대롱편모조식물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
대롱편모조식물은 진핵생물의 일종으로, 세포 구조, 편모, 엽록체의 특징을 가지며, 광합성을 통해 생존한다. 이들은 단세포, 군체, 공통세포성 또는 다세포 생물일 수 있으며, 다양한 환경에서 발견된다. 대롱편모조식물은 크게 카키스타류와 파에이스타류로 분류되며, 규조류, 갈조류, 황조류 등을 포함한다. 또한, 크리시스타와 디아토미스타로 세분화되기도 한다. 황조식물은 스트라메노필라 내의 다양한 분류군을 포함하며, 홍조류와의 2차 공생을 통해 엽록체를 얻었다.
더 읽어볼만한 페이지
| 대롱편모조식물 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
| 학명 | Ochrophytes |
| 이명 | Heterokontophyta Ochrista Stramenochromes |
| 학문적 권위 | Cavalier-Smith 1986 emend. 1996 |
| 모식속 | Fucus |
| 모식속 권위 | Linnaeus, 1753 |
| 화석 범위 | 중기 원생누대 (10억 년 전) ~ 현재 |
| 추정 종 다양성 | 100,000종 이상 |
| 기재된 종 다양성 | 23,314종 |
 | |
| 분류 | |
| 상문 | SAR 슈퍼그룹 |
| 아계 | 스트라메노파일 |
| 하위 분류군 | 크리시스타 (Chrysista) 다이아토미스타 (Diatomista) |
| 크리시스타 하위 분류군 | 크리소피세아에 (Chrysophyceae) 에우스티그마토피세아에 (Eustigmatophyceae) 피코파게아 (Picophagea) 라피도피세아에 (Raphidophyceae) 아우레아레노피세아에 (Aurearenophyceae) 페오탐니오피세아에 (Phaeothamniophyceae) 크리소메로피세아에 (Chrysomerophyceae) 크리소파라독소피세아에 (Chrysoparadoxophyceae) 페오사키오피세아에 (Phaeosacciophyceae) 페오피세아에 (Phaeophyceae) 스키조클라디오피세아에 (Schizocladiophyceae) 크산토피세아에 (Xanthophyceae) 올리스토디스코피세아에 (Olisthodiscophyceae) |
| 다이아토미스타 하위 분류군 | 딕티오코피세아에 (Dictyochophyceae) 펠라고피세아에 (Pelagophyceae) 볼리도피세아에 (Bolidophyceae) 디아토메아에 (Diatomeae) |
| 불확실한 위치 | 악티노프리이다 (Actinophryida) |
2. 세포 구조
황갈조류는 진핵생물의 일종으로, 세포는 껍질이 없거나 비늘, 로리카 또는 세포벽으로 덮여 있다. 이들은 단세포, 군체, 공통세포성 또는 다세포 생물일 수 있으며, 일부 갈조류는 분화된 조직을 가진 큰 다세포 엽상체로 발달하기도 한다.[12] 모든 황갈조류는 관 모양의 미토콘드리아 크리스타를 가지고 있으며, 이는 종속 영양 스트라메노필레스 뿐만 아니라 Rhizaria, Telonemia, Alveolata와 같은 다른 밀접하게 관련된 그룹과 공유하는 특성이다.[4][5]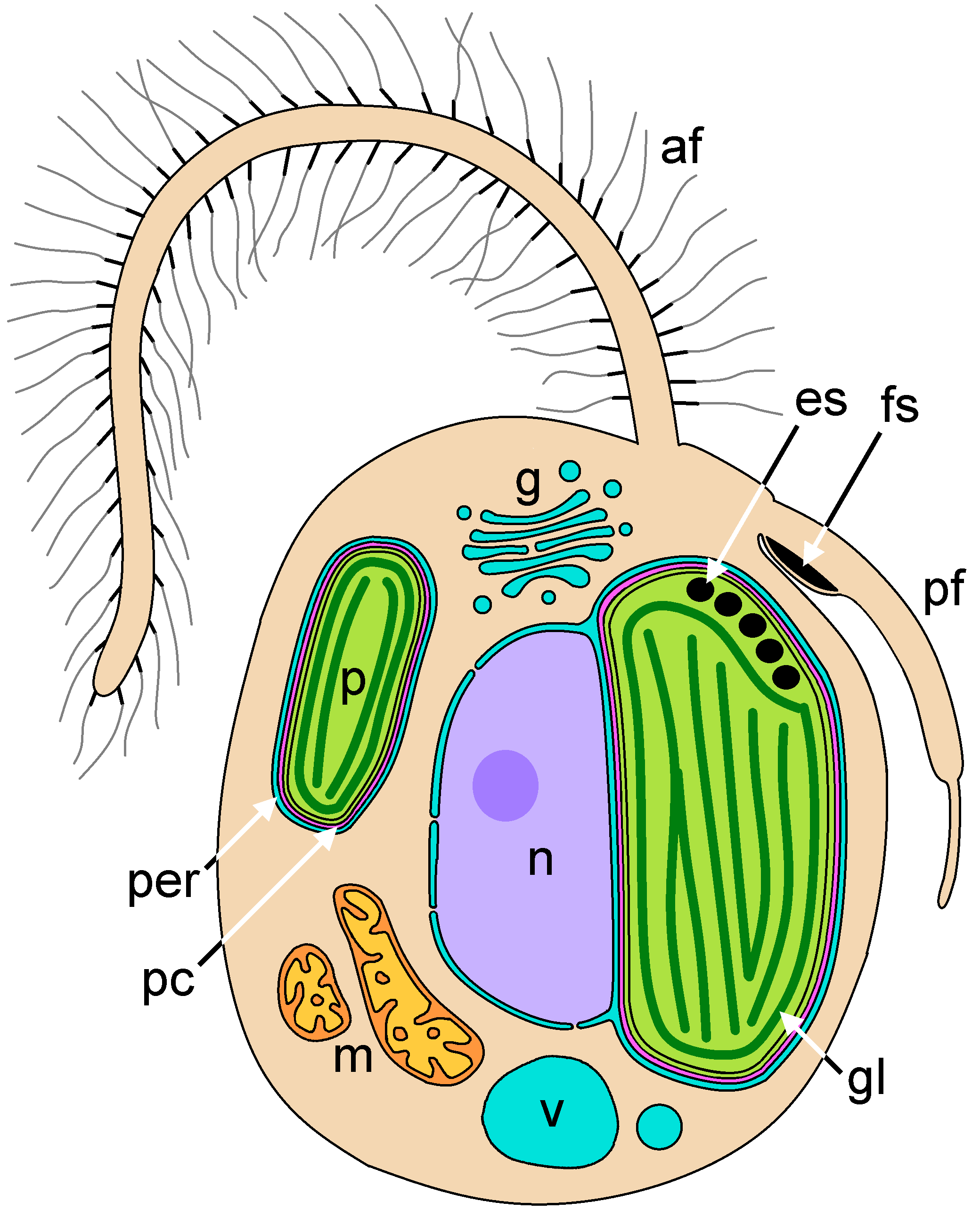
불등편모조류의 세포 구조는 각 강마다 다양하지만, 편모 소모를 가진 전편모와 클로로필 ''a''/''c''를 가진 노란색 엽록체와 같은 기본 구조는 공통적으로 나타난다.
대롱편모조식물의 주요 저장 물질은 β-1,3-글루칸(라미나란, 크리소라미나란)이며, 이는 세포 내 소포(크리소라미나란 소포)에 축적된다. 갈조류는 포도당 분자로 구성된 탄수화물인 크리소라미나린을 세포질 외부, 엽록체 외부에 위치한 소포에 저장한다. 세포질 지질 방울도 흔하게 발견되며, 녹조류와 식물에 흔한 저장 산물인 녹말은 없다.[12]
2. 1. 편모
운동 세포는 두 개의 현저하게 불균등한 편모를 자주 나타낸다. 앞쪽 편모("술")는 마스티고네메라고 하는 짚과 같은 속이 빈 3분할 털이 있고, 뒤쪽 편모("채찍")는 이러한 털이 없는 미성숙한 매끄러운 편모이다. 편모의 섬모 전환 영역은 일반적으로 전환 나선을 가지고 있다.[12] 다른 대롱편모조식물(=이형편모조식물) 생물과 마찬가지로, 이형편모조류도 앞편모에 세 부분(기부, 축부, 선단모)으로 구성된 편모 소모를 갖는다. 갈조류는 해조류이며, 개별 세포는 엽체의 구성 요소로서 운동성을 갖지 않지만, 생식 시 방출되는 유주자는 편모를 갖는다. 이는 규조류나 파에오탐니온 조류에서도 마찬가지이다.2. 2. 엽록체
대롱편모조식물은 대부분 광합성을 한다. 따라서 세포당 하나 이상의 광합성 색소체(엽록체)를 가질 수 있다. 일부 그룹은 광합성 능력과 색소를 잃었지만, 아미노산, 지질, 헴 그룹의 합성에 역할을 하는 것으로 추정되는 백색체를 가진 종을 포함한다. 대롱편모조식물은 다른 조류 그룹과 비교하여 뚜렷한 색소체 미세 구조를 가지고 있다.이들의 엽록체는 홍조류로부터의 이차 공생 사건에서 유래했으며, 이는 4개의 주변 세포막을 초래했다. 즉, 1차 색소체 막에 해당하는 두 개의 내부 막, 홍조류의 원형질막에 해당하는 세 번째 막, 그리고 식포 막에 해당하는 가장 바깥쪽 층이다. 이러한 특징은 엽록체가 단 두 개의 막만 있는 원색소체 조류(글라우코조류, 홍조류 및 녹조류)와 구별된다.[6] 대롱편모조식물 색소체의 두 개의 바깥층은 소포체(ER)와 인접해 있으며, 함께 엽록체 소포체 (CER)를 구성하며, 이는 주변 색소체 소포체 (PER)라고도 하며, 종종 핵막과 연결된다. 스트라메노필의 특징인 세 부분으로 구성된 편모 털은 PER 또는 핵막 내에서 생성된다.
두 번째 층과 세 번째 층 사이의 주변 색소체 구획(PC)은 다른 조류 그룹(예: 은편모조류 및 녹반조류)에서 이차 공생체의 흔적 세포 핵인 핵형체를 포함하는 별도의 영역이지만, 대롱편모조식물 내에서는 핵형체가 알려져 있지 않다. 대신, PC 내에서 합토조류 및 색소체충 조류에서 보이는 것과 유사한 다른 구조가 관찰되었다: PC 단백질이 국소화되는 "덩어리 같은 구조"와 소포 네트워크. CER 내에는 주변 색소체 막과 내부 핵막 사이에 밀접한 직접적인 접촉 부위가 두드러지게 나타나며, 여기서 지질 전달이 발생할 수 있으며, 다른 분자들의 교환도 발생할 수 있다.
일반적으로 색소체 기질 내에서 세 개의 적층된 틸라코이드가 가장 안쪽 막 아래, 색소체의 가장자리를 따라 흐르는 "띠 라멜라"로 분화된다. 나머지 틸라코이드는 세 개의 묶음으로 배열된다. 신크로모조류 및 황금모래조류에서는 각각 두 개 또는 세 개의 내부 막으로 둘러싸인 여러 색소체의 연합체가 공유된 바깥 막으로 둘러싸여 있다.
부등모조류의 엽록체는 홍조류 유래이며, 광합성 색소로 클로로필 ''a''/''c'', 기타 보조 색소로 다양한 카로티노이드를 갖는다. 일반적으로 세포 내에는 엽록체가 두 개 있으며, 사중막에 둘러싸여 있다. 최외막은 핵막과 연결된다. 누클레오모프는 존재하지 않는다.
부등모조류의 엽록체는 해초류의 엽록체와 매우 유사하지만, 부등모조류는 가돌 라멜라를 갖는다는 점에서 다르다. 가돌 라멜라는 엽록체 막 바로 아래에 있는 주머니 모양의 라멜라로, 이 안에 삼중 틸라코이드가 들어가는 형태를 하고 있다. 또한, 부등모조류의 엽록체 DNA는 가돌 라멜라 안쪽에 따라 링 모양으로 분포하고 있다.
부등모조류에는 황금색조강의 스푸멜라(Spumella) 및 파라피소모나스(Paraphysomonas), 디크티오카조강의 프테리도모나스(Pteridomonas) 및 실리오프리스(Ciliophrys) 등, 엽록체를 이차적으로 잃은 생물도 포함된다. 종속영양생물이 엽록체를 이차적으로 잃은 것인지, 아니면 원래 획득하지 않았는지에 대한 판단은 분자 계통수 상에서의 최소 가설 해석에 의한 경우가 많지만, 생물에 엽록체의 흔적 기관이 남아있는 경우에는, 이것이 이차적 손실의 유력한 증거가 된다. 부등모조류에서는, 프테리도모나스 다니카(Pteridomonas danica)로부터는 흔적적인 색소체인 백색체(leucoplast)가, 실리오프리스 인푸시오눔(Ciliophrys infusionum)으로부터는 엽록체 코드 유전자 중 하나인 ''rbc''L이 발견되었다. 이러한 직접적인 근거에 따라, 엽록체를 이차적으로 잃은 부등모조류와, 원래 획득하지 않은 무색 스트라메노필과는 엄밀히 구분된다.
2. 3. 저장 물질
대롱편모조식물의 저장 물질은 β-1,3-글루칸(라미나란, 크리소라미나란)이며, 세포 내 소포(크리소라미나란 소포)에 축적된다. 갈조류는 포도당 분자로 구성된 탄수화물인 크리소라미나린을 저장하는데, 이는 세포질 외부, 엽록체 외부에 위치한 소포에 저장된다는 점에서 다른 조류와 다르다. 세포질 지질 방울도 흔하게 발견된다. 녹조류와 식물에 흔한 저장 산물인 녹말은 없다.[12]3. 분류
대롱편모조식물의 분류는 아직 확립되지 않았지만, 크게 카키스타(Khakista)와 파에이스타(Phaeista)의 두 아문으로 나눌 수 있다.[17] 그러나 각 계통의 단일 계통성에 의문이 제기되고 있으며[18], 국제 원생생물 학회(ISOP)의 2019년도 분류에서는 대신 Chrysista 및 Diatomista라는 하위 분류군이 채택되었다.[19]
다음은 부등편모조류의 계통 분류이다.[30][31][32]
다음은 대롱편모조식물의 계통 분류이다.[33][34]
- 카키스타류 (Khakista)
- * 돌말강 (Bacillariophyceae)
- * 볼리도조강 (Bolidophyceae)
- 파에이스타류 (Phaeista)
- * 마리스타류 (Marista)
- ** 푸키스티아류 (Fucistia)
크리소메리스강 (Chrysomerophyceae)
갈조강 (Phaeophyceae)
파이오탐니오조강 (Phaeothamniophyceae)
스키조클라디아강 (Schizocladiophyceae)
황록조강 (Xanthophyceae)
- ** 히포기리스테아류 (Hypogyristea)
규질편모조강 (Actihrysophyceae 또는 Dictyochophyceae)
펠라고조강 (Pelagophyceae)
- ** 라피도이스티아류 (Raphidoistia)
핀귀오조강 (Pinguiophyceae)
침편모조강/라피도조강 (Raphidophyceae)
- * 림니스타류 (Limnista)
- ** 황조강 (Chrysophyceae)
- ** 진안점조강 (Eustigmatophyceae)
- ** 신크로마류 (Synchromophyceae)
- ** 피코파구스류 (Picophagea/Picophagophyceae)
- * 기타
- ** 시누라조강 (Synurophyceae)
2024년 현재, 황조식물은 23,314종으로 분류되었으며, 490종은 위치가 불확실하다. 그러나 이들은 10만 종이 넘을 것으로 추정되며, 그중 대다수가 규조류이다.[3]


아래는 진핵생물 분류에 대한 2019년 개정안에 따른 황조식물의 현재 분류이며, 이후에 기술된 조류의 강()과 각 강에 대해 기술된 종의 수를 포함한다.
| 계통 분류 | 하위 분류 | 종 수 |
|---|---|---|
| Chrysista | Aurearenophyceae | 1 |
| Chrysoparadoxophyceae | 1 | |
| Chrysophyceae | 1,274 | |
| Eustigmatophyceae | 218 | |
| Olisthodiscophyceae | 2 | |
| Phaeophyceae | 2,124 | |
| Phaeosacciophyceae | 8 | |
| Phaeothamniophyceae | 31 | |
| Raphidophyceae | 58 | |
| Schizocladiophyceae | 1 | |
| Xanthophyceae | 616 | |
| Diatomista | Dictyochophyceae | 217 |
| Pelagophyceae | 31 | |
| Pinguiophyceae | 5 | |
| Bolidophyceae | 18 | |
| Diatomeae | 14,684 | |
| 분류 불확실 | 490 |
3. 1. 크리시스타 (Chrysista)
Chrysista영어는 대롱편모조식물 계통 분류의 하나이다.다음은 대롱편모조식물의 계통 분류이다.[33][34]
SI와 SII의 두 분류군은 모두 Chrysista 계통을 구성한다.
- Chrysista
- * Aurearenophyceae – 1 종.
- * Chrysoparadoxophyceae – 1 종.
- * 황금색조강(Chrysophyceae) – 1,274 종.
- * 진정안점조강(Eustigmatophyceae) – 218 종.
- * Olisthodiscophyceae – 2 종.
- * 갈조강(Phaeophyceae) – 2,124 종.
- * Phaeosacciophyceae – 8 종.
- * Phaeothamniophyceae – 31 종.
- * 라피드조강(Raphidophyceae) – 58 종.
- * Schizocladiophyceae – 1 종.
- * 황록조강(Xanthophyceae) (Heterokontaela; Heteromonadeala; Tribophyceaela) – 616 종.
;황금색조류
: 담수를 중심으로 분포하는 조류로, 대부분 플랑크톤 생활을 한다. 단세포 유영성 생물이 많지만, 사야츠나기와 같은 수지상 군체나 Uroglenala와 같은 구상 군체도 보인다. 규산질 비늘을 세포 표면에 가진 생물도 있으며, 그 중 Synurala 속과 Mallomonasla 속은 시누라조강(Synurophyceae)으로 구분하는 의견도 있다. 옛날에는 팔마조와 펠라고조류(Pelagophyceae), 규질편모조류(Dictyochophyceae)도 황금색조류에 포함되었지만, 현재는 분리되어 있다.
;진정안점조류
: 십여 종의 작은 그룹으로, 주로 담수에서 서식한다. 다른 부등모조류와는 달리 유일하게 클로로필 ''c''를 가지지 않는다. 견고한 세포벽을 가진 부동 세포와 벽을 갖지 않는 유주 세포의 2형을 취하며, 유주 세포의 앞쪽 편모에 작은 털을 가지고 있다. 이름의 유래이기도 한 안점은 엽록체 내에 있는 것이 아니라 세포 앞부분의 편모 기부 근처에 존재하며, 광학 현미경으로도 매우 눈에 띈다.
;갈조류
: 해양에 널리 분포하는 대형 해조류로, 다시마, 미역, 히지키 등 식용으로 되는 것도 많다. 부등모조류 중에서는 가장 인간에게 친숙한 그룹이다.
;파에오탐니온조류
: 수지상의 군체를 형성하는 담수성 조류이다. 약 14속 30종, 대표종은 Phaeothamnion confervicolala이다. 옛날에는 황금색조강에 포함되었지만, 분자 계통 분석에 의해 갈조류나 황록조류에 가깝다는 것이 밝혀져 1998년, Bailey 등에 의해 강으로 독립시키는 의견이 제창되었다. 그러나 갈조류나 황록조류를 포함하는 이 그룹은 경계가 미묘하며, 어디까지를 독립적인 강으로 인정할지에 대해서는 의견이 분분하다.[21]
;라피드조류
: 담수와 해수에 분포한다. 세포 외피로서 세포벽이나 비늘을 갖지 않는 점, 따라서 매우 세포 형태가 불안정하고 변형되기 쉬운 점이 특징이다. 또한, 세포 내에 엽록체가 수 개~수십 개나 많이 보여 다른 조류와 구별하기 쉽다. 라피드조류는 담수에서는 존재감이 적지만, 해양, 특히 연안역에서는 빈번하게 대발생하여 적조를 일으킨다. 적조에서도 알 수 있듯이, 해산 종은 적갈색~오렌지색을 띠지만, 담수산 종은 녹조류와 같은 녹색이다. 이것은 담수종이 카로테노이드 중 푸코잔틴을 결여하기 때문이다.
;시조클라디아조류
: 2003년에 설립되었다. 1속 1종 Schizocladia ischiensisla만 기재되어 있다. 18S의 분자 계통 분석에서는 갈조류에 가깝다. 해산으로 분지상 군체를 형성하지만, 개개의 세포의 원형질은 연락하지 않는다. 또한, 세포벽은 알긴산이 많고, 셀룰로오스를 포함하지 않는다.
;황록조류
: 담수를 중심으로 분포하는 조류이다. 아주 소수가 해수역이나 기수역에 분포한다. 색깔은 황록색이라기보다는 녹색에 가깝고, 종종 녹조류와 구별하는 것이 곤란하지만, 전분을 포함하지 않기 때문에 요오드액과 같은 α-1,4 글루칸 특이적인 염색액으로 염색되지 않아 구별할 수 있다. 체제는 다채롭고, 구형 (Botrydiopsisla), 유영성 (Heterochlorisla), 아메바상 (Rhizochlorisla), 수지상 군체 (Mischococcusla), 사상 군체 (Tribonemala, Ophiocytiumla) 등 다양하다.
;크리소메리스조류
: 1995년에 설립되었다. 약 8속 16종이다. 사상 또는 엽상의 평면적인 군체를 형성한다. 개개의 세포의 원형질은 연락하지 않는다. 또한, 세포벽은 알긴산, 셀룰로오스 모두 포함하지 않는다.
;싱크로마조류
: 2007년에 설립된 강이다. 아메바 모양의 조류인 Synchroma grandela만이 포함된다. 이 조류의 엽록체는 각 세포에 여러 개가 있지만, 이들의 사중막 중 바깥쪽 2장이 여러 엽록체에서 공유된다.[22]
;아우레아레나조류
: 2008년에 설립된 강이다. 생활환이 편모충태와 구형 세포로 이루어진 Aurearena cruciatala만이 포함된다. 편모 장치를 중심으로 한 세포 형태와, 광합성 색소의 조성에 특징을 가진다. 분자 계통 분석에 따르면 본 강은 높은 독립성을 보이는 한편, 갈조강 및 황록조강과의 근연성이 시사되고 있다.[23]
3. 2. 디아토미스타 (Diatomista)
Diatomista는 대롱편모조식물의 계통 분류 중 하나이다.[33][34] Diatomista는 다음 분류를 포함한다.- 볼리도조강 (Bolidophyceae)
- 규조류 또는 돌말강 (Diatomeae, Bacillariophyceae)
- 규질편모조강 (Dictyochophyceae)
- 펠라고조강 (Pelagophyceae)
- 핀귀오조강 (Pinguiophyceae)
2024년 현재, 황조식물은 23,314종으로 분류되었으며, 그중 대다수가 규조류이다.[3] Diatomista에 속하는 각 강(class)에 대해 기술된 종의 수는 다음과 같다.
- 딕티오카조강 (Dictyochophyceae) – 217 종.
- 펠라고조강 (Pelagophyceae) – 31 종.
- 핑구이오조강 (Pinguiophyceae) – 5 종.
- 볼리도조강 (Bolidophyceae) – 18 종.
- 규조강 (Diatomeae) – 14,684 종.
| Diatomista |
|---|
| 볼리도조강 (Bolidophyceae) |
| 규조강 (Diatomeae) |
| 딕티오카조강 (Dictyochophyceae) |
| 펠라고조강 (Pelagophyceae) |
| 핑구이오조강 (Pinguiophyceae) |
; 볼리도조 (Bolidophyceae)
: 해산 피코플랑크톤으로, 1999년 Guillou 등에 의해 강이 설립되었다. 18S rRNA 계통 분석에서는 규조류의 자매군이 된다.[24]
; 규조 (Bacillariophyceae, Diatomeala)
: 규산질 껍질을 가진 조류로, 담수에서 해수까지 널리 분포한다. 단세포 조류로서는 크기가 큰 것도 있으며, 입수가 용이하기 때문에 현미경 관찰의 교재로 자주 사용된다.
; 디크티오카조 (Dictyochophyceae)
: 규산질 내골격을 가진 그룹. 원래는 규산 골격을 가진 디크티오카목만을 포함하는 강이었지만, 후에 방사상 총칭의 체제를 가진 페디넬라목이 추가되었다. 모두 편모는 1개로 앞편모뿐이다. 페디넬라목에는 방사상의 기관인 텐타클을 가진 것이 있는데, 이것이 태양충의 유축 가족과 비슷하기 때문에 Ciliophrysla 등은 종종 소속이 혼란스러웠다. 또한, 이 그룹에는 아메바 모양의 체제를 가진 Rhizochromulinala도 포함된다. 대표 속인 Dictyochala도 장기 배양주에서는 규산 골격이 소실되어 아메바화되어 버린다.[25]
; 펠라고조 (Pelagophyceae)
: Pelagomonasla, Pelagococcusla 등의 해산 외양성 피코플랑크톤으로 이루어진 그룹. 편모는 퇴화되어 뒷편모는 완전히 결실되고 편모근도 남아 있지 않다. 앞편모는 유지하고 있는 것과 이것도 잃은 것이 있으며, 편모를 전혀 가지지 않는 종도 있다.
; 핑기오조 (Pinguiophyceae)
: 2002년, 카와치 등에 의해 신설된 강. 5속 5종, 타입 속은 Pinguiochrysisla이다. 고도 불포화 지방산을 대량으로 생산하는 종이 포함된다.[26]
4. 생태
여러 강의 이질편모조류는 볼리도조강, 펠라고조강, 핑구이오조강 및 분열털조강과 같이 해양 서식지에서만 발견된다. 갈조류(Phaeophyceae)는 거의 전적으로 해양 조류이며, 담수 속은 매우 적다.[7]
황금조강, 황갈조강 및 황록조강은 주로 담수에서 서식하는 강이다. 흐르는 물 서식지(강, 하천)에서 황금색 조류(황금조강)와 황록색 조류(황록조강)는 흔하게 발견되며 때때로 풍부하게 존재한다. 특히 황금조류 속인 ''Hydrurus''는 일부 유역에서 널리 분포할 수 있으며, 차갑고 맑고 빠르게 흐르는 산간 계류에서 단단한 기질에 부착되어 흔히 발견된다. 강에서 흔히 발견되는 황록조강 속에는 자유롭게 떠다니거나 실 모양의 조류와 식물에 부착된 ''Vaucheria'', ''Tribonema'' 및 ''Bumilleria''가 있다.[7] 규조류는 다양성이 더 풍부하며, 60개 이상의 속이 강에서 흔히 발견된다. 많은 강 규조류는 물의 흐름에 의해 이동되는 것을 피하기 위해 기질에 부착하기 위한 다양한 전략을 개발했다. 가장 기본적인 전략은 세포막에서 형성된 다양한 탄수화물 구조인 세포외 고분자 물질을 생성하는 것이다. 더 빠르게 흐르는 물에서는 일부 규조류(예: ''Cocconeis'')가 접착성 막을 통해 기질에 직접 부착되어 자란다. 다른 규조류(예: ''Eunotia'', ''Nitzschia'')는 더 많은 영양분을 얻기 위해 물 기둥 위로 뻗을 수 있는 줄기나 군체 튜브를 생성한다.[8] 갈조류(황갈조강)는 매우 다양하지만 강에는 7개의 종만 존재한다. 이들은 복잡한 다세포 엽상체를 가지고 있지 않으며, 대신 해양 조상으로부터 독립적으로 진화한 저서생물의 실 모양 형태로 존재한다.[9]
부등편모조류는 세계 최대의 조류를 포함하는 그룹이며, 육상 식물에 필적하는 다양성과 생태적 중요성을 가진 그룹이다. 다시마나 미역과 같은 대형 해조류부터 규조류와 같은 미세 조류, 일부는 엽록체를 잃은 원생동물적인 생물도 포함된다.
5. 진화
다음은 부등편모조류의 계통 분류이다.[30][31][32]
{| class="wikitable"
|-
| 부등편모조류
|-
|
{| class="wikitable"
|-
| 플라티술쿠스강
|-
|
{| class="wikitable"
|-
! 사게니스타류
|-
|
| Eogyrea |
| 망형충류 |
|-
!
|-
| 비코소에카류
|-
|
{| class="wikitable"
|-
! 플라시디아류
|-
|
{| class="wikitable"
|-
| 플라시디아강
|-
|
{| class="wikitable"
|-
| 나노모나스강
|-
|
{| class="wikitable"
|-
| 오팔로모나스강
|-
! 오팔리나상강
|-
|
|}
|}
|}
|-
! 기리스타류
|-
|
{| class="wikitable"
|-
| 비기로모나드강
|-
|
{| class="wikitable"
|-
| 난균류
|-
|
{| class="wikitable"
|-
| 역모균류
|-
|
| 피르소니아강 |
| 대롱편모조식물 |
|}
|}
|}
|}
|}
|}
|}
다음은 대롱편모조식물의 계통 분류이다.[33][34]
{| class="wikitable"
|-
! 대롱편모조식물
|-
|
{| class="wikitable"
|-
! 카키스타류
|-
|
|-
! 파에이스타류
|-
|
{| class="wikitable"
|-
! Hypogyrista
|-
|
|-
! Chrysista
|-
|
{| class="wikitable"
|-
! Eustigmista
|-
|
|-
!
|-
! Phagochrysia
|-
|
{| class="wikitable"
|-
| 피코파구스강
|-
|
{| class="wikitable"
|-
| 신크로마강
|-
|
| Leukarachnion |
| 황조류 |
|}
|}
|-
! Xanthophytina
|-
|
{| class="wikitable"
|-
| 침편모조류
|-
|
{| class="wikitable"
|-
| 갈조류
|-
|
{| class="wikitable"
|-
| 크리소메리스강
|-
|
|}
|}
|}
|}
|}
|}
|}
|}
6. 관련 자료
분지학적 분류는 분기도만을 유효한 그룹으로 인식하고 측계통 또는 다계통 그룹의 사용을 거부하는 방법으로, 원생생물학자들이 선호한다. 2019년 국제 원생생물학회(International Society of Protistologists) 최신 개정판은 Ochrophyta를 상위 스트라메노필(Stramenopiles) 그룹 내, SAR 상군 내 유효한 분류군으로 인식한다. Chrysista와 Diatomista 사이 황갈조류 세분은 과학계에서 완전히 받아들여지며 계통 발생 분석으로 뒷받침된다.
다세포 해조류는 갈조강에 속하며, 기원전 3000년경 초기 중국, 기원전 300년경 그리스(테오프라스토스 등), 서기 500년경 일본 기록에서 묘사되었다. 이들에 대한 지식은 기록된 역사 이전 시기부터 식량, 염료, 의약품으로 사용되며 존재했을 것으로 보인다. 스트라메노필 조류에 대한 최초의 공식적인 설명은 린네가 1753년 ''식물의 종''에서 기술한 ''Fucus'' 속이었다. 그 직후, 단세포 황금조류가 오토 프리드리히 뮐러에 의해 기술되었다. 이 시기 과학적 발견 초기 동안, 갈조류는 식물로, 현미경 조류는 유충이라는 이름으로 동물로 취급되었다.
20세기 동안 진화 및 계통 발생에 대한 논의는 이형편모조류를 포함하기 시작했다. 투과 전자 현미경 및 분자 계통 발생학적 분석은 21세기까지 많은 새로운 그룹과 여러 강의 기술로 이어졌다. ''탈라시오시라 슈도나나''(Thalassiosira pseudonana)에 속하는 최초의 황조식물 게놈 시퀀싱은 2002년에 시작되었다.
참조
[1]
서적
"The kingdom Chromista, origin and systematics"
https://books.google[...]
"Round, F.E. and Chapman, D.J. (eds.). Progress in Phycological Research"
1986
[2]
서적
"Biologia e Filogenia das Algas"
https://books.google[...]
"Editora Artmed, Porto Alegre"
2006
[3]
논문
"Stramenopiles"
https://www.scienced[...]
2009-02-17
[4]
Q인용
[5]
Q인용
[6]
논문
"Endosymbiotic Origin of Chloroplasts in Plant Cells' Evolution"
[7]
서적
"River Algae"
"Springer International Publishing"
2016
[8]
서적
"River Algae"
"Springer International Publishing"
2016
[9]
서적
"River Algae"
"Springer International Publishing"
2016
[10]
논문
"Taxonomy and phylogeny of Heliozoa. III. Actinophryids"
http://www.bio-nica.[...]
2001
[11]
논문
"An Enigmatic Stramenopile Sheds Light on Early Evolution in Ochrophyta Plastid Organellogenesis"
2022-04-10
[12]
논문
"Validation of the phylum name ''Heterokontophyta''"
https://www.notulaea[...]
2023-10-11
[13]
서적
"Über Chlorosaccus eine neue Gattung der Süsswasseralgen nebst einiger Bemerkungen zur Systematik verwandter Algen"
"Norstedt"
1899
[14]
서적
"Algae: an introduction to phycology"
https://archive.org/[...]
"University Press"
1995
[15]
서적
"Algae An Introduction to Phycology"
https://books.google[...]
"Cambridge University Press, Cambridge"
1995
[16]
논문
"クリプト藻類・ハプト藻類間の姉妹群関係とクロムアルベオラータ仮説"
http://sourui.org/pu[...]
[17]
논문
"Zooflagellate phylogeny and classification"
https://pubmed.ncbi.[...]
1995
[18]
논문
"Seven gene phylogeny of heterokonts"
http://linkinghub.el[...]
[19]
논문
"Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes"
https://onlinelibrar[...]
2019-01
[20]
웹사이트
"不等毛植物類"
http://www.ujssb.org[...]
"日本分類学会連合"
2021-08-07
[21]
논문
"Phaeothamniophyceae classis nova.: a new lineage of chromophytes based upon photosynthetic pigments, ''rbc''L sequence analysis and ultrastructure"
[22]
논문
"Synchroma grande spec. nov. (Synchromophyceae class. nov., Heterokontophyta): an amoeboid marine alga with unique plastid complexes"
[23]
논문
"Aurearenophyceae classis nova, a New Class of Heterokontophyta Based on a New Marine Unicellular Alga Aurearena cruciata gen. et sp. nov. Inhabiting Sandy Beaches"
[24]
논문
"Bolidomonas: A new genus with two species belonging to a new algal class: Bolidophyceae (Heterokonta)"
[25]
논문
"Vestigial chloroplasts in heterotrophic stramenopiles ''Pteridomonas danica'' and ''Ciliophrys infusionum'' (Dictyochophyceae)"
[26]
논문
"The Pinguiophyceae classis nova, a new class of photosynthetic stramenopiles whose members produce large amounts of omega-3 fatty acids"
[27]
저널 인용
http://linkinghub.el[...]
[28]
웹인용
"Ochrophyta"
http://algaebase.org[...]
"World-wide electronic publication, National University of Ireland, Galway"
[29]
저널 인용
[30]
저널 인용
"Higher Level Classification of All Living Organisms"
[31]
저널 인용
"Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes"
https://hal.archives[...]
[32]
저널 인용
"Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes"
2013
[33]
저널 인용
"Higher Level Classification of All Living Organisms"
[34]
저널 인용
"Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes"
https://hal.archives[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com