케라토사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
케라토사우루스는 후기 쥐라기 시대에 살았던 수각류 공룡의 한 속으로, 특징적인 뿔과 골편을 가지고 있다. 1883년과 1884년에 미국에서 처음 발견되었으며, 이후 북아메리카, 유럽, 아프리카 등지에서 화석이 발견되었다. 케라토사우루스는 이족 보행을 했으며, 세 개의 손가락과 네 개의 손가락을 가진 독특한 특징을 가지고 있었다. 케라토사우루스의 뿔은 공격이나 방어보다는 과시용으로 사용되었을 가능성이 높으며, 먹이로는 다른 공룡이나 수생 동물을 사냥했을 것으로 추정된다. 케라토사우루스는 알로사우루스와 함께 모리슨 층에서 발견되며, 생태적 지위 경쟁에 대한 연구가 이루어지기도 했다.
더 읽어볼만한 페이지
- 1884년 기재된 화석 분류군 - 암모나이트
암모나이트는 멸종된 두족류로, 평면 나선형 껍데기와 격벽, 봉합선 등의 특징을 가지며, 고생대 데본기에 출현하여 중생대를 거쳐 백악기 말에 멸종했고, 화석은 지질 시대의 표준화석으로 사용된다. - 1884년 기재된 화석 분류군 - 메갈리브윌리아
메갈리브윌리아에 대한 정보가 부족하여 요약할 수 없다. - 포르투갈의 화석 - 알로사우루스
알로사우루스는 쥐라기 후기에 살았던 대형 수각류 공룡으로, 짧은 앞다리, 머리의 뿔, 갈고리 발톱을 특징으로 하며, 모리슨층에서 발견되었고, 다른 육식 공룡과 함께 먹이 사슬의 정점에 있었다. - 포르투갈의 화석 - 다켄트루루스
다켄트루루스는 후기 쥐라기 유럽에서 살았던 검룡류 공룡으로, 처음에는 오모사우루스라고 불렸으나 이름이 선점되어 다켄트루루스로 재명명되었고, 단편적인 화석을 통해 분포와 특징이 연구되고 있다. - 탄자니아의 화석 - 글로소프테리스
글로소프테리스는 혀 모양의 잎과 그물맥을 가지는 멸종된 종자 식물로, 곤드와나 대륙에 널리 분포하며 대륙 이동설을 뒷받침하는 화석 증거가 되었다. - 탄자니아의 화석 - 히파리온
히파리온은 마이오세 시대에 번성했던 세 개의 발가락을 가진 중간 크기의 멸종된 말의 한 종류로, 숲과 초원으로 이루어진 사바나 생물 군계에 서식하며 풀을 주식으로 섭취했고, 약 1100만 년 전 코르모히파리온에서 진화하여 구세계에서 기후 변화로 인해 멸종되었다.
| 케라토사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Ceratosaurus |
| 명명자 | 마시, 1884년 |
| 모식종 | †Ceratosaurus nasicornis |
| 모식종 명명자 | 마시, 1884 |
| 다른 종 | †?C. dentisulcatus 매드슨 & 웰스, 2000 †?C. magnicornis 매드슨 & 웰스, 2000 |
| 동의어 | 메갈로사우루스 nasicornis (마시, 1884 [원래 Ceratosaurus]) |
| 어원 | 그리스어 (뿔) + (도마뱀) |
| 크기 | |
| 생물학적 분류 | |
| 상목 | 공룡상목 |
| 목 | 용반목 |
| 목 계급 없음 | 진용반류 |
| 아목 | 수각아목 |
| 하목 | 케라토사우루스하목 |
| 과 | 케라토사우루스과 |
| 시대 | |
| 시기 | 쥐라기 후기 (김메르기지층 ~ 티토누스절), |
2. 역사
1909년부터 1913년까지 베를린 자연사 박물관의 독일 탐험대는 현재 탄자니아에 위치한 텐다구루 지층에서 다양한 공룡 화석을 발굴했다. 1920년, 독일 고생물학자 베르너 야넨슈는 채석장 "TL"에서 발견된 여러 개의 등뼈를 ''케라토사우루스'' sp. (종 불확실)로 분류했고, 1925년에는 채석장 "Mw"에서 발견된 조각난 유해를 바탕으로 ''케라토사우루스''의 새로운 종인 ''C. 로에클링기''를 명명했다.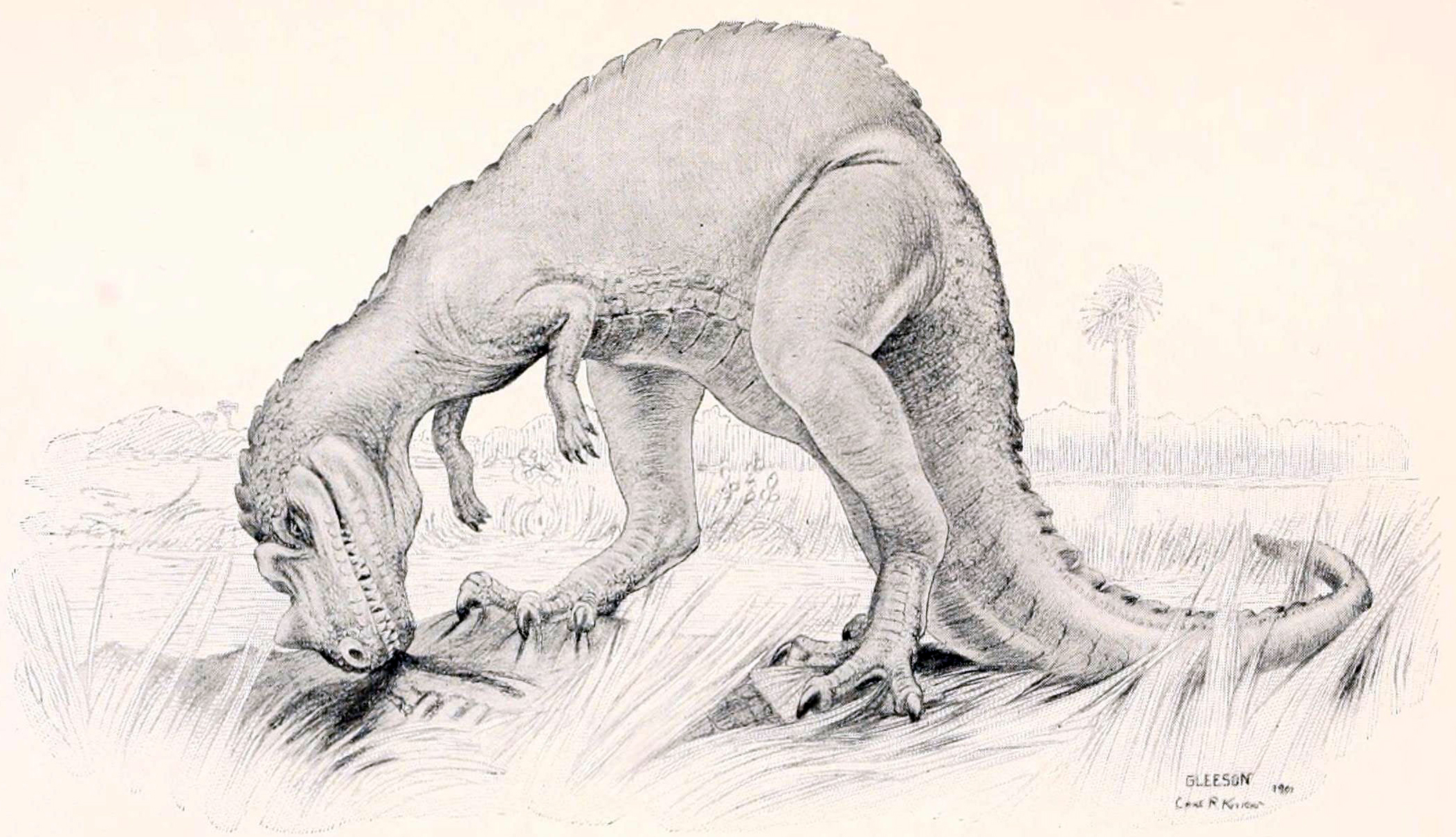
2000년 매드센과 웰스는 이러한 발견을 ''케라토사우루스''로 확인하고, 야넨슈가 원래 라브로사우루스의 한 종으로 묘사했던 여러 개의 이빨을 ''라브로사우루스'' (?) ''스테코비''로 귀속시켰다. 그러나 다른 저자들은 텐다구루에서 발견된 유해를 ''케라토사우루스''로 귀속시키는 것에 의문을 제기했다. 2011년, 라우후트는 ''C. 로에클링기''와 ''라브로사우루스'' (?) ''스테코비'' 모두 케라토사우루스과일 가능성이 있지만, 노멘 두비움 (의심스러운 이름)으로 지정했다.
1990년, 티모시 로우와 자크 고티에는 야넨슈가 1920년에 제안한 ''케라토사우루스 잉겐스''를 언급했지만, 야넨슈는 이 종을 ''메갈로사우루스''로 분류했기에 단순한 복사 오류일 수 있다. 2011년 라우후트는 ''메갈로사우루스 잉겐스''가 카르카로돈토사우루스과를 나타낼 가능성이 있음을 보여주었다.
2000년과 2006년, 옥타비오 마테우스 연구팀은 포르투갈 로리냐 지층에서 발견된 화석(ML 352)을 ''케라토사우루스''의 새로운 표본으로 묘사했다. 2006년 마테우스와 동료들은 이 표본을 ''케라토사우루스 덴티술카투스'' 종으로 분류했지만, 2008년 카라노와 섐슨은 ''케라토사우루스''로는 분류했지만, 특정 종으로의 분류는 불가능하다고 결론지었다. 2015년, 엘리자베테 말라파이아와 동료들은 ''C. 덴티술카투스''의 유효성에 의문을 제기하며, ''케라토사우루스'' aff. ''케라토사우루스 나시코르니스''로 분류했다.
스위스 무티에에서 발견된 단일 이빨은 매드센과 웰스에 의해 ''케라토사우루스'' sp. (종 불확실)로 분류되었다. 2008년, 마티아스 소토와 다니엘 페레아는 우루과이의 타쿠아렘보 지층에서 ''케라토사우루스''에서만 알려진 특징을 보이는 이빨을 묘사했다. 그러나 유해가 희소하여 ''케라토사우루스''로 분류하기는 어렵다고 강조했다. 2020년, 소토와 동료들은 동일 지층에서 추가적인 ''케라토사우루스'' 이빨을 묘사하여 이전의 해석을 뒷받침했다.[4]
2. 1. 발견
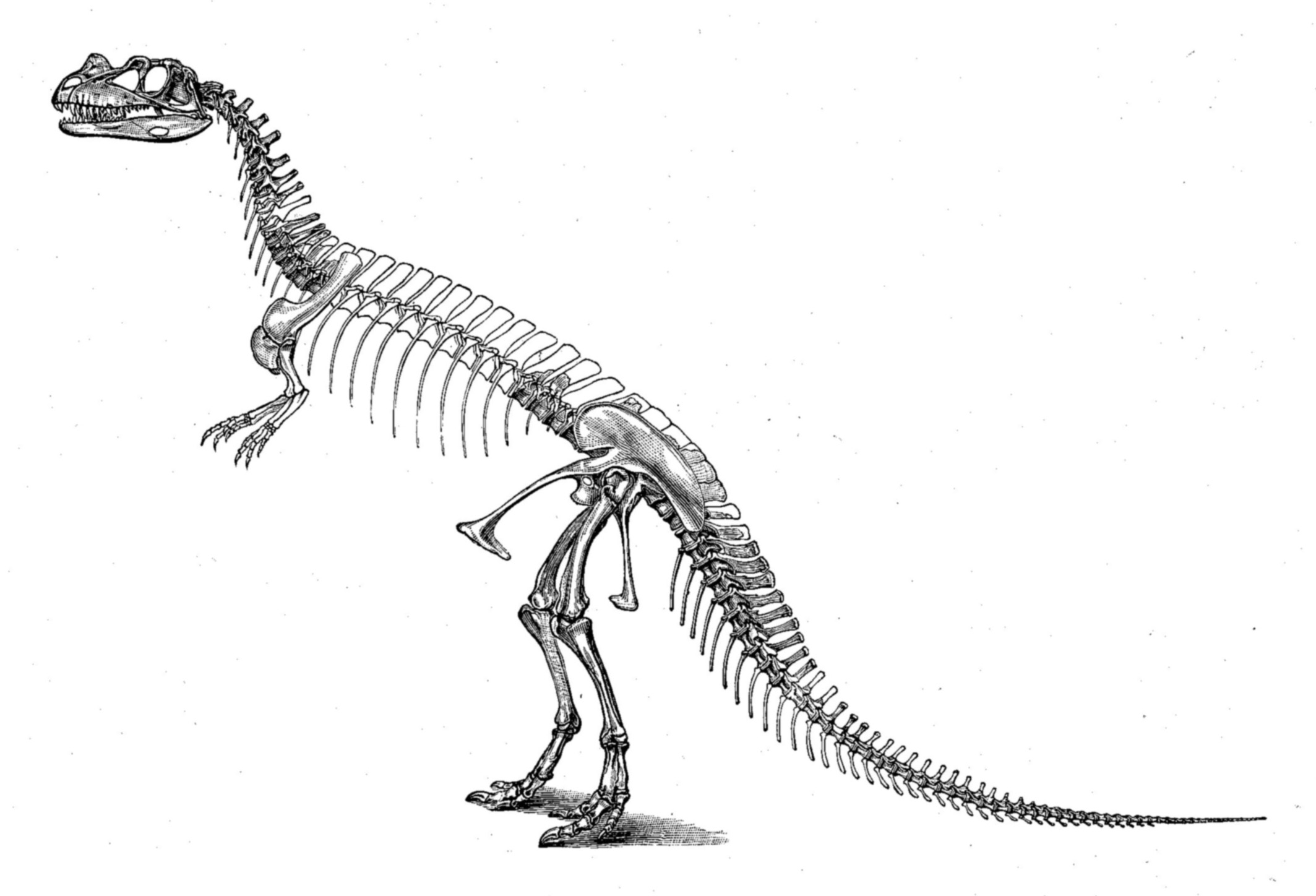
첫 번째 표본(홀로타입 USNM 4735)은 1883년과 1884년 농부 마셜 파커 펠치에 의해 발견, 발굴되었다. 뼈는 서로 연결된 상태로 발견되었으며 두개골을 포함하여 거의 완전했다. 다만, 척추뼈 일부, 몸통 마지막 갈비뼈를 제외한 모든 갈비뼈, 상완골(위팔뼈), 양손의 원위 손가락뼈, 오른쪽 팔 대부분, 왼쪽 다리 대부분, 발 대부분이 발견되지 않았다. 표본은 단단한 사암에 싸여 화석화 과정에서 두개골과 척추가 심하게 변형되었다. 발견 장소는 콜로라도주 캐넌시티 북쪽 가든 파크의 펠치 채석장 1이며, 모리슨 층에서 가장 풍부한 화석이 발견되는 곳 중 하나이다. 이 채석장에서는 ''케라토사우루스'' 발견 전에도 수많은 공룡 화석이 발견되었는데, 특히 1883년과 1884년에 거의 완전한 ''알로사우루스''(USNM 4734) 표본이 발견되었다.
발굴 후 표본은 코네티컷주 뉴헤이븐 피바디 자연사 박물관으로 옮겨져 마쉬에 의해 연구되었고, 1884년에 새로운 속과 종인 ''케라토사우루스 나시코르니스''로 명명되었다. ''케라토사우루스''는 "뿔 도마뱀"(그리스어 κερας/κερατος|케라스/케라토스|뿔grc와 σαυρος|사우로스|도마뱀grc에서 유래), ''나시코르니스''는 "코 뿔"(라틴어 ''nasus''—"코", ''cornu''—"뿔")을 뜻한다.[1] 이 표본은 당시 미국에서 발견된 가장 잘 알려진 수각류였다. 1898년과 1899년에, 이 표본은 마쉬가 처음 기술한 다른 많은 화석들과 함께 워싱턴 D.C. 국립 자연사 박물관으로 옮겨졌다. 이 자료 중 일부만 워싱턴에 도착했을 때 완전히 준비되었고, 이후 준비는 1911년부터 1918년 말까지 이어졌다. 뉴헤이븐에서 워싱턴으로의 포장 및 운송 과정에서 ''케라토사우루스'' 표본에 약간의 손상이 발생했다. 1920년, 찰스 길모어는 이 표본과 같은 채석장에서 발견된 거의 완전한 ''알로사우루스'' 표본을 포함, 뉴헤이븐에서 받은 다른 수각류 표본에 대한 광범위한 재기술을 발표했다.
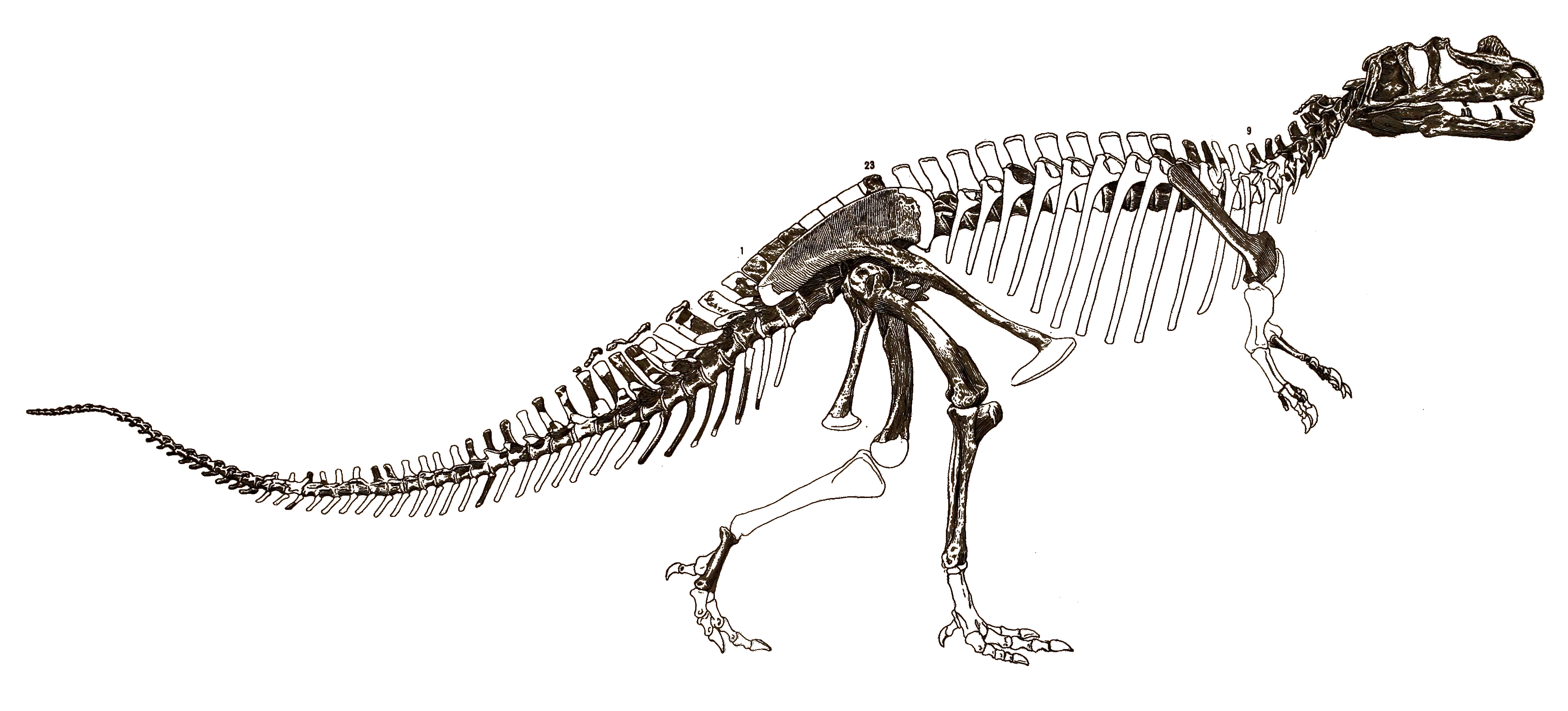
1892년 논문에서 마쉬는 ''케라토사우루스''의 첫 번째 골격 재구성을 발표했는데, 길이는 약 6.71m, 높이는 약 3.66m로 묘사되었다. 1920년 길모어는 이 재구성에서 몸통이 최소 6개의 등 척추뼈를 더 포함하여 너무 길게 묘사되었다고 지적했다. 이 오류는 1899년 찰스 R. 나이트 지시로 프랭크 본이 그린 최초의 생물 재구성을 포함, 여러 후속 출판물에서 반복되었지만 1920년까지 출판되지 않았다. 1901년에 더 정확한 생물 재구성이 나이트 감독하에 조셉 M. 글리슨에 의해 제작되었다. 홀로타입은 1910년과 1911년에 길모어에 의해 부조로 설치되어 국립 자연사 박물관에 전시되었다. 대부분의 초기 재구성은 ''케라토사우루스''를 꼬리가 땅에 끌린 채 직립 자세로 보여주지만, 길모어의 홀로타입 마운트는 시대를 앞서 하지가 아래 다리에 비스듬히 놓인 수평 자세와 꼬리가 땅에 닿지 않는 달리는 동물로 묘사했다. 화석의 강한 평탄화 때문에 길모어는 독립형 골격이 아닌 인공 벽 내의 부조로 설치했고, 뼈가 부분적으로 판에 박혀있어 과학적인 접근이 제한되었다. 2014년과 2019년 사이 박물관 공룡 전시 개조 과정에서 표본은 해체되어 판에서 분리되었다.[2][3] 2019년 개관 예정인 새 전시회에서는 마운트를 독립형 캐스트로 교체하고 원래 뼈는 박물관 컬렉션에 보관하여 과학자들이 자유롭게 접근할 수 있도록 할 계획이었다.[3]

1960년대 초, 제임스 매드슨 팀이 유타 주 클리블랜드-로이드 공룡 채석장에서 두개골(UMNH VP 5278)을 포함한 단편적인 골격을 발굴하면서 ''C. nasicornis'' 홀로타입 발견 이후 상당한 규모의 ''케라토사우루스'' 화석이 발견되었다. 이는 알려진 가장 큰 ''케라토사우루스'' 표본 중 하나이다. 1976년에는 고생물학자 랜스 에릭슨의 아들 토르 에릭슨이 콜로라도주 푸루이타 근처에서 두개골을 포함한 두 번째 연결된 표본(MWC 1)을 발견했다. 비교적 완전하지만 아래턱, 앞다리, 위장늑골이 없다. 두개골은 비교적 완전하지만 분리된 상태로 발견되었고 옆으로 심하게 납작해졌다. 큰 개체였지만 두개골 뼈 사이 봉합선이 융합되지 않아 아직 성체가 아니었다. 박물관 전시용으로 과학적이고 정확한 두개골 3차원 재구성은 개별 원본 뼈의 성형 및 주조, 변형 수정, 누락 부분 재구성, 뼈 주조물 조립, 원래 색상과 일치하도록 페인팅하는 복잡한 과정을 거쳤다.
푸루이타 표본과 클리블랜드-로이드 표본은 매드슨과 사무엘 폴 웰스가 2000년 모노그래프에서 설명했으며, 유타 표본은 ''C. dentisulcatus'', 콜로라도 표본은 ''C. magnicornis''로 명명되었다. 'dentisulcatus'는 앞턱뼈 이와 아래턱 처음 세 개 이 안쪽의 평행한 홈, 'Magnicornis'는 더 큰 코뿔을 가리킨다. 그러나 두 종의 유효성은 이후 간행물에서 의문시되었다. 2000년 브룩스 브리트와 동료들은 ''C. nasicornis'' 홀로타입이 어린 개체이며, 두 개의 더 큰 종은 단일 종의 성체 상태라고 주장했다. 2003년 올리버 라우후트와 2008년 매튜 카라노와 스콧 섐슨은 매드슨과 웰스가 인용한 해부학적 차이가 개체 발생(연령 관련) 또는 개별 변이를 나타낸다고 생각했다.
1992년 유타 주 무어 남동쪽 아게이트 베이슨 채석장에서 또 다른 표본(BYUVP 12893)이 발견되었지만 아직 설명되지 않았다. 이 표본은 속에서 가장 큰 것으로 여겨지며 두개골 앞쪽 절반, 일곱 개의 단편적 골반 등쪽 척추, 연결된 골반 및 천골을 포함한다. 1999년 브리트는 어린 개체의 ''케라토사우루스'' 골격 발견을 보고했다. 와이오밍 주 본 캐빈 채석장에서 발견되었으며, ''C. nasicornis'' 홀로타입보다 34% 작고 완전한 두개골과 완전한 골반을 포함한 골격의 나머지 부분 중 30%로 구성되어 있다.
이 다섯 개의 골격 발견 외에도, 단편적인 ''케라토사우루스'' 유해가 모리슨 층의 2, 4-6번 층서대에서 보고되었으며, 층의 주요 화석 유적지도 포함된다. 유타 주 공룡 국립 기념물에서는 고립된 오른쪽 앞턱뼈(DNM 972)가 발견되었다. 큰 어깨뼈(견갑골)가 와이오밍 주 코모 블러프에서 보고되었다. 또 다른 표본은 콜로라도 주 드라이 메사 채석장에서 나왔으며 왼쪽 견갑골과 척추 및 사지 뼈 조각을 포함한다. 콜로라도 주 마이게이트 무어 채석장에서는 이빨로 이 속을 알 수 있다.
2. 2. 분류 논란과 현재의 학설
베를린 자연사 박물관의 독일 탐험대가 1909년부터 1913년까지 탄자니아 텐다구루 지층에서 다양한 공룡 화석을 발굴했다. 1920년, 독일 고생물학자 베르너 야넨슈는 여러 개의 등뼈를 ''케라토사우루스'' sp. (종 불확실)로 분류했고, 1925년에는 ''케라토사우루스''의 새로운 종인 ''C. 로에클링기''를 명명했다.2000년 매드센과 웰스는 이러한 발견을 ''케라토사우루스''로 확인하고, 야넨슈가 라브로사우루스의 한 종으로 묘사했던 여러 개의 이빨을 ''라브로사우루스'' (?) ''스테코비''로 귀속시켰다. 그러나 다른 저자들은 텐다구루에서 발견된 유해를 ''케라토사우루스''로 귀속시키는 것에 의문을 제기했다. 2011년, 라우후트는 ''C. 로에클링기''와 ''라브로사우루스'' (?) ''스테코비'' 모두 케라토사우루스과일 가능성이 있지만, 노멘 두비움 (의심스러운 이름)으로 지정했다.
1990년, 티모시 로우와 자크 고티에는 야넨슈가 1920년에 제안한 ''케라토사우루스 잉겐스''를 언급했지만, 야넨슈는 이 종을 ''메갈로사우루스''로 분류했기에 단순한 복사 오류일 수 있다. 2011년 라우후트는 ''메갈로사우루스 잉겐스''가 카르카로돈토사우루스과를 나타낼 가능성이 있음을 보여주었다.
2000년과 2006년, 옥타비오 마테우스 연구팀은 포르투갈 로리냐 지층에서 발견된 화석(ML 352)을 ''케라토사우루스''의 새로운 표본으로 묘사했다. 2006년 마테우스와 동료들은 이 표본을 ''케라토사우루스 덴티술카투스'' 종으로 분류했지만, 2008년 카라노와 섐슨은 ''케라토사우루스''로는 분류했지만, 특정 종으로의 분류는 불가능하다고 결론지었다. 2015년, 엘리자베테 말라파이아와 동료들은 ''C. 덴티술카투스''의 유효성에 의문을 제기하며, ''케라토사우루스'' aff. ''케라토사우루스 나시코르니스''로 분류했다.
스위스 무티에에서 발견된 단일 이빨은 매드센과 웰스에 의해 ''케라토사우루스'' sp. (종 불확실)로 분류되었다. 2008년, 마티아스 소토와 다니엘 페레아는 우루과이의 타쿠아렘보 지층에서 이빨을 묘사했는데, 이는 ''케라토사우루스''에서만 알려진 특징을 보였다. 그러나 유해가 희소하여 ''케라토사우루스''로 분류하기는 어렵다고 강조했다. 2020년, 소토와 동료들은 동일 지층에서 추가적인 ''케라토사우루스'' 이빨을 묘사하여 이전의 해석을 뒷받침했다.[4]

마쉬는 ''Ceratosaurus nasicornis'' 홀로타입 최초 기술에서 다른 수각류에서는 알려지지 않은 특징을 언급하며, ''Ceratosaurus''를 새로운 과인 Ceratosauridae와 새로운 하목인 Ceratosauria의 유일한 구성원으로 만들었다. 1892년 에드워드 드링커 코프는 코뿔 같은 특징은 뚜렷한 종임을 보여줄 뿐, 뚜렷한 속을 정당화하기에는 불충분하다고 주장하며 ''C. nasicornis''를 ''Megalosaurus'' 속으로 할당했다.
''Ceratosaurus''는 이후 분석에서 뚜렷한 속으로 유지되었지만, 그 관계는 논란이 많았다. 여러 학자들이 ''Ceratosaurus''를 다양한 분류군에 포함시켰다. 1920년 길모어는 이 속이 트라이아스기 이후 가장 기저 수각류라고 주장하며 Ceratosauridae 과가 필요하다고 했다.
1985년, Abelisaurus와 ''Carnotaurus''가 ''Ceratosaurus''와 밀접한 관련이 있는 것으로 밝혀졌다. 1986년 고티에는 Coelophysoidea가 ''Ceratosaurus''와 가깝다고 보았지만, 최신 분석에서는 Ceratosauria 밖에 속한다. Ceratosauria는 조류로 이어지는 진화 계통에서 일찍 분기되어 수각류 내에서 기저로 간주된다. Ceratosauria는 Noasauridae와 Abelisauridae를 포함하는 Abelisauroidea와, ''Elaphrosaurus'', ''Deltadromeus'', ''Ceratosaurus'' 등 여러 기저 구성원을 포함한다.
기저 세라토사우루스 내 ''Ceratosaurus''의 위치는 논쟁 중이다. 일부는 ''Ceratosaurus''를 가장 파생된 기저 구성원으로 보고 Abelisauroidea의 자매 분류군으로 간주한다. 2004년 올리버 라우후트는 ''Genyodectes''를 ''Ceratosaurus''의 자매 분류군으로 제안하고, 이들을 Ceratosauridae 과로 묶었다.
2017년 쉬오 왕과 동료들은 Noasauridae가 Abelisauroidea에 속하지 않고 ''Ceratosaurus''보다 기저에 있다고 결론지었다. 2018년 라파엘 델코트는 Abelisauroidea를 Ceratosauroidea로 대체해야 한다고 지적하고, Abelisauridae는 ''Ceratosaurus''를 제외하는 새로운 정의를 제안했다. 왕과 동료들은 ''Ceratosaurus''와 ''Genyodectes''가 ''Eoabelisaurus''와 함께 분류군을 형성한다고 보았고, 델코트는 이 분류군을 Ceratosauridae로 명명하고, ''Ceratosaurus''보다 ''Carnotaurus''에 더 가까운 모든 분류군을 포함하도록 정의했다.

''Ceratosaurus''의 관계를 보여주는 다음 분기도는 2012년 디에고 폴과 올리버 라우후트의 계통 발생 분석을 기반으로 한다:
영국 쥐라기 중기 시대 두개골은 ''Ceratosaurus''와 유사한 코뿔을 가진 것으로 보인다. 1926년 프리드리히 폰 후에네는 이 두개골을 프로세라토사우루스로 기술하고 ''Ceratosaurus''의 선행자라고 가정했다. 오늘날 ''Proceratosaurus''는 티라노사우루스상과의 기저 구성원으로 간주된다. 코뿔은 두 속에서 독립적으로 진화했을 것이다. 2010년 올리버 라우후트와 동료들은 ''Proceratosaurus''를 Proceratosauridae로 분류하고, 코뿔이 불완전하게 보존되어 ''Guanlong'' 같은 다른 프로세라토사우루스과의 머리 볏 앞부분일 가능성을 지적했다.
3. 특징
케라토사우루스는 전형적인 대형 수각류 공룡의 신체 구조를 따랐다.[25] 이족보행을 했으며, 강력한 다리로 움직이고 팔은 크기가 줄어들었다. 완모식표본 USNM 4735는 여러 자료에 따르면 길이 5.3m 또는 5.69m의 개체였다.[26][27] 이 동물이 완전히 자랐는지는 불분명하다.[28] 오스니얼 찰스 마시는 1884년에 이 표본의 무게가 알로사우루스의 절반 정도일 것이라고 제안했다.[29] 이후 이 표본의 무게는 418kg, 524kg, 또는 670kg으로 수정되었다.[30] 20세기 후반에 발견된 세 개의 추가 골격은 훨씬 더 컸다.
''케라토사우루스 나시코르니스'' 모식 표본 척추에는 여러 틈이 있어 정확한 척추 수는 알 수 없다. 적어도 20개의 척추가 천골 앞쪽의 목과 등을 형성했다. 목의 중간 부분에서 척추 중심체(몸통)는 높이와 길이가 같았지만, 앞쪽과 뒤쪽 부분에서는 높이보다 짧았다. 위쪽으로 튀어나온 신경 가시는 비교적 컸고, 등쪽(등) 척추에서 척추 중심체만큼 높았다. 6개의 융합된 천골 척추로 구성된 천골은 위쪽으로 아치형을 이루며, 척추 중심체가 중간 부분에서 높이가 강하게 감소했는데, 이는 다른 일부 케라토사우루스류에서도 나타나는 현상이다. 꼬리는 약 50개의 꼬리 척추로 구성되었으며 동물 전체 길이의 약 절반이었다. 모식 표본에서는 약 2.84m로 추정되었다. 꼬리는 높은 신경 가시와 길쭉한 셰브론(척추 중심체 아래에 위치한 뼈) 때문에 위에서 아래까지 깊었다. 다른 공룡과 마찬가지로 몸의 균형을 잡고 있으며, 근육 수축 시 상단 허벅지를 뒤쪽으로 당겨 이동 중 앞으로 나아가는 추진력을 담당하는 caudofemoralis 근육을 포함하고 있었다.
어깨뼈는 부리돌기와 융합되어 두 개의 요소를 구분할 수 없는 단일 뼈를 형성했다. ''C. nasicornis'' 모식 표본은 불완전한 손을 포함하는 연결된 왼쪽 팔과 함께 발견되었다. 준비 과정에서 분리되었지만, 뼈의 원래 위치를 기록하기 위해 화석의 주형이 미리 만들어졌다. 손목뼈는 어떤 표본에서도 알려져 있지 않아 일부 학자들은 이 속에서 손실되었다고 추정했다. 2016년 매튜 카라노와 조나 초이니에르는 전완골과 중수골 사이의 틈새와 표면 질감으로 보아 하나 이상의 연골성(뼈가 아닌) 손목뼈가 존재했을 가능성이 있다고 제안했다. 대부분의 파생형질 수각류는 각 손에 세 개의 손가락(I–III)만 보이지만, ''케라토사우루스''는 네 개의 손가락을 유지했으며, IV 손가락은 크기가 감소했다. 첫 번째와 네 번째 중수골은 짧았고, 두 번째는 세 번째보다 약간 더 길었다. 중수골과 특히 첫 번째 발가락뼈는 다른 대부분의 기저 수각류와는 달리 비례적으로 매우 짧았다. 모식 표본에는 II, III, IV 손가락의 첫 번째 발가락뼈만 보존되어 있다. 발가락뼈와 발톱의 총수는 알려져 있지 않다. 중수골 I의 해부학적 구조는 이 손가락에도 원래 발가락뼈가 존재했음을 나타낸다. 발은 두 번째에서 네 번째까지 세 개의 체중 부하 손가락으로 구성되었다. 수각류에서 일반적으로 땅에 닿지 않는 퇴화된 발톱으로 축소되는 I 손가락은 모식 표본에서는 보존되지 않았다. 마시는 1884년 원본 설명에서 이 손가락이 ''케라토사우루스''에서 손실되었다고 가정했지만, 찰스 W. 길모어는 1920년 모노그래프에서 이 손가락의 존재를 보여주는 두 번째 중족골에 부착 영역이 있다고 언급했다.
수각류 중 특이하게도 ''케라토사우루스''는 몸 중앙선에 작고 길쭉하며 불규칙한 형태의 골편(피부뼈)을 가지고 있었다. 이러한 골편은 경추 4와 5, 꼬리 척추 4에서 10까지의 신경 가시 위에서 발견되었으며, 두개골 기저부에서 꼬리의 대부분까지 확장되었을 가능성이 있는 연속적인 열을 형성했을 것이다. 길모어가 1920년에 제안한 바와 같이, 바위 매트릭스에서의 위치는 살아있는 동물의 정확한 위치를 반영할 가능성이 높다. 꼬리 위 골편은 신경 가시와 25mm에서 38mm로 분리되어 있었는데, 아마도 그 사이에 존재하는 피부와 근육을 설명하는 것이며, 목의 골편은 신경 가시에 훨씬 더 가까웠다. 몸 중앙선 외에도 피부에는 추가적인 골편이 있었는데, 이는 모식 표본과 함께 발견된 58mm x 70mm 크기의 대략 사변형 판으로 나타난다. 그러나 이 판의 몸 위치는 알려져 있지 않다. 표본 UMNH VP 5278에서도 여러 개의 골편이 발견되었으며, 형태가 무정형으로 묘사되었다. 이러한 골편 대부분은 골격에서 최대 5m 떨어진 곳에서 발견되었지만, ''C. nasicornis'' 모식 표본과는 달리 어떤 척추와도 직접적으로 관련되지 않아 이 표본에서는 몸 위치를 추론할 수 없다.
몸길이 약 4.5~6미터의 중형 육식 공룡으로, 코 위에 있는 각질의 뿔 모양 돌기, 안와 상부에 있는 융기, 등면 정중선상에 늘어선 골질의 판 등이 눈에 띄는 특징이다.
3. 1. 두개골
두개골은 몸의 나머지 부분에 비해 상당히 컸다. ''C. nasicornis'' 모식 표본의 두개골 길이는 55cm이며, 주둥이 끝에서 첫 번째 경추에 연결되는 occipital condyle|후두과영어까지 측정되었다. 이 두개골의 너비는 심하게 왜곡되어 재구성하기 어려웠으며, 길모어(Gilmore)의 1920년 재구성은 나중에 너무 넓은 것으로 밝혀졌다. 비교적 완전한 표본 MWC 1의 두개골은 길이가 60cm이고 너비가 16cm인 것으로 추정되었다. 이 두개골은 모식 표본보다 약간 더 길었다. 두개골 뒤쪽은 다른 대형 수각류보다 두개골 구멍이 많아 가볍게 구성되었지만, 턱은 비율적으로 큰 치아를 지탱하기 위해 깊었다. 눈물뼈는 전안와창의 뒤쪽 가장자리뿐만 아니라 눈과 골성 콧구멍|골성 콧구멍영어 사이의 큰 구멍의 상단 가장자리 일부를 형성했는데, 이는 관련 종인 아벨리사우루스과의 구성원과는 달랐다. 턱 관절을 형성하기 위해 하악에 연결된 사각골은 턱 관절이 후두과와 관련하여 뒤쪽으로 이동하도록 기울어졌다. 이것은 또한 눈 뒤에 있는 큰 구멍인 측두하창의 바닥을 넓히는 결과를 가져왔다.
가장 뚜렷한 특징은 골성 콧구멍 뒤 두개골 정중선에 위치한 눈에 띄는 뿔이었으며, 이는 왼쪽 및 오른쪽 비골의 융합된 돌기에서 형성되었다. 화석에서는 뼈 뿔의 핵심만 알려져 있다. 살아있는 동물에서 이 핵심은 케라틴 외피를 지탱했을 것이다. 뿔 핵심의 기저부는 매끄러운 반면, 상부 2/3는 주름이 져 있었고 살아있을 때 혈관이 들어있을 홈이 있었다. 모식 표본에서 뿔 핵심은 길이가 13cm이고 기저부의 너비는 2cm이지만, 위로 올라갈수록 1.2cm로 좁아지고 높이는 7cm이다. MWC 1의 두개골에서는 더 길고 낮다. 살아있는 동물에서 뿔은 케라틴 외피로 인해 더 길어졌을 것이다. 비골 뿔 뒤에서 비골은 타원형 홈을 형성했다. 이 홈과 비골 뿔은 모두 ''케라토사우루스''(Ceratosaurus)를 관련 속과 구별하는 특징으로 작용한다. 큰 비골 뿔 외에도 ''케라토사우루스''는 각 눈 앞에 작은 반원형 뼈 능선을 가지고 있었으며, 이는 ''알로사우루스''(Allosaurus)의 것과 유사하다. 이 능선은 눈물뼈에 의해 형성되었다. 유체에서는 세 뿔 모두 성체보다 작았고 비골 뿔 핵심의 두 부분이 아직 융합되지 않았다.

주둥이 끝을 형성하는 전상악골은 각 측면에 단지 3개의 치아를 포함하고 있었으며, 이는 대부분의 다른 수각류보다 적었다. 위턱의 상악골|상악골영어은 모식 표본에서 각 측면에 15개의 칼날 모양의 치아로 줄지어 있었다. 이 치아 중 처음 8개는 매우 길고 튼튼했지만, 아홉 번째 치아부터는 점차 크기가 감소했다. 수각류의 전형적인 특징과 같이, 모식 표본에서 5mm당 약 10개의 치아 돌기가 포함된 미세하게 톱니 모양|톱니 모양영어의 가장자리가 특징이었다. 표본 MWC 1은 각 상악골에 11~12개의 치아만 보였고, 표본 UMNH VP 5278은 12개의 치아를 보였다. 후자의 표본에서는 치아가 더 튼튼하고 더 구부러졌다. 모든 표본에서 위턱의 치관은 예외적으로 길었다. 표본 UMNH VP 5278에서는 최대 9.3cm 길이였으며, 이는 하악의 최소 높이와 같다. 모식 표본에서는 길이가 7cm인데, 이는 하악의 최소 높이보다도 길다. 다른 수각류에서는, 비슷한 치아 길이는 아마도 밀접하게 관련된 ''게니오덱테스''(Genyodectes)에서만 알려져 있다. 반대로, 아벨리사우루스과 구성원 중 일부는 치관이 매우 짧은 특징을 가지고 있다. 모식 표본에서, 하악골|하악골영어의 치아를 지탱하는 뼈인 치골|치골영어의 각 부분에는 15개의 치아가 장착되었지만, 보존 상태가 좋지 않다. 표본 MWC 1과 UMNH VP 5278은 각 치골에 11개의 치아만 보이며, 후자의 표본에서 알 수 있듯이 위턱의 치아보다 약간 더 곧고 튼튼하지 않았다.
일반적으로 케라토사우루스의 특징으로 널리 알려진 것은 학명의 유래가 된 머리 꼭대기의 뿔(케라토)이다. 그러나 뿔이라는 명칭과는 달리, 그 두께는 얇고 오히려 볏에 가까운 물건이었다. 이러한 빈약한 "뿔"은 주로 라이벌이나 이성에게 과시하기 위해 사용되었을 가능성이 높다[17][18] 케라토사우루스의 두개골 화석을 보면, 뿔 부분에 주름과 같은 요철이 보이므로 뿔의 표면을 케라틴이 덮고 있었을 가능성도 있지만, 화석 증거는 발견되지 않았다.
이러한 빈약한 뿔보다 본 종의 특징을 두드러지게 하는 것은 체격에 어울리지 않게 발달한 세이버 모양의 위턱 이빨이다. 이 이빨은 자신보다 거대한 알로사우루스보다 길고[19], 카르카로돈토사우루스과와 매우 흡사하며, 먹이에 치명적인 열상을 입힐 수 있었다. 그 때문에 케라토사우루스는 중형(전장 6~7m)이면서, 약간 큰 용각류 등을 먹이로 삼았을 가능성도 있다[20][21][22] 이 이빨은 입을 닫으면 아래턱에서 튀어나올 정도로 길기 때문에, 최근 수각류 복원에서 주류가 되고 있는 "입술 복원"이 이 공룡에게 적절하지 않을 가능성을 시사하고 있다.
3. 2. 골격
케라토사우루스는 대형 수각류 공룡의 전형적인 신체 구조를 따랐다.[25] 이족보행을 하면서 강력한 다리로 움직였고 팔의 크기는 줄어들었다. 케라토사우루스의 골격이자 완모식표본인 USNM 4735 표본은 서로 다른 출처에 따르면 또는 의 개별 개체였다.[26][27] 이 동물이 완전히 자랐는지 여부는 불분명하다.[28] 1884년 오스니얼 찰스 마시는 이 표본의 무게가 현대 알로사우루스보다 약 절반 정도 무겁다고 제안했다.[29] 최근 기록에 따르면 이 표본의 무게는 , , 또는 으로 수정되었다.[30] 20세기 후반에 추가로 발견된 세 개의 골격은 훨씬 더 컸다.척추의 정확한 수는 ''케라토사우루스 나시코르니스''(Ceratosaurus nasicornis)의 모식 표본 척추에 여러 틈이 있어 알려져 있지 않다. 적어도 20개의 척추가 천골 앞쪽의 목과 등을 형성했다. 목의 중간 부분에서 척추의 중심체(몸통)는 높이와 길이가 같았지만, 목의 앞쪽과 뒤쪽 부분에서는 중심체가 높이보다 짧았다. 위쪽으로 튀어나온 신경 가시는 비교적 컸고, 등쪽(등) 척추에서 척추 중심체만큼 높았다. 6개의 융합된 천골 척추로 구성된 천골은 위쪽으로 아치형을 이루며, 척추 중심체가 중간 부분에서 높이가 강하게 감소했는데, 이는 다른 일부 케라토사우루스류에서도 나타나는 현상이다. 꼬리는 약 50개의 꼬리 척추로 구성되었으며 동물의 전체 길이의 약 절반이었다. 모식 표본에서는 로 추정되었다. 꼬리는 높은 신경 가시와 길쭉한 세브론(척추 중심체 아래에 위치한 뼈) 때문에 위에서 아래까지 깊었다. 다른 공룡과 마찬가지로 몸의 균형을 잡고 있으며, caudofemoralis 근육을 포함하고 있었는데, 이 근육은 이동 중 앞으로 나아가는 추진력을 담당했으며, 근육 수축 시 상단 허벅지를 뒤쪽으로 당겼다.
어깨뼈는 부리돌기와 융합되어 두 개의 원래 요소를 구분할 수 없는 단일 뼈를 형성했다. ''C. nasicornis'' 모식 표본은 불완전한 손을 포함하는 연결된 왼쪽 팔과 함께 발견되었다. 준비 과정에서 분리되었지만, 뼈의 원래 상대적인 위치를 기록하기 위해 화석의 주형이 미리 만들어졌다. 손목뼈는 어떤 표본에서도 알려져 있지 않아 일부 저자는 이 속에서 손실되었다고 추정했다. 2016년 논문에서 매튜 카라노와 조나 초이니에르는 전완골과 중수골 사이의 틈새와 이 틈새의 표면 질감에서 나타나는 것으로 미루어 보아 하나 이상의 연골성(뼈가 아닌) 손목뼈가 존재했을 가능성이 있다고 제안했다. 각 손에 세 개의 손가락(손가락 I–III)만 보이는 대부분의 파생형질 수각류와는 대조적으로, ''케라토사우루스''는 네 개의 손가락을 유지했는데, 손가락 IV가 크기가 감소했다. 첫 번째와 네 번째 중수골은 짧았고, 두 번째는 세 번째보다 약간 더 길었다. 중수골과 특히 첫 번째 발가락뼈는 다른 대부분의 기저 수각류와는 달리 비례적으로 매우 짧았다. 모식 표본에는 손가락 II, III, IV의 첫 번째 발가락뼈만 보존되어 있다. 발가락뼈와 발톱의 총수는 알려져 있지 않다. 중수골 I의 해부학적 구조는 이 손가락에도 원래 발가락뼈가 존재했음을 나타낸다. 발은 두 번째에서 네 번째까지 세 개의 체중 부하 손가락으로 구성되었다. 수각류에서 일반적으로 땅에 닿지 않는 퇴화된 발톱으로 축소되는 손가락 I은 모식 표본에서는 보존되지 않았다. 마시는 1884년 원본 설명에서 이 손가락이 ''케라토사우루스''에서 손실되었다고 가정했지만, 찰스 W. 길모어는 1920년 모노그래프에서 이 손가락의 존재를 보여주는 두 번째 중족골에 부착 영역이 있다고 언급했다.
수각류 중에서 특이하게도 ''케라토사우루스''는 몸의 중앙선에 작은, 길쭉하고 불규칙한 형태의 골편(피부뼈)을 가지고 있었다. 이러한 골편은 경추 4와 5, 꼬리 척추 4에서 10까지의 신경 가시 위에서 발견되었으며, 두개골 기저부에서 꼬리의 대부분까지 확장되었을 가능성이 있는 연속적인 열을 형성했을 것이다. 길모어가 1920년에 제안한 바와 같이, 바위 매트릭스에서의 위치는 살아있는 동물의 정확한 위치를 반영할 가능성이 높다. 꼬리 위의 골편은 신경 가시와 에서 로 분리되어 있었는데, 아마도 그 사이에 존재하는 피부와 근육을 설명하는 것이며, 목의 골편은 신경 가시에 훨씬 더 가까웠다. 몸의 중앙선 외에도, 피부에는 추가적인 골편이 포함되어 있었는데, 이는 모식 표본과 함께 발견된 by 크기의 대략 사변형 판으로 나타난다. 그러나 이 판의 몸에 대한 위치는 알려져 있지 않다. 표본 UMNH VP 5278에서도 여러 개의 골편이 발견되었으며, 형태가 무정형으로 묘사되었다. 이러한 골편의 대부분은 골격에서 최대 5m 떨어진 곳에서 발견되었지만, ''C. nasicornis'' 모식 표본과는 달리 어떤 척추와도 직접적으로 관련되지 않아 이 표본에서는 몸에 대한 원래 위치를 추론할 수 없다.
4. 고생물학
모리슨층과 로리나층에서 케라토사우루스 화석은 토르보사우루스[31], 알로사우루스와 같은 대형 수각류 화석과 함께 발견되는 경우가 많다. 여러 연구에서 이러한 공존 종들이 어떻게 직접적인 경쟁을 줄일 수 있었는지 설명하려고 시도했다.[32][33][34] 케라토사우루스는 희귀한 수각류 동물군으로, 이들이 공존하는 지역에서 알로사우루스에 비해 평균 7.5 대 1의 비율로 수적으로 뒤처져 있었다.[35]
1998년 도널드 헨더슨은 케라토사우루스가 짧은 주둥이, 높고 넓은 두개골, 짧고 뒤로 뻗은 이빨을 가진 형태와 긴 주둥이, 낮은 두개골, 길고 수직인 이빨을 가진 형태의 두 가지 알로사우루스와 공존했다고 주장했다. 헨더슨은 짧은 주둥이 알로사우루스 형태가 긴 주둥이 형태 및 케라토사우루스와 다른 생태적 틈새를 차지한다고 결론 내렸다. 짧은 두개골은 물 때 발생하는 굴곡 모멘트를 줄여 물기 힘을 증가시켰다. 반면 케라토사우루스와 다른 알로사우루스 형태는 긴 주둥이 두개골을 가졌고, 더 긴 이빨은 날카로운 물기를 가하기 위해 송곳니처럼 사용되었을 것이며, 좁은 두개골 때문에 물기 힘은 작은 영역에 집중되었을 것이다. 헨더슨은 케라토사우루스와 긴 주둥이 알로사우루스 형태 사이의 두개골 모양의 유사성은 이들이 서로 직접적인 경쟁을 벌였음을 나타낸다고 보았다. 따라서 케라토사우루스는 긴 주둥이 형태가 지배하는 서식지에서 밀려났을 수 있다.[32]
케라토사우루스는 몸길이 약 4.5~6미터의 중형 육식 공룡으로, 코 위에 있는 각질의 뿔 모양 돌기, 안와 상부의 융기, 등면 정중선상에 늘어선 골질의 판 등이 특징이다. 뿔 모양 돌기를 가진 수각류는 몇몇 알려져 있지만, 등면에 골판을 가진 수각류는 케라토사우루스 뿐이다. 케라토사우루스의 뿔은 얇고 볏에 가까워, 주로 라이벌이나 이성에게 과시하기 위해 사용되었을 가능성이 높다.[17][18]
케라토사우루스는 체격에 비해 발달한 세이버 모양의 위턱 이빨을 가졌다. 이 이빨은 알로사우루스보다 길고[19], 카르카로돈토사우루스과와 매우 흡사하며, 먹이에 치명적인 열상을 입힐 수 있었다. 중형(전장 6~7m)이면서, 약간 큰 용각류 등을 먹이로 삼았을 가능성도 있다.[20][21][22] 이빨은 입을 닫으면 아래턱에서 튀어나올 정도로 길었다. 앞다리에는 4개의 손가락이 있지만, 발톱이 자라난 것은 세 번째 손가락까지였고, 네 번째 손가락은 흔적 정도였다. 꼬리는 유연하고 폭이 넓고 두꺼웠다.
''케라토사우루스''는 대형 수각류 공룡의 전형적인 몸 구조를 따랐다. 두 발 보행을 하는 종으로, 강력한 다리로 이동했으며 팔은 크기가 감소했다. 최초로 발견된 골격이자 ''케라토사우루스 나시코르니스''의 모식 표본인 USNM 4735 표본은 길이가 5.3m 또는 5.69m인 개체였다. 이 동물이 완전히 성장했는지는 불분명하다.
두개골은 몸의 나머지 부분에 비해 상당히 컸다. 가장 뚜렷한 특징은 골성 콧구멍 뒤 두개골 정중선에 위치한 눈에 띄는 뿔이었으며, 이는 왼쪽 및 오른쪽 비골의 융합된 돌기에서 형성되었다. 큰 비골 뿔 외에도 ''케라토사우루스''는 각 눈 앞에 작은 반원형 뼈 능선을 가지고 있었으며, 이는 ''알로사우루스''(Allosaurus)와 유사하다. 이 능선은 눈물뼈에 의해 형성되었다.
주둥이 끝을 형성하는 전상악골은 각 측면에 단지 3개의 치아를 포함하고 있었으며, 이는 대부분의 다른 수각류보다 적었다. 위턱의 상악골은 모식 표본에서 각 측면에 15개의 칼날 모양의 치아로 줄지어 있었다.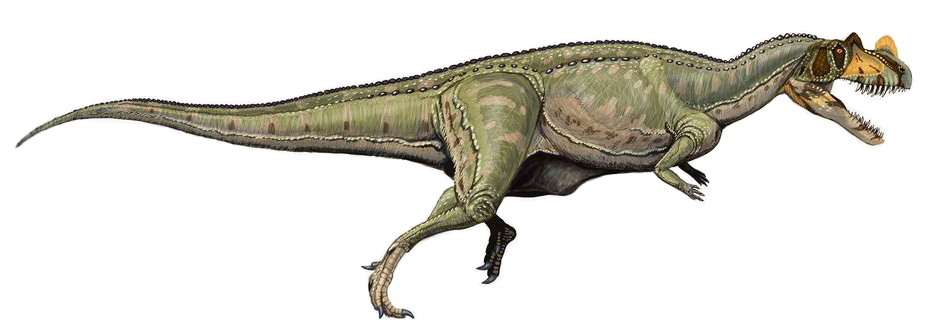
척추의 정확한 수는 ''케라토사우루스 나시코르니스'' 모식 표본 척추에 여러 틈이 있어 알려져 있지 않다. 꼬리는 약 50개의 미추로 구성되었으며 동물의 전체 길이의 약 절반이었다.
어깨뼈는 부리돌기와 융합되어 두 개의 원래 요소를 구분할 수 없는 단일 뼈를 형성했다. 수각류 중에서 특이하게도 ''케라토사우루스''는 몸의 중앙선에 작은, 길쭉하고 불규칙한 형태의 골편(피부뼈)을 가지고 있었다.




4. 1. 생태 및 먹이
모리슨층과 로리나층에서 케라토사우루스 화석은 토르보사우루스[31], 알로사우루스와 같은 대형 수각류 화석과 함께 발견되는 경우가 많다. 여러 연구에서 이러한 공존 종들이 어떻게 직접적인 경쟁을 줄일 수 있었는지 설명하려고 시도했다.[32][33][34] 케라토사우루스는 희귀한 수각류 동물군으로, 이들이 공존하는 지역에서 알로사우루스에 비해 평균 7.5 대 1의 비율로 수적으로 뒤처져 있었다.[35]1998년 도널드 헨더슨은 케라토사우루스가 두 가지 형태의 알로사우루스와 공존했다고 주장했다. 짧은 주둥이, 높고 넓은 두개골, 짧고 뒤로 뻗은 이빨을 가진 형태와 긴 주둥이, 낮은 두개골, 길고 수직인 이빨을 가진 형태이다. 헨더슨은 짧은 주둥이 알로사우루스 형태가 긴 주둥이 형태 및 케라토사우루스와 다른 생태적 틈새를 차지한다고 결론 내렸다. 짧은 두개골은 물 때 발생하는 굴곡 모멘트를 줄여 물기 힘을 증가시켰다. 반면 케라토사우루스와 다른 알로사우루스 형태는 긴 주둥이 두개골을 가졌고, 더 긴 이빨은 날카로운 물기를 가하기 위해 송곳니처럼 사용되었을 것이며, 좁은 두개골 때문에 물기 힘은 작은 영역에 집중되었을 것이다. 헨더슨은 케라토사우루스와 긴 주둥이 알로사우루스 형태 사이의 두개골 모양의 유사성은 이들이 서로 직접적인 경쟁을 벌였음을 나타낸다고 보았다. 따라서 케라토사우루스는 긴 주둥이 형태가 지배하는 서식지에서 밀려났을 수 있다.[32]
헨더슨은 케라토사우루스가 다른 먹이를 선호함으로써 경쟁을 피했을 수 있다고 제안했다. 또한 케라토사우루스의 길어진 이빨은 같은 종을 인식하거나 다른 사회적 기능을 수행하는 데 도움이 되는 시각적 신호 역할을 했을 수 있다. 이러한 수각류의 큰 크기는 경쟁을 감소시키는 경향이 있었을 것이다.[32]
2006년 연구에서 포스터와 다니엘 추어는 케라토사우루스와 알로사우루스가 일반적으로 동일한 서식지를 공유하고 동일한 유형의 먹이를 사냥했으며, 다른 먹이 전략을 가지고 있었을 것이라는 헨더슨의 주장에 동의했다. 그러나 두 알로사우루스 형태 사이의 구분은 일부 후기 연구에서 의문을 제기했다. 2010년 연구에서 케네스 카펜터는 클리블랜드 로이드 채석장의 짧은 주둥이 알로사우루스 개체는 별개의 분류군이 아닌 극심한 개체 변이의 경우를 나타낸다고 결론지었다.
2004년 연구에서 로버트 배커와 게리 비어는 케라토사우루스가 주로 폐어, 악어, 거북과 같은 수생 먹이에 특화되었다고 제안했다. 코모 블러프 주변 50개 지역에서 수집된 이빨 분석에 따르면, 케라토사우루스와 메갈로사우루스과의 이빨은 젖은 범람원, 호수 가장자리, 늪과 같은 수역 주변과 서식지에서 가장 흔하게 발견되었다. 연구자들은 케라토사우루스와 메갈로사우루스과가 주로 수역 근처에서 사냥을 했을 것이며, 케라토사우루스는 가끔 더 큰 공룡의 시체를 먹었을 것이라고 결론지었다. 또한 케라토사우루스의 길고, 낮고, 유연한 몸과 꼬리 앞쪽 척추뼈의 신경 가시가 더 높고 수직인 점, 꼬리 아래쪽의 깊은 쉐브론 뼈는 수영에 적응했을 가능성이 있는 깊고 "악어 같은" 꼬리를 나타낸다고 보았다. 반대로, 알로사우루스과는 다리가 더 길고, 짧고, 더 높고, 더 뻣뻣한 몸체를 가지고 있어, 열린 지형에서 빠르게 달리는 데 적응했을 것이라고 추측했다. 그러나 2019년 윤은 케라토사우루스가 완전한 반수생이라기보다는 모리슨 층의 다른 수각류보다 수생 먹이를 사냥하는 데 더 능숙했을 뿐이라고 제안했다.[6]
알로사우루스의 치골 발은 케라토사우루스 또는 토르보사우루스의 이빨에 의해 생긴 흔적을 보여준다. 이는 알로사우루스가 시체를 먹고 있었음을 나타낸다.[7] 상부 쥐라기 미가트-무어 채석장의 뼈 조립체는 비정상적으로 높은 수각류 물린 자국 발생률을 보존하는데, 그 중 대부분은 알로사우루스와 케라토사우루스에 기인할 수 있다.[8]
케라토사우루스는 몸길이 약 4.5~6미터의 중형 육식 공룡으로, 코 위에 있는 각질의 뿔 모양 돌기, 안와 상부의 융기, 등면 정중선상에 늘어선 골질의 판 등이 특징이다. 뿔 모양 돌기를 가진 수각류는 몇몇 알려져 있지만, 등면에 골판을 가진 수각류는 케라토사우루스 뿐이다.
케라토사우루스의 뿔은 얇고 볏에 가까워, 주로 라이벌이나 이성에게 과시하기 위해 사용되었을 가능성이 높다.[17][18] 케라토사우루스의 두개골 화석에는 뿔 부분에 주름과 같은 요철이 보여 각질(케라틴)이 뿔 표면을 덮고 있었을 가능성도 있지만, 화석 증거는 발견되지 않았다.
케라토사우루스는 체격에 비해 발달한 세이버 모양의 위턱 이빨을 가졌다. 이 이빨은 알로사우루스보다 길고[19], 카르카로돈토사우루스과와 매우 흡사하며, 먹이에 치명적인 열상을 입힐 수 있었다. 케라토사우루스는 중형(전장 6~7m)이면서, 약간 큰 용각류 등을 먹이로 삼았을 가능성도 있다.[20][21][22] 이 이빨은 입을 닫으면 아래턱에서 튀어나올 정도로 길다.
앞다리에는 4개의 손가락이 있지만, 발톱이 자라난 것은 세 번째 손가락까지였고, 네 번째 손가락은 흔적 정도였다. 꼬리는 유연한 구조로 폭이 넓고 두꺼웠다. 로버트 배커는 케라토사우루스가 반수생이었을 것이라고 지적했지만, 이에 대한 이견도 많다.[24]
4. 2. 기능
''케라토사우루스''는 대형 수각류 공룡의 전형적인 몸 구조를 따랐다. 두 발 보행을 하는 종으로, 강력한 다리로 이동했으며 팔은 크기가 감소했다. 최초로 발견된 골격이자 ''케라토사우루스 나시코르니스''의 모식 표본인 USNM 4735 표본은 길이가 5.3m 또는 5.69m인 개체였다. 이 동물이 완전히 성장했는지는 불분명하다. 오트니엘 찰스 마시는 1884년에 이 표본의 무게가 동시대의 ''알로사우루스''의 절반 정도일 것이라고 추정했다. 최근에는 이 무게가 418kg, 524kg, 또는 670kg으로 수정되었다.
20세기 후반에 발견된 세 개의 추가 골격은 실질적으로 더 컸다. UMNH VP 5278 표본은 제임스 헨리 매드슨에 의해 길이가 약 8.8m로 추정되었지만, 나중에는 7m로 추정되었다. 무게는 서로 다른 연구에서 980kg, 452kg, 700kg으로 계산되었다. 두 번째 골격 MWC 1은 UMNH VP 5278보다 약간 작았으며 무게는 275kg일 수 있다. 세 번째, 아직 기술되지 않은 BYUVP 12893 표본은 가장 큰 것으로 발견되었다고 주장했지만, 추정치는 발표되지 않았다. 2000년 포르투갈에서 발견된 ML 352 표본은 길이가 6m이고 무게는 600kg으로 추정되었다.
두개골은 몸의 나머지 부분에 비해 상당히 컸다. ''C. nasicornis'' 모식 표본의 두개골 길이는 55cm이며, 주둥이 끝에서 첫 번째 경추에 연결되는 후두과까지 측정되었다. 이 두개골의 너비는 심하게 왜곡되어 재구성하기 어려웠으며, 길모어(Gilmore)의 1920년 재구성은 나중에 너무 넓은 것으로 밝혀졌다. 비교적 완전한 MWC 1 표본의 두개골은 길이가 60cm이고 너비가 16cm인 것으로 추정되었다. 이 두개골은 모식 표본보다 약간 더 길었다. 두개골 뒤쪽은 다른 대형 수각류보다 두개골 구멍이 많아 가볍게 구성되었지만, 턱은 비율적으로 큰 치아를 지탱하기 위해 깊었다. 눈물뼈는 전안와창의 뒤쪽 가장자리뿐만 아니라 눈과 골성 콧구멍 사이의 큰 구멍의 상단 가장자리 일부를 형성했는데, 이는 관련 종인 아벨리사우루스과의 구성원과는 달랐다. 턱 관절을 형성하기 위해 하악에 연결된 사각골은 턱 관절이 후두과와 관련하여 뒤쪽으로 이동하도록 기울어졌다. 이것은 또한 눈 뒤에 있는 큰 구멍인 측두하창의 바닥을 넓히는 결과를 가져왔다.
가장 뚜렷한 특징은 골성 콧구멍 뒤 두개골 정중선에 위치한 눈에 띄는 뿔이었으며, 이는 왼쪽 및 오른쪽 비골의 융합된 돌기에서 형성되었다. 화석에서는 뼈 뿔의 핵심만 알려져 있다. 살아있는 동물에서 이 핵심은 케라틴 외피를 지탱했을 것이다. 뿔 핵심의 기저부는 매끄러운 반면, 상부 2/3는 주름이 져 있었고 살아있을 때 혈관이 들어있을 홈이 있었다. 모식 표본에서 뿔 핵심은 길이가 13cm이고 기저부의 너비는 2cm이지만, 위로 올라갈수록 1.2cm로 좁아지고 높이는 7cm이다. MWC 1의 두개골에서는 더 길고 낮다. 살아있는 동물에서 뿔은 케라틴 외피로 인해 더 길어졌을 것이다. 비골 뿔 뒤에서 비골은 타원형 홈을 형성했다. 이 홈과 비골 뿔은 모두 ''케라토사우루스''(Ceratosaurus)를 관련 속과 구별하는 특징으로 작용한다. 큰 비골 뿔 외에도 ''케라토사우루스''는 각 눈 앞에 작은 반원형 뼈 능선을 가지고 있었으며, 이는 ''알로사우루스''(Allosaurus)의 것과 유사하다. 이 능선은 눈물뼈에 의해 형성되었다. 유체에서는 세 뿔 모두 성체보다 작았고 비골 뿔 핵심의 두 부분이 아직 융합되지 않았다.
주둥이 끝을 형성하는 전상악골은 각 측면에 단지 3개의 치아를 포함하고 있었으며, 이는 대부분의 다른 수각류보다 적었다. 위턱의 상악골은 모식 표본에서 각 측면에 15개의 칼날 모양의 치아로 줄지어 있었다. 이 치아 중 처음 8개는 매우 길고 튼튼했지만, 아홉 번째 치아부터는 점차 크기가 감소했다. 수각류의 전형적인 특징과 같이, 모식 표본에서 5mm당 약 10개의 치아 돌기가 포함된 미세하게 톱니 모양의 가장자리가 특징이었다. MWC 1 표본은 각 상악골에 11~12개의 치아만 보였고, UMNH VP 5278 표본은 12개의 치아를 보였다. 후자의 표본에서는 치아가 더 튼튼하고 더 구부러졌다. 모든 표본에서 위턱의 치관은 예외적으로 길었다. UMNH VP 5278 표본에서는 최대 9.3cm 길이였으며, 이는 하악의 최소 높이와 같다. 모식 표본에서는 길이가 7cm인데, 이는 하악의 최소 높이보다도 길다. 다른 수각류에서는, 비슷한 치아 길이는 아마도 밀접하게 관련된 ''게니오덱테스''(Genyodectes)에서만 알려져 있다. 반대로, 아벨리사우루스과 구성원 중 일부는 치관이 매우 짧은 특징을 가지고 있다. 모식 표본에서, 하악골의 치아를 지탱하는 뼈인 치골의 각 부분에는 15개의 치아가 장착되었지만, 보존 상태가 좋지 않다. MWC 1과 UMNH VP 5278 표본은 각 치골에 11개의 치아만 보이며, 후자의 표본에서 알 수 있듯이 위턱의 치아보다 약간 더 곧고 튼튼하지 않았다.
척추의 정확한 수는 ''케라토사우루스 나시코르니스''(Ceratosaurus nasicornis)의 모식 표본 척추에 여러 틈이 있어 알려져 있지 않다. 적어도 20개의 척추가 천골 앞쪽의 목과 등을 형성했다. 목의 중간 부분에서 척추의 중심체(몸통)는 높이와 길이가 같았지만, 목의 앞쪽과 뒤쪽 부분에서는 중심체가 높이보다 짧았다. 위쪽으로 튀어나온 신경 가시는 비교적 컸고, 등쪽(등) 척추에서 척추 중심체만큼 높았다. 6개의 융합된 천골 척추로 구성된 천골은 위쪽으로 아치형을 이루며, 척추 중심체가 중간 부분에서 높이가 강하게 감소했는데, 이는 다른 일부 케라토사우루스류에서도 나타나는 현상이다. 꼬리는 약 50개의 미추로 구성되었으며 동물의 전체 길이의 약 절반이었다. 모식 표본에서는 약 2.84m로 추정되었다. 꼬리는 높은 신경 가시와 길쭉한 쉐브론, 즉 척추 중심체 아래에 위치한 뼈 때문에 위에서 아래까지 깊었다. 다른 공룡과 마찬가지로 몸의 균형을 잡고 있으며, caudofemoralis 근육을 포함하고 있었는데, 이 근육은 이동 중 앞으로 나아가는 추진력을 담당했으며, 근육 수축 시 상단 허벅지를 뒤쪽으로 당겼다.
어깨뼈는 부리돌기와 융합되어 두 개의 원래 요소를 구분할 수 없는 단일 뼈를 형성했다. ''C. nasicornis'' 모식 표본은 불완전한 손을 포함하는 연결된 왼쪽 팔과 함께 발견되었다. 준비 과정에서 분리되었지만, 뼈의 원래 상대적인 위치를 기록하기 위해 화석의 주형이 미리 만들어졌다. 손목뼈는 어떤 표본에서도 알려져 있지 않아 일부 저자는 이 속에서 손실되었다고 추정했다. 2016년 논문에서 매튜 카라노와 조나 초이니에르는 전완골과 중수골 사이의 틈새와 이 틈새의 표면 질감에서 나타나는 것으로 미루어 보아 하나 이상의 연골성(뼈가 아닌) 손목뼈가 존재했을 가능성이 있다고 제안했다. 각 손에 세 개의 손가락(손가락 I–III)만 보이는 대부분의 파생형질 수각류와는 대조적으로, ''케라토사우루스''는 네 개의 손가락을 유지했는데, 손가락 IV가 크기가 감소했다. 첫 번째와 네 번째 중수골은 짧았고, 두 번째는 세 번째보다 약간 더 길었다. 중수골과 특히 첫 번째 지골은 다른 대부분의 기저 수각류와는 달리 비례적으로 매우 짧았다. 모식 표본에는 손가락 II, III, IV의 첫 번째 지골만 보존되어 있다. 지골과 발톱의 총수는 알려져 있지 않다. 중수골 I의 해부학적 구조는 이 손가락에도 원래 지골이 존재했음을 나타낸다. 발은 두 번째에서 네 번째까지 세 개의 체중 부하 손가락으로 구성되었다. 수각류에서 일반적으로 땅에 닿지 않는 퇴화된 발톱으로 축소되는 손가락 I은 모식 표본에서는 보존되지 않았다. 마시는 1884년 원본 설명에서 이 손가락이 ''케라토사우루스''에서 손실되었다고 가정했지만, 찰스 W. 길모어는 1920년 모노그래프에서 이 손가락의 존재를 보여주는 두 번째 중족골에 부착 영역이 있다고 언급했다.
수각류 중에서 특이하게도 ''케라토사우루스''는 몸의 중앙선에 작은, 길쭉하고 불규칙한 형태의 골편(피부뼈)을 가지고 있었다. 이러한 골편은 경추 4와 5, 미추 4에서 10까지의 신경 가시 위에서 발견되었으며, 두개골 기저부에서 꼬리의 대부분까지 확장되었을 가능성이 있는 연속적인 열을 형성했을 것이다. 길모어가 1920년에 제안한 바와 같이, 바위 매트릭스에서의 위치는 살아있는 동물의 정확한 위치를 반영할 가능성이 높다. 꼬리 위의 골편은 신경 가시와 25mm에서 38mm로 분리되어 있었는데, 아마도 그 사이에 존재하는 피부와 근육을 설명하는 것이며, 목의 골편은 신경 가시에 훨씬 더 가까웠다. 몸의 중앙선 외에도, 피부에는 추가적인 골편이 포함되어 있었는데, 이는 모식 표본과 함께 발견된 58mm x 70mm 크기의 대략 사변형 판으로 나타난다. 그러나 이 판의 몸에 대한 위치는 알려져 있지 않다. UMNH VP 5278 표본에서도 여러 개의 골편이 발견되었으며, 형태가 무정형으로 묘사되었다. 이러한 골편의 대부분은 골격에서 최대 5m 떨어진 곳에서 발견되었지만, ''C. nasicornis'' 모식 표본과는 달리 어떤 척추와도 직접적으로 관련되지 않아 이 표본에서는 몸에 대한 원래 위치를 추론할 수 없다.
모리슨 층과 로리냐 층에서, ''세라토사우루스'' 화석은 거대한 수각류, 이를테면 메갈로사우루스과의 ''토르보사우루스''[5] 와 알로사우루스과인 ''알로사우루스''와 함께 발견되는 경우가 많다. 콜로라도 주의 가든 파크 지역에서는 ''세라토사우루스'' 외에도 ''알로사우루스''로 여겨지는 화석이 발견되었다. 콜로라도 주의 드라이 메사 채석장, 유타 주의 클리블랜드 로이드 채석장과 공룡 국립 기념지에서는 각각 ''세라토사우루스'', ''알로사우루스'', 그리고 ''토르보사우루스''의 잔해가 발견되었다.[14] 마찬가지로, 와이오밍 주의 코모 블러프와 인근 지역에서도 ''세라토사우루스'', ''알로사우루스'', 그리고 적어도 하나의 거대한 메갈로사우루스과의 잔해가 발견되었다. ''세라토사우루스''는 수각류 동물군에서 드문 존재였는데, 공존하는 지역에서 ''알로사우루스''보다 평균 7.5 대 1의 비율로 수가 적었다.
여러 연구에서는 이러한 공존 종들이 어떻게 직접적인 경쟁을 줄일 수 있었는지 설명하려고 시도했다. 1998년, 도널드 헨더슨은 ''세라토사우루스''가 두 개의 별개의 ''알로사우루스'' 종, 즉 "형태"와 공존했다고 주장했는데, 그는 짧은 주둥이, 높고 넓은 두개골, 짧고 뒤로 뻗은 이빨을 가진 형태와 더 긴 주둥이, 낮은 두개골, 길고 수직인 이빨을 가진 형태로 구분했다. 일반적으로, 형태, 생리학 및 행동과 관련하여 공존 종 사이의 유사성이 클수록, 이들 종 간의 경쟁은 더 치열해질 것이다. 헨더슨은 짧은 주둥이 ''알로사우루스'' 형태가 긴 주둥이 형태와 ''세라토사우루스'' 모두와 다른 생태적 지위를 차지했다는 결론에 도달했다. 이 형태의 짧은 두개골은 물 때 발생하는 굴곡 모멘트를 줄여 물기 힘을 증가시켰으며, 이는 고양이에서 보이는 상황과 유사했다. 그러나 ''세라토사우루스''와 다른 ''알로사우루스'' 형태는 긴 주둥이 두개골을 가지고 있었는데, 이는 개의 것과 더 유사하다. 더 긴 이빨은 날카롭고 찢는 물기를 가하기 위해 송곳니처럼 사용되었을 것이며, 좁은 두개골 때문에 물기 힘은 작은 영역에 집중되었다. 헨더슨에 따르면, ''세라토사우루스''와 긴 주둥이 ''알로사우루스'' 형태 사이의 두개골 모양의 유사성은 이들 형태가 서로 직접적인 경쟁을 벌였음을 나타낸다. 따라서 ''세라토사우루스''는 긴 주둥이 형태가 지배하는 서식지에서 밀려났을 수 있다. 실제로 ''세라토사우루스''는 긴 주둥이 ''알로사우루스'' 형태를 포함하는 클리블랜드 로이드 채석장에서는 매우 드물지만, 짧은 주둥이 형태와 공존하는 가든 파크와 드라이 메사 채석장에서는 더 흔하게 나타난다.
더욱이 헨더슨은 ''세라토사우루스''가 다른 먹이를 선호함으로써 경쟁을 피했을 수 있다고 제안했다. 그것의 극도로 길어진 이빨의 진화는 긴 주둥이 ''알로사우루스'' 형태와의 경쟁의 직접적인 결과였을 수 있다. 두 종 모두 청소부 역할을 할 때 시체의 다른 부분을 선호했을 수도 있다. ''세라토사우루스''의 길어진 이빨은 같은 종의 구성원을 인식하거나 다른 사회적 기능을 수행하는 데 도움이 되는 시각적 신호 역할을 했을 수 있다. 또한, 이러한 수각류의 큰 크기는 가능한 먹이의 수가 크기에 따라 증가하기 때문에 경쟁을 감소시키는 경향이 있었을 것이다.
2006년의 연구에서 포스터와 다니엘 추어는 ''세라토사우루스''와 ''알로사우루스''가 일반적으로 동일한 서식지를 공유하고 동일한 유형의 먹이를 사냥했으며, 이는 경쟁을 피하기 위해 다른 먹이 전략을 가지고 있었을 것이라는 헨더슨의 주장에 동의했다. 이 연구자들에 따르면, 이는 두개골, 이빨 및 팔의 다른 비율에 의해서도 입증된다. 그러나 두 개의 ''알로사우루스'' 형태 사이의 구분은 일부 후기 연구에서 의문을 제기했다. 2010년의 연구에서 케네스 카펜터는 클리블랜드 로이드 채석장의 짧은 주둥이 ''알로사우루스'' 개체는 별개의 분류군이 아닌 극심한 개체 변이의 경우를 나타낸다고 결론지었다. 더욱이, 헨더슨이 짧은 주둥이 형태에 대한 분석의 근거가 된 가든 파크 지역의 USNM 4734 두개골은 나중에 너무 짧게 재구성되었음이 밝혀졌다.
2004년의 연구에서 로버트 T. 배커와 게리 비어는 ''세라토사우루스''가 주로 폐어, 악어, 그리고 거북이와 같은 수생 먹이에 특화되었다고 제안했다. 코모 블러프 주변 50개의 개별 지역에서 수집된 이빨에 대한 통계적 분석에 따르면, ''세라토사우루스''와 메갈로사우루스과의 이빨은 젖은 범람원, 호수 가장자리, 늪과 같은 수역 주변과 서식지에서 가장 흔하게 발견되었다. ''세라토사우루스''는 또한 가끔 육상 지역에서도 발견되었다. 그러나 알로사우루스과는 육상 및 수생 서식지에서 동일하게 흔하게 발견되었다. 이러한 결과로부터 배커와 비어는 ''세라토사우루스''와 메갈로사우루스과가 주로 수역 근처와 내에서 사냥을 했을 것이며, ''세라토사우루스''는 가끔 더 큰 공룡의 시체를 먹었을 것이라고 결론지었다. 연구자들은 또한 ''세라토사우루스''와 메갈로사우루스과의 길고, 낮고, 유연한 몸을 언급했다. 다른 모리슨 수각류에 비해 ''세라토사우루스''는 꼬리 앞쪽 척추뼈의 신경 가시가 더 높았고, 뒤쪽을 향하지 않고 수직이었다. 꼬리 아래쪽의 깊은 쉐브론 뼈와 함께, 그들은 수영에 적응했을 가능성이 있는 깊고 "악어 같은" 꼬리를 나타낸다. 반대로, 알로사우루스과는 다리가 더 길고, 짧고, 더 높고, 더 뻣뻣한 몸체를 가지고 있다. 그들은 열린 지형에서 빠르게 달리고 초식 동물인 용각류와 스테고사우루스와 같은 대형 초식 공룡을 사냥하는 데 적응했을 것이지만, 배커와 비어가 추측한 바와 같이, 계절에 따라 대형 초식 동물이 없을 때는 수생 먹이로 전환했을 것이다. 그러나 이 이론은 2019년 윤에 의해 반박되었는데, 그는 ''세라토사우루스''가 완전한 반수생이라기보다는 모리슨 층의 다른 수각류보다 수생 먹이를 사냥하는 데 더 능숙했을 뿐이라고 제안했다.[6]
1986년의 대중적인 저서 ''공룡 이설''에서 배커는 위턱뼈가 주변 두개골 뼈에 느슨하게 부착되어 두개골 내에서 어느 정도의 움직임을 허용하며, 이를 두개 운동성이라고 칭했다. 마찬가지로, 아래턱 뼈는 서로와 사각형 뼈에 대해 움직일 수 있었고, 사각형 뼈는 바깥쪽으로 흔들려 턱 관절에서 아래턱을 벌릴 수 있었다. 이러한 특징들을 모두 합하면, 이 동물은 더 큰 먹이를 삼키기 위해 턱을 넓힐 수 있었을 것이다. 2008년의 연구에서 케이시 홀리데이와 로렌스 위트머는 다른 공룡에 대해 유사한 주장을 재평가하여, 근육으로 구동되는 두개 운동성의 존재는 어떤 공룡 종에서도 증명될 수 없으며 대부분의 경우에 부재했을 것이라고 결론지었다.
''알로사우루스''의 치골 발은 아마도 ''세라토사우루스'' 또는 ''토르보사우루스''의 이빨에 의해 생긴 흔적을 보여준다. 몸의 뼈 위치(몸통의 하단 가장자리를 따라 있고 다리에 의해 부분적으로 가려짐)와 골격에서 가장 거대한 뼈 중 하나였다는 사실은 ''알로사우루스''가 시체를 먹고 있었음을 나타낸다.[7] 상부 쥐라기 미가트-무어 채석장의 뼈 조립체는 비정상적으로 높은 수각류 물린 자국 발생률을 보존하는데, 그 중 대부분은 ''알로사우루스''와 ''세라토사우루스''에 기인할 수 있으며, 다른 자국은 줄무늬의 크기를 고려할 때 ''사우로파가낙스'' 또는 ''토르보사우루스''에 의해 만들어졌을 수 있다. 초식 공룡에 있는 물린 자국의 위치는 포식 또는 잔해에 대한 조기 접근과 일치하는 반면, ''알로사우루스'' 재료에서 발견된 물린 자국은 다른 수각류 또는 다른 ''알로사우루스''로부터의 시체 청소를 시사한다. 다른 조립체에 비해 비정상적으로 높은 수각류 물린 자국의 농도는 수각류에 의한 건기 동안 자원의 더 완전한 활용 또는 다른 지역의 수집 편견으로 설명될 수 있다.[8]
1884년, 마시는 ''케라토사우루스''의 코뿔이 공격과 방어 모두에 사용되는 "매우 강력한 무기"라고 생각했으며, 1920년 길모어도 이 해석에 동의했다. 현재는 뿔을 무기로 사용했을 가능성은 일반적으로 낮게 여겨진다. 1985년 데이비드 노먼은 뿔이 "다른 포식자로부터의 방어용은 아닐 것"이며, 대신 번식 권리를 놓고 다투는 수컷 케라토사우루스 간의 동족 간의 싸움에 사용되었을 가능성이 있다고 생각했다. 그레고리 S. 폴은 1988년 비슷한 기능을 제안하며 치명적이지 않지만 머리 부딪히기 싸움을 하는 두 마리의 ''케라토사우루스''를 묘사했다. 1990년, 로우와 고티에는 ''케라토사우루스''의 코뿔이 "아마도 전시 목적으로만 사용되었을 것이며" 물리적인 충돌에는 아무런 역할도 하지 않았을 것이라고 주장했다. 전시용으로 사용되었다면, 뿔은 밝은 색이었을 것이다. 몸 중앙선을 따라 내려오는 골판의 열 역시 전시 기능이 제안되었다.
''케라토사우루스''의 현저하게 짧아진 중수골과 지골은 손이 다른 기초 수각류에게서 추정되는 잡는 기능을 유지했는지에 대한 의문을 제기한다. 케라토사우루스 내에서 팔이 원래 기능을 상실한 아벨리사우루스과와 ''리무사우루스''에서 훨씬 더 극단적인 손 축소를 관찰할 수 있다. 2016년 ''케라토사우루스'' 손의 해부학에 관한 논문에서 카라노와 조나 초이니에르는 손의 형태학적 유사성이 다른 기초 수각류의 손과 매우 유사하다는 점을 강조하며, 짧아졌음에도 불구하고 원래의 잡는 기능을 여전히 수행했을 가능성을 시사했다. 첫 번째 지골만 보존되어 있지만, 잘 발달된 관절면으로 미루어 보아 두 번째 지골은 움직일 수 있었을 것이며, 손가락은 다른 기초 수각류와 유사한 정도의 움직임을 허용했을 것이다. 아벨리사우루스과를 제외한 다른 수각류에서처럼 첫 번째 손가락은 굴곡되었을 때 약간 안쪽으로 돌아갔을 것이다.
4. 3. 두뇌와 감각
마쉬의 감독하에 홀로타입의 뇌강 주형이 만들어졌다. 마쉬는 이를 통해 뇌가 "중간 크기였지만, 초식 공룡보다 비교적 훨씬 컸다"고 결론 내렸다. 그러나 그 후 두개골 뼈가 서로 접착되었기 때문에, 이 주형의 정확성은 후속 연구에서 검증될 수 없었다.
콜로라도주 프루이타에서 표본 MWC 1과 함께 두 번째로 잘 보존된 뇌두가 발견되었으며, CT 스캔을 거쳐 내이, 뇌의 거시적 영역, 그리고 뇌에서 혈액을 운반하는 경막 정맥동의 재구성이 가능해졌다. 2005년, 연구자들은 ''케라토사우루스''가 기초적 수각류에 전형적인 뇌강을 가지고 있으며, ''알로사우루스''와 유사하다는 결론을 내렸다. 후각을 담당하는 후각 구의 인상은 잘 보존되어 있으며, ''알로사우루스''와 유사하지만, 매우 예민한 후각을 갖춘 것으로 여겨지는 ''티라노사우루스''보다는 작았다. 균형 감각을 담당하는 반고리관은 습관적인 머리 방향과 이동에 대한 추론을 가능하게 하며, 다른 수각류에서 발견되는 것과 유사하다. 수각류에서 이러한 구조는 일반적으로 보존적이며, 이는 이동 중 기능적 요구 사항이 종에 걸쳐 유사했음을 시사한다. 반고리관 중 가장 앞쪽이 확대되었으며, 이는 일반적으로 이족 보행 동물에서 발견되는 특징이다. 측면 반고리관의 방향은 머리와 목이 중립 위치에서 수평으로 유지되었음을 나타낸다.
4. 4. 병리학적 증거
''C. nasicornis''의 모식 표본은 왼쪽 2~4 중족골이 융합된 채로 발견되었다. 1884년, 오트니엘 찰스 마시는 당시 공룡에게서는 알려지지 않았던 이 특징에 대한 짧은 논문을 썼으며, 현대 조류에서 보이는 상태와 매우 유사하다고 지적했다. 이러한 특징이 ''케라토사우루스''에 존재한다는 사실은 1890년에 게오르그 바우어가 모식 표본의 융합이 골절의 결과로 치유된 것이라고 추측하면서 논란이 되었다. 이 주장은 1892년 코프에 의해 반복되었는데, 그는 ''C. nasicornis''가 이러한 속 사이의 해부학적 차이가 불충분하다는 이유로 ''메갈로사우루스''의 종으로 분류되어야 한다고 주장했다.그러나 병리학적 기원이 아닌 공룡의 융합된 중족골의 예가 그 이후로 기술되었으며, 여기에는 ''케라토사우루스''보다 기저적인 분류군도 포함된다. 1920년, 오스본은 비정상적인 골 성장이 보이지 않으며, 융합은 특이하지만 병리학적이지 않을 가능성이 높다고 설명했다. 1976년, 로널드 라트케비치는 이 융합이 동물의 달리기 능력을 제한했다고 주장했지만, 이 주장은 1988년 폴에 의해 기각되었는데, 그는 동일한 특징이 오늘날의 지상 조류와 유제류를 포함한 많은 빠른 움직임을 보이는 동물들에게서 나타난다고 지적했다.
1999년 대런 탱크와 브루스 로스차일드의 분석은 융합이 실제로 병리학적이며, 바우어의 초기 주장을 확인한다고 제안했다. 다른 병리학적 보고서에는 이 속에 할당된 발뼈의 스트레스 골절과 골절 이후 추가적인 마모의 징후를 보이는 미확인 ''케라토사우루스'' 종의 부러진 치아가 포함된다.
5. 고환경과 고지리 분포
북아메리카에서 발견된 모든 ''케라토사우루스'' 화석은 모리슨 층에서 발견되었는데, 이곳은 얕은 해양 및 충적 평원 퇴적암으로 구성되어 있으며 북미 대륙에서 공룡 뼈가 가장 많이 발견되는 곳이다. 방사성 연대 측정법에 따르면, 모리슨 층은 1억 5630만 년 전[9]부터 1억 4680만 년 전[10] 사이로, 후기 쥐라기의 옥스포드절 후기, 킴머리지절, 초기 티토니아절에 해당한다. ''케라토사우루스''는 킴머리지절과 티토니아절 지층에서 발견되었다. 모리슨 층은 뚜렷한 우기와 건기가 있는 반건조 기후 환경이었으며, 모리슨 분지는 뉴멕시코에서 앨버타와 서스캐처원까지 뻗어 있었다. 로키 산맥의 전신이 서쪽으로 솟아오르면서 동쪽을 향한 배수 유역에서 유입된 퇴적물은 하천과 강에 의해 늪지 저지대, 호수, 강바닥, 충적 평원에 퇴적되었다.[11] 이 지층은 포르투갈의 로린냐 층과 탄자니아의 텐다구루 층과 연대가 비슷하다.[16]
모리슨 층은 거대한 용각류 공룡이 지배하던 환경이었다.[12] 모리슨 층에서는 수각류 ''코파리온'', ''스토케소사우루스'', ''오르니톨레스테스'', ''알로사우루스'', ''토르보사우루스'', 용각류 ''아파토사우루스'', ''브라키오사우루스'', ''카마라사우루스'', ''디플로도쿠스'', 조반류 ''캄프토사우루스'', ''드라요사우루스'', ''나노사우루스'', ''가고일레오사우루스'', ''스테고사우루스'' 등이 발견되었다.[13] ''알로사우루스''는 전체 수각류 표본의 70~75%를 차지하며 모리슨 먹이 그물의 최상위 영양 단계에 있었다.[14] 이 환경에는 경골어류, 개구리, 도롱뇽, ''울롭스''와 같은 거북, 스페노돈트, 도마뱀, 육상 및 수생 악어(예: ''호플로수쿠스''), 여러 종의 익룡(예: ''하르팍토그나투스'', ''메사닥틸루스'')도 서식했다. 이매패류와 수생 달팽이 껍질도 흔하며, 녹조류, 곰팡이, 이끼, 속새, 소철, 은행나무, 여러 과의 구과식물 화석이 발견되었다. 식생은 고사리와 양치식물로 이루어진 강변 숲(갤러리 숲)에서 아라우카리아와 유사한 구과식물인 ''브라키필룸''과 같은 나무가 드문드문 있는 양치식물 사바나까지 다양했다.[15]
''케라토사우루스''의 부분적인 표본은 포르투갈 로린냐 층의 포르투 노보 구성원에서 발견되었다. 로린냐 층의 많은 공룡들은 모리슨 층과 동일한 속이거나 밀접한 관계를 가진 종이다.[16] ''케라토사우루스'' 외에도 포르투갈에서는 ''알로사우루스''와 ''토르보사우루스''가 발견되었으며, ''로린하노사우루스''는 포르투갈에서만 보고되었다. 포르투 노보 구성원의 초식 공룡으로는 ''디네이로사우루스''와 ''즈비''를 비롯한 용각류와 스테고사우루스인 ''미라가이아'' 등이 있다. 후기 쥐라기 동안 유럽은 좁은 대서양으로 북아메리카와 분리되어 있었고, 이베리아 반도의 일부인 포르투갈은 유럽의 다른 지역과 분리되어 있었다. 마테우스(Mateus)와 동료들은 포르투갈과 북미 수각류의 유사성이 임시 육교의 존재를 나타낸다고 보았다. 그러나 말라파이아(Malafaia)와 동료들은 용각류, 거북, 악어와 같은 다른 그룹이 포르투갈과 북미에서 뚜렷하게 다른 종 구성을 보이기 때문에 더 복잡한 시나리오를 제시했다. 즉, 이들 동물 군의 초기 분리는 일부 그룹에서는 교환을, 다른 그룹에서는 동소적 종분화를 초래했을 수 있다.
참조
[1]
서적
Cassell's Latin Dictionary
Cassell Ltd.
[2]
뉴스
David Koch Donates $35 Million to National Museum of Natural History for Dinosaur Hall
https://www.washingt[...]
2012-05-03
[3]
웹사이트
The Good, Bad and Ugly: Dismantling Historic Fossil Displays, Part Two
http://nmnh.typepad.[...]
Smithsonian National Museum of Natural History
2017-12-04
[4]
논문
"Ceratosaurus" (Theropoda, Ceratosauria) teeth from the Tacuarembó Formation (Late Jurassic, Uruguay)
2020-11-01
[5]
논문
Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods
2014-03-05
[6]
논문
Comments on the ecology of Jurassic theropod dinosaur ''Ceratosaurus'' (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle
https://yadda.icm.ed[...]
[7]
논문
Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic
https://www.academia[...]
[8]
논문
High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem
2020-05-27
[9]
논문
Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations
[10]
서적
The Morrison Formation: An Interdisciplinary Study
Taylor and Francis Group
[11]
서적
An Odyssey in Time: Dinosaurs of North America
NorthWord Press
[12]
서적
Jurassic West: The Dinosaurs of the Morrison Formation and Their World
Indiana University Press
[13]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[14]
서적
Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.
New Mexico Museum of Natural History and Science
[15]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[16]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[17]
문서
Comments on the ecology of Jurassic theropod dinosaur Ceratosaurus (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle
[18]
문서
The evolution of animal weapons
[19]
문서
Comments on the ecology of Jurassic theropod dinosaur Ceratosaurus (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle
[20]
문서
Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues
[21]
문서
世界の巨大恐竜博2006 図録 P63
[22]
웹사이트
狙うは大物!ケラトサウルス
https://ameblo.jp/ol[...]
2006-07-07
[23]
문서
ジュニア自然図鑑 恐竜と古代生物 ジュラ紀〜ケラトサウルスの項
[24]
문서
Comments on the ecology of Jurassic theropod dinosaur Ceratosaurus (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle
[25]
논문
Restorations of ''Claosaurus'' and ''Ceratosaurus''
https://babel.hathit[...]
[26]
논문
Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera ''Antrodemus'' (''Allosaurus'') and ''Ceratosaurus''
http://repository.si[...]
[27]
서적
Predatory Dinosaurs of the World
https://archive.org/[...]
Simon & Schuster
[28]
서적
The Dinosauria: Second Edition
University of California Press
[29]
논문
Principal characters of American Jurassic dinosaurs, part VIII: The order Theropoda
http://marsh.dinodb.[...]
[30]
논문
My theropod is bigger than yours ... or not: estimating body size from skull length in theropods
[31]
논문
Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods
2014-03-05
[32]
논문
Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods
[33]
서적
Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.
New Mexico Museum of Natural History and Science
[34]
서적
Feathered Dragons: Studies on the Transition from Dinosaurs to Birds
Indiana University Press
[35]
논문
Hindlimb allometry in the Late Jurassic theropod dinosaur ''Allosaurus'', with comments on its abundance and distribution
2006
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com