켈렌켄
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
켈렌켄은 약 1,500만 년 전인 중신세에 살았던 멸종된 대형 조류로, 아르헨티나 파타고니아 지역에서 발견된 화석을 통해 알려졌다. 포루스라코스과에 속하며, 알려진 포루스라코스과 조류 중 가장 큰 종으로, 머리 높이가 최대 3m에 달했을 것으로 추정된다. 거의 완전한 두개골 화석을 통해, 켈렌켄은 튼튼한 부리와 강력한 턱 근육을 가지고 있었으며, 다른 포루스라코스과에 비해 두개골의 특징적인 차이를 보였다. 육상 포식자 또는 청소동물로서, 날지 못하고 긴 다리를 이용하여 먹이를 사냥했을 것으로 추정된다. 켈렌켄이 발견된 콜론 쿠라 지층은 반건조 기후의 초원 환경이었으며, 다양한 포유류, 조류, 파충류 화석이 발견된다.
더 읽어볼만한 페이지
- 공포새과 - 티타니스
티타니스는 플라이오세와 플라이스토세 시대에 북미와 남미에 서식했던 포루스라코스과의 육식성 거대 조류로, 튼튼한 골격과 발톱, 퇴화한 날개를 지녔으며, 빠른 속도로 달리고 강력한 다리와 턱으로 사냥했을 것으로 추정되나, 대형 포유류와의 경쟁, 기후 변화, 인류의 영향 등으로 멸종된 것으로 보이며 문화 콘텐츠 소재로도 활용된다. - 마이오세 남아메리카의 새 - 포루스라코스
포루스라코스는 '공포새'라는 별칭으로 불리는 멸종된 거대 육식 조류로, 남아메리카에서 6,200만 년 전부터 180만 년 전까지 생존했으며, 강력한 부리와 다리를 가진 시조새류에 속한다. - 절멸한 날지 못하는 새 - 도도아과
도도아과는 날지 못하는 거대한 몸집의 도도와 로드리게스도도를 포함하는 비둘기과의 아과로, 인간의 남획으로 멸종되어 생태계 보존의 중요성을 보여주며, 니코바르비둘기와 계통 분류학적으로 가까운 관계를 가진다. - 절멸한 날지 못하는 새 - 레이산뜸부기
레이산뜸부기는 하와이 제도 레이산 섬의 고유종으로, 날지 못하고 무척추동물을 먹었으며, 서식지 파괴와 쥐의 유입으로 인해 1944년에 멸종되었다.
| 켈렌켄 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 화석 범위 | 신제3기중신세중기 중신세 (~15백만 년 전) |
| 학명 | Kelenken guillermoi |
| 명명자 | Bertelli et al., 2007 |
| 생물 분류 | |
| 계 | 진핵생물 |
| 문 | 척삭동물문 |
| 강 | 조강 |
| 목 | 노간모지목 |
| 과 | 포루스라코스과 |
| 아과 | 포루스라코스아과 |
| 속 | 케렌켄속 |
| 특징 | |
| 추정 키 | 약 3미터 |
| 두개골 길이 | 716 밀리미터 |
| 부리 길이 | 437 밀리미터 |
| 추정 무게 | 100 킬로그램 |
2. 발견 및 명명
포루스라코스과(Phorusrhacidae)는 신생대 남아메리카 대륙에 서식했던 거대한 육식성 날지 못하는 새 그룹으로, 흔히 '공포새'라고도 불린다. 이들은 긴 뒷다리, 좁은 골반, 상대적으로 작은 날개, 그리고 크고 강력한 갈고리 모양의 부리를 가진 두개골이 특징이다.[6][28]
2004년경, 아르헨티나의 고등학생 기예르모 아기레-사발라가 아르헨티나 파타고니아 지역 리오네그로주 북서부에 위치한 작은 마을 코말로의 철도에서 약 100m 떨어진 곳에서 포루스라코스류(공포새)의 화석을 발견했다. 이 화석이 발견된 노두는 콜론 쿠라 지층에 속한다. 아기레-사발라는 직접 화석 처리 작업을 했으며, 이 발견은 그가 심리학에서 고생물학 및 지구과학으로 학문적 관심을 바꾸는 계기가 되었다.[1][2][3]
발견된 표본은 리오네그로주에 있는 바릴로체 고생물학 협회 박물관(Museo Asociación Paleontológica Bariloche)에 소장되었고, 표본 번호 BAR 3877-11로 분류되었다.[3] 이 동물이 학명을 받기 전인 2006년, 아르헨티나의 고생물학자 루이스 M. 치아페와 사라 베르텔리가 짧은 논문을 통해 이 표본에 대해 보고하고 논의했다.[3][10] 2007년, 베르텔리, 치아페, 클라우디아 탐부시는 이 표본을 ''켈렌켄 길리에르모이''(''Kelenken guillermoi'')의 모식 표본(holotype)으로 지정하여 정식으로 명명했다. 속명 '켈렌켄'은 파타고니아 테우엘체족 신화에 등장하는 거대한 맹금의 모습을 한 정령의 이름에서 따왔으며, 종명 '길리에르모이'는 발견자인 기예르모 아기레-사발라를 기리기 위해 붙여졌다.[3]
모식 표본이자 현재까지 알려진 유일한 표본은 거의 완전한 두개골로 이루어져 있다. 이 두개골은 위아래로 약간 눌린 상태이며, 안와(눈구멍) 뒤쪽의 구개골 대부분은 소실되었지만, 안와의 대부분, 두개골 지붕, 뇌두, 왼쪽 사두골 등은 잘 보존되어 있다. 표본에는 두개골 외에도 관련된 왼쪽 부척골(새의 다리뼈), 발 말절골(발가락뼈)의 윗부분 일부, 그리고 몇 개의 불확실한 뼛조각이 포함되어 있다. 연구자들은 이 뼈들이 함께 발견되었고 주변에 다른 화석이 섞여 있지 않았으며, 색상이나 질감 등 전반적인 보존 상태가 유사하고, 큰 포루스라코스류의 특징과 형태학적으로 일치한다는 점을 근거로 이 뼈들이 모두 단일 개체에 속한다고 결론지었다. 이 표본은 발견 당시까지 알려진 대형 포루스라코스류의 두개골 중 가장 완전한 것이었다. 이전에는 ''데빈첸지아''(''Devincenzia'')나 ''포루스라코스''(''Phorusrhacos'') 같은 다른 대형 포루스라코스류의 두개골은 단편적인 형태로만 알려져 있었고, 특히 ''포루스라코스''의 두개골은 발견 당시 부리 끝부분만 남고 부서져서 ''켈렌켄''이 발견되기 전까지는 서로 다른 크기의 포루스라코스류 분류군 간의 비교 연구가 어려웠다.[3]
3. 분류 및 진화
2004년경, 아르헨티나의 고등학생 기예르모 아기레-사발라(Guillermo Aguirre-Zabala)는 파타고니아 지역 리오네그로주 북서부의 작은 마을 코말로(Comallo)에서 포루스라코스류(Phorusrhacidae, 일명 '공포새') 화석을 발견했다. 이 화석은 기찻길에서 약 100m 떨어진 두 집 사이에서 발견되었으며, 해당 노두는 콜론 쿠라 지층(Collón Curá Formation)에 속한다. 아기레-사발라는 직접 화석 처리를 했고, 이 발견을 계기로 고생물학 및 지구 과학 연구로 진로를 바꾸었다.[1][2][3]
발견된 표본은 리오네그로의 바릴로체 고생물학 협회 박물관(Museo Asociación Paleontológica Bariloche) 소장품(표본 번호 BAR 3877-11)이 되었다.[3] 아르헨티나 고생물학자 루이스 M. 치아페(Luis M. Chiappe)와 사라 베르텔리(Sara Bertelli)는 2006년 짧은 논문에서 이 표본을 보고했다.[3][10] 2007년, 베르텔리, 치아페, 클라우디아 탐부시(Claudia Tambussi)는 이 표본을 켈렌켄 길리에르모이(''Kelenken guillermoi'')의 모식 표본(holotype)으로 공식 지정했다. 속명 '켈렌켄'은 파타고니아 테우엘체족 신화에 등장하는 거대한 맹금류 정령의 이름에서 유래했으며, 종명 '길리에르모이'는 발견자인 아기레-사발라를 기리기 위해 붙여졌다.[3]
모식 표본이자 현재까지 알려진 유일한 표본은 거의 완전한 두개골(약간 눌린 상태), 왼쪽 부척골(tarsometatarsus), 발가락뼈(phalanx) 일부 등으로 구성된다. 발견 당시 이 두개골은 대형 포루스라코스류 중에서 가장 완전한 상태였으며, 이전에는 ''데빈첸지아(Devincenzia)''나 ''포루스라코스(Phorusrhacos)''의 파편적인 두개골만 알려져 있었다.[3]
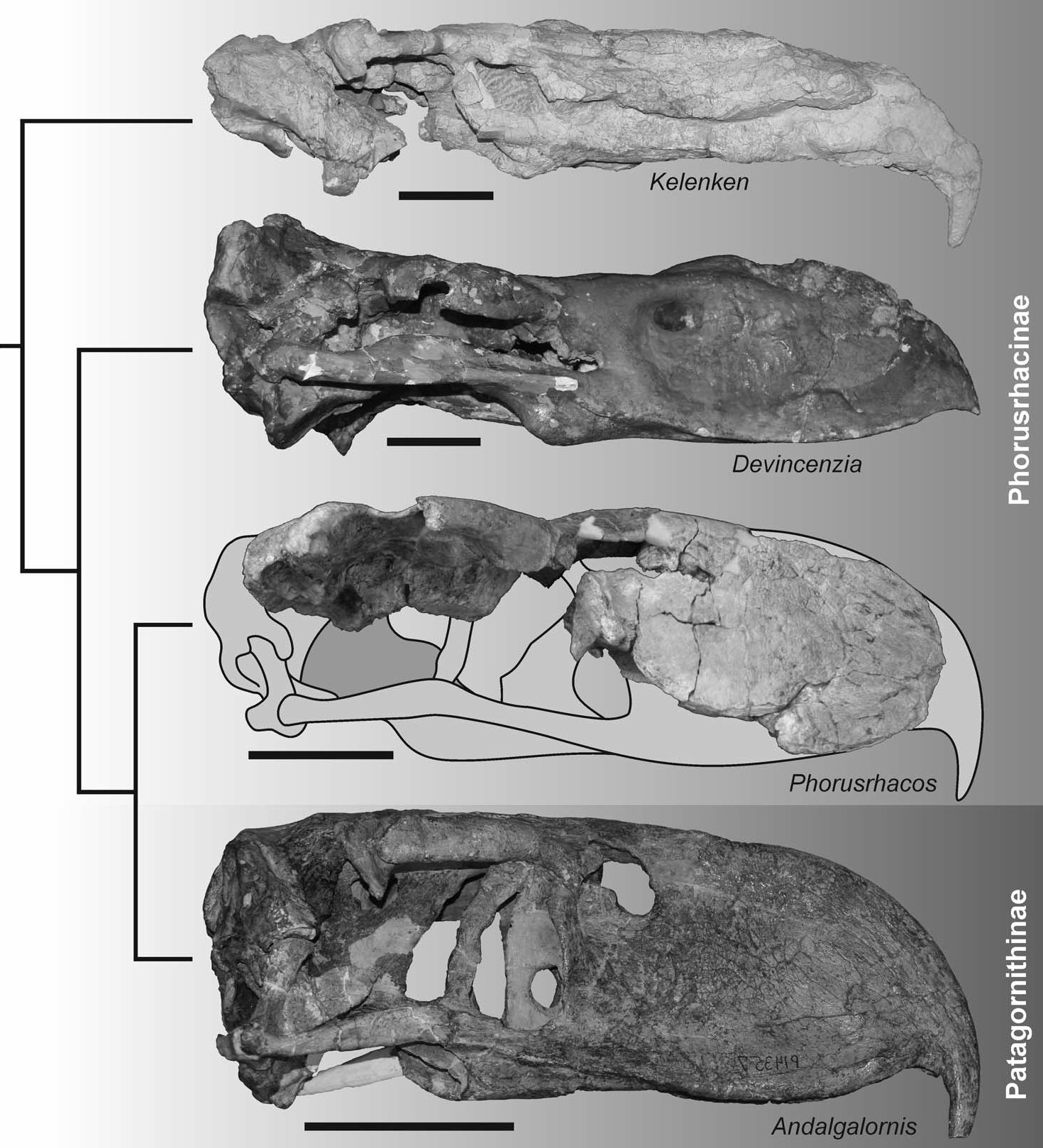
2007년 발표된 연구에서 베르텔리와 동료들은 켈렌켄을 거대한 크기, 옆으로 납작하고 강력한 갈고리 모양의 부리, 볼록한 윗부리 등을 근거로 포루스라코스과(Phorusrhacidae)로 분류했다. 당시 포루스라코스과는 5개의 아과(Brontornithinae, Phorusrhacinae, Patagornithinae, Mesembriornithinae, Psilopterinae)로 나뉘었으나, 분지 분석을 통해 이 분류의 타당성이 완전히 확인되지는 않았다. 연구자들은 켈렌켄이 전통적으로 포루스라코스아과(Phorusrhacinae)로 분류되던 그룹과 가장 유사하다고 보았다. 포루스라코스아과와 공유하는 특징으로는 두개골 뒤쪽이 낮고 압축된 형태, 넓은 후두부 평면(occipital table), 뭉툭한 후안와 돌기(postorbital process) 등이 있으며, 부척골은 하단 융기 표면이 평평하다는 점에서 ''티타니스(Titanis)''와 유사했다. 그러나 포루스라코스아과 자체에 대한 해부학적 정보 부족으로 더 자세한 비교는 어려웠다.[3][11]
2011년 브라질 고생물학자 에르쿨라누 알바렌가(Herculano Alvarenga)와 동료들이 발표한 계통 발생 분석 결과, 켈렌켄은 ''데빈첸지아(Devincenzia)''와 가장 가까운 자매 분류군으로 나타났다. 이 분석은 5개 아과의 존재를 지지했지만, 분기도 상에서는 Brontornithinae, Phorusrhacinae, Patagornithinae 아과가 명확히 구분되지 않았다.[4] 2015년 아르헨티나 고생물학자 페데리코 J. 데그란헤(Federico J. Degrange)와 동료들은 ''라라와비스(Llallawavis)''를 기술하며 새로운 계통 발생 분석을 수행했다. 이 연구에서 포루스라코스아과(Phorusrhacinae)는 다계통군(polyphyletic group), 즉 자연적인 분류군이 아님이 밝혀졌다. 2015년 분석에 따르면, 켈렌켄은 데빈첸지아, 티타니스 등 다른 대형 포루스라코스류와 함께 분기하지만, 기존의 포루스라코스아과는 여러 계통이 섞인 다계통군으로 나타났다.[5]
비조류 공룡 멸종 이후 초기 신생대 동안 포유류가 진화적 방산을 겪는 한편, 일부 조류 그룹들도 거대동물로 진화하는 경향을 보였다. 여기에는 가스토르니스과(Gastornithidae), 드로모르니스과(Dromornithidae), 고악류(Palaeognathae), 그리고 포루스라코스과가 포함된다.[4][8] 포루스라코스과는 느시사촌목(Cariamiformes)에 속하는 멸종된 그룹으로, 현존하는 유일한 친척은 느시사촌과(Cariamidae)에 속하는 2종의 세리에마(seriema)뿐이다. 포루스라코스과는 느시사촌목 내에서 가장 종 다양성이 높은 그룹이었지만, 화석 기록이 불완전하여 그룹 내 상호 관계는 아직 명확하지 않다.[6]
포루스라코스과는 시신세(Eocene)부터 남아메리카 대륙(당시 고립된 섬 대륙)에 존재했으며 플라이스토세(Pleistocene)까지 생존했다. 이들은 플리오세(Pliocene) 말 아메리카 대륙간 대교환 시기에 북아메리카로 진출하기도 했다. 유럽에서 발견된 일부 화석이 포루스라코스과로 분류되기도 했으나 논란이 있다. 이 그룹의 기원이 남아메리카인지, 혹은 다른 지역에서 유입되었는지는 불분명하다.[11][4] 포루스라코스과는 플라이스토세까지 생존하여, 플리오세에 멸종한 남아메리카 고유의 포식성 유대류인 틸라코스밀루스과(Thylacosmilidae) 등보다 더 오랫동안 번성했으며, 북아메리카에서 유입된 태반류 포식자들과 생태적으로 경쟁했을 가능성이 있다.[7]
켈렌켄 자체는 포루스라코스과가 멸종하기 훨씬 전인 약 1,500만 년 전, 중기 마이오세(Middle Miocene)에 살았다. 이 시기는 아직 남아메리카에 대형 육식성 태반류 포유류가 진출하기 전이며, 틸라코스밀루스와 같은 토착 포식 동물들이 존재하던 때였다.[3]
4. 특징
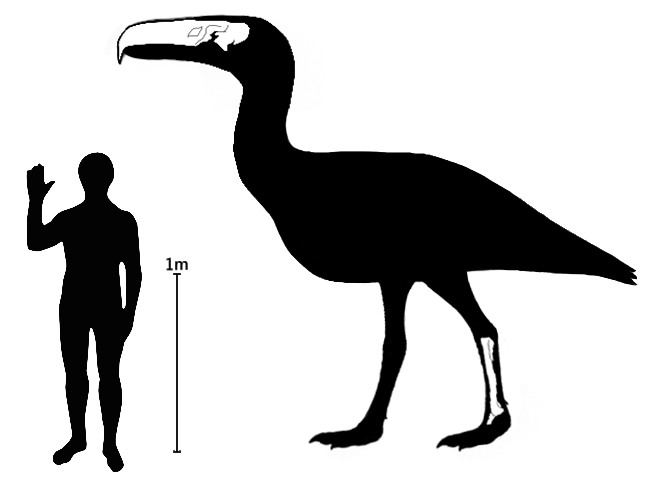
켈렌켄(''Kelenken guillermoi'')은 지금까지 알려진 포루스라코스과 중에서 가장 큰 종이다.[1][23] 이전에 가장 큰 종으로 알려졌던 포루스라코스(Phorusrhacos)보다 약 10% 더 컸던 것으로 추정된다.[3] 켈렌켄의 키는 최대 3m에 달했을 것으로 보이며,[8][30] 이는 현생 세리에마(seriema, 높이 90cm)와 비교하면 엄청난 크기이다.[8][31] 켈렌켄의 두개골 길이는 약 71.6cm로,[3][25] 이는 현존하는 모든 조류를 통틀어 가장 크며, 심지어 말의 두개골 크기와 비슷하다.[1][3][23][25] 발목과 발바닥뼈가 합쳐진 부척골의 길이는 43.7cm에 달했다.[3][25] 정확한 체중 추정치는 없지만, 100kg은 넘었을 것으로 여겨진다.[8][9][30][31]
켈렌켄의 유일하게 알려진 표본(BAR 3877-11)은 아르헨티나 파타고니아의 중신세 지층인 콜론 쿠라 지층(Collón Curá Formation)에서 발견되었으며, 거의 완전한 두개골과 일부 다리뼈로 이루어져 있다.[3][25] 이 표본, 특히 거의 완전한 상태로 발견된 두개골은 거대 포루스라코스과의 형태 연구에 중요한 기여를 했다.[3][10][25][32] 상세한 두개골과 다리뼈의 형태는 하위 섹션에서 다룬다.
4. 1. 두개골
켈렌켄의 모식 표본(Holotype)이자 유일하게 알려진 표본(BAR 3877-11)은 거의 완전한 두개골로 구성되어 있다. 이 두개골은 위아래로 약간 눌린 상태이며, 대부분의 안와(눈구멍), 두개골 지붕, 뇌두개, 왼쪽 방형골이 보존되어 있으나, 안와 뒤쪽의 구개골 대부분은 유실되었다. 표본은 아르헨티나 리오네그로주 바릴로체에 있는 '무세오 아소시아시온 팔레온톨로히카 바릴로체'(Museo Asociación Paleontológica Bariloche)에 소장되어 있다.[3][25] 표본에는 두개골 외에도 왼쪽 부척골(새의 발목과 발바닥뼈가 합쳐진 뼈), 발가락뼈(지골)의 윗부분 일부, 그리고 몇 개의 불확실한 뼛조각이 포함되어 있다. 이 뼈들은 함께 발견되었고 보존 상태(색, 질감 등)가 비슷하며, 모두 대형 포루스라코스과 조류의 특징을 보여 단일 개체의 것으로 여겨진다.[3][25]
이 두개골은 발견 당시 알려진 대형 포루스라코스류의 것 중 가장 완전한 상태였다. 이전에는 데빈첸지아(Devincenzia)나 포루스라코스(Phorusrhacos) 같은 대형 종의 두개골은 매우 단편적인 상태로만 알려져 있었고, 특히 포루스라코스의 두개골은 발견 과정에서 부리 끝만 남기고 부서져 버렸기 때문에, 켈렌켄의 발견은 이 그룹 내 다양한 크기의 종들을 비교 연구하는 데 중요한 계기가 되었다.[3][25] 켈렌켄이 발견되기 전까지, 대형 포루스라코스류의 두개골은 프실로프테루스(Psilopterus)나 파타고르니스(Patagornis)처럼 더 작지만 완전한 표본이 알려진 근연종의 두개골을 단순히 확대한 모습으로 복원되곤 했다. 예를 들어, 1895년에 그려진 포루스라코스 두개골 스케치는 파타고르니스를 기반으로 했는데, 이는 매우 높은 부리, 둥글고 높은 눈구멍, 아치형 뇌 상자를 특징으로 했다. 그러나 켈렌켄의 두개골은 이 그룹의 큰 종과 작은 종 사이에 상당한 형태적 차이가 있음을 보여주었다.[3][10][25][32]

켈렌켄의 완모식 두개골은 매우 거대하며, 위에서 보면 삼각형 모양이고 뒤쪽 부분이 위아래로 눌려 있다. 두개골 길이는 부리 끝에서 뒤통수까지 약 71.6cm에 달하며, 이는 알려진 모든 조류의 두개골 중 가장 큰 것이다.[23][25][30] 위턱 부리는 매우 길어서 두개골 전체 길이의 절반 이상(약 56%)을 차지하며, 이는 메셈브리오르니스(Mesembriornis), 파타고르니스, 포루스라코스보다도 긴 비율이다. 위아래로 눌린 점을 감안해도 부리는 높고 매우 튼튼하지만, 파타고르니스, 안드레우소르니스(Andrewsornis), 안달갈로르니스(Andalgalornis)와 같은 파타고르니스아과(Patagornithinae) 조류만큼 극단적으로 높지는 않았던 것으로 보인다.[3][25]
전상악골(부리 끝부분 뼈)의 앞쪽 끝은 날카롭고 아래로 강하게 구부러진 갈고리 모양으로 돌출되어 있다. 이는 작은 프실로프테루스아과(Psilopterinae)의 약한 돌출과는 다르며, 포루스라코스, 파타고르니스, 안드레우소르니스, 안달갈로르니스 같은 중대형 포루스라코스류와 유사한 특징이다. 위턱 부리 아랫면에는 두 개의 두드러진 능선이 있으며, 이는 부리의 날카로운 가장자리(tomium)에서 홈으로 분리된다. 이 능선들은 또한 부리 중앙의 세로 홈에 의해 서로 분리된다. 파타고르니스도 입 앞부분에서 비슷한 구조를 보인다. 위턱 부리 측면 대부분에는 신경 혈관이 지나갔을 것으로 추정되는 작은 불규칙한 구멍들이 덮여 있다. 부리 뒤쪽 2/3에는 가장자리와 평행하게 뻗은 뚜렷한 홈이 파여 있다.[3][25]
콧구멍은 작고 직사각형이며, 파타고르니스아과처럼 위턱 부리의 위쪽 뒤편에 위치한다(더 큰 포루스라코스아과나 브론토르니스아과에서는 콧구멍의 크기와 위치가 불분명하다). 콧구멍은 높이보다 앞뒤로 더 길어 보이지만, 이는 화석화 과정에서의 압착 때문에 과장되었을 수 있다. 콧구멍 뒤쪽 가장자리는 비골의 상악골을 향한 돌기에 의해 형성된다. 콧구멍 사이에 중격이 없어 서로 연결되었는지는 불분명하다. 눈구멍 앞의 구멍인 전안와창은 양쪽이 약간 눌렸음에도 사다리꼴 모양임이 분명하다. 앞쪽 경계는 콧구멍 뒤쪽 가장자리와 거의 같은 높이에 있고, 아래쪽 경계는 직선이다. 튼튼한 누골이 전안와창의 뒤쪽 경계를 이루며, 이 뼈는 협골궁(눈구멍 아래쪽 경계)과 전두골(이마뼈)의 바깥쪽 가장자리에 비해 뒤로 들어가 있다. 전안와창은 파타고르니스의 것보다 비례적으로 작다.[3][10][25][32]
눈구멍(안와)의 모양은 위아래 압착의 영향을 받았을 수 있지만, 원래는 위아래로 낮고 거의 직사각형이었을 가능성이 높다. 위쪽 가장자리는 오목하고 아래쪽 경계는 약간 볼록하다. 눈구멍 위쪽은 두껍고 둥근 가장자리(상안와 골화)로 둘러싸여 있으며, 그 뒤쪽 부분은 옆에서 볼 때 아래로 돌출되어 보인다. 파타고르니스에서는 비슷한 구조가 누골의 돌기로 제안되었는데, 켈렌켄에서도 이 구조는 누골의 연장일 가능성이 높다. 이 상안와 골화는 후안와 돌기를 형성하는 전두골 일부에 의해 만들어진 소켓 안에 자리 잡고 있는데, 이는 다른 포루스라코스과에서는 알려지지 않은 특징이다. 눈구멍 아래쪽 가장자리는 매우 높고 좌우로 납작하며 튼튼한 협골궁으로 형성되어 있다. 이 협골궁은 데빈첸지아의 것보다 크며, 눈구멍 아래 중앙 부분에서의 높이가 두께의 약 4배에 달해 다른 포루스라코스과보다 훨씬 높다.[3][25]
전두골은 윗면이 평평했던 것으로 보인다. 전두골과 전상악골이 만나는 부분은 손상되어 봉합선을 확인할 수 없지만, 전두골과 비골, 두정골 사이의 봉합선은 완전히 융합되어 있다. 이 때문에 이 뼈들이 두개골 지붕을 어떻게 구성했는지 정확히 알기 어렵지만, 뭉툭하고 튼튼한 후안와 돌기는 주로 전두골에 의해 형성되었을 것이다. 각 전두골 아랫면에는 턱 근육이 부착되었던 큰 함몰부가 있다. 후안와 돌기는 튼튼한 관골 돌기(zygomatic process)와 좁은 간격으로 분리되어 있으며, 이 두 돌기가 좁은 측두창(관자놀이 부분의 구멍)을 둘러싼다. 후안와 돌기에는 거대한 턱 근육이 남긴 흉터가 있으며, 이 흉터 중 일부는 두정골 높이에서 두개골 지붕 대부분을 침범했다. 관골 돌기 뒤쪽, 측두골을 따라서는 턱을 닫는 근육에 해당하는 잘 발달된 함몰부가 있다. 그 뒤의 후측두와(subtemporal fossa)는 넓고, 뒤쪽 경계는 옆으로 뻗은 뭉툭한 후두 능선으로 정의된다.[3][25]
상악골은 넓은 입천장(구개)을 형성한다. 구개의 측면 가장자리는 위턱 부리 길이의 대부분에서 거의 평행하며, 눈구멍 근처에서 옆으로 넓어진다. 파타고르니스처럼, 상악골은 중앙선에서 뚜렷한 세로 방향 함몰부로 나뉘며, 구개 뒤쪽 절반에서는 이 함몰부가 상악골의 일부에 의해 양옆으로 둘러싸인다. 상악골 뒤쪽 측면 가장자리는 협골과 명확한 봉합선으로 연결된다. 눈구멍 뒤쪽의 두개골 지붕은 평평하며, 측두근의 발달로 인한 흉터가 있다. 후두 테이블(occipital table)은 데빈첸지아처럼 매우 넓고 낮아서 뒤에서 보면 직사각형 모양이다. 후두과(머리 뒤쪽에서 첫 번째 경추와 연결되는 둥근 돌기)는 둥글며, 윗면에서 시작해 거의 중앙까지 이어지는 수직 홈이 있다. 대공(두개골 바닥에서 척수가 지나가는 큰 구멍)은 독특하게 거의 삼각형 모양이며 위쪽 꼭지점이 뭉툭하고, 후두과보다 약간 작다. 대공 바로 위에는 능선 같은 돌출부가 수직으로 뻗어 대공 가장자리에서 후두 능선까지 이어진다. 후두과 아래의 움푹 파인 부분(와)은 보이지 않는데, 이는 와가 뚜렷한 파타고르니스나 데빈첸지아와 다른 점이다.[3][10][25][32]
4. 2. 다리뼈
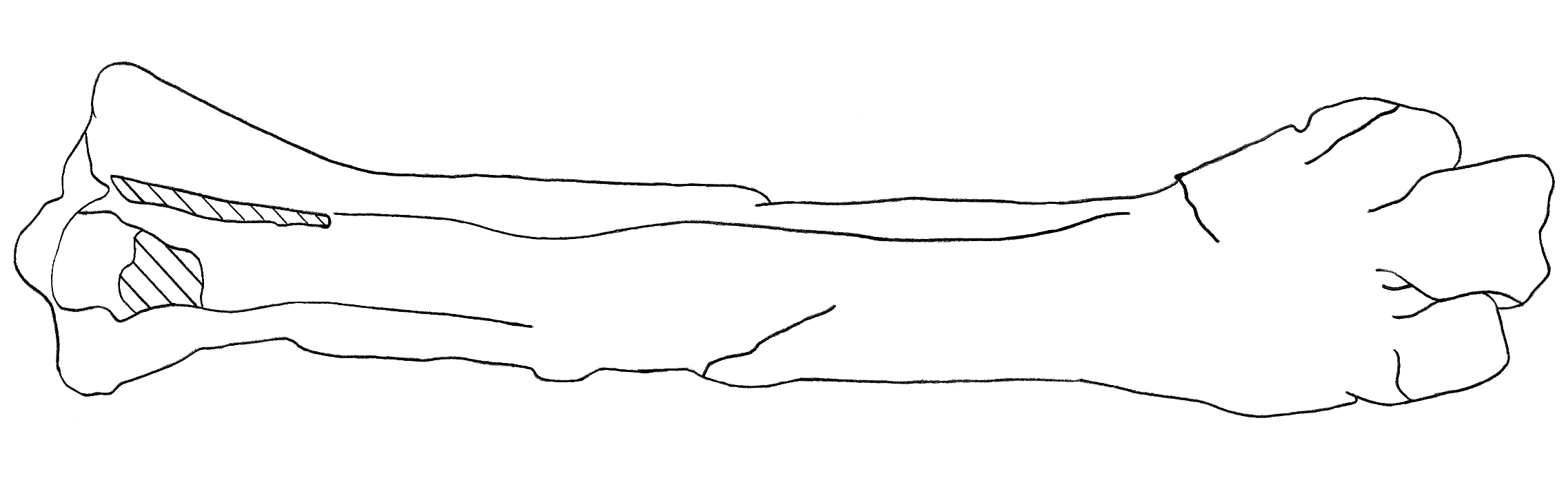
족근중족골의 몸통(축부)은 다소 가늘고 길며, 중간 부분의 단면은 거의 직사각형 형태로 포루스라코스와 유사하다.[3][25] 윗면의 위쪽 2/3는 오목하고 아래쪽 1/3은 더 평평하다.[3][25] 족근중족골 위쪽 끝에는 거의 타원형이고 깊게 파인 두 개의 관절와(cotylae)가 있다. 바깥쪽의 외측 관절와는 안쪽의 내측 관절와보다 작고 약간 더 아래쪽에 위치한다.[3][25] 관절와 사이의 관절와간 융기는 다른 포루스라코스과와 마찬가지로 잘 발달되어 있고 튼튼하다.[3][25] 켈렌켄만의 특징으로, 외측 관절와의 안쪽(내측)면 모서리에 둥근 결절이 있으며, 이 결절은 관절와간 융기보다 높이가 낮다.[3][25]
족근중족골 몸통의 중간 부분 단면은 불규칙한 사각형인데, 이는 매우 넓은 직사각형 단면을 가진 브론토르니스아과(Brontornithinae)와는 다른 점이다.[25] 발가락뼈와 연결되는 족근중족골 아래쪽 끝의 세 개의 활차(trochleae) 중, 세 번째 중족골과 연결되는 가운데 활차가 다른 두 개(두 번째와 네 번째)보다 훨씬 크고 더 아래쪽으로 돌출되어 있다.[3][25] 또한 네 번째 활차가 두 번째 활차보다 크다.[3][25] 네 번째 활차는 불규칙한 사각형 모양으로, 직사각형 활차를 가진 데비첸지아와는 대조적이다.[25] 족근중족골 아래쪽 앞면에 있는 구멍인 원위 혈관 공(distal vascular foramen)은 세 번째와 네 번째 활차의 위쪽 중앙에 위치한다.[3][25]
5. 고생물학

2004년경, 아르헨티나 리오네그로 주 북서부의 작은 마을 코말요(Comallo) 인근 철도에서 약 100m 떨어진 지점에서 당시 고등학생이었던 기예르모 아기레-사발라(Guillermo Aguirre-Zabala)가 포루스라코스과 조류의 화석을 발견했다. 이 표본이 발견된 노두는 콜론 쿠라 지층(Collón Curá Formation)에 속한다. 아기레-사발라는 직접 화석을 처리했으며, 이 발견을 계기로 전공을 심리학에서 고생물학 및 지구과학으로 변경했다.[23][24][25]
표본은 리오네그로 주 바릴로체 고생물학 협회 박물관(Museo Asociación Paleontológica Bariloche)에 소장되었고, 표본 번호 BAR 3877-11을 부여받았다.[25] 2007년, 사라 베르텔리(Sara Bertelli)와 동료들은 이 표본을 모식 표본으로 하여 새로운 속과 종인 ''켈렌켄 기예르모이''(''Kelenken guillermoi'')를 명명했다. 속명 '켈렌켄'은 파타고니아 테우엘체족 신화에 등장하는 거대한 맹금류의 모습을 한 영혼의 이름에서 유래했으며, 종소명 '기예르모이'는 발견자인 아기레-사발라를 기리기 위해 붙여졌다.[25]
모식 표본이자 현재까지 알려진 유일한 표본은 거의 완전한 두개골, 왼쪽 족근중족골(tarsometatarsus), 뒷다리 지골 일부, 그리고 몇몇 미확인 뼛조각으로 구성된다. 이 뼈들은 함께 발견되었고 보존 상태가 동일하며, 형태적으로도 대형 포루스라코스과 조류의 특징과 일치하여 단일 개체의 것으로 여겨진다. 발견 당시 이 두개골은 알려진 대형 포루스라코스과 조류의 두개골 중 가장 완전한 것이었다. 이전까지 알려진 ''데빈켄지아''(Devincenzia)나 ''포루스라코스''(Phorusrhacos)의 두개골은 단편적이어서 분류군 간 비교가 어려웠다.[25]
베르텔리와 동료들은 표본의 크기, 옆으로 납작하고 강하게 구부러진 부리 형태 등을 근거로 켈렌켄을 포루스라코스과로 분류했다. 특히 두개골 뒷부분이 낮고 넓으며, 눈 뒤쪽의 후안와 돌기(postorbital process)가 뭉툭하고, 족근중족골의 형태가 ''티타니스''(Titanis)와 유사하다는 점에서 전통적으로 포루스라코스아과(Phorusrhacinae)로 분류되던 그룹과 가장 유사하다고 보았다.[25][33] 이후 알바렌가(Alvarenga) 등이 2011년에 수행한 계통 분석에서는 켈렌켄이 ''데빈켄지아''와 가장 가까운 자매군 관계에 있음을 시사했다.[26] 그러나 2015년 데그란지(Degrange) 등의 연구에서는 기존의 포루스라코스아과가 다계통군일 가능성을 제기하며 켈렌켄을 포함한 대형 포루스라코스과 그룹의 분류학적 위치에 대한 추가 연구의 필요성을 시사했다.[27]
포루스라코스과는 신생대에 번성했던 날지 못하는 새 그룹으로, 뜸부기목에 속한다. 이들은 공룡 멸종 이후 포유류가 다양해지던 시기에 남아메리카 대륙에서 독자적으로 진화하여 대형화된 육상 포식자 그룹 중 하나이다.[26][30] 고제3기 에오세 무렵 남아메리카에 처음 등장하여 제4기 플라이스토세까지 생존했으며, 신제3기 플라이오세 말에는 아메리카 대륙간 대교환을 통해 북아메리카까지 진출했다.[33][26] 켈렌켄은 포루스라코스과가 멸종하기 훨씬 전인 약 1,500만 년 전, 신제3기 중기 마이오세에 살았다. 이 시기는 아직 북아메리카의 육식성 태반류가 남아메리카로 진출하기 전이었으며, 후수하강 포식 동물들이 존재하던 때였다.[25]
켈렌켄 화석이 발견된 콜론 쿠라 지층은 아르헨티나 파타고니아 지역에 분포하며, 주로 화산 활동으로 생성된 화산쇄설암, 응회암, 석회암, 사암 등으로 이루어져 있다. 이 지층은 강이나 호수와 같은 대륙성 담수 환경에서 퇴적되었다.[25][42] 당시 이 지역의 연대는 약 1,500만 년 전(중기 마이오세, 콜론쿠란절)으로 추정된다.[25][30][41] 이 시기 파타고니아 지역은 오늘날의 반건조 기후 아래 온대 관목지나 삼림과 유사하게 식생 밀도가 낮고 개방된 환경이었을 것으로 보인다. 이는 초기 마이오세 후기의 나무에 사는 동물이 많은 울창한 숲 환경과는 대조적이며, 남아메리카 육상 포유류 시대(SALMA) 중 프리아스절(Friasian) 초기에 점진적으로 일어난 것으로 보인다.[17][18][39][40] 이러한 환경 변화는 중기 마이오세 기후 최적기 이후 진행된 전 지구적인 냉각화 및 건조화 추세와 관련이 있을 수 있다.[18][40] 개방된 환경은 켈렌켄과 같이 달리기에 적응한 대형 동물들이 서식하기에 유리했다.[17][18]
콜론 쿠라 지층에서는 켈렌켄 외에도 다양한 포유류 화석이 발견되었다. 여기에는 피갑류(예: ''Megathericulus'', ''Prepotherium'', ''Prozaedyus'', ''Paraeucinepeltus''), 불유치목(예: ''Protypotherium''), 아스트라포테리움목(예: ''Astrapotherium''), 스파라소돈트목(예: ''파타고스밀루스'', ''클라도식티스''), 유대류(예: ''Abderites''), 영장류(예: ''Proteropithecia''), 그리고 다양한 설치류(예: ''Maruchito'', ''Protacaremys'', ''Neoreomys'', ''Prolagostomus'') 등이 포함된다. 포유류 외에도 조류, 파충류, 양서류, 어류 화석도 함께 발견되어 당시의 풍부한 생물 다양성을 보여준다.[19][20][21][22][17][41][42][43][44]
5. 1. 식성 및 사냥 방식

포루스라코스과는 육상 포식자 또는 청소동물로 여겨지며, 태반류 포유류 포식자가 없었던 신생대 남아메리카 생태계의 최상위 포식자로 간주되기도 한다. 이들은 일부 대형 육식성 보르히에나과 포유류와 공존했다.[6][15] 포루스라코스과의 식성에 관한 초기 가설은 주로 갈고리 모양의 부리를 가진 큰 두개골을 근거로 추론되었으며, 달리기 능력이나 포식 행동에 대한 상세한 가설과 생체역학 연구는 21세기 초부터 시작되었다.[6][15]
알바렌가(Alvarenga)와 엘리자베스 횔플링(Elizabeth Höfling)은 2003년 연구에서 포루스라코스과의 습성에 대한 일반적인 견해를 제시했다. 이들은 날개 크기와 체중 비율로 볼 때 날지 못하는 새였으며, 특히 몸집이 큰 종류일수록 날개가 더 작았다. 연구자들은 좁은 골반, 상악골, 흉곽 등이 키 큰 풀숲이나 울퉁불퉁한 지형에서 작은 동물을 찾고 잡는 데 유리한 적응이었을 수 있다고 보았다. 또한, 눈 위에 돌출된 눈물뼈는 현대의 매처럼 강한 햇빛으로부터 눈을 보호하고 예리한 시력을 유지하게 하여, 그늘진 숲보다는 햇볕이 내리쬐는 개활지에서 시각에 의존해 사냥했음을 시사한다고 주장했다.[11][33]
2010년, 데그란지(Degrange)와 동료들은 중형 포루스라코스과인 안달갈로르니스(''Andalgalornis'')를 대상으로 CT 스캔과 유한 요소 해석을 이용해 물기 힘과 두개골의 응력 분포를 연구했다. 연구 결과, 부리 끝에서의 물기 힘은 133 N으로 추정되었으며, 켈렌켄과 같은 다른 대형 포루스라코스과처럼 두개골 뼈 사이의 움직임(두개내 운동성)이 상당히 줄어들어 두개골의 강성이 높아진 것으로 나타났다. 이는 두개골 강화를 위한 적응으로 해석되었다. 현대의 붉은다리세리마나 흰꼬리수리와 비교했을 때, 포루스라코스과의 두개골은 좌우 방향의 힘에는 비교적 약했지만, 위아래 방향의 힘이나 뒤로 당기는 힘에는 강한 저항력을 보였다. 이러한 측면 방향의 취약성 때문에 연구진은 안달갈로르니스가 크고 버둥거리는 먹이를 부리로 제압하는 위험한 행동을 했을 가능성은 낮다고 보았다. 대신, 작은 먹이를 통째로 삼키거나, 큰 먹이를 사냥할 때는 부리로 여러 번 정밀하게 타격하는 "치고 빠지기" 전술을 사용했을 것으로 제안했다. 날카로운 발톱은 없었지만, 발로 먹이를 제압했을 수도 있다.[15][37]
2012년 탐부시(Tambussi)와 동료들은 안달갈로르니스의 목뼈 형태 분석을 통해 목의 유연성을 연구하여, 목이 세 부분으로 나뉜다는 것을 발견했다. 연구진은 목뼈를 수동으로 조작하여 안달갈로르니스의 목 근육과 골격이 큰 머리를 지탱하고, 아래로 공격한 후 머리를 다시 들어 올리는 동작에 적응했다고 결론지었다. 이러한 특징은 켈렌켄을 포함한 다른 대형 포루스라코스과에도 적용될 수 있다고 추정했다.[16][38] 2020년 데그란지의 포루스라코스과 두개골 형태 연구에서는 세리마와 유사한 조상으로부터 두 가지 주요 두개골 형태가 진화했음을 밝혔다. 하나는 조상 형태에 가까운 "프실롭테린 두개골 유형"이고, 다른 하나는 켈렌켄 및 다른 대형 포루스라코스과를 포함하며 더 단단하고 뻣뻣하게 특수화된 "테러 버드 두개골 유형"이다. 이러한 차이에도 불구하고 두 유형 모두 유사한 방식으로 먹이를 처리했지만, "테러 버드" 유형의 더 강한 두개골과 그에 따른 더 강한 물기 힘은 더 큰 먹이를 사냥하기 위한 적응이었을 것으로 여겨진다.[6][28]
5. 2. 이동 방식

2005년, 루데마르 에르네스토 블랑코(Rudemar Ernesto Blanco)와 워싱턴 W. 존스(Washington W. Jones)는 포루스라코스과의 경골(정강이뼈) 강도를 조사하여 속도를 추정하려 시도했다. 다만, 이러한 추정 방식은 현재 살아있는 동물에게 적용해도 신뢰성이 떨어질 수 있음을 인정했다.[13][35] 연구 결과, ''파타고르니스''(Patagornis)와 특정되지 않은 대형 포루스라코스과의 경골 강도는 14m/s의 속도를 나타냈고, ''메셈브리오르니스''(Mesembriornis)는 27m/s의 속도를 보였다. 후자의 속도는 현생 타조보다 빠르며, 치타의 최고 속도인 29m/s에 근접하는 수치이다.[13][12][35][34]
그러나 블랑코와 존스는 이 새들의 거대한 몸집 때문에 이렇게 빠른 속도로 달리는 것은 현실적으로 어려웠을 것이라고 보았다. 대신, 그들은 이 강한 다리뼈가 사이가나 톰슨가젤 크기의 중간 크기 포유류의 긴 뼈를 부러뜨리는 데 사용되었을 수 있다고 제안했다. 이는 뼈 속의 골수를 먹기 위함이거나, 일부 현생 육상 조류처럼 다리를 발로 차는 무기로 사용했을 가능성을 시사한다. 이러한 추측은 일부 포루스라코스과에서 발견되는 크고 구부러졌으며 옆으로 납작한 발톱의 형태와도 일치한다. 또한, 그들은 향후 연구를 통해 부리와 발톱이 아르마딜로나 글립토돈과 같이 단단한 방어 체계를 갖춘 포유류를 사냥하는 데 사용될 수 있었는지 조사할 필요가 있다고 덧붙였다.[13][35]
2006년, 루이스 키아페(Luis M. Chiappe)와 사라 베르텔리(Sara Bertelli)는 켈렌켄의 발견이 포루스라코스과의 크기와 민첩성 사이에 반비례 관계가 있다는 기존의 통념, 즉 덩치가 클수록 느리고 달리기에 덜 적합하다는 생각에 의문을 제기한다고 주장했다.[10][32] 켈렌켄의 길고 가느다란 발목뼈(족근중족골)는 이 새가 더 작지만 땅딸막하고 느렸던 ''브론토르니스''(Brontornis)보다 훨씬 민첩했을 가능성을 보여준다.[10][32] 켈렌켄 발견에 대한 2006년 뉴스 기사에서 키아페는 켈렌켄이 타조만큼 빠르지는 않았을지라도, 길고 가는 다리뼈(현생 레아와 유사)를 바탕으로 기존에 대형 포루스라코스과에 대해 추정했던 것보다 훨씬 빠르게 달릴 수 있었을 것이라고 언급했다. 이 기사는 켈렌켄이 작은 포유류나 파충류를 쫓아 사냥했을 가능성을 제시했다.[1][23]
또 다른 2006년 뉴스 기사에서 키아페는 켈렌켄이 그레이하운드만큼 빠를 수 있었을 것이라고 추정했다. 당시 남아메리카에는 다른 대형 포식자도 있었지만, 그 수가 제한적이었고 포루스라코스과만큼 빠르고 민첩하지는 않았을 것으로 보았다. 또한, 당시 풍부했던 다양한 초식동물들이 켈렌켄에게 충분한 먹잇감을 제공했을 것이라고 덧붙였다. 키아페는 포루스라코스과가 거대한 머리, 매우 작은 앞다리, 그리고 매우 긴 다리를 가졌다는 점에서 초기 육식 공룡인 ''티라노사우루스''와 외형적으로 유사하며, 비슷한 방식의 포식 적응을 거쳤을 것이라고 설명했다.[14][36]
6. 고환경
''켈렌켄''은 아르헨티나 파타고니아 코말요 남동쪽 모서리의 콜론 쿠라 지층(Collón Curá Formation)에 속하는 화산쇄설암 노두에서 발견되었다. 이 지역은 흰색 응회암으로 덮여 있다.[3][8][19] 해당 지층은 서로 연결되지 않은 여러 개의 구조 분지가 특징인 무너진 전연 분지 시스템에 퇴적되었으며, 주로 충적 환경(흐르는 물에 의해 퇴적됨)과 호수 환경에 쌓인 화산쇄설성 석회암과 사암으로 구성되어 있다.[20][42]
당시 이 지역의 층서는 예비적으로만 연구되었고 퇴적물의 연대는 정확히 결정되지 않았으나, 콜론 쿠라 지층의 여러 지역에서 얻은 방사성 연대 측정 결과와 다른 남아메리카 육상 포유류 시대 화석층과의 비교를 통해 약 1,500만 년 전 중기 마이오세의 콜론쿠란 시대에 해당하는 것으로 추정된다.[3][8][19][25][30][41]
콜론 쿠라 지층과 남아메리카의 콜론쿠란 시대는 식물 덮개가 줄어든 비교적 개방된 환경이 우세했던 시기를 나타낸다. 이는 반건조하고 온화하거나 따뜻하며 건조한 삼림 또는 관목지와 유사했을 것으로 보인다. 이러한 개방적인 환경은 달리기에 적응한 동물과 큰 동물이 서식하기에 유리했으며, 나무 위에서 생활하는 동물이 많은 발달된 숲이 존재했던 초기 마이오세 후반의 이전 환경과는 대조적이었다. 당시 숲은 산맥의 계곡 지역에 제한적으로 분포했고 나무에 서식하는 종은 거의 없었을 것으로 추정된다. 이러한 환경 변화는 점진적으로 일어났으며,[17][18][39][40] 중기 마이오세 기후 전환과 관련된 기후 변화와 동시에 발생했을 가능성이 있다. 이 시기의 지구 냉각 사건은 남아메리카 대륙에 건조한 영향을 미쳤을 것으로 보인다.[18][40]
콜론 쿠라 지층에서는 다양한 포유류 화석이 발견되었으며, 최소 24개 분류군 이상이 보고되었다. 이 외에도 조류, 파충류, 양서류, 어류의 화석도 발견된다.[17][39] 주요 포유류 화석은 다음과 같다.[19][20][21][22][41][42][43][44]
| 분류 | 속명 |
|---|---|
| 피갑류 | Megathericulus, Prepotherium, Prozaedyus, Paraeucinepeltus |
| 불요치목 | Protypotherium |
| 아스트라포테리아 | 아스트라포테리움 |
| 스파라소돈트 | 파타고스밀루스, 클라도식티스 |
| 유대류 | Abderites |
| 영장류 | Proteropithecia |
| 설치류 | Maruchito, Protacaremys, Neoreomys, Prolagostomus |
참조
[1]
웹사이트
Fossil found of a Big Bird Kermit wouldn't like
https://www.nytimes.[...]
2006-10-31
[2]
뉴스
Teen finds fossil skull of biggest bird known
http://news.google.c[...]
2006-10-26
[3]
간행물
A new phorusrhacid (Aves: Cariamae) from the middle Miocene of Patagonia, Argentina
https://www.research[...]
2007
[4]
서적
Living Dinosaurs
2011
[5]
간행물
A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds
https://www.research[...]
2015
[6]
간행물
A revision of skull morphology in Phorusrhacidae (Aves, Cariamiformes)
https://www.research[...]
2020
[7]
간행물
The youngest large carnassial bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene-Early Pleistocene of Uruguay)
https://www.jstor.or[...]
1999
[8]
간행물
Flights of fancy in avian evolution
2014
[9]
서적
South American and Antarctic Continental Cenozoic Birds
Springer
2013
[10]
간행물
Skull morphology of giant terror birds
https://www.research[...]
2006
[11]
간행물
Systematic revision of the Phorusrhacidae (Aves: Ralliformes)
2003
[12]
간행물
High speed galloping in the cheetah (''Acinonyx jubatus'') and the racing greyhound (''Canis familiaris''): spatio-temporal and kinetic characteristics
2012
[13]
간행물
Terror birds on the run: a mechanical model to estimate its maximum running speed
https://www.research[...]
2005
[14]
웹사이트
Huge "terror bird" fossil discovered in Patagonia
https://www.npr.org/[...]
2006-10-25
[15]
간행물
Mechanical analysis of feeding behavior in the extinct "terror bird" ''Andalgalornis steulleti'' (Gruiformes: Phorusrhacidae)
2010
[16]
간행물
Flexibility along the neck of the Neogene terror bird ''Andalgalornis steulleti'' (Aves Phorusrhacidae)
2012
[17]
서적
The Late Cenozoic of Patagonia and Tierra del Fuego
Elsevier Science
[18]
간행물
Middle Miocene climate transition as reflected by changes in ichnofacies and palaeosols from Patagonia, Argentina
2022
[19]
간행물
"Astrapotherium" from the Middle Miocene Collón Cura Formation and the Decline of Astrapotheres in Southern South America
2019
[20]
간행물
Evolutive implications of ''Megathericulus patagonicus'' (Xenarthra, Megatheriinae) from the Miocene of Patagonia Argentina
2020
[21]
간행물
A new pitheciin primate from the middle Miocene of Argentina.
https://www.research[...]
1998
[22]
간행물
''Cladosictis patagonica'' (Metatheria, Sparassodonta) from the Collón Cura Formation (Middle Miocene), Río Negro, Argentina
2021
[23]
웹사이트
Fossil found of a Big Bird Kermit wouldn't like
https://www.nytimes.[...]
2006-10-31
[24]
뉴스
Teen finds fossil skull of biggest bird known
http://news.google.c[...]
2006-10-26
[25]
간행물
A new phorusrhacid (Aves: Cariamae) from the middle Miocene of Patagonia, Argentina
https://www.research[...]
2007
[26]
서적
Living Dinosaurs
2011
[27]
간행물
A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds
https://www.research[...]
2015
[28]
간행물
A revision of skull morphology in Phorusrhacidae (Aves, Cariamiformes)
https://www.research[...]
2020
[29]
간행물
The youngest large carnassial bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene-Early Pleistocene of Uruguay)
https://www.jstor.or[...]
1999
[30]
간행물
Flights of fancy in avian evolution
2014
[31]
서적
South American and Antarctic Continental Cenozoic Birds
Springer
2013
[32]
간행물
Skull morphology of giant terror birds
https://www.research[...]
2006
[33]
간행물
Systematic revision of the Phorusrhacidae (Aves: Ralliformes)
2003
[34]
간행물
High speed galloping in the cheetah (''Acinonyx jubatus'') and the racing greyhound (''Canis familiaris''): spatio-temporal and kinetic characteristics
2012
[35]
간행물
Terror birds on the run: a mechanical model to estimate its maximum running speed
https://www.research[...]
2005
[36]
웹사이트
Huge "terror bird" fossil discovered in Patagonia
https://www.npr.org/[...]
2006-10-25
[37]
논문
Mechanical analysis of feeding behavior in the extinct "terror bird" ''Andalgalornis steulleti'' (Gruiformes: Phorusrhacidae)
2010
[38]
논문
Flexibility along the neck of the Neogene terror bird ''Andalgalornis steulleti'' (Aves Phorusrhacidae)
2012
[39]
서적
The Late Cenozoic of Patagonia and Tierra del Fuego
Elsevier Science
[40]
논문
Middle Miocene climate transition as reflected by changes in ichnofacies and palaeosols from Patagonia, Argentina
2022
[41]
논문
"Astrapotherium" from the Middle Miocene Collón Cura Formation and the Decline of Astrapotheres in Southern South America
2019
[42]
논문
Evolutive implications of ''Megathericulus patagonicus'' (Xenarthra, Megatheriinae) from the Miocene of Patagonia Argentina
2020
[43]
논문
A new pitheciin primate from the middle Miocene of Argentina.
https://www.research[...]
1998
[44]
논문
"Cladosictis patagonica" (Metatheria, Sparassodonta) from the Collón Cura Formation (Middle Miocene), Río Negro, Argentina
2021
[45]
논문
A new phorusrhacid (Aves: Cariamae) from the middle Miocene of Patagonia, Argentina.
2007
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com