 계산 예측 (Computational prediction)
상위성을 감지하고 그 특성을 밝히기 위해 여러 계산 방법이 개발되었다. 이 방법들 중 상당수는 선형 회귀 같은 단순한 통계 방식으로는 찾아내기 어려운 비가산적 효과(non-additive effect)를 감지하기 위해 머신 러닝 기술을 활용한다.[57]예를 들어, 다요인 차원 축소법(MDR, Multifactor Dimensionality Reduction)은 특정 모델에 얽매이지 않고 비모수적인 방식으로 인간 집단 내에서 질병 상태와 같은 표현형을 예측하는 유전자 변이 조합을 찾아내도록 설계된 방법이다.[58][59] 이러한 계산적 접근 방식들에 대해서는 이미 많은 연구와 검토가 이루어졌다.[60]
최근에는 이론 전산학(Hadamard 변환[61]이나 압축 센싱[62][63] 등)이나 최대 우도 추론[64] 같은 분야의 아이디어를 활용하는 방법들이 등장했다. 이러한 방법들은 유전자형-표현형 관계에서 나타나는 전반적인 비선형성 속에서 순수한 상위성 효과만을 구분해내는 데 효과적인 것으로 밝혀졌다.[65] 또한, 환자의 생존 데이터를 분석하여 비선형적 상호작용을 식별하는 연구도 진행되고 있다.[66]
7. 상위성의 예
생쥐의 털색은 여러 유전자에 의해 결정된다. 자연 상태의 생쥐는 각 털에 무늬가 있어 전체적으로 회색을 띤다. 생쥐 털색에 관여하는 주요 유전자는 다음과 같다.
| 염색체 위치 | 유전자 | 역할 | 발현 특징 |
|---|---|---|---|
| A | AA | 색소 형성을 지시한다 | 우성 동형접합 유전자 |
| Aa | 색소 형성을 지시한다 | 우성 대립유전자 | |
| aa | 색소 형성을 막는다 (흰색) | 열성 동형접합 유전자 | |
| B | BB | 회색의 무늬를 만든다 | 우성 동형접합 유전자 |
| Bb | 회색의 무늬를 만든다 | 우성 대립 유전자 | |
| bb | 무늬 형성을 억제하여 검은색이 되게 한다 | 열성 동형접합 유전자 |
유전형이 AABB인 회색 생쥐와 aabb인 흰색 생쥐를 교배하면, 첫 번째 자식 세대(F1)의 유전형은 모두 AaBb가 되어 회색 털색을 나타낸다. 이 F1 세대를 다시 교배하여 얻은 두 번째 자식 세대(F2)에서는 털색 발현 비율이 다음 표와 같이 9(회색) : 3(검은색) : 4(흰색)로 나타난다.
| X | AB | Ab | aB | ab |
|---|---|---|---|---|
| AB | AABB (회색) | AABb (회색) | AaBB (회색) | AaBb (회색) |
| Ab | AABb (회색) | AAbb (검은색) | AaBb (회색) | Aabb (검은색) |
| aB | AaBB (회색) | AaBb (회색) | aaBB (흰색) | aaBb (흰색) |
| ab | AaBb (회색) | Aabb (검은색) | aaBb (흰색) | aabb (흰색) |
이 결과는 색소 형성 자체를 막는 열성 유전자형 aa가 털 무늬를 결정하는 B/b 유전자보다 상위에서 작용함을 보여준다. 즉, aa 유전자형을 가지면 B 유전자의 종류와 관계없이 무조건 흰색 털을 갖게 된다. 이는 A/a 유전자가 만드는 효소가 먼저 작용한 후, B/b 유전자가 관여하는 효소가 작용하기 때문이다.[72] 이러한 상위성 때문에 해당 유전자들은 멘델의 유전법칙 중 독립의 법칙을 따르지 않는 분리비를 보인다.
상위성은 두 좌위 간의 상호작용으로 나타나며, 표현형의 분리비를 변화시킨다. 주요 상위성의 종류는 다음과 같다.
- '''피복 유전자 (우성 상위, dominant epistasis)'''
- 하나의 우성 유전자(A)가 다른 좌위 유전자(B/b)의 형질 발현을 억제한다.
- A가 존재하면 B/b의 표현형이 드러나지 않아 AB와 Ab의 표현형이 같아진다.
- 분리비: (AB+Ab) : aB : ab = 12 : 3 : 1
- 예시: 호박의 과피색 유전
- '''조건 유전자 (열성 상위, recessive epistasis)'''
- 하나의 열성 유전자형(aa)이 다른 좌위 유전자(B/b)의 형질 발현을 억제한다. (위의 생쥐 털색 예시 해당)
- aa일 경우 B/b의 표현형이 드러나지 않아 aB와 ab의 표현형이 같아진다.
- 분리비: AB : Ab : (aB+ab) = 9 : 3 : 4
- 예시: 양파의 색 유전
- '''보충 유전자 (Complementary Genes)'''
- 두 좌위의 우성 유전자(A와 B)가 함께 존재해야 특정 형질이 발현된다. A나 B 중 하나라도 없으면 다른 형질이 나타난다. 각 유전자 자체에는 우열 관계가 없다.
- 분리비: AB : (Ab+aB+ab) = 9 : 7
- 예시: 스위트피의 꽃 색깔 유전
- '''억제 유전자 (Inhibitory Gene)'''
- 하나의 우성 유전자(A)가 다른 좌위의 우성 유전자(B)의 발현을 억제한다. B/b 자체는 형질을 나타내지만, A가 존재하면 B의 효과가 나타나지 않는다.
- 분리비: (AB+Ab+ab) : aB = 13 : 3
- 예시: 누에나방의 고치 색깔 유전
- '''동의 유전자 (Duplicate Genes)'''
- 두 좌위의 우성 유전자(A 또는 B) 중 하나라도 존재하면 동일한 특정 형질이 발현된다. 둘 다 열성(aabb)일 때만 다른 형질이 나타난다.
- 분리비: (AB+Ab+aB) : ab = 15 : 1
- 예시: 황새냉이의 열매 모양 유전
과거에는 분리비가 12:3:1이 되는 피복 유전자의 경우만을 엄밀한 의미의 상위성으로 보았으나, 현재는 위와 같이 유전자 간 상호작용으로 인해 멘델의 독립 법칙에서 예상되는 분리비(9:3:3:1)를 변화시키는 모든 경우를 넓은 의미의 상위성으로 부른다.
참조
[1]
서적
Biologie
Spektrum-Verlag Heidelberg-Berlin
2003
[2]
논문
The Evolution of Epistasis and its Links with Genetic Robustness, Complexity and Drift in a Phenotypic Model of Adaptation
2009-05
[3]
서적
A Glossary of Genetics and Cytogenetics: Classical and Molecular
Springer-Verlag
[4]
논문
Women in the Early History of Genetics: William Bateson and the Newnham College Mendelians, 1900–1910
The History of Science Society
[5]
논문
Quantitative analyses of empirical fitness landscapes
2013-01-16
[6]
논문
Evolution of robustness in digital organisms
2004
[7]
논문
Genome-wide interaction and pathway-based identification of key regulators in multiple myeloma
2019
[8]
서적
Proceedings of the 5th ACM Conference on Bioinformatics, Computational Biology, and Health Informatics
[9]
논문
iLOCi: a SNP interaction prioritization technique for detecting epistasis in genome-wide association studies
2012-12-13
[10]
논문
Epistasis--the essential role of gene interactions in the structure and evolution of genetic systems
2008-11
[11]
논문
Viral quasispecies evolution
2012-06
[12]
논문
Stability effects of mutations and protein evolvability
2009-10
[13]
논문
Prevalent positive epistasis in Escherichia coli and Saccharomyces cerevisiae metabolic networks
2010-03
[14]
서적
Evolution
Blackwell Publishing.
[15]
서적
Elements of Evolutionary Genetics
Roberts and Company Publishers
[16]
논문
Crystal structure of an ancient protein: evolution by conformational epistasis
2007-09
[17]
논문
Epistasis: what it means, what it doesn't mean, and statistical methods to detect it in humans
2002-10
[18]
서적
The Origins of Order: Self-organization and Selection in Evolution
Oxford University Press
1993
[19]
논문
Engineering the third wave of biocatalysis
2012-05
[20]
논문
Sexual reproduction selects for robustness and negative epistasis in artificial gene networks
2006-03
[21]
논문
Evidence for positive epistasis in HIV-1
2004-11
[22]
논문
Perspective: Sign epistasis and genetic constraint on evolutionary trajectories
2005-06
[23]
논문
Empirical fitness landscapes reveal accessible evolutionary paths
2007-01
[24]
논문
Computational Complexity as an Ultimate Constraint on Evolution
2019-05-01
[25]
논문
Interaction of morphogenetic genes of bacteriophage T4
1970
[26]
웹사이트
Synthetic Lethal Mutations
http://www.sci.sdsu.[...]
Department of Microbiology, University of Illinois, Urbana
2017-08-03
[27]
서적
An introduction to genetic statistics
Iowa State University Press
[28]
논문
The biochemical architecture of an ancient adaptive landscape
2005-10
[29]
논문
General nature of the genetic code for proteins
1961
[30]
논문
Protein structure and evolutionary history determine sequence space topology
2005-03
[31]
논문
Evolutionary biochemistry: revealing the historical and physical causes of protein properties
2013-08
[32]
논문
Recent developments in disulfide bond formation
[33]
논문
Robustness-epistasis link shapes the fitness landscape of a randomly drifting protein
2006-12
[34]
논문
Protein sectors: evolutionary units of three-dimensional structure
2009-08
[35]
논문
The conversion of serine at the active site of subtilisin to cysteine: a "chemical mutation"
1966-11
[36]
논문
A theoretical study of the active sites of papain and S195C rat trypsin: implications for the low reactivity of mutant serine proteinases
1996-07
[37]
논문
Thiol-beta-lactamase: replacement of the active-site serine of RTEM beta-lactamase by a cysteine residue
1982-12
[38]
논문
Intragenic Complementation Among Temperature Sensitive Mutants of Bacteriophage T4D
1965
[39]
논문
The Theory of Inter-Allelic Complementation
1964
[40]
논문
Reciprocal sign epistasis is a necessary condition for multi-peaked fitness landscapes
https://hal.archives[...]
2011-03
[41]
논문
Environmental changes bridge evolutionary valleys
2016-01
[42]
논문
Constructing and analyzing the fitness landscape of an experimental evolutionary process
2008-09
[43]
논문
Darwinian evolution can follow only very few mutational paths to fitter proteins
2006-04
[44]
논문
Stability-mediated epistasis constrains the evolution of an influenza protein
2013-05
[45]
논문
Shifting Fitness and Epistatic Landscapes Reflect Trade-offs along an Evolutionary Pathway
https://zenodo.org/r[...]
2016-07
[46]
논문
The importance of molecular hierarchy in information processing
Edinburgh University Press Edinburgh
[47]
논문
Bootstrapping on the adaptive landscape
[48]
서적
Die Ordnung des Lebendigen: Systembedingungen der Evolution
Parey
[49]
논문
A systems-analytical approach to macroevolutionary phenomena
[50]
논문
Predictability of evolutionary trajectories in fitness landscapes
2011-12
[51]
논문
An epistatic ratchet constrains the direction of glucocorticoid receptor evolution
2009-09
[52]
논문
Deleterious mutations and the evolution of sexual reproduction
1988-12
[53]
논문
Coevolution of robustness, epistasis, and recombination favors asexual reproduction
2007-07
[54]
논문
Survival of the sparsest: robust gene networks are parsimonious
2008-08
[55]
논문
Cyto-nuclear epistasis: two-locus random genetic drift in hermaphroditic and dioecious species
2006-04
[56]
논문
Double-mutant cycles: a powerful tool for analyzing protein structure and function
[57]
서적
Epistasis
2021-03-18
[58]
서적
Epistasis
2015-01-01
[59]
서적
Epistasis: Methods and Protocols
Springer
[60]
논문
Detecting gene-gene interactions that underlie human diseases
2009-06
[61]
논문
Should evolutionary geneticists worry about higher-order epistasis?
2013-12
[62]
논문
The Context-Dependence of Mutations: A Linkage of Formalisms
2016-06
[63]
논문
Learning the pattern of epistasis linking genotype and phenotype in a protein
2019-09
[64]
논문
Inferring the shape of global epistasis
2018-08
[65]
논문
Detecting High-Order Epistasis in Nonlinear Genotype-Phenotype Maps
2017-03
[66]
논문
Beyond Synthetic Lethality: Charting the Landscape of Pairwise Gene Expression States Associated with Survival in Cancer
[67]
서적
ハートウェル遺伝学
[68]
서적
エッセンシャル遺伝学
培風館
[69]
서적
チャート式シリーズ 新生物 生物基礎・生物
数研出版
[70]
서적
遺伝学キーノート
シュプリンガー・フェアラーク東京
[71]
논문
Epistasis: what it means, what it doesn't mean, and statistical methods to detect it in humans
2002
[72]
서적
생명 생물의 과학
교보문고
2006
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com