켄트로사우루스 (검룡)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
켄트로사우루스는 스테고사우루스과에 속하는 초식 공룡으로, 몸길이 2.5~5m, 몸무게 400kg~1.5t으로 추정된다. 켄트로사우루스는 작은 머리, 긴 목, 짧은 앞다리와 긴 뒷다리, 길고 수평하며 근육질의 꼬리를 특징으로 하며, 등에는 골판과 가시가 2열로 배열되어 있다. 꼬리 가시는 방어 수단으로 사용되었으며, 켄트로사우루스는 텐다구루 지층에서 발견되었으며, 스테고사우루스와 근연 관계에 있다.
더 읽어볼만한 페이지
- 쥐라기 후기 아프리카의 공룡 - 케라토사우루스
케라토사우루스는 쥐라기 후기에 북미, 유럽, 아프리카 등지에서 서식했으며 머리 뿔과 몸 중앙선의 골편, 강력한 뒷다리를 가진 이족보행 육식 공룡으로, 다른 대형 수각류와 공존하며 먹이 경쟁을 피했을 것으로 추정된다. - 쥐라기 후기 아프리카의 공룡 - 알로사우루스
알로사우루스는 쥐라기 후기에 살았던 대형 수각류 공룡으로, 짧은 앞다리, 머리의 뿔, 갈고리 발톱을 특징으로 하며, 모리슨층에서 발견되었고, 다른 육식 공룡과 함께 먹이 사슬의 정점에 있었다. - 1915년 기재된 화석 분류군 - 기간토피테쿠스
기간토피테쿠스는 거대한 어금니 화석을 토대로 명명된 멸종된 유인원 속으로, 큰 덩치와 초식 또는 잡식성 식습관을 가진 것으로 추정되지만, 오랑우탄과 더 가까운 관계이며 기후 변화 등으로 멸종했을 가능성이 있고, 미확인 동물의 정체로도 거론된다. - 1915년 기재된 화석 분류군 - 스피노사우루스
스피노사우루스는 백악기 전기부터 후기 북아프리카에 살았던 스피노사우루스과의 수각류 공룡으로, 긴 신경배돌기("돛")가 있는 등뼈가 특징이며 가장 큰 수각류 공룡 중 하나로 여겨지지만 식성, 수생 생활 여부, 이동 방식 등에 대한 논쟁이 있다. - 스테고사우루스과 - 우에르호사우루스
우에르호사우루스는 약 1억 3천만 년 전 전기 백악기 아시아에 서식했으며 넓은 몸통과 확장된 골반이 특징인 최대 7m 길이의 검룡류 공룡 속으로, 한때 스테고사우루스와 같은 속으로 분류되었으나 재분류되었다. - 스테고사우루스과 - 파란토돈
파란토돈은 백악기 초기에 남아프리카 공화국에서 서식했으며, 부분적인 두개골과 이빨 화석으로 알려진 검룡류 공룡의 한 속으로, 투오장고사우루스, 로리카토사우루스와 같은 다른 검룡류와 가까운 관계를 갖는 것으로 나타난다.
| 켄트로사우루스 (검룡) - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Kentrosaurus |
| 명명자 | Hennig, 1915 |
| 모식종 | Kentrosaurus aethiopicus |
| 모식종 명명자 | Hennig, 1915 |
| 동의어 | Kentrurosaurus (Hennig, 1916) Doryphorosaurus (Nopcsa, 1916) |
| 발음 | IPA: /ˈkɛntroʊˈsɔːrəs/ |
| 어원 | 가시 도마뱀 |
| 크기 | 4–4.5 m |
| 무게 | 700–1600 kg |
| 생물학적 분류 | |
| 상목 | 공룡상목 |
| 목 | 조반목 |
| 목 (계통발생학적 위치) | 게나사우리아 |
| 아목 | 장순아목 |
| 하목 | 검룡하목 |
| 과 | 스테고사우루스과 |
| 속 | Kentrosaurus |
| 시간적 범위 | |
| 시대 | 쥐라기 후기 (티토니아절), |
| 시기 | 약 1억 5,200만 년 전 |
| 분포 | |
| 지역 | 탄자니아 린디 지역 |
2. 형태

스테고사우루스와 같은 속에 속하지만, 크기나 몸의 유연성, 갑옷 등의 형태를 볼 때 다소 차이가 있다. 켄트로사우루스의 몸 전체 크기는 약 2.5~5m, 체중은 400kg~1.5t이었을 것으로 추정되어 스테고사우루스보다는 소형으로 보인다.[28]
''켄트로사우루스''는 작은 스테고사우루스였다. 켄트로사우루스는 작은 머리, 긴 목, 짧은 앞다리와 긴 뒷다리, 길고 수평하며 근육질의 꼬리로 특징지어지는 전형적인 공룡의 몸 구조를 가지고 있었다. 전형적인 스테고사우루스과의 특징으로는 머리의 길고 납작한 형태, 앞다리의 강력한 골격, 기둥과 같은 형태의 곧게 뻗은 뒷다리, 그리고 동물의 상단 중앙선을 따라 양쪽에 배열된 골판과 가시가 있었다.[15]
'''골격'''
''켄트로사우루스 아에티오피쿠스''는 비교적 작은 검룡류로, 몸길이는 4~4.5m에 달하고 몸무게는 700~1600kg 정도였다.[3] 일부 표본을 보면 비교적 더 큰 개체도 존재했을 수 있다. 이 표본들은 발달된 주두돌기 측면에서 일부 ''스테고사우루스'' 표본과 유사하다.[4]
''켄트로사우루스''의 긴 꼬리는 질량 중심이 사족 보행 동물로서는 이례적으로 뒤쪽에 위치하게 한다. 질량 중심은 엉덩이 바로 앞에 위치하는데, 이것은 보통 이족 보행 공룡에서 볼 수 있는 위치이다. 그러나 ''켄트로사우루스''의 넙다리뼈는 일반적인 이족 보행 공룡과는 달리 곧게 뻗어 있으며, 이는 다리가 곧고 수직으로 뻗은 자세를 나타낸다. 따라서 뒷다리는 긴 엉덩뼈에 붙어 있는 거대한 허벅지 근육의 힘을 받았지만, 동물을 혼자 지탱하지 못했으며, 매우 튼튼한 앞다리가 체중의 10~15%를 지탱했다. ''켄트로사우루스''나 ''스테고사우루스''의 골판(피부의 뼈 구조)은 질량 중심에 큰 영향을 미치지 않아, 이들은 무장했음에도 불구하고 이동성을 유지할 수 있었다. 뒷다리의 허벅지 근육은 매우 강력하여, ''켄트로사우루스''가 뒷다리와 꼬리로 삼각대 자세를 취할 수 있었다.[5]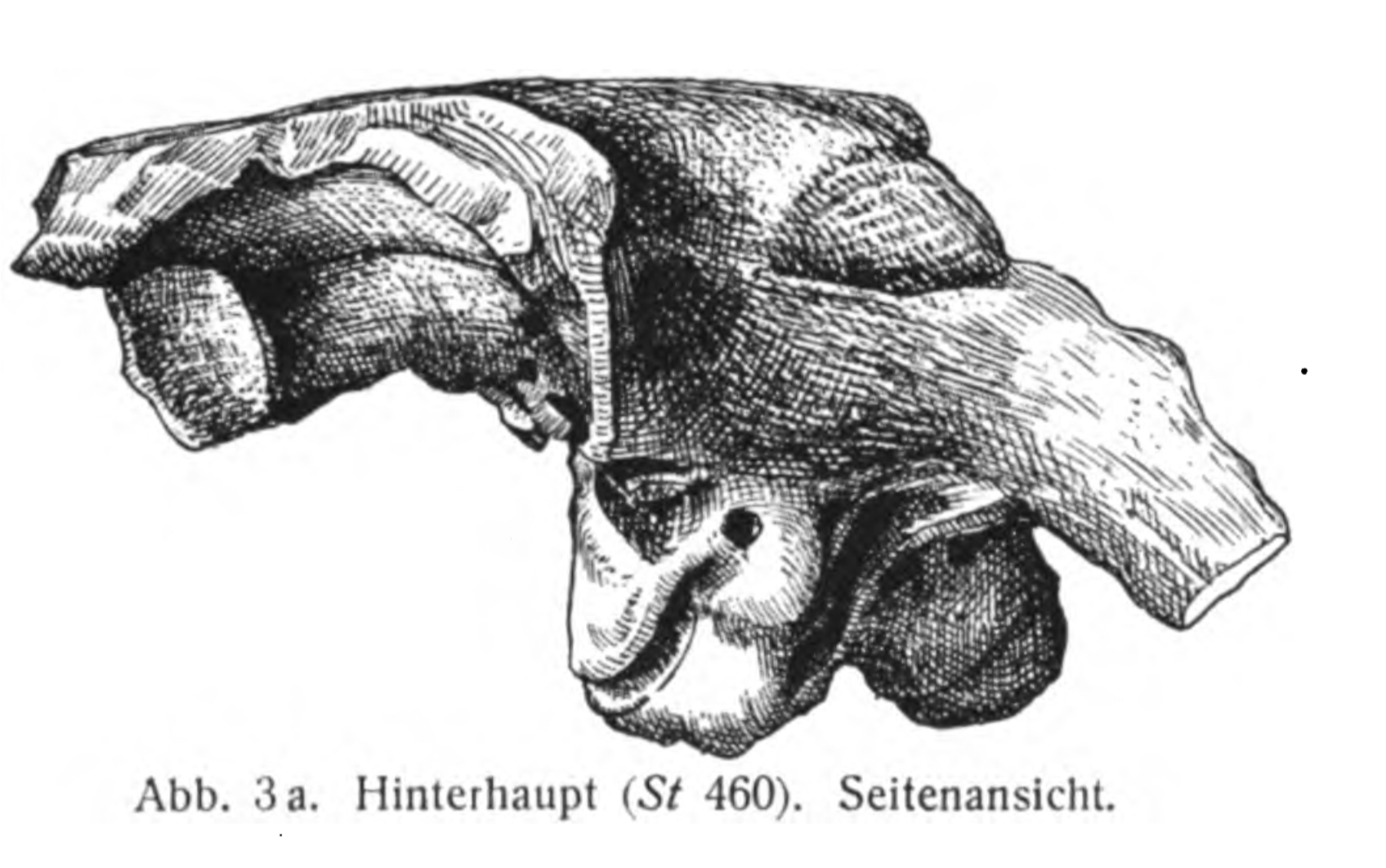
길고 좁은 두개골은 몸에 비해 작았다. 코와 눈 사이의 구멍인 작은 안와전개구가 있었지만, 현존하는 악어에서는 사라졌다. 두개골의 낮은 위치는 ''켄트로사우루스''가 낮게 자라는 식물을 먹었을 가능성을 시사한다. 이러한 해석은 앞니가 없고, 그 대신 뿔로 된 부리 또는 구치로 대체되었을 가능성이 있다는 점에서 뒷받침된다. 부리가 턱의 대부분을 따라 뻗어 있어 검룡류의 볼이 없을 수 있다.[8] ''켄트로사우루스''는 ''스테고사우루스''와 같이 광범위한 부리를 가졌을 가능성은 낮으며, 대신 턱 끝에 제한된 부리를 가졌을 것이다.[9][10]
하악골 (아래턱)에서 ''켄트로사우루스''는 불완전한 오른쪽 치골만 알려져 있다.[12] 깊은 치골은 ''스테고사우루스''와 거의 동일한 모양이다. 이빨은 일반적인 검룡류 이빨로 작고 넓어진 바닥과 수직 홈이 있다. 검룡류 이빨은 작고 삼각형이며 평평했다. 마모면은 그들이 음식을 갈았다는 것을 보여준다.[13]
[[파일:KentrosaurusDorsalVertebra.jpg|left|thumb|옆면 및 앞면에서 본 ''켄트로사우루스''의 등(뒷면) 척추.]
목은 13개의 목 척추로 구성되었으며, 축추가 뒤따른다. 등뼈는 13개의 등 척추로 구성된다. 천골(척추가 있는 골반 부분)은 6개의 융합된 중심을 이루며, 천골 늑골의 등쪽 부분과 융합되어 단단한 천골판을 형성한다. 미추 (꼬리) 척추는 29개이지만, 27-29개는 사고마이저 (꼬리 가시)에 부착하기 위해 함께 융합된다. ''켄트로사우루스''는 꼬리에서 대부분의 공룡과 달리 하위 평행하게 뻗어 있지 않은 척추의 많은 돌기로 인해 스테고사우루스의 다른 구성원과 구별할 수 있다.
[[파일:KentrosaurusSacrum(HMNSt439).jpg|thumb|배면에서 본 ''켄트로사우루스''의 천골.]
어깨뼈는 아랫부분이 사각형이며 튼튼한 날개를 가지고 있다. 오훼돌기는 반원형이다.[14] 앞다리는 튼튼한 뒷다리보다 훨씬 짧아서 특이한 자세를 취하게 되었다. 뒷다리는 대퇴골 (넓적다리뼈)은 몸에서 가장 긴 요소로, 알려진 가장 큰 대퇴골은 근위에서 원위 끝까지 665mm이다. 발은 3개의 발가락으로 끝났으며, 모두 발굽과 같은 발톱이 있었다.[15][16]
'''피부 갑옷'''
켄트로사우루스는 일반적인 검룡류와 마찬가지로, 광범위한 골편(피부의 뼈 구조)을 가지고 있었으며, 작은 판과 다양한 모양의 가시를 포함했다. 판은 가운데 부분이 두꺼워 마치 변형된 가시와 같았다. 가시와 판은 각질로 덮여 있었을 것이다. 꼬리 끝 부분과 연결된 짝을 이룬 가시가 발견되었으며, 여러 가시가 연결된 꼬리의 경로를 따라 짝을 이루어 규칙적으로 배치된 것으로 보인다.
Hennig과 Janensch는 피부 갑옷 요소를 네 가지 뚜렷한 유형으로 분류하면서, 앞쪽의 짧고 평평한 판이 뒤쪽으로 갈수록 길고 뾰족한 가시로 점차 바뀌는 것을 인식했으며, 이는 15쌍의 가시가 전체 몸통을 따라 끊김 없이 분포되었음을 시사했다. 각 유형의 골편이 좌우 대칭으로 발견되었기 때문에, 모든 유형의 골편이 동물의 등 뒤를 따라 두 줄로 분포되었을 가능성이 높다. 이는 스테고사우루스와 뚜렷한 대조를 이룬다. 강하게 비대칭적이며 매우 넓은 기저부를 가진 가시 유형이 하나 있는데, 뼈의 형태 때문에 고전적인 재구성은 이를 엉덩이에 위치시켰지만, 많은 최근 재구성은 이를 어깨에 위치시키는데, 그 이유는 중국 검룡류인 기가스피노사우루스와 화양고사우루스에서도 비슷한 모양의 가시가 어깨에 존재했던 것으로 알려져 있기 때문이다.
스테고사우루스와 근연종이지만, 크기, 갑옷 형태, 몸의 유연성에서 차이가 인정된다. 크기는 스테고사우루스가 몸길이 7.4미터, 몸무게 3,500킬로그램에 달하는 데 비해 켄트로사우루스는 몸길이 2.5 - 5미터, 몸무게 400킬로그램 - 1.5톤으로 훨씬 소형이다.[28] 등 쪽에 있는 골판은 스테고사우루스에서는 마름모꼴 골판이 2열로 어긋나게 배치되어 있지만, 켄트로사우루스에서는 2열 대칭으로 배치되어 있으며, 게다가 그 골판은 유럽의 렉소비사우루스(Lexovisaurus) 등과 마찬가지로 허리 부근부터 가늘게 뻗은 가시 모양으로 변해 있다. 스테고사우루스의 골판이 체온 조절 기능을 가지고 있었던 데 반해, 켄트로사우루스의 골판과 가시의 용도는 오로지 방어에만 사용되었던 것으로 보인다. 또한 켄트로사우루스의 허리 부근에는 굽어진 날카로운 가시가 있다. 이 가시에 대해서는, 투오장고사우루스(Tuojiangosaurus) 등 중국에서 발견된 다른 종에서는 어깨에 붙어 있었다는 증거가 얻어졌기 때문에, 켄트로사우루스의 복원에 대해서도 재검토해야 한다는 의견이 있다.
스테고사우루스의 척추뼈를 특징짓는 허리부터 꼬리까지의 척추뼈의 현저한 극돌기가 켄트로사우루스에는 없었다. 스테고사우루스는 강인한 근육으로 인해 뒷다리만으로 일어서서 나무 잎을 먹을 수도 있었지만, 켄트로사우루스에게는 불가능했던 것으로 보인다. 발의 다른 부분에 대한 대퇴골 길이의 비율이 커서 이 공룡이 느리고 비활발한 공룡이었음을 보여준다.
두개골에 대해서는 단편적인 화석밖에 얻어지지 않았지만, 근연종으로부터의 추정에 따르면 두개골은 작고 가늘고 긴 형태이며, 이를 잃고 대신 각질의 부리를 가진 주둥이가 있었다고 여겨진다.[28]
2. 1. 골격
스테고사우루스와 같은 속에 속하지만, 크기나 몸의 유연성, 갑옷 등의 형태를 볼 때 다소 차이가 있다. 켄트로사우루스의 몸 전체 크기는 약 2.5~5m, 체중은 400kg~1.5t이었을 것으로 추정되어 스테고사우루스보다는 소형으로 보인다.[28]
''켄트로사우루스 아에티오피쿠스''는 비교적 작은 검룡류로, 몸길이는 4~4.5m에 달하고 몸무게는 700~1600kg 정도였다.[3] 일부 표본을 보면 비교적 더 큰 개체도 존재했을 수 있다. 이 표본들은 발달된 주두돌기 측면에서 일부 ''스테고사우루스'' 표본과 유사하다.[4]
''켄트로사우루스''의 긴 꼬리는 질량 중심이 사족 보행 동물로서는 이례적으로 뒤쪽에 위치하게 한다. 질량 중심은 엉덩이 바로 앞에 위치하는데, 이것은 보통 이족 보행 공룡에서 볼 수 있는 위치이다. 그러나 ''켄트로사우루스''의 넙다리뼈는 일반적인 이족 보행 공룡과는 달리 곧게 뻗어 있으며, 이는 다리가 곧고 수직으로 뻗은 자세를 나타낸다. 따라서 뒷다리는 긴 엉덩뼈에 붙어 있는 거대한 허벅지 근육의 힘을 받았지만, 동물을 혼자 지탱하지 못했으며, 매우 튼튼한 앞다리가 체중의 10~15%를 지탱했다. ''켄트로사우루스''나 ''스테고사우루스''의 골판(피부의 뼈 구조)은 질량 중심에 큰 영향을 미치지 않아, 이들은 무장했음에도 불구하고 이동성을 유지할 수 있었다. 뒷다리의 허벅지 근육은 매우 강력하여, ''켄트로사우루스''가 뒷다리와 꼬리로 삼각대 자세를 취할 수 있었다.[5]
길고 좁은 두개골은 몸에 비해 작았다. 코와 눈 사이의 구멍인 작은 안와전개구가 있었지만, 현존하는 악어에서는 사라졌다. 두개골의 낮은 위치는 ''켄트로사우루스''가 낮게 자라는 식물을 먹었을 가능성을 시사한다. 이러한 해석은 앞니가 없고, 그 대신 뿔로 된 부리 또는 구치로 대체되었을 가능성이 있다는 점에서 뒷받침된다. 부리가 턱의 대부분을 따라 뻗어 있어 검룡류의 볼이 없을 수 있다.[8] ''켄트로사우루스''는 ''스테고사우루스''와 같이 광범위한 부리를 가졌을 가능성은 낮으며, 대신 턱 끝에 제한된 부리를 가졌을 것이다.[9][10]
하악골 (아래턱)에서 ''켄트로사우루스''는 불완전한 오른쪽 치골만 알려져 있다.[12] 깊은 치골은 ''스테고사우루스''와 거의 동일한 모양이다. 이빨은 일반적인 검룡류 이빨로 작고 넓어진 바닥과 수직 홈이 있다. 검룡류 이빨은 작고 삼각형이며 평평했다. 마모면은 그들이 음식을 갈았다는 것을 보여준다.[13]
[[파일:KentrosaurusDorsalVertebra.jpg|left|thumb|옆면 및 앞면에서 본 ''켄트로사우루스''의 등(뒷면) 척추.]
목은 13개의 목 척추로 구성되었으며, 축추가 뒤따른다. 등뼈는 13개의 등 척추로 구성된다. 천골(척추가 있는 골반 부분)은 6개의 융합된 중심을 이루며, 천골 늑골의 등쪽 부분과 융합되어 단단한 천골판을 형성한다. 미추 (꼬리) 척추는 29개이지만, 27-29개는 사고마이저 (꼬리 가시)에 부착하기 위해 함께 융합된다. ''켄트로사우루스''는 꼬리에서 대부분의 공룡과 달리 하위 평행하게 뻗어 있지 않은 척추의 많은 돌기로 인해 스테고사우루스의 다른 구성원과 구별할 수 있다.
[[파일:KentrosaurusSacrum(HMNSt439).jpg|thumb|배면에서 본 ''켄트로사우루스''의 천골.]
어깨뼈는 아랫부분이 사각형이며 튼튼한 날개를 가지고 있다. 오훼돌기는 반원형이다.[14] 앞다리는 튼튼한 뒷다리보다 훨씬 짧아서 특이한 자세를 취하게 되었다. 뒷다리는 대퇴골 (넓적다리뼈)은 몸에서 가장 긴 요소로, 알려진 가장 큰 대퇴골은 근위에서 원위 끝까지 665mm이다. 발은 3개의 발가락으로 끝났으며, 모두 발굽과 같은 발톱이 있었다.[15][16]
스테고사우루스와 근연종이지만, 크기, 갑옷 형태, 몸의 유연성에서 차이가 인정된다. 크기는 스테고사우루스가 몸길이 7.4미터, 몸무게 3,500킬로그램에 달하는 데 비해 켄트로사우루스는 몸길이 2.5 - 5미터, 몸무게 400킬로그램 - 1.5톤으로 훨씬 소형이다.[28] 켄트로사우루스의 골판과 가시의 용도는 오로지 방어에만 사용되었던 것으로 보인다.
스테고사우루스의 척추뼈를 특징짓는 허리부터 꼬리까지의 척추뼈의 현저한 극돌기가 켄트로사우루스에는 없었다. 스테고사우루스는 강인한 근육으로 인해 뒷다리만으로 일어서서 나무 잎을 먹을 수도 있었지만, 켄트로사우루스에게는 불가능했던 것으로 보인다.
두개골에 대해서는 단편적인 화석밖에 얻어지지 않았지만, 근연종으로부터의 추정에 따르면 두개골은 작고 가늘고 긴 형태이며, 이를 잃고 대신 각질의 부리를 가진 주둥이가 있었다고 여겨진다.[28]
2. 2. 피부 갑옷
켄트로사우루스는 일반적인 검룡류와 마찬가지로, 광범위한 골편(피부의 뼈 구조)을 가지고 있었으며, 작은 판과 다양한 모양의 가시를 포함했다. 판은 가운데 부분이 두꺼워 마치 변형된 가시와 같았다. 가시와 판은 각질로 덮여 있었을 것이다. 꼬리 끝 부분과 연결된 짝을 이룬 가시가 발견되었으며, 여러 가시가 연결된 꼬리의 경로를 따라 짝을 이루어 규칙적으로 배치된 것으로 보인다.Hennig과 Janensch는 피부 갑옷 요소를 네 가지 뚜렷한 유형으로 분류하면서, 앞쪽의 짧고 평평한 판이 뒤쪽으로 갈수록 길고 뾰족한 가시로 점차 바뀌는 것을 인식했으며, 이는 15쌍의 가시가 전체 몸통을 따라 끊김 없이 분포되었음을 시사했다. 각 유형의 골편이 좌우 대칭으로 발견되었기 때문에, 모든 유형의 골편이 동물의 등 뒤를 따라 두 줄로 분포되었을 가능성이 높다. 이는 스테고사우루스와 뚜렷한 대조를 이룬다. 강하게 비대칭적이며 매우 넓은 기저부를 가진 가시 유형이 하나 있는데, 뼈의 형태 때문에 고전적인 재구성은 이를 엉덩이에 위치시켰지만, 많은 최근 재구성은 이를 어깨에 위치시키는데, 그 이유는 중국 검룡류인 기가스피노사우루스와 화양고사우루스에서도 비슷한 모양의 가시가 어깨에 존재했던 것으로 알려져 있기 때문이다.
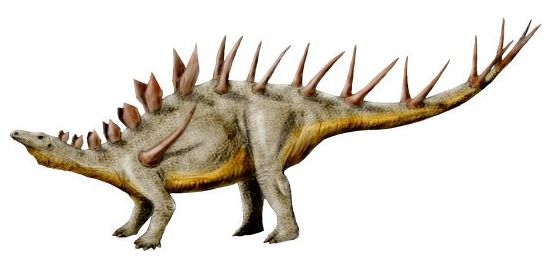
스테고사우루스와 근연종이지만, 갑옷 형태에서 차이가 확인된다. 스테고사우루스는 마름모꼴 골판이 2열로 어긋나게 배치되어 있지만, 켄트로사우루스에서는 2열 대칭으로 배치되어 있으며, 허리 부근부터 가늘게 뻗은 가시 모양으로 변해 있다. 스테고사우루스의 골판이 체온 조절 기능을 가지고 있었던 데 반해, 켄트로사우루스의 골판과 가시의 용도는 오로지 방어에만 사용되었던 것으로 보인다. 또한 켄트로사우루스의 허리 부근에는 굽어진 날카로운 가시가 있다. 이 가시에 대해서는, 투오장고사우루스 등 중국에서 발견된 다른 종에서는 어깨에 붙어 있었다는 증거가 있기 때문에, 켄트로사우루스의 복원에 대해서도 재검토해야 한다는 의견이 있다.
3. 발견 및 연구사

''켄트로사우루스''(Kentrosaurus)의 첫 번째 화석은 1909년 독일 텐다구루 탐험대에 의해 발견되었으며, 1910년 7월 24일 탐험대장 베르너 야넨슈에 의해 검룡류에 속하는 것으로 인식되었고, 1915년 독일 고생물학자 에드윈 헤닝에 의해 기술되었다. ''켄트로사우루스''라는 이름은 헤닝이 지었으며, "뾰족한 점" 또는 "가시"를 의미하는 그리스어 ''kentrongrc-Latn''/κέντρονgrc과 "도마뱀"을 의미하는 ''saurosgrc-Latn''/σαῦροςgrc에서 유래되었다. 헤닝은 종명으로 아프리카에서 발견된 것을 나타내기 위해 ''aethiopicus''를 추가했다.
설명 직후, 검룡의 이름에 대한 논란이 일어났는데, 이 이름은 각룡류인 ''켄트로사우루스''(Centrosaurus)와 매우 유사했기 때문이다. 생물 명명법 규칙에 따르면, 동음이의어를 금지하므로 두 동물에게 동일한 이름을 부여할 수 없다. 헤닝은 1916년에 자신의 검룡 이름을 "뾰족한 꼬리 도마뱀"을 의미하는 ''켄트루로사우루스''(Kentrurosaurus)로 변경했고, 헝가리 고생물학자 프란츠 노프차는 같은 해에 속명을 "창을 가진 도마뱀"을 의미하는 ''도리포로사우루스''(Doryphorosaurus)로 변경했다. 개명이 필요했다면, 헤닝의 것이 우선권을 가졌을 것이다. 그러나 철자가 다르기 때문에 ''도리포로사우루스''와 ''켄트루로사우루스''는 불필요한 대체 이름이다; ''켄트로사우루스''는 ''켄트루로사우루스''와 ''도리포로사우루스''가 주니어 객관적 동의어가 되면서 속명으로 유효한 이름을 유지하고 있다.[11]
완전한 개체는 발견되지 않았지만, 거의 완전한 꼬리, 엉덩이, 여러 등뼈 및 한 개체의 일부 사지 골격을 포함하여 일부 재료가 함께 발견되었다. 이것들은 야넨슈가 자연사 박물관에 설치한 마운트의 핵심을 형성한다. 이 마운트는 2006/2007년 박물관 개조 기간 동안 해체되었고, Research Casting International에 의해 개선된 자세로 다시 설치되었다. 뇌 케이스와 척추를 포함한 다른 일부 자료는 제2차 세계 대전 중에 잘못 놓이거나 파괴된 것으로 생각되었다. 그러나 잃어버린 것으로 추정되었던 모든 두개골 재료는 나중에 지하실 옷장의 서랍에서 발견되었다.
1909년 이후, ''켄트로사우루스'' 유해는 ''미틀레 자우리어슈텐''(Mittlere Saurierschichten, 중부 사우리아층)의 네 개 채석장과 ''오베레 자우리어슈텐''(Obere Saurierschichten, 상부 사우리아층)의 한 개 채석장에서 발견되었다. 네 번의 현장 조사 기간 동안 독일 탐험대는 약 50마리의 개체에 속하는 1200개 이상의 ''켄트로사우루스'' 뼈를 발견했는데, 그 중 많은 수가 제2차 세계 대전 중에 파괴되었다. 오늘날, 거의 모든 남은 재료는 베를린의 자연사 박물관에 보관되어 있으며(약 350개의 표본이 남아 있음), 튀빙겐 대학교 지구과학 연구소 박물관에는 복합 마운트가 보관되어 있으며, 약 50%가 원래 뼈이다.
원래 설명에서 헤닝은 모식표본을 지정하지 않았다. 그러나 1925년에 ''켄트로사우루스''의 골학, 체계적 위치 및 고생물학에 대한 상세한 모노그래피에서, 헤닝은 오늘날 'MB.R.4800.1~MB.R.4800.37'로 등록된 가장 완전한 부분 골격을 렉토타입(lectotype, 신표본)으로 선택했다.[15][1] 이 자료에는 거의 완전한 꼬리 척추, 등뼈 여러 개, 5개의 천추 척추와 양쪽 장골, 양쪽 넙다리뼈 및 자뼈가 포함되어 있으며, 독일 베를린의 자연사 박물관에 마운트된 골격에 포함되어 있다. 기준 산지는 텐다구루 언덕 북쪽에 있는 탄자니아의 킨도페이다.[1]
헤닝이 이미 렉토타입을 정의했다는 사실을 알지 못한 채, 피터 갤턴은 헤닝의 1915년 설명에 그림으로 나와 있는 자료에서 MB.R.1930과 MB.R.1931이라는 두 개의 등뼈를 '모식표본'으로 선택했다. 이러한 모식표본의 정의는 헤닝의 선택이 우선권을 갖기 때문에 유효하지 않다. 2011년, 하인리히 맬리슨은 헤닝이 1915년에 알고 있던 모든 자료, 즉 헤르만 헥이 마지막 독일 발굴을 완료한 1912년 이전에 발견된 모든 뼈는 준렉토타입이며 MB.R.4800이 올바른 렉토타입이라고 명확히 밝혔다.[2]
1909년 - 1912년의 독일에 의한 독일령 동아프리카 원정은 몇몇 새로운 공룡 종의 발견을 가져왔다. 이 원정에 참여한 세 명의 고생물학자 중 한 명인 에드윈 헤니히영어가 1915년에 켄트로사우루스를 기재했다. 거의 완전한 골격이 복원되어 베를린 대학교의 훔볼트 자연사 박물관에 전시되었지만, 박물관이 제2차 세계 대전 중 폭격을 받아 대부분 소실되었다.
3. 1. 분류
검룡의 골판과 가시는 기저 갑옷룡류의 특징인 낮은 융기된 골반에서 진화했다.[17] 피터 갤턴 (2019)은 인도의 하부 쥐라기 (시네뮤리안 - 플리엔스바키안) 하부 코타 지층에서 발견된 갑옷 공룡의 판을 곡룡류 화석으로 해석하며, 이는 곡룡류와 검룡류 모두 초기 전기 쥐라기에 기원했을 가능성을 시사한다.[18]대부분의 검룡류 공룡은 쥐라기 후반과 백악기 초기에 살았던 검룡과에 속하며, 폴 세레노는 ''스테고사우루스''보다 ''화양고사우루스''와 더 밀접하게 관련된 모든 검룡류로 정의했다.[19] 이 그룹은 북반구, 아프리카, 그리고 아마도 남아메리카에 걸쳐 널리 분포되어 있다.[20] 아르헨티나 추부트주에서 발견된 남아메리카 유해는 부분적인 상완골만 있지만, ''켄트로사우루스''와 매우 유사하며 둘 다 후기 쥐라기에 해당한다. 계통 분석에서 추부트 검룡류는 기저 검룡류로서 ''켄트로사우루스''와 다분지 분류로 복원되어 밀접한 관련이 있음을 시사한다.[20]
헤닝은 1915년 ''켄트로사우루스''를 검룡과에 할당했다. 이는 현대의 분지학적 분석으로 확인되었지만, 1915년 당시 검룡과는 현재 곡룡류로 분류되는 일부 분류군을 포함하는 훨씬 더 포괄적인 개념이었다. ''켄트로사우루스''는 1980년대까지 전형적인 "원시적인" 검룡류로 간주되었지만, 최근 분석에서는 ''켄트로사우루스''가 일반적으로 이 과에서 가장 기저에 있는 속 중 하나이지만 검룡과에 속한다는 것을 일관되게 발견했다.[21][20][22] ''켄트로사우루스''는 다른 검룡과에서 보이지 않지만 기저 검룡류에서 보이는 많은 특징을 가지고 있다.
''켄트로사우루스''의 기준종이자 유일하게 인정되는 종은 헤닝이 1915년에 명명한 ''켄트로사우루스 에티오피쿠스''이다. 찰스 W. 길모어가 1914년에 ''스테고사우루스 롱기스피누스''로 명명한 와이오밍의 단편적인 화석 자료는 1993년에 ''켄트로사우루스''의 북미 종으로 분류되었으나,[23] 고생물학계에서 받아들여지지 않았고, ''S. 롱기스피누스''는 자체 속인 ''알코바사우루스''에 할당되었다.[24][25]
4. 생태
''켄트로사우루스''는 현재 탄자니아인 후기 쥐라기 텐다구루 지층에서 서식했다. 주요 ''켄트로사우루스'' 채석장은 킴메리지안 상부에 해당하는 중부 사우리아 지층에 위치해 있었다. 일부 유해는 티토니안에 해당하는 상부 사우리아 지층에서 발견되었다. 2012년부터 킴메리지안과 티토니안 사이의 경계는 1억 5210만 년 전으로 측정되었다.
텐다구루 생태계는 주로 3가지 유형의 환경으로 구성되었다. 얕은 석호와 같은 해양 환경, 조수 갯벌과 낮은 해안 환경, 그리고 초목이 우거진 내륙 환경이었다. 해양 환경은 파고대 위에 존재했으며 규질쇄설암 및 사상암 장벽 뒤에 있었다. 염분 수준의 변화가 거의 없었고 조수와 폭풍을 겪었던 것으로 보인다. 해안 환경은 기수 해안 호수, 연못 및 웅덩이로 구성되었다. 이 환경은 식물이 거의 없었으며, 초식 공룡들이 주로 가뭄 기간 동안 방문했을 것이다. 초목이 우거진 내륙은 구과식물이 지배적이었다. 전반적으로 후기 쥐라기 텐다구루 기후는 아열대에서 열대 기후였으며 계절성 강우와 뚜렷한 건조 기간이 있었다. 초기 백악기 동안 텐다구루는 더욱 습해졌다. 텐다구루 지층은 해양 사이 지층을 제외하고 모리슨 층과 유사하다.
''켄트로사우루스''는 동족의 조반목 공룡인 ''디살로토사우루스''와 공존했을 것이며, 용각류인 ''기라파티탄'', ''디크라에오사우루스'', ''자넨스키아'', ''텐다구리아'', ''토르니에리아'', 수각류인 "알로사우루스", "세라토사우루스", "토르보사우루스", ''엘라프로사우루스'', ''베테루프리스티사우루스'', ''오스타프리카사우루스'' 그리고 익룡인 ''텐다구리프테루스''와도 공존했을 것이다. 텐다구루에 서식했던 다른 생물체로는 산호, 극피동물, 두족류, 이매패류, 복족류, 십각류, 상어, 신기어류, 악어류, 그리고 ''브란카테룰룸''과 같은 작은 포유류가 있었다.
북아메리카 대륙 로키 산맥의 모리슨 층에서 발견된 스테고사우루스와 아프리카 대륙 탄자니아의 텐다구루 층에서 발견된 켄트로사우루스가 근연 관계에 있다는 것은, 양자의 공통 조상이 하나의 대륙 판게아 (이후 북반부가 로라시아 대륙, 남반부가 곤드와나 대륙으로 분열)에 서식했음을 나타낸다. 반면에, 양자의 상이점은 같은 조상을 가진 동물이 분열된 각 대륙에서 각기 다른 진화 과정을 거쳤음을 보여준다. 또한, 이 두 지층의 유사성은 브라키오사우루스류에서도 현저하게 나타난다.
'''식성'''
모든 조반류 공룡과 마찬가지로, ''켄트로사우루스''는 초식동물이었다. 먹이는 거의 씹지 않고 큰 덩어리로 삼켰을 것이다. 검룡류의 식단에 대한 한 가설은 이들이 지면 근처에서 잎과 낮은 곳에 열리는 과실을 먹는, 다양한 비(非) 개화 식물을 섭취하는 저지대 섭취자였을 것이라고 본다.[2] ''켄트로사우루스''는 네 발로 설 때 1.7m 높이까지 먹이를 먹을 수 있었다. 또한 뒷다리로 일어서서 나무의 더 높은 곳에 있는 식물을 섭취하는 것도 가능했을 것이다.
질량 중심이 뒷다리에 가까이 있어, 이 공룡은 일어설 때 스스로를 지탱할 수 있었을 것이다. 엉덩이는 약 60도 정도의 수직 몸통 회전을 허용했을 가능성이 있으며, 꼬리는 이 움직임을 막지 않도록 완전히 들어 올리거나, 지면에 닿을 수 있도록 충분한 곡률을 가졌을 것이다. 따라서 추가적인 지지력을 제공했을 수 있지만, 이러한 유연성 때문에 실제로 얼마나 많은 지지력이 제공되었는지는 확실하지 않다. 로버트 토마스 배커가 제안했던 것처럼 "세 번째 다리" 역할을 할 정도로 뻣뻣하지는 않았을 것이다. 이러한 자세에서 ''켄트로사우루스''는 3.3m 높이의 먹이를 섭취할 수 있었을 것이다.
'''방어'''
꼬리에는 최소 40개의 꼬리 척추가 있었기 때문에, 매우 유연했다. 180도의 아크로 휘두를 수 있어서 뒤쪽 반원을 모두 커버할 수 있었을 것이다. 꼬리 끝의 휘두르는 속도는 시속 50km에 달했을 수 있으며, 계속되는 빠른 휘두르기는 공격자의 피부를 찢거나 연조직을 찔러 갈비뼈나 얼굴 뼈를 부러뜨릴 수 있었을 것이다. 더 직접적인 타격은 둔탁한 외상으로 인해 스파이크 측면에서 다리의 튼튼한 긴 뼈가 부러지는 결과를 초래했을 것이며, 이러한 공격은 작고 중간 크기의 수각류를 불구로 만들었을 것이고, 심지어 큰 수각류에게도 약간의 피해를 입혔을 수 있다. ''켄트로사우루스''의 방어 행동에 대한 초기 해석에는 동물이 현대의 고슴도치처럼 척추로 공격자를 관통하기 위해 뒤로 돌진했을 수 있다는 제안이 포함되었다.
''켄트로사우루스''는 다른 공룡처럼 앞다리를 세운 자세로 서 있었을 가능성이 높지만, 방어할 때는 엎드린 자세를 취했을 것으로 추정된다. 목은 유연해서 주둥이로 몸 옆을 짚고 뒤를 볼 수 있어서 포식자를 계속 볼 수 있었다. 또한, 질량 중심이 뒤쪽에 위치한 것은 빠른 이동에는 유리하지 않았을 수 있지만, 팔로 옆으로 밀어 엉덩이 주변을 빠르게 회전시켜 꼬리를 공격자를 향하게 할 수 있었다는 것을 의미한다. 그럼에도 불구하고 ''켄트로사우루스''는 무적이 아니었다. 빠른 포식자는 꼬리가 지나갈 때 꼬리 기저부(충격 속도가 훨씬 낮을 것)까지 도달할 수 있었고, 꼬리가 휘둘러지지 않으면 목과 몸 윗부분이 보호받지 못했을 것이다. ''켄트로사우루스''의 포식 성공은 집단 사냥이 필요했을 수 있다. ''스테고사우루스''의 더 튼튼한 스파이크에 비해 ''켄트로사우루스''의 더 얇은 스파이크는 구부러질 위험이 더 컸다.
'''성적 이형성'''
홀리 바든과 수재너 메이드먼트는 켄트로사우루스의 대퇴골(넙다리뼈) 크기 비율 차이를 통해 성적 이형성을 보였을 것이라고 추정했다. 대퇴골은 튼튼한 형태와 가늘어진 형태로 나뉘며, 발생 비율은 2:1이었다. 더 높은 비율을 가진 튼튼한 형태의 동물이 암컷이었을 가능성이 높다고 여겨지며, 이는 켄트로사우루스 수컷이 여러 암컷과 짝짓기를 하는 사회 구조를 가졌을 것이라는 추정을 가능하게 한다. 이러한 행동은 다른 척추동물에서도 발견된다.
하지만 연구된 표본들이 같은 장소에서 죽었지만, 갑작스러운 대량 죽음은 아니었기 때문에 단일 무리나 동시대 집단을 나타내지 않는다는 문제가 제기된다. 튼튼한 동물이 화석화되거나 발견될 가능성이 더 높아 결과가 왜곡되었을 수 있다. 1982년 갈턴의 연구에서는 켄트로사우루스와 다켄트루루스의 천골 늑골 수의 개별 차이가 이형성의 지표일 수 있다고 제안했다. 암컷은 장골에 연결된 첫 번째 천골 척추뼈를 포함하여 천골 늑골 한 쌍을 더 가졌을 것으로 추정된다.
4. 1. 식성
모든 조반류 공룡과 마찬가지로, ''켄트로사우루스''는 초식동물이었다. 먹이는 거의 씹지 않고 큰 덩어리로 삼켰을 것이다. 검룡류의 식단에 대한 한 가설은 이들이 지면 근처에서 잎과 낮은 곳에 열리는 과실을 먹는, 다양한 비(非) 개화 식물을 섭취하는 저지대 섭취자였을 것이라고 본다.[2] ''켄트로사우루스''는 네 발로 설 때 1.7m 높이까지 먹이를 먹을 수 있었다. 또한 뒷다리로 일어서서 나무의 더 높은 곳에 있는 식물을 섭취하는 것도 가능했을 것이다.질량 중심이 뒷다리에 가까이 있어, 이 공룡은 일어설 때 스스로를 지탱할 수 있었을 것이다. 엉덩이는 약 60도 정도의 수직 몸통 회전을 허용했을 가능성이 있으며, 꼬리는 이 움직임을 막지 않도록 완전히 들어 올리거나, 지면에 닿을 수 있도록 충분한 곡률을 가졌을 것이다. 따라서 추가적인 지지력을 제공했을 수 있지만, 이러한 유연성 때문에 실제로 얼마나 많은 지지력이 제공되었는지는 확실하지 않다. 로버트 토마스 배커가 제안했던 것처럼 "세 번째 다리" 역할을 할 정도로 뻣뻣하지는 않았을 것이다. 이러한 자세에서 ''켄트로사우루스''는 3.3m 높이의 먹이를 섭취할 수 있었을 것이다.
4. 2. 방어
꼬리에는 최소 40개의 꼬리 척추가 있었기 때문에, 매우 유연했다. 180도의 아크로 휘두를 수 있어서 뒤쪽 반원을 모두 커버할 수 있었을 것이다. 꼬리 끝의 휘두르는 속도는 시속 50km에 달했을 수 있으며, 계속되는 빠른 휘두르기는 공격자의 피부를 찢거나 연조직을 찔러 갈비뼈나 얼굴 뼈를 부러뜨릴 수 있었을 것이다. 더 직접적인 타격은 둔탁한 외상으로 인해 스파이크 측면에서 다리의 튼튼한 긴 뼈가 부러지는 결과를 초래했을 것이며, 이러한 공격은 작고 중간 크기의 수각류를 불구로 만들었을 것이고, 심지어 큰 수각류에게도 약간의 피해를 입혔을 수 있다. ''켄트로사우루스''의 방어 행동에 대한 초기 해석에는 동물이 현대의 고슴도치처럼 척추로 공격자를 관통하기 위해 뒤로 돌진했을 수 있다는 제안이 포함되었다.
''켄트로사우루스''는 다른 공룡처럼 앞다리를 세운 자세로 서 있었을 가능성이 높지만, 방어할 때는 엎드린 자세를 취했을 것으로 추정된다. 목은 유연해서 주둥이로 몸 옆을 짚고 뒤를 볼 수 있어서 포식자를 계속 볼 수 있었다. 또한, 질량 중심이 뒤쪽에 위치한 것은 빠른 이동에는 유리하지 않았을 수 있지만, 팔로 옆으로 밀어 엉덩이 주변을 빠르게 회전시켜 꼬리를 공격자를 향하게 할 수 있었다는 것을 의미한다. 그럼에도 불구하고 ''켄트로사우루스''는 무적이 아니었다. 빠른 포식자는 꼬리가 지나갈 때 꼬리 기저부(충격 속도가 훨씬 낮을 것)까지 도달할 수 있었고, 꼬리가 휘둘러지지 않으면 목과 몸 윗부분이 보호받지 못했을 것이다. ''켄트로사우루스''의 포식 성공은 집단 사냥이 필요했을 수 있다. ''스테고사우루스''의 더 튼튼한 스파이크에 비해 ''켄트로사우루스''의 더 얇은 스파이크는 구부러질 위험이 더 컸다.
4. 3. 성적 이형성
홀리 바든과 수재너 메이드먼트는 켄트로사우루스의 대퇴골(넙다리뼈) 크기 비율 차이를 통해 성적 이형성을 보였을 것이라고 추정했다. 대퇴골은 튼튼한 형태와 가늘어진 형태로 나뉘며, 발생 비율은 2:1이었다. 더 높은 비율을 가진 튼튼한 형태의 동물이 암컷이었을 가능성이 높다고 여겨지며, 이는 켄트로사우루스 수컷이 여러 암컷과 짝짓기를 하는 사회 구조를 가졌을 것이라는 추정을 가능하게 한다. 이러한 행동은 다른 척추동물에서도 발견된다.하지만 연구된 표본들이 같은 장소에서 죽었지만, 갑작스러운 대량 죽음은 아니었기 때문에 단일 무리나 동시대 집단을 나타내지 않는다는 문제가 제기된다. 튼튼한 동물이 화석화되거나 발견될 가능성이 더 높아 결과가 왜곡되었을 수 있다. 1982년 갈턴의 연구에서는 켄트로사우루스와 다켄트루루스의 천골 늑골 수의 개별 차이가 이형성의 지표일 수 있다고 제안했다. 암컷은 장골에 연결된 첫 번째 천골 척추뼈를 포함하여 천골 늑골 한 쌍을 더 가졌을 것으로 추정된다.
5. 고생물지리학적 의의
''켄트로사우루스''는 현재 탄자니아인 후기 쥐라기 텐다구루 지층에서 서식했다. 주요 ''켄트로사우루스'' 채석장은 킴메리지안 상부에 해당하는 중부 사우리아 지층에 위치해 있었으며, 일부 유해는 티토니안에 해당하는 상부 사우리아 지층에서 발견되었다. 2012년부터 킴메리지안과 티토니안 사이의 경계는 1억 5210만 년 전으로 측정되었다.
텐다구루 생태계는 얕은 해양 환경, 해안 환경, 내륙 환경으로 구성되었다. 해양 환경은 파고대 위에 존재했으며 염분 변화가 거의 없었다. 해안 환경은 기수 해안 호수, 연못 등으로 구성되었고 식물이 거의 없었다. 내륙은 구과식물이 지배적이었다. 텐다구루 기후는 아열대에서 열대 기후였으며 계절성 강우와 건조 기간이 있었다. 초기 백악기 동안 텐다구루는 더욱 습해졌다. 텐다구루 지층은 해양 사이 지층을 제외하고 모리슨 층과 유사하다.
''켄트로사우루스''는 ''디살로토사우루스'', 용각류인 ''기라파티탄'', ''디크라에오사우루스'', ''자넨스키아'', ''텐다구리아'', ''토르니에리아'', 수각류인 "알로사우루스", "세라토사우루스", "토르보사우루스", ''엘라프로사우루스'', ''베테루프리스티사우루스'', ''오스타프리카사우루스'' 그리고 익룡인 ''텐다구리프테루스'' 등과 공존했을 것이다. 그 외에도 산호, 극피동물, 두족류, 이매패류, 복족류, 십각류, 상어, 신기어류, 악어류, 그리고 작은 포유류 등이 텐다구루에 서식했다.
북아메리카 대륙 로키 산맥의 모리슨 층에서 발견된 스테고사우루스와 아프리카 대륙 탄자니아의 텐다구루 층에서 발견된 켄트로사우루스가 근연 관계에 있다는 것은, 양자의 공통 조상이 판게아에 서식했음을 나타낸다. 양자의 상이점은 같은 조상을 가진 동물이 분열된 각 대륙에서 각기 다른 진화 과정을 거쳤음을 보여준다. 또한, 이 두 지층의 유사성은 브라키오사우루스류에서도 현저하게 나타난다.
6. 대중 문화
참조
[1]
논문
Ein aufgestelltes skelett des stegosauriers Kentrurosaurus aethiopicus E. Hennig aus den Tendaguru-schichten Deutsch-Ostafrikas
Palaeontographica-Supplementbände
1925
[2]
논문
The real lectotype of Kentrosaurus aethiopicus HENNIG, 1915
Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen
2011
[3]
간행물
Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage
2014-05-06
[4]
간행물
Two New Stegosaur Specimens from the Upper Jurassic Morrison Formation of Montana, USA
[5]
간행물
Osteoderm distribution has low impact on the centre of mass of stegosaurs
https://fr.copernicu[...]
2014-03-07
[6]
논문
Skull bones and endocranial casts of stegosaurian dinosaur Kentrosaurus Hennig, 1915 from Upper Jurassic of Tanzania, East Africa.
https://www.research[...]
Geologica et Palaeontologica
1988
[7]
논문
Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia)
Journal of Systematic Palaeontology
2008
[8]
간행물
The beaked jaws of stegosaurs and their implications for other ornithischians
[9]
간행물
Buccal soft anatomy in Lesothosaurus (Dinosauria: Ornithischia)
https://www.research[...]
[10]
서적
Tooth wear and possible jaw action of Scelidosaurus harrisonii Owen and a review of feeding mechanisms in other thyreophoran dinosaurs
Indiana University Press
2001
[11]
서적
Stegosauria
University of California Press
[12]
논문
Ein dentale von Kentrurosaurus aethiopicus Hennig
Palaeontographica-Supplementbände
1936
[13]
서적
Stegosauria: Hot Plates
Cambridge University Press
[14]
간행물
The Postcranial Skeleton of an Exceptionally Complete Individual of the Plated Dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, U.S.A.
2015-10-14
[15]
논문
Kentrurosaurus aethiopicus; die Stegosaurierfunde vom Tendaguru, Deutsch-Ostafrika
Palaeontographica-Supplementbände
1925
[16]
논문
The postcranial anatomy of stegosaurian dinosaur Kentrosaurus from the Upper Jurassic of Tanzania, East Africa
1982
[17]
서적
Scelidosaurus, the earliest complete dinosaur
Indiana University Press
2001
[18]
간행물
Earliest record of an ankylosaurian dinosaur (Ornithischia: Thyreophora): Dermal armor from Lower Kota Formation (Lower Jurassic) of India
[19]
논문
A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria
Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen
1998
[20]
간행물
First osteological record of a stegosaur (Dinosauria, Ornithischia) from the Upper Jurassic of South America
https://www.tandfonl[...]
2020-12-10
[21]
간행물
New stegosaurs from the Middle Jurassic Lower Member of the Shaximiao Formation of Chongqing, China
2022-03-30
[22]
간행물
A new phylogeny of Stegosauria (Dinosauria, Ornithischia)
https://onlinelibrar[...]
2017
[23]
간행물
The Origin and Evolution of the Stegosaurs
[24]
웹사이트
Evolution of the stegosaurs (Dinosauria; Ornithischia). Dinologia
http://dinoweb.narod[...]
[25]
문서
Natronasaurus longispinus, 100 years with another name
2014
[26]
간행물
North Africa's first stegosaur: Implications for Gondwanan thyreophoran dinosaur diversity
https://www.scienced[...]
2020-01-01
[27]
간행물
The socio-sexual behaviour of extant archosaurs: Implications for understanding dinosaur behaviour
http://doc.rero.ch/r[...]
[28]
서적
恐竜博物図鑑
新樹社
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com