질형목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
질형목은 유두윤충강, 단성강, 세이손강으로 구성되었으나, 분자 계통 발생 연구를 통해 극두동물이 추가되어 윤형동물문으로 재분류되었다. 윤형동물문 내에서 유두윤충강의 위치는 논쟁의 여지가 있으며, 세이손강과 극두동물이 자매 분류군이고 단성강이 외부군으로 여겨진다. 질형목은 윤형동물강에 속하며, 세 개의 목과 약 450종으로 나뉜다. 이들은 무성생식을 하며, 형태학적 데이터와 분자 데이터를 혼합하여 종을 동정한다. 질형목은 섭식, 이동, 생식, 스트레스 유발 행동을 보이며, 특히 무성생식과 건조 상태에서의 생존 능력, 수평 유전자 이동 등의 특징을 가진다.
더 읽어볼만한 페이지
| 질형목 - [생물]에 관한 문서 | |
|---|---|
| 분류 정보 | |
| 형태 및 생태 | |
| 기타 정보 | |
2. 진화적 관계
윤형동물문은 전통적으로 유두윤충강, 단성강, 세이손강의 세 강으로 분류되었으나, 분자 계통 발생 연구를 통해 극두동물이 윤형동물문과 밀접한 관계를 가지는 네 번째 분지군임이 밝혀졌다.[10] 극두동물은 원래 별도의 문으로 분류되었으나, 1994년과 2014년 사이에 축적된 분자 및 형태학적 증거는 극두동물이 유두윤충강, 단성강, 세이손강과 함께 단계통군을 형성한다는 것을 보여준다.[8][11]
2. 1. 윤형동물문 내의 계통 발생
질형목은 전통적으로 유두윤충강, 단성강, 세이손강의 세 강으로 구성되었다.[6] 1990년 이전의 형태학적 계통 발생 연구에서는 유두윤충류의 자매군이 단성강이고, 세이손류는 초기에 분기된 외부군으로 나타났다.[7]
현대 분자 계통 발생 연구는 '윤형동물'에 대한 이러한 고전적인 이해가 불완전하며, 네 번째 분지군인 가시머리벌레 (극두동물)을 포함하지 않는다는 것을 보여준다.[10] 극두동물은 원래 별도의 문으로 분류되었으나, 1994년과 2014년 사이에 축적된 분자 및 형태학적 증거는 극두동물이 유두윤충강, 단성강, 세이손강과 함께 단계통군을 형성한다는 것을 보여준다.[8][11] 이 발견을 수용하기 위해 일부 학자들은 '윤형동물'이라는 용어를 확장하여 유두윤충, 단성, 세이손 윤형동물 외에 고도로 변형된 기생성 '극두동물 윤형동물'을 포함시킨다.[12] 다른 학자들은 네 분류군 그룹이 공유하는 합포체 표피에서 파생된 용어인 윤형동물문이라고 부른다.[11]
윤형동물문(또는 윤형동물) 내에서 유두윤충강의 위치는 명확하지 않다. 가능한 계통 발생 관계는 첨부된 분류도에 설명되어 있다. 2014년 현재, "윤형동물 관계에 대한 가장 포괄적인 계통 유전체 분석"은 네 그룹 모두의 전사체 데이터를 기반으로 하며,[8] 그림의 왼쪽 하단에 표시된 가설에 대해 "강력한 지지"를 제공한다. 이 연구는 세이손강과 극두동물이 자매 분류군이며, 단성강은 이 세 종 모두의 외부군임을 보여준다. 즉, 유두윤충류의 가장 가까운 현존하는 친척은 이전에 생각했던 단성 윤형동물이 아니라, 고도로 변형된 형태임에도 불구하고 세이손 윤형동물과 극두동물이다.
3. 분류 및 동정
윤형동물문의 강인 질형강은 Philodinavida, Philodinida, Adinetida의 세 가지 목으로 구성된다.[13] 이 목들은 4개의 과와 약 450종으로 나뉜다.[14]
질형동물은 살아있을 때만 눈으로 식별할 수 있는데, 이는 분류에 중요한 많은 특징이 먹이 섭취 및 이동과 관련이 있기 때문이다. 그러나 죽은 개체에 대해서도 유전자적 식별이 가능하다. 일단 보존되면 개체는 "덩어리"로 수축되어 분석을 제한한다.[18] 현재 세 가지 형태학적 식별 방법이 있으며, 그중 두 가지는 오래된 것으로 간주된다. Bartoš (1951)[17]와 Donner (1965)[14]가 그것이며, 세 번째 방법은 1995년 Shiel에 의해 개발된 진단 키이다.[18]
3. 1. 종 동정의 어려움
질형강은 무성 생식을 하기 때문에, 종을 비옥한 자손을 낳을 수 있는 생물 집단으로 정의하는 일반적인 방식은 적용되지 않는다. 따라서 이 생물들의 종 개념은 형태학적 데이터와 분자 데이터를 혼합하여 사용한다. DNA 연구에 따르면, 다양성은 원래 형태학적 분류가 제시하는 것보다 훨씬 더 크다.[15][16]3. 2. 형태학적 특징
질형목의 몸은 머리, 몸통, 발의 세 가지 주요 부위로 구성된다. 종에 따라 신체 부위의 명칭은 같지만, 그 형태는 매우 다를 수 있다. 질형목은 일반적으로 잘 발달된 관(corona)을 가지고 있으며, 이는 접이식 머리(retractile head)에 두 부분으로 나뉜다.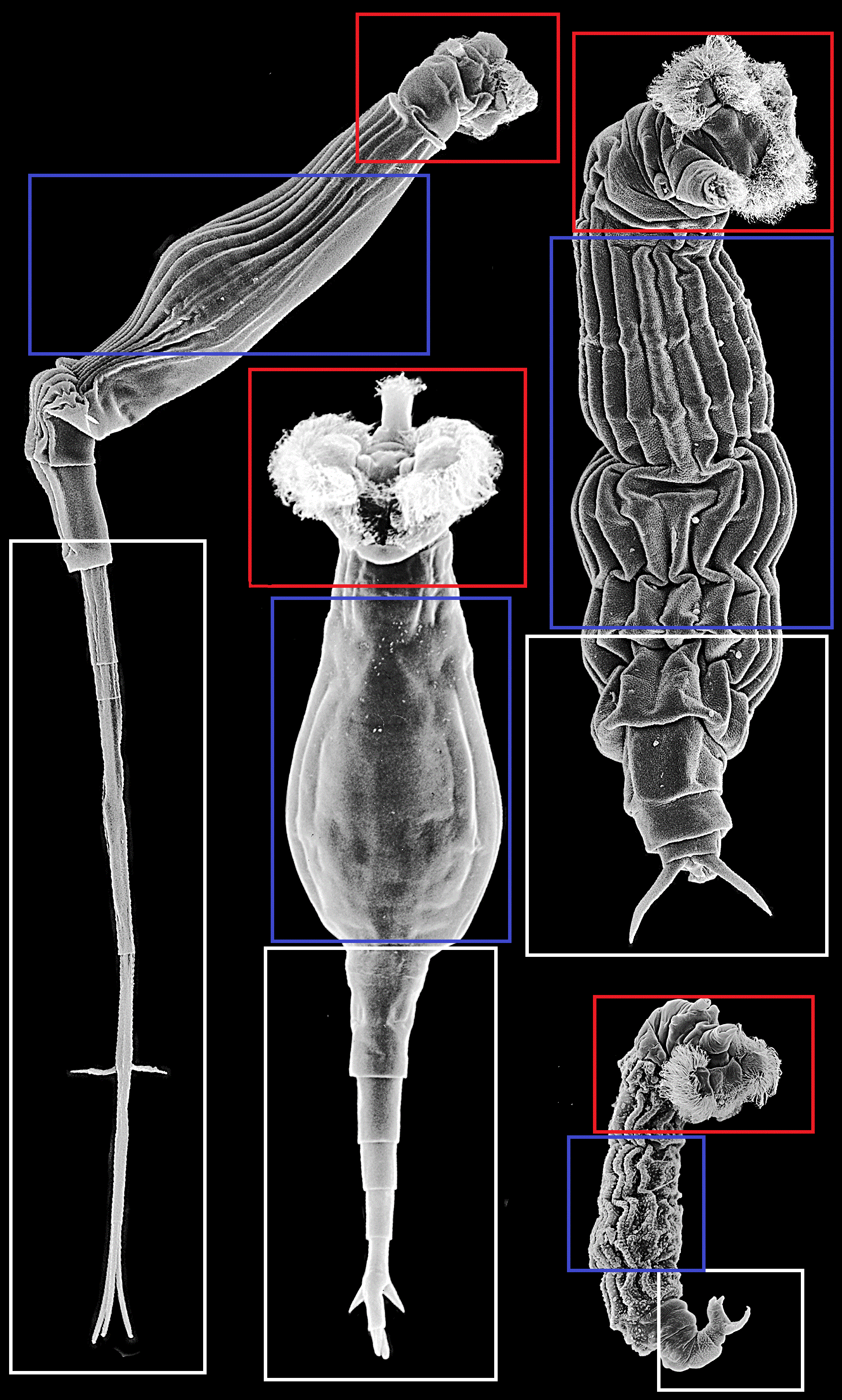
질형목의 식별 특징은 다음과 같다.
- 잘 발달된 발샘[18]
- 긴 식도가 있는 입[18]
- 강한 이빨 (치아 지수로 표시)[18]
- 많은 섬모[18]
- 종에 따라 다른 윗입술 모양[18]
- 목(order)별 관 유형[3]
- Philodinida: 두 개의 섬모판으로 구성
- Adinetida: 복부 섬모장으로 구성
- Philodinavida: 작은 관을 가짐
질형목의 소화 및 생식 기관은 몸통 부분에서 발견되며, 위는 가장 눈에 띄는 기관이다. 특정 속( ''Habrotrocha'', ''Otostephanos'', ''Scepanotrocha'')에서는 질형목이 위 내에 뚜렷한 구형 펠렛 모양으로 식별될 수 있는데, 이는 대변으로 배출된다. 다른 속들은 느슨한 물질 형태로 대변을 배출하기 때문에 이는 구별되는 특징이 된다.[3]
대부분의 질형목은 먹이를 먹는 동안 발을 움츠리지만, ''Adineta'', ''Bradyscela'', ''Henoceros'', ''Philodinavus''의 네 속은 발이 없다. 이는 먹이 섭취 방식뿐만 아니라 이동 방식에도 영향을 미친다. 예를 들어 ''Adineta''와 ''Bradyscela''는 미끄러지듯 움직이는 반면, 다른 속들은 고리 모양으로 움직인다.[3]
4. 행동
질형강의 행동은 섭식, 이동, 생식, 스트레스 유발 행동으로 나눌 수 있다.
- '''섭식:''' 대부분 코로나 기관의 섬모 고리를 사용하여 물의 흐름을 만들어 입을 통해 음식을 마스탁스 기관으로 불어넣어 음식을 갈도록 특별히 적응시켰다.[19] 음식에는 부유 세균, 조류, 부스러기 및 기타 물질이 포함된다.
- '''이동:''' 이동에는 자유 수영, 기질을 따라 꿈틀거리며 이동하기, 고착성의 세 가지 주요 방식이 있다. 꿈틀거림(또는 기어다니기)은 특정 거머리처럼 머리와 꼬리로 번갈아 단계를 밟는 것을 포함하며, 이로 인해 이 그룹의 이름이 붙여졌다.(βδέλλα|bdellagrc, 거머리라는 뜻)[1]
- '''생식:''' 질형강은 수컷이 관찰된 적이 없고,[20] 암컷이 수정 없이 배아가 성장하고 발달하는 무성생식의 한 형태인 처녀생식을 통해서만 번식하기 때문에 성의 진화 연구에 흥미로운 대상이다.[21] 각 개체는 한 쌍의 생식선을 가지고 있다.
- '''스트레스 유발 행동:''' 질형강은 무수생명(휴면 상태)을 통해 건조와 같은 환경 스트레스에 저항한다. 이 휴면 상태에서 최대 9년까지 생존 가능하며, 유리한 조건이 돌아오면 몇 시간 안에 빠르게 활성화된다.[28]
4. 1. 섭식
대부분은 코로나 기관의 섬모 고리를 사용하여 물의 흐름을 만들어 입을 통해 음식을 마스탁스 기관으로 불어넣어 음식을 갈도록 특별히 적응시켰다.[19] 음식에는 부유 세균, 조류, 부스러기 및 기타 물질이 포함된다.4. 2. 이동
이동에는 자유 수영, 기질을 따라 꿈틀거리며 이동하기, 고착성의 세 가지 주요 방식이 있는 것으로 보인다. 꿈틀거림(또는 기어다니기)은 특정 거머리처럼 머리와 꼬리로 번갈아 단계를 밟는 것을 포함하며, 이로 인해 이 그룹의 이름이 붙여졌다. (βδέλλα|bdellagrc, 거머리라는 뜻)[1]4. 3. 생식
질형강은 수컷이 관찰된 적이 없고,[20] 암컷이 수정 없이 배아가 성장하고 발달하는 무성생식의 한 형태인 처녀생식을 통해서만 번식하기 때문에 성의 진화 연구에 흥미로운 대상이다.[21] 각 개체는 한 쌍의 생식선을 가지고 있다.그러나 새로운 연구에서는 고대 무성생식으로 여겨졌던 종인 ''Adineta vaga''에서 개체 간 유전자 교환 및 재조합의 증거를 제시했다.[22]
''Adineta vaga''는 비감수 감수 분열에 의해 DNA 복구를 수행할 수 있다.[23] 생식 계열 DNA 복구는 상동 염색체가 감수 분열과 유사한 위치에 놓이는 난자 형성의 특정 기간에 발생한다.[1] 이 생식 계열 DNA 복구는 자손에게 전달되는 유전 물질을 정확하게 재구성한다.
4. 4. 스트레스 유발 행동
질형강은 무수생명(휴면 상태)을 통해 건조와 같은 환경 스트레스에 저항한다. 이 휴면 상태에서 최대 9년까지 생존 가능하며, 유리한 조건이 돌아오면 몇 시간 안에 빠르게 활성화된다.[28] 건조 스트레스는 일부 종의 적합성과 다산성을 향상시키기도 한다.[29]4. 5. 수평 유전자 이동
질형강은 박테리아, 곰팡이, 식물로부터 수평 유전자 이동을 통해 유전자를 획득한 것으로 알려져 있으며, 이는 윤형동물 진화의 중요한 요인일 수 있다.[33] 환형동물에서 수평 유전자 이동이 어떻게, 왜 발생하는지는 현재 많은 논쟁의 대상이다. 특히 외래 유전자와 건조 과정 사이의 가능한 연결뿐만 아니라 환형동물의 고대 무성생식과의 가능한 연결과 관련하여 논쟁이 이루어지고 있다.완전히 건조되면 DNA가 여러 조각으로 분해된다. 그리고 재수화를 거쳐 다시 생명을 되찾을 때, 이종 DNA 조각이 게놈에 들어갈 기회를 만든다. 이 과정은 6천만 년 전에 이 방식으로 박테리아 유전자를 포착했을 때 개선되었으며, 이를 통해 새로운 유전자 조절 시스템을 얻었다. 새로운 시스템은 전이 인자를 제어하는 데 사용되었다.[34]
5. DNA 복구 능력
질형강은 휴면 상태에서 사용되는 DNA 보존 적응을 통해 이온화 방사선으로 인한 손상에 매우 강하다.[35] 이러한 적응에는 DNA 이중 가닥 파손을 복구하기 위한 매우 효율적인 메커니즘이 포함된다.[36] 이 복구 메커니즘은 ''Adineta vaga''[36] 및 ''Philodina roseola''[37] 두 종의 환형동물에서 연구되었으며, 각 종 내의 상동 DNA 영역 간의 유사분열 재조합을 포함하는 것으로 보인다.
6. 의무적 처녀생식의 진화
2003년, 윤충류의 무성 생식 방식은 완전히 밝혀지지 않았다.[24] 윤충류에서 의무적 처녀생식이 어떻게 발생했는지에 대한 한 가지 이론은 처녀생식 계통이 성 유도 신호에 반응하는 능력을 상실했고, 이것이 이 계통이 무성 생식을 유지하는 이유라는 것이다.[25] 이후 의무적 처녀생식 윤충류 계통은 성 유도 신호를 생성하지만, 그 신호에 반응하는 능력을 상실했으며, 처녀생식을 하는 의무적 처녀생식 개체에서 성 유도 신호에 반응하지 못하는 것은 유전자 ''op''의 단순한 멘델 유전에 의해 발생한다는 것이 밝혀졌다.[26]
7. 갤러리
참조
[1]
서적
Bestimmungsbücher zur Bodenfauna Europas, volume 6
Akademie Verlag
[1]
학술지
Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution
http://www.mapress.c[...]
[2]
서적
Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution
http://www.mapress.c[...]
Magnolia Press
2007
[3]
학술지
Key to the identification of the genera of bdelloid rotifers
[4]
학술지
Bdelloid rotifers in Dominican amber: Evidence for parthenogenetic continuity
[5]
뉴스
This Tiny Creature Survived 24,000 Years Frozen in Siberian Permafrost - The microscopic animals were frozen when woolly mammoths still roamed the planet, but were restored as though no time had passed.
https://www.nytimes.[...]
2021-06-07
[6]
학술지
Evolutionary Dynamics of 'the' Bdelloid and Monogonont Rotifer Life-history Patterns
2005-09
[7]
학술지
Phylogenetic relationships within phylum Rotifera: orders and genus Notholca
1989-12
[8]
학술지
Transcriptome Data Reveal Syndermatan Relationships and Suggest the Evolution of Endoparasitism in Acanthocephala via an Epizoic Stage
2014-02-10
[9]
학술지
A Mitogenomic Re-Evaluation of the Bdelloid Phylogeny and Relationships among the Syndermata
2012-08-23
[10]
학술지
Early contributions of molecular phylogenetics to understanding the evolution of Rotifera
[11]
학술지
Epidermal ultrastructure of ''Seison nebaliae'' and ''Seison annulatus'', and a comparison of epidermal structures within the Gnathifera
[12]
서적
Animal evolution : interrelationships of the living phyla
Oxford University Press
2012
[13]
학술지
Rotatory apparatus in Bdelloids
[14]
서적
Ordnung Bdelloidea
Akademie-Verlag
[15]
학술지
Inconsistent estimates of diversity between traditional and DNA taxonomy in bdelloid rotifers
[16]
학술지
Extreme levels of hidden diversity in microscopic animals (Rotifera) revealed by DNA taxonomy
[17]
학술지
The Czechoslovak Rotatoria of the order Bdelloidea
[18]
서적
A guide to identification of rotifers, cladocerans and copepods from Australian inland waters
Co-operative Research Centre for Freshwater Ecology
[19]
학술지
The hard parts (trophi) of the rotifer mastax do contain chitin: evidence from studies on Brachionus plicatilis
[20]
학술지
Ancient asexual scandals
[21]
간행물
Bdelloids: No sex for over 40 million years
http://www.thefreeli[...]
2002-11-01
[22]
학술지
Genomic signatures of recombination in a natural population of the bdelloid rotifer Adineta vaga
2020-12-18
[23]
학술지
DNA repair during nonreductional meiosis in the asexual rotifer Adineta vaga
2022
[24]
학술지
Phylogenetic relationships between parthenogens and their sexual relatives: the possible routes to parthenogenesis in animals
[25]
학술지
Obligate asex in a rotifer and the role of sexual signals
[26]
학술지
Loss of Sexual Reproduction and Dwarfing in a Small Metazoan
2010-09-20
[27]
학술지
Anhydrobiosis: an unsolved problem
[28]
학술지
Long-term anhydrobiotic survival in semi-terrestrial micrometazoans
http://lup.lub.lu.se[...]
[29]
학술지
The importance of being a bdelloid: ecological and evolutionary consequences of dormancy
[30]
학술지
Stress and fitness in parthenogens: is dormancy a key feature for bdelloid rotifers?
[31]
학술지
Anciently Asexual Bdelloid Rotifers Escape Lethal Fungal Parasites by Drying Up and Blowing Away
2010-01-29
[32]
학술지
Spatial and temporal escape from fungal parasitism in natural communities of anciently asexual bdelloid rotifers
2013-08-22
[33]
학술지
Foreign genes and novel hydrophilic protein genes participate in the desiccation response of the bdelloid rotifer ''Adineta ricciae''
[34]
웹사이트
60-M-Year-Old Bacteria Shed Light on 'New' DNA Modification System in Animals
https://www.laborato[...]
[35]
학술지
Extreme resistance of bdelloid rotifers to ionizing radiation
2008-04-01
[36]
논문
Gateway to genetic exchange? DNA double-strand breaks in the bdelloid rotifer Adineta vaga submitted to desiccation
https://hal-pasteur.[...]
2014-07
[37]
논문
Evidence for degenerate tetraploidy in bdelloid rotifers
2008-04-01
[38]
논문
Massive horizontal gene transfer in bdelloid rotifers
http://nrs.harvard.e[...]
2008-05-30
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com