고대고래아목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
고대고래아목은 멸종된 고래의 한 분류로, 고래하목에 속한다. 파키케투스과, 암불로케투스과, 레밍턴케투스과, 프로토케투스과, 바실로사우루스과를 포함하며, 화석 기록을 통해 초기 고래의 진화 과정을 보여준다. 이들은 육상 생활에서 수중 생활로 적응하는 과정을 거쳤으며, 콧구멍의 위치 변화, 사지의 퇴화 등의 특징을 나타낸다. 에오세와 올리고세 경계에서 대부분 멸종되었으며, 기후 변화와 경쟁, 생태적 지위 변화 등이 멸종에 영향을 미쳤다.
더 읽어볼만한 페이지
- 고대 고래 - 파키케투스
파키케투스는 5천만 년 전 에오세 초 파키스탄에 살았던 늑대 크기의 육상 포유류 고래류로, 육지 생활에 적응했으나 고래류 특유의 내이 구조와 치아 배열을 가져 고래 진화의 중요한 중간 단계이며, 고래가 육상 우제류에서 진화했음을 보여주는 중요한 증거이다. - 고대 고래 - 스콸로돈
스콸로돈은 멸종된 이빨고래의 속으로, 조상 고래와 현대 이빨고래의 특징을 모두 가지며, 원시적인 치열과 반향정위의 기원을 보이는 특징을 나타낸다. - 측계통군 - 맹금류
맹금류는 날카로운 발톱과 부리를 가진 사나운 새를 통칭하며, 생태계 최상위 포식자로서 뛰어난 시력과 비행 능력을 갖추고 있으며, 수리, 매, 독수리, 올빼미 등이 대표적이고, 서식지 파괴 등의 위협에 직면해 있다. - 측계통군 - 돌고래
돌고래는 이빨고래류에 속하는 해양 포유류의 총칭으로, 다양한 종이 존재하며 높은 지능과 사회성을 가지지만, 서식지 파괴, 환경 오염 등으로 멸종 위기에 처해 있다.
| 고대고래아목 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Archaeoceti |
| 명명자 | 플라워, 1883 |
| 화석 범위 | '' |
| 시대 | 에오세 초기 ~ 올리고세 후기 |
| 분류 | |
| 상위 분류 | 고래하목 |
| 하위 분류 | 아래 텍스트 참조 |
| 과 및 분기군 | 파키케투스과 암불로케투스과 레밍토노케투스과 프로토케투스과 바실로사우루스과 |
2. 하위 분류
- '''고래하목''' (Cetacea)
- * † '''고대고래소목''' (Archaeoceti)
- ** '''파키케투스과''' (Pakicetidae)
파키케투스속 (''Pakicetus'')
날라케투스속 (''Nalacetus'')
이크티올레스테스속 (''Ichthyolestes'')
- ** '''암불로케투스과''' (Ambulocetidae)
암불로케투스속 (''Ambulocetus'')
간다카시아속 (''Gandakasia'')
히말라야케투스속 (''Himalayacetus'')
- ** '''레밍톤케투스과''' (Remingtonocetidae)
앤드류시피우스속 (''Andrewsiphius'')
아토키케투스속 (''Attockicetus'')
달라니스테스속 (''Dalanistes'')
쿠트키케투스속 (''Kutchicetus'')
레밍토노케투스속 (''Remingtonocetus'')
- ** '''프로토케투스과''' (Protocetids)
조지아케투스아과 (Georgiacetinae)
* 바비아케투스속 (''Babiacetus'')
* 카롤리나케투스속 (''Carolinacetus'')
* 에오케투스속 (''Eocetus'')
* 조지아케투스속 (''Georgiacetus'')
* 낫치토치아속 (''Natchitochia'')
* 팝포케투스속 (''Pappocetus'')
마카라케투스아과 (Makaracetinae)
* 마카라케투스속 (''Makaracetus'')
프로토케투스아과 (Protocetinae)
* 아르티오케투스속 (''Artiocetus'')[22]
* 크레나토케투스속 (''Crenatocetus'')
* 가비아케투스속 (''Gaviacetus'')
* 인도케투스속 (''Indocetus'')
* 마이아케투스속 (''Maiacetus'')
* 프로토케투스속 (''Protocetus'')
* 콰이스라케투스속 (''Qaisracetus'')
* 로드호케투스속 (''Rodhocetus'')
* 타크라케투스속 (''Takracetus'')
- ** '''바실로사우루스과''' (Basilosauridae)
바실로사우루스아과(Basilosaurinae)
* 바실로사우루스속 (''Basilosaurus'')
* 바실로테루스속 (''Basiloterus'')
도루돈아과 (Dorudontinae)
* 안카레케투스속 (''Ancalecetus '')
* 크리소케투스속 (''Chrysocetus'')
* 킨시아케투스속 (''Cynthiacetus'')
* 도루돈속 (''Dorudon'')
* 사가케투스속 (''Saghacetus'')
* 지고르히자속 (''Zygorhiza'')
케케노돈아과 (Kekenodontinae)
* 케케노돈속 (''Kekenodon'')
* 포코케투스속 (''Phococetus'')
스트로메리우스아과 (Stromeriinae)
* 스트로메리우스속 (''Stromerius'')[23]
케케노돈과 (Kekenodontinae)는 고대고래아목에 속하는지, 수염고래아목[14] 또는 참돌고래상과 (Delphinoidea)에 속하는지에 대한 논쟁이 있다.[15]
2. 1. 파키케투스과 (Pakicetidae)
Pakicetidae영어는 고대고래아목에 속하는 멸종된 과이다. 파키케투스과는 West에 의해 처음 고래로 확인되었으며,[7] 가장 오래된 고래로 알려져 있다. 이들은 길고 가느다란 다리와 길고 좁은 꼬리를 가지고 있었으며, 현대 늑대 크기까지 자랄 수 있었다. 인도 북서부와 파키스탄 북부의 담수 하천 퇴적물에서만 발견되었으며, 수영보다는 물속을 걸어 다니는 동물이었을 것으로 추정된다.[7]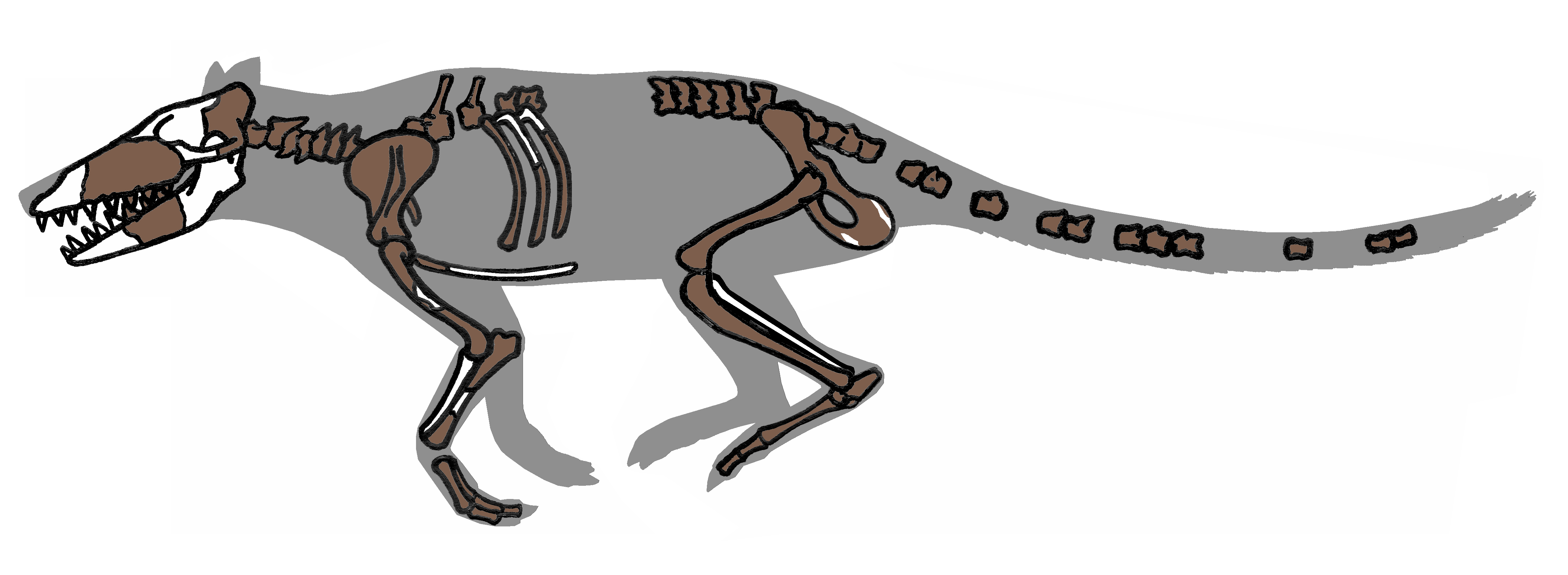
파키케투스과는 파키케투스(''Pakicetus''), 날라케투스(''Nalacetus''), 이크티올레스테스(''Ichthyolestes'')의 세 속(屬)으로 구성된다.[21]
수십 개의 화석이 알려져 있지만 두개골, 이빨, 턱 조각뿐이며, 완전한 골격은 발견되지 않았다. 치아의 형태는 다양했는데, 가장 작은 종은 현대의 물고기 포식자와 같은 이빨을 가졌고, 가장 큰 종은 현대의 하이에나와 더 비슷했다. 파키케투스과는 포식자이거나 썩은 고기를 먹는 동물이었을 수 있다. 파키케투스과의 두개골과 치아는 현대 고래와 닮은 점이 없지만, S자 돌기, 이중골, 골질 비대(조밀한) 회전된 작은뼈는 여전히 고래류의 특징을 보여준다.[2]

- 파키케투스 : 약 5,300만 년 전에 서식했으며, 몸길이는 약 1.8m로 추정된다. 파키스탄에서 발견되었다. 긴 꼬리를 가지고 있으며, 육상 포유류로서의 사지와 발굽 형태의 발을 가지고 있다. 내이골의 전체적인 형태는 고래와 유사하며, 뼈 등의 두께로 보아 수중에서의 골전도에 의한 청각을 가졌을 것으로 생각된다. 또한 내이골의 일부인 등골이 우제목의 특징을 가지고 있다는 점에서, 초기 고래우제목에서 분기되어 고래가 되는 과정의 생물로 여겨진다.
2. 2. 암불로케투스과 (Ambulocetidae)
암불로케투스과는 크고, 이미 완전한 수생 생물이었으며,[8] 큰 발과 튼튼한 꼬리를 가진 악어와 같은 모습을 하고 있었다. 퇴적물은 이들이 해안 지역에 살았으며, 그들의 골경화증(뼈 경화)는 이들이 빠른 추격자라기보다는 매복 포식자였음을 시사한다. 파키스탄과 인도에서만 알려진 암불로케투스과에는 가장 오래된 고래로 여겨지는 ''히말라야케투스''가 포함되어 있으며, 이는 다른 암불로케투스과 구성원들보다 약 400만 년 더 오래된 것이다.[7]10개 미만의 화석이 설명되었는데, 그 중 하나인 ''암불로케투스 나탄스''는 거의 완전하며 초기 고래류 진화에 대한 주요 정보원이다. 수컷 바다사자 크기였으며, 길고 주둥이가 길고 튼튼하며 심하게 닳은 이빨을 가진 큰 머리를 가지고 있었다. 아래턱은 ''암불로케투스''가 턱 뒤쪽과 중이(中耳)를 연결하는 특이한 연조직을 가지고 있었음을 보여주는데, 이는 현대 이빨고래류의 큰 소리 수신 지방 패드와 작은 유사성을 가진다. 눈은 머리 윗쪽에 위치했지만 옆을 향하고 있었다. 머리, 목, 등의 근육은 강했고, 지느러미가 없는 꼬리는 길었다. 뒷다리는 짧았지만 긴 발이 있었다. 앞다리 역시 짧았고 5개의 짧은 발굽이 있었다. ''암불로케투스''는 현대 수달처럼 뒷발로 헤엄쳤을 것이며, 육지에서 자체적인 무게를 지탱할 수 없었다. 그것은 아마도 현대 악어처럼 매복 사냥꾼이었을 것이다.[2]
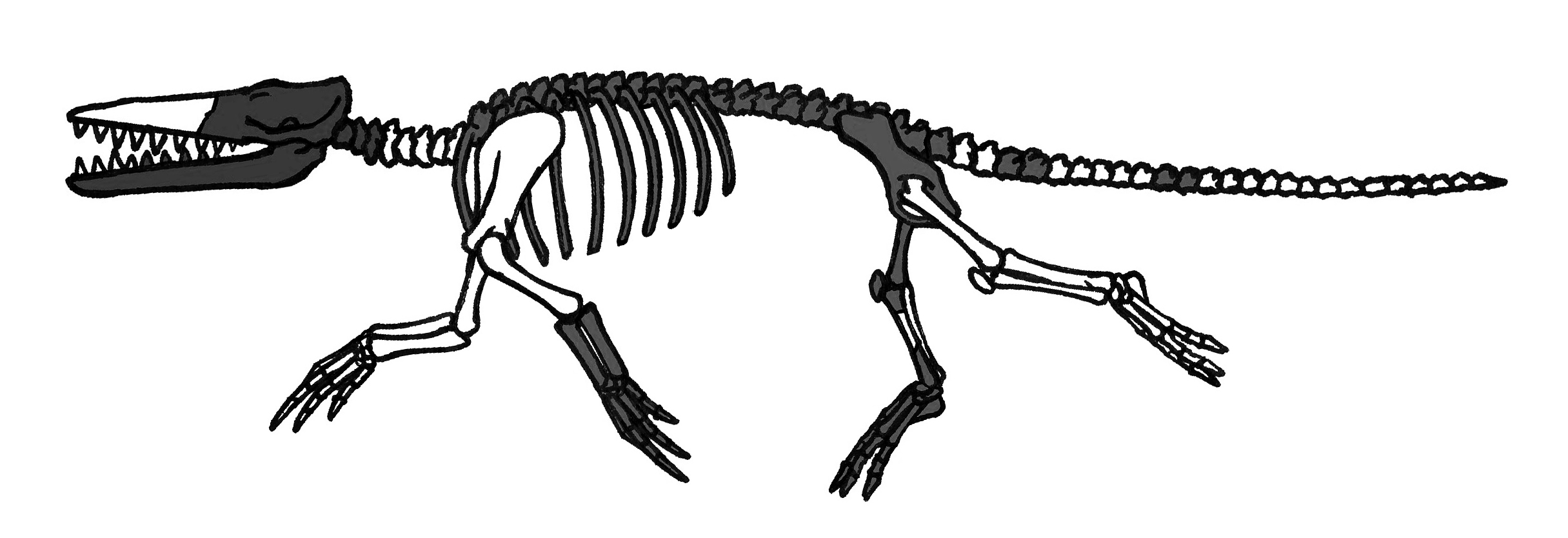

- 암불로케투스 : 약 5,000만 ~ 4,900만 년 전에 서식했으며, 몸길이는 약 3m로 추정된다. 암불로케투스과에 속하며, 인도·파키스탄 지방에서 발견되었다. 학명은 '헤엄쳐 걷는 고래'라는 뜻이다. 대표적인 종으로는 암불로케투스 나탄스가 있다. 긴 꼬리를 가지고 있으며, 사지도 있지만 사지와 전체 골격의 비율은 파키케투스보다 작고 짧다. 또한 뒷발에는 물갈퀴가 있었을 것으로 생각된다. 튼튼한 골격을 가지고 있으며, 복원 상상도는 하마와 악어의 중간과 같다. 머리와 턱의 골격이 크게 발달했으며, 이빨도 육식에 적합한 형태를 띠고 있어, 물가에서 악어와 같은 포식 활동을 했을 것으로 여겨진다.
2. 3. 레밍턴케투스과 (Remingtonocetidae)
레밍턴케투스과(Remingtonocetidae)는 짧은 사지와 납작한 척추뼈를 가진 강력하고 힘있는 꼬리를 가지고 있었다. 긴 주둥이, 작은 눈, 그리고 귀의 형태는 이들의 시력이 좋지 않았고 청각이 지배적인 감각이었음을 시사한다. 이들은 파키스탄과 인도에서만 발견되었으며, 퇴적물은 이들이 해안 지역의 탁한 물에서 살았음을 알려준다. 레밍턴케투스과는 육지에서도 살 수 있었을 것이지만, 꼬리를 사용하여 수영했을 것으로 보인다.[7]수십 개의 화석이 묘사되었지만, 대부분은 치아 및 두개골 이외의 골격 잔해는 거의 없이 두개골과 하악골뿐이다. 레밍턴고래과는 크기가 다양했을 것이며, 가장 작은 종은 파키케투스(Pakicetus)와 비슷하고 가장 큰 종은 암불로케투스(Ambulocetus)와 비슷했을 것이다. 레밍턴고래과는 다른 고대 고래류보다 더 긴 주둥이를 가지고 있었지만, 두개골 형태 역시 상당히 달랐으며, 이는 아마도 이들이 다른 먹이를 먹었음을 보여주는 것으로 보인다. 눈은 작았지만 귀는 크고 멀리 떨어져 있었는데, 이는 수중 청각에 대한 강조가 증가했음을 반영하는 것으로 보인다. 레밍턴고래과 두개골 이외 골격의 단편적인 잔해는 이들이 긴 목과 육상에서 체중을 지탱할 수 있었을 큰 뒷다리를 가지고 있었음을 시사한다.[2]
레밍턴케투스과에는 다음 속들이 포함된다.
- 앤드루시피우스(Andrewsiphius)
- 아토키케투스(Attockicetus)
- 달라니스테스(Dalanistes)
- 쿠트키케투스(Kutchicetus)
- 레밍토노케투스(Remingtonocetus)
- ''라야니스테스''(Rayanistes)

2. 4. 프로토케투스과 (Protocetidae)

원시고래과(Protocetidae)는 아프리카와 아메리카에서 발견되는, 뒷다리와 강력한 꼬리를 가진 다양한 과이다. 얕고 따뜻한 바다(예: 산호초)에 서식했으며, 헤엄을 잘 쳤음을 알 수 있다. 이들은 지구의 바다로 퍼져 나가면서 고래류의 진화에 큰 영향을 미쳤다.[7] 긴 주둥이와 큰 눈을 가지고 있었고, 코 구멍은 초기 고대 고래보다 머리 위쪽에 위치해 있었다. 이는 현대 고래와 유사하게 머리를 수평으로 유지하며 숨을 쉴 수 있었음을 시사하며, 호흡공으로 가는 첫 단계였다. 치열은 다양했지만, 현대 고래의 저작하지 않는 이빨로 진화하기 시작했으며, 아마도 활동적인 사냥꾼이었을 것이다.
육상에서 이동하는 능력은 다양했던 것으로 보인다. ''로드호케투스''(Rodhocetus)와 ''페레고케투스''(Peregocetus)는 천장관절을 가지고 있어 육상에서 이동할 수 있었음을 나타낸다.[10] 반면 ''조지아케투스''(Georgiacetus)와 ''에기케투스''(Aegicetus)는 골반이 척추와 연결되지 않아 뒷다리가 몸무게를 지탱할 수 없었을 것으로 보인다.[2] 일부 속(''로드호케투스'')은 큰 물갈퀴를 형성하는 큰 뒷발을 가지고 있었고, ''에기케투스''는 물속에서 추진력을 얻기 위해 꼬리에 더 의존했던 것으로 보인다.[11]
프로토케투스과는 아래와 같이 3개의 아과로 나뉜다.[21]
| 아과 | 속 |
|---|---|
| 조지아케투스아과 (Georgiacetinae) | |
| 마카라케투스아과 (Makaracetinae) | |
| 프로토케투스아과 (Protocetinae) |
2. 5. 바실로사우루스과 (Basilosauridae)
바실로사우루스과는 뒷다리가 작고 지느러미 모양의 앞다리를 가지고 있었으며, 필연적으로 수생 생활을 했고 바다를 지배하게 되었다. 그들은 여전히 현대의 이빨고래와 수염고래가 가진 반향정위와 수염을 갖추지 못했다.[7] 바실로사우루스과와 도루돈과는 전체 골격이 알려진 가장 오래된 완전 수생 고래류이다. 이들은 이전의 고대고래류에는 없는 많은 수생 적응을 보여준다:[12] 척주에서 목뼈는 짧고, 가슴뼈와 허리뼈는 길이가 비슷하며, 천추는 융합되지 않았고, 천장관절은 없으며, 짧은 꼬리에는 구상 척추가 있다(꼬리지느러미의 존재를 나타냄). 어깨뼈는 넓고 부채꼴 모양이며 앞쪽의 견봉과 작은 극상와를 가지고 있다. 척골은 크고 가로로 평평한 주두를 가지고 있으며, 손목과 원위 전완은 손의 평면에 평평하고 뒷다리는 작다.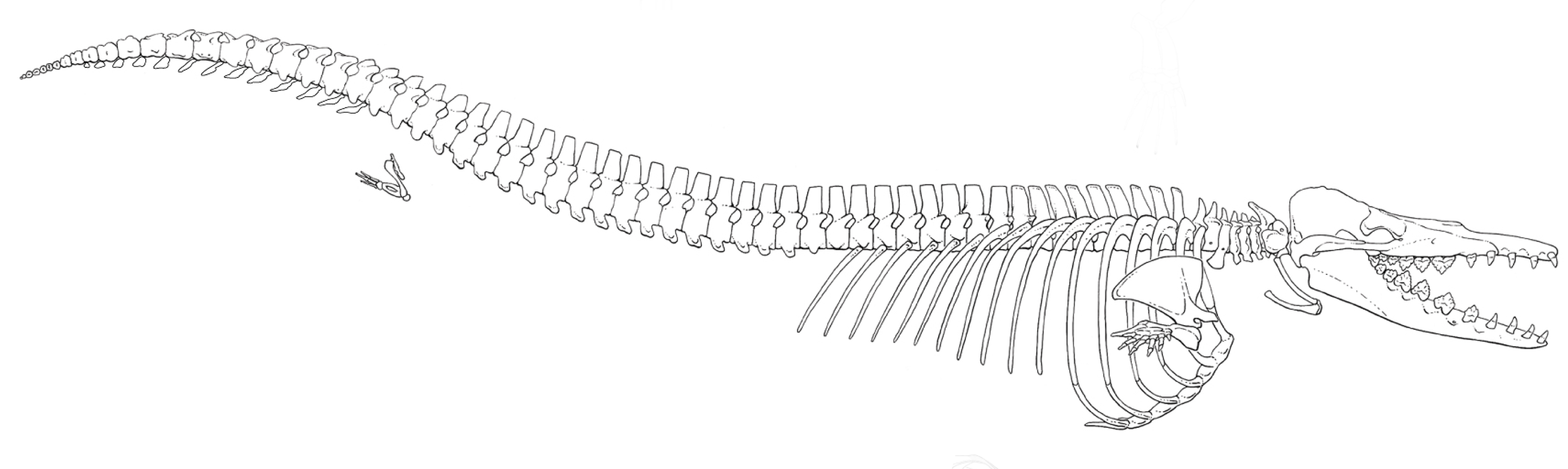
- 바실로사우루스아과(Basilosaurinae)
- * 바실로사우루스속 (''Basilosaurus'')
- * 바실로테루스속 (''Basiloterus'')
- 도루돈아과 (Dorudontinae)
- * 안카레케투스속 (''Ancalecetus '')
- * 크리소케투스속 (''Chrysocetus'')
- * 킨시아케투스속 (''Cynthiacetus'')
- * 도루돈속 (''Dorudon'')
- * 사가케투스속 (''Saghacetus'')
- * 지고르히자속 (''Zygorhiza'')
- 케케노돈아과 (Kekenodontinae)
- * 케케노돈속 (''Kekenodon'')
- * 포코케투스속 (''Phococetus'')
- 스트로메리우스아과 (Stromeriinae)
- * 스트로메리우스속 (''Stromerius'')[23]

- 바실로사우루스 : 약 4,000만 ~ 3,400만 년 전에 서식했으며, 몸길이는 약 20m로 추정된다. 이집트에서 발견되었다. 고래목으로 분류되기 전에는 어룡목으로 분류되어 바실로사우루스라고 명명되었지만, 분류 변경에 따라 '''제우글로돈'''으로 변경하려는 시도가 있었다. 앞발은 지느러미가 되었고, 뒷발은 거의 퇴화하여 3개의 손가락만 남아있다. 꼬리가 매우 길지만, 골격만으로는 지느러미 모양의 부분이 있었는지는 알 수 없다. 하지만, 학술적인 추론에 따라 복원 상상도에서는 다양한 지느러미가 꼬리에 그려져 있다. 머리에는 콧구멍이 2개 있으며, 텔레스코핑 현상은 나타나지 않는다. 또한 머리는 튼튼하고 가늘며 상하 턱에는 육식에 적합한 이빨이 다수 있어, 이 점 등으로 어룡으로 분류된 하나의 요인으로 여겨진다.
3. 계통 발생
경우제류(Cetartiodactyla)에서 갈라져 나온 고대고래아목은 분자계통학과 형태학적 연구를 통해 우제류의 하마과와 자매 관계에 있다는 견해가 지배적이다. 고대고래류의 조상은 백악기 후기 또는 시생세 초기에 우제류에서 분화되었으며, 낙타나 멧돼지로 진화하는 계통보다는 늦게, 반추류로 이어지는 계통보다는 빠르게 분기했다. 이후 고래의 조상은 하마 계통과도 분리되어 해양 포유류로 진화했다. 백악기 말기의 대량 절멸로 생긴 해양 생태계의 공백을 메우는 형태로 대형 해양 동물로 적응해 나갔다.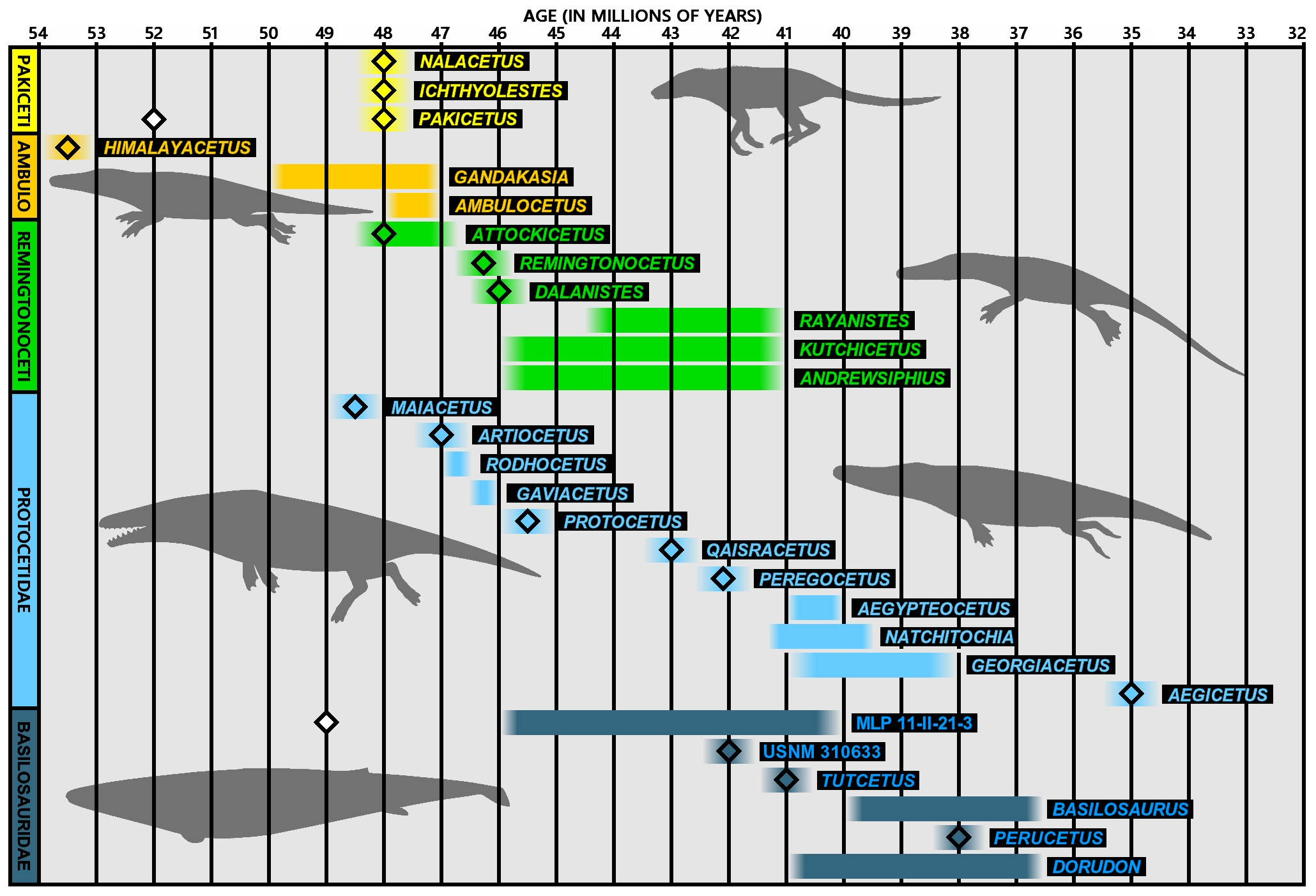
대부분의 고대고래류는 뒷다리를 가지고 있어 현생 고래류와 다르다. 초기에는 튼튼한 사지를 가진 육상 동물이었으며, 가장 오래된 고래로 여겨지는 파키케투스가 이에 해당한다. 시신세에 고대고래류는 테티스 해를 중심으로 번성하며 빠르게 진화했다. 사지는 지느러미로 바뀌고, 육상에 적합한 반고리관은 퇴화하고 해양에 적합한 골전도 구조를 가진 고래류 특유의 이골이 발달했다. 이러한 빠른 진화 과정을 통해, 바실로사우루스 단계에 이르러서는 육상에서 해양이라는 전혀 다른 환경에 대한 적응을 완료했다.
아래는 고대고래의 계통 발생을 나타낸 분기도이다.








4. 진화
고대고래아목의 진화는 경우제류(Cetartiodactyla)에서 시작되었다. 초기에는 우제류(Artiodactyla)에 속했으며, 이후 고래하목(Cetacea)으로 분화되었다. 고래하목은 다시 파키케투스과(Pakicetidae), 암불로케투스과(Ambulocetidae), 레밍톤고래과(Remingtonocetidae), 프로토케투스과(Protocetidae), 바실로사우루스아과(Basilosaurinae), 도루돈아과(Dorudontinae)를 거쳐 이빨고래(Odontoceti)와 수염고래(Mysticeti)로 진화했다.
과거에는 고대고래류가 메소니키아목에서 진화했다고 생각되었으나, 최근에는 분자계통학과 형태학적 연구를 통해 우제목의 하마과와 자매 관계에 있다는 견해가 지배적이다. 고대고래류의 조상은 백악기 후기 또는 팔레오세 초기에 우제류에서 분화되었으며, 낙타나 멧돼지보다는 늦게, 반추류보다는 빠른 시기에 분기했다. 이후 고래의 조상은 하마 계통과 갈라져 해양 포유류로 진화했다. K-Pg 멸종 이후, 고래의 조상은 해양 생태계의 공백을 메우며 대형 해양 동물로 적응해 나갔다.
대부분의 고대고래류는 뒷다리를 가지고 있었지만, 에오세를 거치며 테티스 해를 중심으로 번성하고 빠르게 진화했다. 이 과정에서 사지는 지느러미로 바뀌고, 육상 생활에 적합했던 반고리관은 퇴화하고 골전도 구조를 가진 이골이 발달하는 등 해양 생활에 적합한 형태로 변화했다. 이러한 진화는 매우 빠르게 진행되어, 바실로사우루스 단계에 이르러서는 초기 고래류와는 현저하게 다른 수준의 적응을 달성했다.
4. 1. 육상 생활에서 수중 생활로의 적응
파키케투스과는 가장 오래된 고래로, 길고 가느다란 다리와 좁은 꼬리를 가지고 있었으며, 현대 늑대 크기까지 자랄 수 있었다. 이들은 인도 북서부와 파키스탄 북부의 담수 하천 퇴적물에서만 발견되었으며, 수영보다는 물속을 걸어 다니는 동물이었을 것이다.[7]수십 개의 화석이 알려져 있지만 두개골, 이빨, 턱 조각뿐이며, 완전한 골격은 발견되지 않았다. 치아의 형태는 다양했는데, 가장 작은 종은 현대의 물고기 포식자와 같은 이빨을 가졌고, 가장 큰 종은 현대의 하이에나와 더 비슷했다. 파키케투스과는 포식자이거나 썩은 고기를 먹는 동물이었을 수 있다. 파키케투스과의 두개골과 치아는 현대 고래와 닮은 점이 없지만, S자 돌기, 이중골, 골질 비대(조밀한) 회전된 작은뼈는 여전히 고래류의 특징을 보여준다.[2]
고래의 다음 분기 과인 암불로케투스과는 크고, 이미 완전한 수생 생물이었으며,[8] 큰 발과 튼튼한 꼬리를 가진 악어와 같은 모습을 하고 있었다. 퇴적물은 이들이 해안 지역에 살았으며, 그들의 골경화증(뼈 경화)는 이들이 빠른 추격자라기보다는 매복 포식자였음을 시사한다. 파키스탄과 인도에서만 알려진 암불로케투스과에는 가장 오래된 고래로 여겨지는 ''히말라야케투스''가 포함되어 있으며, 이는 다른 암불로케투스과 구성원들보다 약 400만 년 더 오래된 것이다.[7]
10개 미만의 화석이 설명되었는데, 그 중 하나인 ''Ambulocetus natans|암불로케투스 나탄스''는 거의 완전하며 초기 고래류 진화에 대한 주요 정보원이다. 수컷 바다사자 크기였으며, 길고 주둥이가 길고 튼튼하며 심하게 닳은 이빨을 가진 큰 머리를 가지고 있었다. 아래턱은 ''암불로케투스''가 턱 뒤쪽과 중이(中耳)를 연결하는 특이한 연조직을 가지고 있었음을 보여주는데, 이는 현대 이빨고래류의 큰 소리 수신 지방 패드와 작은 유사성을 가진다. 눈은 머리 윗쪽에 위치했지만 옆을 향하고 있었다. 머리, 목, 등의 근육은 강했고, 지느러미가 없는 꼬리는 길었다. 뒷다리는 짧았지만 긴 발이 있었다. 앞다리 역시 짧았고 5개의 짧은 발굽이 있었다. ''암불로케투스''는 현대 수달처럼 뒷발로 헤엄쳤을 것이며, 육지에서 자체적인 무게를 지탱할 수 없었다. 그것은 아마도 현대 악어처럼 매복 사냥꾼이었을 것이다.[2]
레밍턴고래과(Remingtonocetidae)는 짧은 사지와 납작한 척추뼈를 가진 강력하고 힘있는 꼬리를 가지고 있었다. 긴 주둥이, 작은 눈, 귀의 형태는 시력이 좋지 않았고 청각이 지배적인 감각이었음을 시사한다. 이들 또한 파키스탄과 인도에서만 발견되었으며, 퇴적물은 이들이 해안 지역의 탁한 물에서 살았음을 시사한다. 육지에서도 살 수 있었을 것이지만, 꼬리를 사용하여 수영했을 것으로 보인다.[7]
수십 개의 화석이 묘사되었지만, 대부분은 치아 및 두개골 이외의 골격 잔해는 거의 없이 두개골과 하악골뿐이다. 레밍턴고래과는 크기가 다양했을 것이며, 가장 작은 종은 ''파키케투스''(Pakicetus)와 비슷하고 가장 큰 종은 ''암불로케투스''(Ambulocetus)와 비슷했을 것이다. 레밍턴고래과는 다른 고대 고래류보다 더 긴 주둥이를 가지고 있었지만, 두개골 형태 역시 상당히 달랐으며, 이는 아마도 다른 식단을 반영하는 것으로 보인다. 눈은 작았지만 귀는 크고 멀리 떨어져 있었는데, 이는 수중 청각에 대한 강조가 증가했음을 반영하는 것으로 보인다. 레밍턴고래과 두개골 이외 골격의 단편적인 잔해는 긴 목과 육상에서 체중을 지탱할 수 있었을 큰 뒷다리를 가지고 있었음을 시사한다.[2]
나머지 과와 후기 왕관 고래류는 여섯 개의 파생형질(Synapomorphy)에 의해 묶인 분지군을 형성한다:[9] 바깥 콧구멍의 앞쪽 가장자리가 세 번째 위쪽 앞니 위 또는 뒤에 위치하고, 코는 넓으며, 안와상 돌기는 존재하지만 짧고, 안와의 앞쪽 가장자리는 두 번째 또는 세 번째 위쪽 어금니 위에 위치하고, 후안와 돌기는 시상 능선과 90° 각도를 이루며, 목뼈는 짧다.
아프리카와 아메리카에서 발견되는 원시고래과(Protocetidae)는 뒷다리와 강력한 꼬리를 가진 다양한 과로, 얕고 따뜻한 바다(예: 산호초)에 서식하며 헤엄을 잘 쳤음을 나타낸다. 이들은 지구의 바다로 퍼져 나가면서 고래류의 진화에 큰 영향을 미쳤다.[7] 그들은 긴 주둥이, 큰 눈을 가지고 있었고, 코 구멍은 초기 고대 고래보다 머리 위쪽에 위치해 있었는데, 이는 현대 고래와 유사하게 머리를 수평으로 유지하며 숨을 쉴 수 있었음을 시사하며, 이는 호흡공으로 가는 첫 단계였다. 그들의 치열은 다양했지만, 현대 고래의 저작하지 않는 이빨로 진화하기 시작했으며, 아마도 활동적인 사냥꾼이었을 것이다. 육상에서 이동하는 능력은 다양했던 것으로 보이는데, ''로도케투스''(Rodhocetus)와 ''페레고케투스''(Peregocetus)는 천장관절을 가지고 있어 육상에서 이동할 수 있었음을 나타낸다.[10] 다른 속(''조지아케투스''(Georgiacetus)와 ''에기케투스''(Aegicetus))에서는 골반이 척추와 연결되지 않아 뒷다리가 몸무게를 지탱할 수 없었을 것으로 보인다.[2] 일부 속(''로도케투스'')은 큰 물갈퀴를 형성하는 큰 뒷발을 가지고 있었고, 반면 ''에기케투스''는 물속에서 추진력을 얻기 위해 꼬리에 더 의존했던 것으로 보인다.[11]
바실로사우루스과는 뒷다리가 작고 지느러미 모양의 앞다리를 가지고 있었으며, 필연적으로 수생 생활을 했고 바다를 지배하게 되었다. 그들은 여전히 현대의 이빨고래와 수염고래가 가진 반향정위와 수염을 갖추지 못했다.[7] 바실로사우루스과와 도루돈과는 전체 골격이 알려진 가장 오래된 완전 수생 고래류이다. 이들은 이전의 고대고래류에는 없는 많은 수생 적응을 보여준다:[12] 척주에서 목뼈는 짧고, 가슴뼈와 허리뼈는 길이가 비슷하며, 천추는 융합되지 않았고, 천장관절은 없으며, 짧은 꼬리에는 구상 척추가 있다(꼬리지느러미의 존재를 나타냄). 어깨뼈는 넓고 부채꼴 모양이며 앞쪽의 견봉과 작은 극상와를 가지고 있다. 척골은 크고 가로로 평평한 주두를 가지고 있으며, 손목과 원위 전완은 손의 평면에 평평하고 뒷다리는 작다.
대부분의 고대고래류는 뒷다리를 갖추고 있으며, 현생 고래류와는 확연히 다르다. 시원적인 종은 튼튼한 사지를 갖춘 완전한 육상 동물이었다고 생각되며, 현재 가장 그것과 가깝다고 여겨지는 것은 가장 오래된 고래라고 여겨지는 사지를 가진 동물인 파키케투스이다. 시신세를 맞아, 고대고래류는 따뜻하고 광대한 얕은 바다인 테티스 해를 중심으로 크게 번성하고, 다양하고 빠르게 진화해 갔다고 보인다. 사지는 지느러미로 바뀌고, 육상에 적합한 반고리관은 퇴화하여 해양에 적합한 골전도 구조를 가진 고래류 특유의 이골 등이 그 자리를 대신한다. 이 진화의 흐름은 매우 빨라서, 같은 세 후기 초두에는 초기 고래류와는 현저하게 다른 수준의 적응을 달성하여 장대한 체구를 가진 바실로사우루스 단계에까지 도달했다. 즉, 불과 800만 년 정도의 짧은 기간에 고래류는 육상에서 해양이라는 전혀 다른 환경에 대한 적응 프로세스를 기본적으로 완료했던 것이다.
4. 2. 콧구멍의 이동
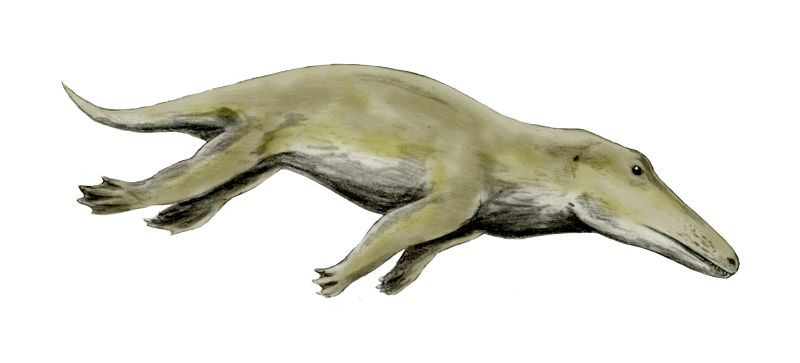
- -|]]|thumb|upright=1.5|마이아케투스(Maiacetus), 프로토케투스과(Protocetidae)]]
원시고래과(Protocetidae)는 아프리카와 아메리카에서 발견되며, 뒷다리와 강력한 꼬리를 가진 다양한 고대 고래 무리였다. 이들은 얕고 따뜻한 바다, 예를 들어 산호초와 같은 곳에 서식하며 헤엄을 잘 쳤다. 이들은 지구의 바다로 퍼져 나가면서 고래류의 진화에 큰 영향을 미쳤다.[7]
원시고래류는 긴 주둥이와 큰 눈을 가지고 있었고, 코 구멍은 초기 고대 고래보다 머리 위쪽에 위치해 있었다. 이는 현대 고래와 유사하게 머리를 수평으로 유지하며 숨을 쉴 수 있었음을 시사하며, 이는 호흡공으로 가는 첫 단계였다. 이들의 치열은 다양했지만, 현대 고래처럼 먹이를 씹지 않고 삼키는 형태로 진화하기 시작했으며, 아마도 활동적인 사냥꾼이었을 것이다.
육상에서 이동하는 능력은 다양했던 것으로 보인다. ''로도케투스''(Rodhocetus)와 ''페레고케투스''(Peregocetus)는 천장관절을 가지고 있어 육상에서 이동할 수 있었음을 나타낸다.[10] 반면, ''조지아케투스''(Georgiacetus)와 ''에기케투스''(Aegicetus)에서는 골반이 척추와 연결되지 않아 뒷다리가 몸무게를 지탱할 수 없었을 것으로 보인다.[2] 일부 속(''로도케투스'')은 큰 물갈퀴를 형성하는 큰 뒷발을 가지고 있었고, ''에기케투스''는 물속에서 추진력을 얻기 위해 꼬리에 더 의존했던 것으로 보인다.[11]
5. 멸종
고대 고래류는 에오세와 올리고세를 가르는 멸종기를 넘지 못하고, 아마도 바실로사우루스과 중 도루돈아과를 유일한 예외로 하여 다른 종들은 모두 사라졌다. 바실로사우루스과 또한 다음 시대에는 볼 수 없었고 멸종을 면하지 못했지만, 진정수염고래류라는 자손을 남기고 사라졌다.[20] 고대 고래류의 멸종은 고도로 진화한 진정수염고래류의 출현에 따른 도태압도 있지만, 에오세 말에 일어난 기후 변동에 의한 해수 온도의 저하나 해퇴(에오세 종말 사건), 이에 따른 생물량의 감쇠가 크게 영향을 미친 것으로 보이며, 어쩌면 그 둘 모두가 관련되어 있다고도 한다.
고대 고래류가 멸종한 올리고세의 생태계에서는 얕은 바다의 중형 해양 포식 동물의 니치 (생태적 지위)가 "공석"이 되었지만, 곰과 가까운 수륙 양서적 경향이 강한 육상 고양이목 식육류인 헤미키온과(en)의 일부가 이를 메우기 위해 진화를 시작하여 분기했고, 이어지는 중신세까지에는 본격적인 적응을 이루어 기각류(물범이나 바다사자의 무리)라는 동물군을 확고하게 자리 잡았다. 그들은 특히 원시적인 고대 고래류가 얻었던 수륙 양서의 중형 포식 동물로서의 지위를 차지하게 되었다. 또한, 후에 외해에서 진화한 돌고래 (소형 고래류)가 분포를 넓혀가는 가운데 얕은 바다나 담수역까지 진출함으로써, 고대 고래류의 멸종으로 인해 고래류의 손에서 벗어난 형태가 된 "얕은 바다의 중형 해양 포식 동물의 니치"는 대폭 되찾아졌다.
고대 고래류의 콧구멍 (분기공)은 파키케투스과에서는 머리 앞쪽에 위치해 있던 것이 차츰 뒤쪽으로 이동하여, 최말기의 종 중 하나인 도루돈[20]에서는 주둥이의 중간 위치에 이르렀다. 그러나 수면에서의 호흡을 용이하게 하는 머리 꼭대기로의 콧구멍의 완전한 이동은 진정수염고래류의 등장 이후에 일어났다. 더 나아가 해양으로의 적응 진화이다.
참조
[1]
웹사이트
Archaeoceti
https://paleodb.org/[...]
Paleobiology Database
2018-09-20
[2]
문헌
[3]
문헌
[4]
문헌
[5]
문헌
[6]
문헌
[7]
문헌
[8]
학술지
Farewell to life on land - thoracic strength as a new indicator to determine paleoecology in secondary aquatic mammals
[9]
문헌
[10]
간행물
An amphibious whale from the Middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans
[11]
간행물
Aegicetus gehennae, a new late Eocene protocetid (Cetacea, Archaeoceti) from Wadi Al Hitan, Egypt, and the transition to tail-powered swimming in whales
https://journals.plo[...]
[12]
문헌
[13]
문헌
[14]
문헌
[15]
문헌
[16]
학술기사
哺乳類の高次分類群および分類階級の日本語名称の提案について
https://doi.org/10.1[...]
日本哺乳類学会
[17]
학술기사
いくつかの日本産鯨類化石の再検討 ―起源の時期と古生物地理の観点から―
https://dl.ndl.go.jp[...]
福井県立恐竜博物館
[18]
문서
[19]
문서
[20]
문서
[21]
서적
The Beginning of the Age of Mammals
http://books.google.[...]
JHU Press
[22]
학술기사
Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan
http://adsabs.harvar[...]
[23]
저널
Stromerius nidensis, new archaeocete (Mammalia, Cetacea) from the Upper Eocene Qasr El-Sagha Formation, Fayum, Egypt
http://deepblue.lib.[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com