미포자충류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
미포자충류는 미토콘드리아 대신 미토솜을 가지며 편모와 같은 운동성 구조가 없는 기생성 진핵생물이다. 이들은 매우 저항성이 강한 포자를 생성하며, 포자의 형태는 종을 구별하는 데 사용된다. 생활환은 숙주 내에서 포자가 발아하여 감염성 내용물을 방출하고, 숙주 세포 내에서 분열하거나 플라즈모디움을 형성하며, 무성생식 또는 유성생식을 통해 새로운 포자를 생성하는 과정을 포함한다. 미포자충류는 곤충, 갑각류, 어류, 포유류 등 다양한 숙주에 감염되어 질병을 유발하며, 특히 곤충에서는 수직 전파를 통해 난소에서 알로 전달되기도 한다. 사람에게는 미포자충증을 일으키며, 게놈은 작고 수평적 유전자 이동이 관찰되며, 균류와 계통학적으로 연관되어 있다.
더 읽어볼만한 페이지
- 생물 계통도에 관한 - 눈표범
눈표범은 고양이과 동물로 중앙아시아와 남아시아의 고산 지대에 서식하며, 길고 두꺼운 털, 큰 발, 긴 꼬리가 특징이며, 유전자 분석 결과 호랑이와 가장 가까운 종으로 밝혀졌으며, 밀렵과 서식지 파괴로 인해 개체 수가 감소하여 여러 국가에서 보호 노력이 진행 중이다. - 생물 계통도에 관한 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.
| 미포자충류 - [생물]에 관한 문서 | |
|---|---|
| 분류 정보 | |
| 계 | 진핵생물 |
| 역 | 오피스토콘타 |
| 계 | 균류 |
| 문 | 미포자충문 (Microsporidia) |
| 문 명명 | Sprague, 1977 또는 Balbiani, 1882 |
| 하위 분류 명칭 | 강 |
| 하위 분류 | 본문 참조 |
| 학술적 분류 | |
| 학명 | Microsporidiomycota |
| 명명자 | Benny, 2007 |
| 하위 분류 계급 | 강 및 목 |
| 하위 분류 참고 | |
| 하위 분류 | Paramicrosporidiales Morellosporales Nucleophagales Chytridiopsidea Hesseida Chytridiopsida Metchnikovellea Metchnikovellida Microsporidea Neopereziida Ovavesiculida Amblyosporida Glugeida Nosematida |
| 동의어 | |
| 명칭 | Microsporidia Balbiani, 1882 |
| 명칭 | Microsporidiida Labbé, 1899 |
| 명칭 | Cnidosporidia Doflein 190? |
| 명칭 | Microsporea Delphy, 1936 [1963], Levine et al., 1980Delphy, J. 1936. Sous-règne des Protozoaires. In: Perrier, R. (ed.). La Faune de la France en tableaux synoptiques illustrés, vol 1A. Delagrave: Paris. |
| 명칭 | Microsporidea Corliss & Levine 1963 |
| 명칭 | Microspora Sprague, 1969, 1977Sprague, V. (1977). Classification and phylogeny of the Microsporidia. In: Comparative pathobiology. vol. 2, Systematics of the Microsporidia. Lee A. Bulla & Thomas C. Cheng (ed.). pp. 1–30. New York: Plenum Press, [https://books.google.com/books?id=wpnwAAAAMAAJ]. |
| 명칭 | Microsporida Tuzet et al. 1971 |
| 이미지 | |
 | |
| 일반 정보 | |
| 한국어 이름 | 미포자충 |
| 영어 이름 | Microsporidia |
2. 형태
미포자충류는 미토콘드리아가 없는 대신 미토솜이라는 구조를 가지고 있으며, 편모와 같은 운동 기관도 없다.
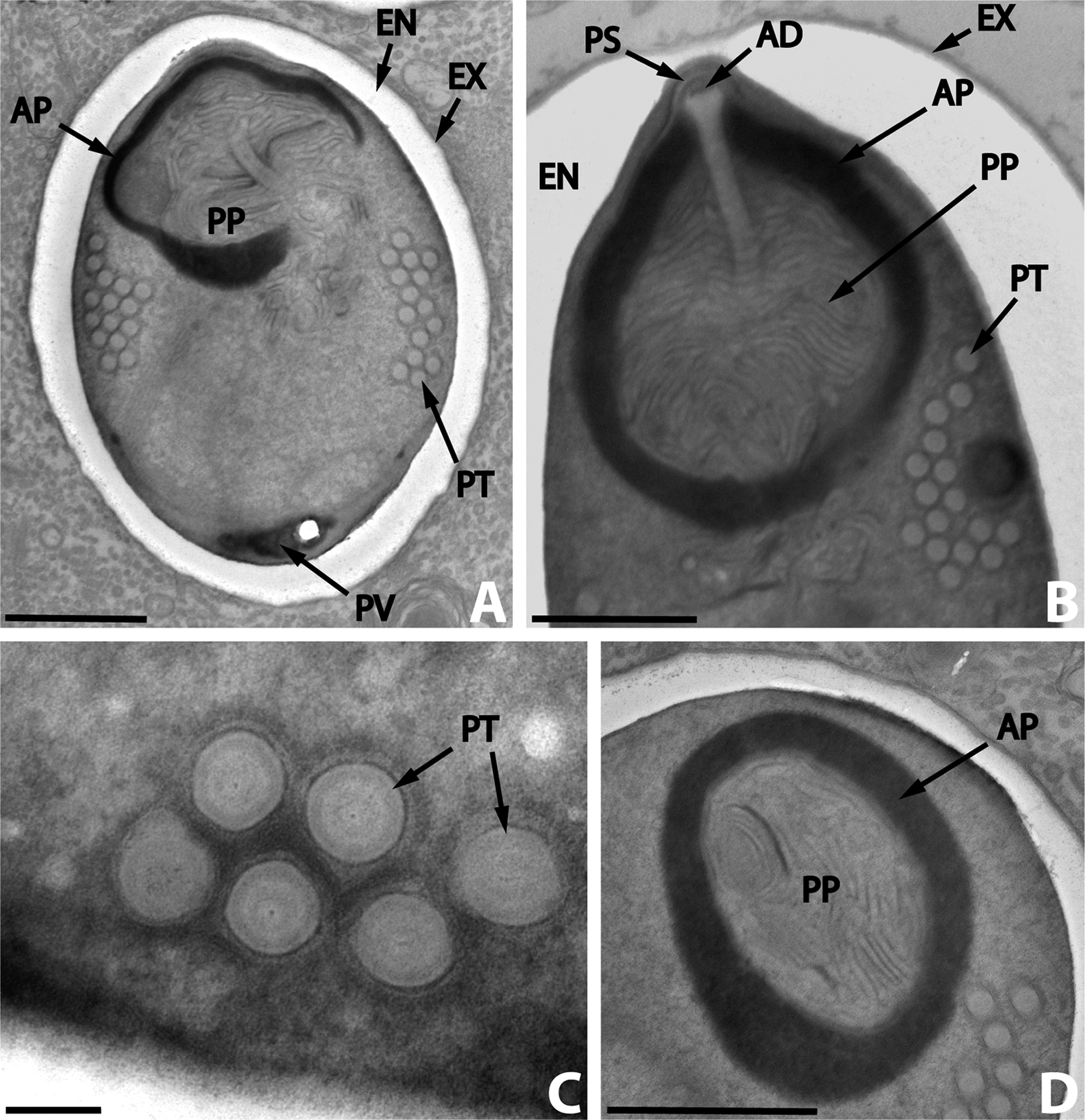
이들은 매우 저항력이 강한 포자를 만드는데, 이 포자는 숙주 몸 밖에서도 수년 동안 생존할 수 있다. 포자의 형태는 종마다 다양하여 종을 구별하는 중요한 기준이 된다. 대부분의 미포자충류 포자는 타원형이나 배 모양이지만, 막대 모양이나 구형 포자를 가진 종도 드물게 발견된다. 몇몇 속(속)은 그 속에서만 나타나는 독특한 모양의 포자를 생성하기도 한다. 포자의 크기는 대략 1~40 μm이다.
포자는 단백질과 키틴으로 이루어진 두꺼운 세포벽으로 보호된다. 이 벽은 세 개의 층으로 구성된다.
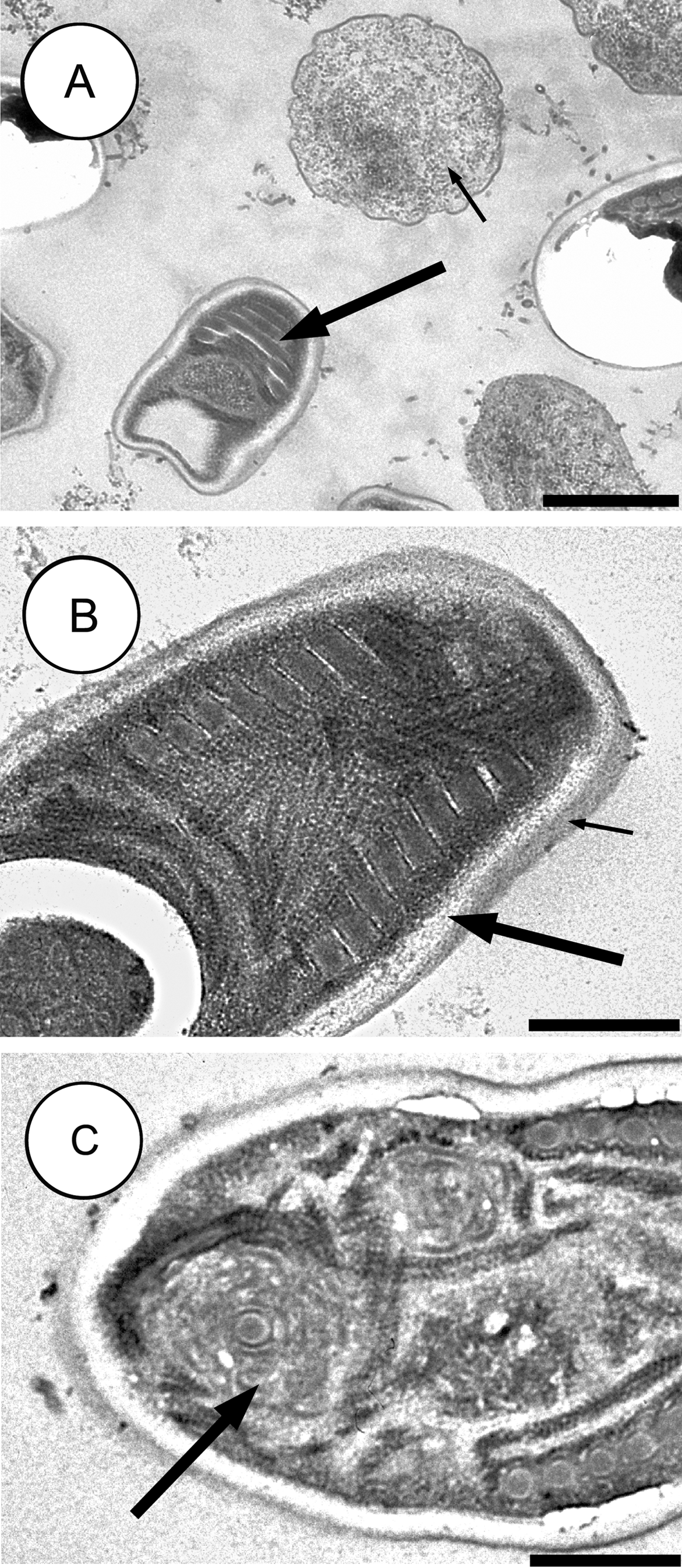
- 바깥쪽은 전자가 빽빽하게 모여 어둡게 보이는 ''외포자'' 층이다.
- 중간층은 넓고 특별한 구조가 없어 보이는 ''내포자'' 층으로, 키틴을 포함하고 있다.
- 가장 안쪽은 얇은 ''원형질막'' 층이다.
포자 내부에는 대부분 두 개의 세포핵이 서로 밀접하게 붙어 있는 '''이핵체'''(diplokaryon) 상태로 존재하지만, 핵이 하나만 있는 경우도 있다. 포자의 앞쪽 절반에는 길고 실 같은 ''극사''(polar filament)가 작살 모양의 구조를 이루고 있으며, 이 극사는 포자의 뒤쪽 절반에 코일처럼 감겨 있다. 극사의 앞쪽 끝은 ''극소체''라는 얇은 막 구조로 둘러싸여 있고, 극사의 뒤쪽에는 뒤쪽 ''액포''가 있다.[10] 이 극사는 숙주 세포에 침투할 때 사용되는 중요한 구조이다. 일부 자료에서는 이 구조를 믹소스포리디아류의 극사와 구분하기 위해 '''극관'''(polar tube)이라고 부르기도 한다.
3. 생활환
미포자충류의 생활환은 종에 따라 매우 다양하다. 일반적으로 경구 감염을 통해 수평적으로 전파되지만, 곤충을 숙주로 삼는 경우에는 숙주의 난소에서 알로 감염 물질이 전달되는 경난소 감염, 즉 수직 전파가 일어나기도 한다.
숙주의 장 내에서 포자는 특정 조건 하에 발아한다. 포자 종류에 따라 높은 pH, 폴리글루탐산, 혹은 뮤신의 존재 등이 발아를 유발하는 요인으로 작용할 수 있다. 이러한 조건이 충족되면 포자 내부의 삼투압이 급격히 높아져 포자 첨단의 가장 얇은 벽 부분이 파열된다. 동시에 후부 액포가 팽창하면서 '극사'(polar filament) 또는 '극관'(polar tube)이라 불리는 관 구조물이 빠르게 방출된다. 이 극관은 마치 호스를 뒤집어 꺼내듯이 숙주 세포를 향해 사출되어 세포막을 관통하며, 피하 주사 바늘과 같은 역할을 수행한다. 극관은 수백 μm까지 길게 뻗을 수 있으며, 이를 통해 포자 내부의 감염성 내용물인 포자질(sporoplasm)이 불과 수십 초라는 짧은 시간 안에 숙주 세포의 세포질 내부로 주입된다. 일부 경우에는 숙주 세포가 포자를 탐식하는 과정을 통해서도 감염이 이루어지기도 한다.
숙주 세포 안으로 들어간 포자질은 성장하기 시작한다. 이후의 증식 과정은 종류에 따라 다른데, 단순한 세포 분열을 반복하거나 여러 개의 핵을 가진 다핵체 상태의 플라즈모디움 또는 변형체를 형성하기도 한다. 감염된 숙주 세포는 미포자충류의 증식에 필요한 영양분을 충분히 공급하기 위해 대사 활동이 증가하거나 세포 자체가 비정상적으로 커져 표면적이 넓은 종양과 유사한 형태를 띠는 등 다양한 변화를 겪게 된다.
증식 단계를 거친 후에는 새로운 포자를 형성하는데, 이때 유성 생식이 관찰되는 경우가 있다. 미포자충류의 전체 생활환은 매우 다양하여, 어떤 종은 단순하게 무성 생식만으로 생활환을 완결하는 반면,[15] 다른 종들은 여러 종류의 숙주를 거치거나 무성 생식과 유성 생식 단계를 모두 포함하는 복잡한 생활환을 가지기도 한다. 또한, 생활사의 각 단계에서 서로 다른 종류의 포자가 만들어질 수 있는데, 이는 숙주 개체 내에서의 추가적인 확산(자가 감염)이나 숙주 간 전파 등 각기 다른 기능을 수행하기 위한 것으로 추정된다.
4. 병원성
미포자충류는 감염된 동물이나 인간에게 치명적이기보다는 만성적인 쇠약 질환을 유발하는 경우가 많으며, 이로 인해 숙주의 수명, 생식력, 체중, 전반적인 활력이 저하될 수 있다.[16][17][18][19] 수직 전파 또한 빈번하게 보고된다.
곤충, 갑각류, 어류에 기생하는 미포자충류는 각각 농업이나 어업에 심각한 피해를 줄 수 있는 병원체로 알려져 있다.
포유류에서는 자연적으로 치유되는 설사의 원인이 되기도 하며, 특히 면역력이 약화된 개체에게 기회감염을 일으키는 병원체로 알려져 있다. 사람에게 감염될 경우 미포자충증이라는 질병을 일으키는데, 최소 8개 속에 걸쳐 14종 이상의 미포자충이 사람에게 병을 일으키는 것으로 확인되었다.[23] 이는 가축, 가금, 애완동물 등과의 접촉을 통해 전파되는 인수공통감염병의 형태를 띨 수 있다.
이 외에도 다른 기생충인 점액포자충류나 그레가리나류 등에 기생하는 미포자충류도 알려져 있다.
4. 1. 곤충
곤충 숙주에서 미포자충류 감염은 치명적이기보다는 만성적인 쇠약 질환을 유발하는 경우가 많다. 감염된 곤충은 수명 단축, 생식력 저하, 체중 감소, 전반적인 활력 저하 등의 영향을 받는다.[16][17][18][19] 일부 미포자충류는 농업에 심각한 피해를 주는 병원체로 알려져 있다.곤충 숙주에서는 수직 전파, 특히 난소 전파가 빈번하게 보고된다. 이는 미포자충 기생충이 암컷 숙주의 난소에서 알로 이동하여 감염된 유충에서 증식하는 방식이다. 모기 ''Culex salinarius'' Coquillett에 감염되는 ''Amblyospora salinaria''와 모기 ''Culex tarsalis'' Coquillett에 감염되는 ''Amblyospora californica''는 이러한 난소 전파의 대표적인 예이다.[16][17][18][19]
일부 미포자충류는 말라리아 방제에 활용될 가능성이 있어 연구되고 있다. 예를 들어, 모기에 감염되는 ''Vavraia culicis''는 '진화에 영향받지 않는' 말라리아 방제법으로 주목받고 있다.[20] 말라리아의 주요 매개체인 ''Anopheles gambiae'' 모기가 미포자충에 감염되면, 모기 내 말라리아 기생충(Plasmodium falciparum)의 수가 감소하고 모기의 수명 또한 단축된다.[21] 대부분의 말라리아 감염 모기는 말라리아 기생충이 사람에게 전파될 수 있을 만큼 성숙하기 전에 자연적으로 죽기 때문에, 미포자충 감염을 통해 모기 사망률을 높이면 결과적으로 사람에게 말라리아가 전파되는 것을 줄일 수 있다. 2020년 5월에는 ''Anopheles arabiensis'' 모기의 중장과 난소에 공생하는 ''Microsporidia MB''라는 미포자충류가 ''P. falciparum''의 전파를 상당히 저해하며, 숙주 모기의 건강에는 뚜렷한 악영향을 주지 않고 수직 전파(유전)된다는 연구 결과가 보고되었다.[22]
''Nosema'' 속 미포자충류는 특정 곤충에게 질병을 일으키는 것으로 잘 알려져 있다. ''Nosema bombycis''는 누에에 미립자병을 일으켜 영양 불량으로 쇠약하게 만들고 결국 죽게 한다. ''Nosema apis''는 서양뒤영벌에 노제마병을 일으키는 병원체로 유명하다.
4. 2. 어류
어류에 기생하여 어업에 심각한 영향을 미치는 미포자충류 병원체가 알려져 있다.;''Heterosporis''
근육 세포 안에 기생하며 주머니 모양의 미세한 포자낭을 형성하고 그 안에서 포자를 만든다. 일본에서는 양식 뱀장어에 베코병(べこ病|베코뵤일본어)을 일으키는 ''Heterosporis anguillarum''이 알려져 있다. 이 기생충은 포자 형성이 끝나면 주변 근육 조직을 녹여 물고기 몸 곳곳을 움푹 패이게 만들어 상품 가치를 떨어뜨린다. 감염된 뱀장어는 성장하면서 회복되기도 하지만, 성장 지연만으로도 양식업자에게는 심각한 문제가 된다. 양식 기술의 발달로 최근에는 발생 빈도가 줄어들었다.
;''Glugea''
내장 등에 수 mm 크기의 둥근 낭종(cyst)을 형성한다. 숙주에게 직접적인 해를 끼치지 않는 경우도 많지만, 상품 가치를 떨어뜨리기 때문에 심각한 병원체로 간주된다. 일본에서는 산천어에 글루게아증(グルゲア症|구루게아쇼일본어)을 일으키는 ''Glugea plecoglossi'' 등이 알려져 있다.
4. 3. 포유류
미포자충류는 포유류를 포함한 동물과 인간에게 만성적인 쇠약 질환을 유발하는 경우가 많다.[16][17][18][19] 감염된 숙주는 수명, 생식력, 체중 및 전반적인 활력이 저하될 수 있다. 수직 전파도 빈번하게 보고된다.1950년대까지 미포자충류는 사람에게 병원성을 나타내지 않는다고 여겨졌으나, 1959년 처음으로 인간 감염 사례가 보고되었다. 이후에도 보고는 드물었지만, 이는 건강한 성인에게 감염되어도 자연 치유되거나 심각한 증상이 없어 잘 알려지지 않았기 때문이다. 실제 감염률은 인구의 2%에서 많게는 70%에 달할 정도로 흔한 병원체로 추정된다. 특히 영아나 장기 이식, 후천성면역결핍증(AIDS) 등으로 면역력이 저하된 사람에게는 심각한 증상을 유발할 수 있어 기회감염증으로 주목받고 있다.
사람에게 감염되는 주요 경로는 가축, 가금, 애완동물(개, 새 등)과의 접촉으로 인한 인수공통감염병 형태이다. 동물보호시설에서는 개의 30%가 감염된 적이 있으며, 강아지 집단 폐사의 원인이 되기도 한다. 주된 감염 경로는 경구 감염으로 추정되며, 음용수가 감염원이 될 수 있다. 일부 종류는 성접촉이나 호흡기를 통한 감염 가능성도 제기된다. 동물에서는 선천성 감염이 알려져 있지만, 사람에게서는 아직 보고된 바 없다. 각막의 외상성 감염 사례도 있다. 감염 부위는 소화기관이 가장 흔하지만, 생식기, 호흡기, 배설기, 신경계 등 다양한 기관에서 발견될 수 있다.
사람에게 병원성을 나타내는 미포자충류는 8속 13종이 알려져 있다. 대표적인 종과 유발 증상은 다음과 같다.
;''엔세팔리토존 쿠니쿨리'' (''Encephalitozoon cuniculi'')
: 간염, 뇌염, 전신성 감염
;''엔세팔리토존 인테스티날리스'' (''Encephalitozoon intestinalis'')
: 설사, 전신성 감염, 표층 각결막염 (과거 ''Septata intestinalis''로 불림)
;''엔세팔리토존 헬렘'' (''Encephalitozoon hellem'')
: 표층 각결막염, 부비강염, 호흡기 질환, 전립선 농양, 전신성 감염
;''브라키올라 알제라에'' (''Brachiola algerae'')
: 피부 감염 (과거 ''Nosema algerae''로 불림)
;''엔테로사이토존 비에네우시'' (''Enterocytozoon bieneusi'')
: 흡수 불량, 설사, 담관염
이 외에도 ''Trachipleistophora'', ''Pleistophora'', ''Nosema'', ''Vittaforma'', ''Microsporidium'' 속의 일부 종들이 간질성 각막염이나 근염을 일으키는 것으로 알려져 있다.
5. 초기생
미포자충류는 다양한 숙주를 감염시킬 수 있는데, 이때 숙주 자신이 다른 생물에 기생하는 기생충인 경우도 있다. 이러한 경우 미포자충류는 초기생(hyperparasite), 즉 기생충에 기생하는 기생충이 된다. 예를 들어, 디게네아(Digenea) (기생성 편형동물)에 기생하는 미포자충류가 18종 이상 알려져 있다. 이 디게네아들은 다양한 척추동물과 연체동물에서 기생 생활을 한다. 이들 초기생 미포자충류 중 8종은 ''Nosema'' 속에 속한다.[24] 또 다른 예로, 미포자충류의 일종인 ''Toguebayea baccigeri''는 디게네아의 일종인 파우스툴리드과(Faustulidae) ''Bacciger israelensis''에 기생한다. 이 ''Bacciger israelensis''는 바닷물고기인 보그(''Boops boops'', 농어목 도미과)의 장에 기생하는 기생충이다.[25]
6. 게놈
미포자충류는 알려진 진핵생물 중 가장 작은 핵 게놈을 가진다.[26] 기생 생활 방식에 적응하면서 많은 미토콘드리아 및 골지체 관련 유전자를 잃어버렸고, 심지어 리보솜 RNA의 크기조차 대부분의 다른 진핵생물에 비해 줄어들었다.[26]
그 결과, 미포자충류의 게놈 크기는 다른 진핵생물보다 훨씬 작다. 현재까지 알려진 게놈 크기는 2.5~11.6 메가베이스(Mb)이며, 이는 1,848~3,266개의 단백질을 암호화하는 수준으로 많은 세균과 비슷한 범위이다.[26] 특히 ''엔세팔리토존'' 속(''Encephalitozoon'')의 경우 게놈 크기가 3 Mb 미만으로, 원핵생물인 대장균보다도 작다.
2001년에는 ''엔세팔리토존 쿠니쿨리''(''Encephalitozoon cuniculi'')의 전체 게놈 서열이 해독되면서 극단적으로 압축된 게놈 구조가 밝혀졌다. *E. cuniculi*의 게놈은 11개의 염색체로 이루어져 있으며, 각 염색체는 22만~32만 염기쌍으로 매우 작다. 주목할 만한 특징은 모든 염색체의 양쪽 말단에 리보솜 RNA 유전자가 배열되어 있다는 점이다. 이러한 특징은 2차 공생 조류의 핵형질체에서도 발견되는데, 이는 극단적으로 작은 게놈 크기와 리보솜 RNA 유전자의 특정 배열 사이에 연관성이 있을 가능성을 시사한다.
수평적 유전자 이동(HGT) 역시 미포자충류 진화에 영향을 미친 것으로 보인다. 예를 들어, ''엔세팔리토존 로말레아이''(''Encephalitozoon romaleae'')와 ''트라키플레이스토포라 호미니스''(''Trachipleistophora hominis'')의 게놈에는 동물, 세균, 일부 균류에서 유래한 것으로 보이는 유전자가 포함되어 있다.[26]
또한 미포자충류는 DNA 손상을 복구하는 기구를 가지고 있다. DNA 손상 부위에 모여 검사점 신호 전달 연쇄 반응(cascade)을 활성화하는 데 기여하는 것으로 여겨지는 Rad9-Rad1-Hus1 단백질 복합체(9-1-1 복합체)를 암호화하는 유전자가 존재한다.[27] 9-1-1 복합체 외에도 다른 DNA 복구 기구의 구성 요소들도 존재하여 미포자충류에서 DNA 손상 복구가 일어날 가능성을 시사한다.[1][27]
7. 계통 분류
미포자충류는 단세포 기생 생물로, 최근 분자계통학 연구 결과 균류에 속하거나 균류와 가까운 자매군으로 분류된다.[34][35][36][37] 이는 기생 생활에 적응하며 고도로 특수화된 결과로 이해된다.[33]
후편모생물 내에서 미포자충류의 계통학적 위치는 홀로미코타(Holomycota) 그룹에 속하며, 특히 로젤라아계(Rozellomycota) 또는 은진균문(Cryptomycota) 내에서 로젤라목(Rozellales) 등과 가까운 관계에 있는 것으로 보인다.[43][44][45][46][28][29] 후편모생물의 계통 분류 연구에 따르면, 미포자충류는 로젤라아계/은진균문 분기군 내에서 Namako-37 그룹과 자매군을 이루며, 이들은 다시 로젤라목과 더 큰 분기군을 형성한다. 이 전체 그룹은 BCG1이라는 다른 미분류 그룹과 자매 관계에 있으며, 이들은 진균류 및 아펠리디움류와 함께 오피스토스포리디아(Opistosporidia) 또는 넓은 의미의 균류를 구성하는 것으로 제안된다.
이러한 계통학적 발견은 미포자충류의 분류 체계 재정립으로 이어지고 있으며, 학명 사용 등에서도 변화가 나타나고 있다.
7. 1. 하위 분류
미포자충류의 분류는 역사적으로 많은 변화를 겪었다. 최초로 기술된 ''노세마속''(Nosema)은 나겔리에 의해 일부 세균 및 효모와 함께 균류 그룹 쉬조미세테스(Schizomycetes)로 분류되었다.[30][31] 이후 오랫동안 미포자충류는 매우 원시적인 진핵생물로 여겨져 원생동물 그룹 크니도스포라(Cnidospora)에 속하는 것으로 간주되었다.[4] 특히 미토콘드리아가 없다는 특징 때문에 디플로모나드류, 파라바살리드류, 고세균류 등 다른 원생동물과 함께 아르케조아(Archezoa) 그룹으로 분류되기도 했다.[32]그러나 최근 연구들은 이러한 초기 기원설을 반증하고 있다. 현재 미포자충류는 숙주로부터 필요한 자원을 얻으면서 불필요해진 기능을 퇴화시킨, 고도로 진화하고 특수화된 생물로 여겨진다.[33] 또한, 포자를 형성하는 생물들이 일반적으로 그렇듯, 미포자충류 역시 원시적이라고 보기 어려운 복잡한 유성 및 무성 생식 체계를 가지고 있다. 2000년대 중반 이후의 연구들은 미포자충류가 균류 내에 속하거나 균류와 자매군을 이루는 공통 조상을 가진다고 본다.[34][35][36][37]
과거에는 서식지나 형태적 특징에 기반한 다양한 분류 체계가 제안되었다. 예를 들어 Vossbrinck과 Debrunner-Vossbrinck는 서식지에 따라 아쿠아스포리디아(Aquasporidia), 마리노스포리디아(Marinosporidia), 테레스포리디아(Terresporidia)의 세 강으로 나누자고 제안했다.[38] Cavalier-Smith는 1993년에 형태적 특징을 바탕으로 다음과 같은 분류 체계를 제안하기도 했다.[39]
| 분류 단계 | 분류명 |
|---|---|
| 아문 | 루디미크로스포라(Rudimicrospora) |
| 강 | 미니스포레아(Minisporea) |
| 목 | 미니스포리다(Minisporida) |
| 강 | 메치니코벨레아(Metchnikovellea) |
| 목 | 메치니코벨리다(Metchnikovellida) |
| 아문 | 폴라로플라스타(Polaroplasta) |
| 강 | 플레이스토포리데아(Pleistophoridea) |
| 목 | 플레이스토포리다(Pleistophorida) |
| 강 | 디스포레아(Disporea) |
| 아강 | 유핵아강(Unikaryotia) |
| 아강 | 이핵아강(Diplokaryotia) |
현재는 분자계통학적 연구 결과를 바탕으로 분류 체계가 계속해서 재정립되고 있다. Wijayawardene 등이 2020년에 제안한 분류 체계[41][42]는 다음과 같은 주요 분류군을 포함한다.
- 목 파라미크로스포리달레스(Paramicrosporidiales)
- * 과 파라미크로스포리디아세아에(Paramicrosporidiaceae)
- 목 모렐로스포랄레스(Morellosporales)
- * 과 미토스포리디아세아에(Mitosporidiaceae)
- * 과 모렐로스포라세아에(Morellosporaceae)
- 목 뉴클레오파갈레스(Nucleophagales)
- * 과 뉴클레오파가세아에(Nucleophagaceae)
- 강 키트리디오프시데아(Chytridiopsidea)
- * 목 헤세이다(Hesseida)
- ** 과 헤세이다에(Hesseidae)
- * 목 키트리디오프시다(Chytridiopsida)
- ** 과 벅스테후데이다에(Buxtehudeidae)
- ** 과 키트리디오프시다에(Chytridiopsidae)
- 강 메치니코벨레아(Metchnikovellea)
- * 목 메치니코벨리다(Metchnikovellida)
- ** 과 암피아칸티다에(Amphiacanthidae)
- ** 과 메치니코벨리다에(Metchnikovellidae)
- 강 마이크로스포리디아(Microsporidea)
- * 과 아벨스포리다에(Abelsporidae)
- * 과 아레오스포리다에(Areosporidae)
- * 과 부렐레리다에(Burenellidae)
- * 과 버키다에(Burkeidae)
- * 과 쿠구르델리다에(Cougourdellidae)
- * 과 실린드로스포리다에(Cylindrosporidae)
- * 과 두보스키이다에(Duboscqiidae)
- * 과 골베르기이다에(Golbergiidae)
- * 과 마이크로필리다에(Microfilidae)
- * 과 네오노세모이디다에(Neonosemoidiidae)
- * 과 플레이스토스포리디이다에(Pleistosporidiidae)
- * 과 슈도플레이스토포리다에(Pseudopleistophoridae)
- * 과 스트리아토스포리다에(Striatosporidae)
- * 과 텔로믹시다에(Telomyxidae)
- * 과 톡소글루게이다에(Toxoglugeidae)
- * 과 투제티다에(Tuzetiidae)
- * 목 암블리오스포리다(Amblyosporida)
- ** 과 암블리오스포리다에(Amblyosporidae)
- ** 과 카우도스포리다에(Caudosporidae)
- ** 과 걸리이다에(Gurleyidae)
- * 목 네오페레지이다(Neopereziida)
- ** 과 야나체키이다에(Janacekiidae)
- ** 과 베르왈디이다에(Berwaldiidae)
- ** 과 네오페레지이다에(Neopereziidae)
- ** 과 튜불리노세마티다에(Tubulinosematidae)
- * 목 오바베시쿨리다(Ovavesiculida)
- ** 과 오바베시쿨리다에(Ovavesiculidae)
- * 목 글루게이다(Glugeida)
- ** 과 파실리스포리다에(Facilisporidae)
- ** 과 글루게이다에(Glugeidae)
- ** 과 미오스포리다에(Myosporidae)
- ** 과 페레지이다에(Pereziidae)
- ** 과 플레이스토포리다에(Pleistophoridae)
- ** 과 스프레게이다에(Spragueidae)
- ** 과 텔로하니이다에(Thelohaniidae)
- ** 과 유카리오니다에(Unikaryonidae)
- * 목 노세마티다(Nosematida)
- ** 과 엔세팔리토조오니다에(Encephalitozoonidae)
- ** 과 엔테로사이토조오니다에(Enterocytozoonidae)
- ** 과 헤테로베시쿨리다에(Heterovesiculidae)
- ** 과 므라제키이다에(Mrazekiidae)
- ** 과 노세마티다에(Nosematidae)
- ** 과 오르도스포리다에(Ordosporidae)
참조
[1]
논문
Sur les microsporidies ou psorospermies des Articulés
https://www.biodiver[...]
[2]
서적
Sous-règne des Protozoaires. In: Perrier, R. (ed.). La Faune de la France en tableaux synoptiques illustrés, vol 1A.
Delagrave
[3]
논문
A Newly Revised Classification of the Protozoa
https://www.research[...]
[4]
논문
Establishment of the Microsporidea as a new class in the protozoan subphylum Cnidospora
[5]
서적
Classification and phylogeny of the Microsporidia. In: Comparative pathobiology. vol. 2, Systematics of the Microsporidia. Lee A. Bulla & Thomas C. Cheng (ed.).
https://books.google[...]
Plenum Press
[6]
논문
How do Microsporidia invade cells?
[7]
논문
A higher level phylogenetic classification of the Fungi
http://www.umich.edu[...]
[8]
서적
Protistes Eucaryotes : Origine, Evolution et Biologie des Microbes Eucaryotes
https://hal.archives[...]
HAL
[9]
논문
The magnitude of fungal diversity: The 1.5 million spices estimate revisited
[10]
웹사이트
Cytology and taxonomy of the microsporidia
http://www.cob.lu.se[...]
[11]
논문
Microsporidiosis: an emerging and opportunistic infection in humans and animals
2005-04
[12]
서적
Les Psorospermies des Articulés ou Microsporidies
https://www.biodiver[...]
Doin
[13]
서적
Parasites of North American Freshwater Fishes
https://books.google[...]
University of California Press
[14]
논문
Molecular and ultrastructural characterization of Dictyocoela diporeiae n. sp. (Microsporidia), a parasite of Diporeia spp. (Amphipoda, Gammaridea)
[15]
논문
Multiple losses of sex within a single genus of Microsporidia
[16]
논문
Development, ultrastructure, and mode of transmission of Amblyospora sp. (Microspora) in the mosquito
1979-08
[17]
논문
Significance of transovarial infections of Amblyospora sp. (Microspora:Thelohaniidae) in relation to parasite maintenance in the mosquito Culex salinarius
1979-09
[18]
논문
A comparison of the life cycles of two Amblyospora (Microspora: Amblyosporidae) in the mosquitoes Culex salinarius and Culex tarsalis Coquillett
[19]
논문
Amblyospora salinaria n. sp. (Microsporidia: amblyosporidae), parasite of Culex salinarius (Diptera: culicidae): its life cycle stages in an intermediate host
https://zenodo.org/r[...]
1998-05
[20]
서적
Chapter 12 Microsporidians as Evolution-Proof Agents of Malaria Control?
[21]
논문
A Possible Mechanism for the Suppression of Plasmodium berghei Development in the Mosquito Anopheles gambiae by the Microsporidian Vavraia culicis
[22]
논문
A microsporidian impairs Plasmodium falciparum transmission in Anopheles arabiensis mosquitoes
[23]
논문
The genome of the obligate intracellular parasite Trachipleistophora hominis: New insights into microsporidian genome dynamics and reductive evolution
2012-10
[24]
논문
Ultrastructure and development of Nosema podocotyloidis n. sp. (Microsporidia), a hyperparasite of Podocotyloides magnatestis (Trematoda), a parasite of Parapristipoma octolineatum (Teleostei)
[25]
논문
Ultrastructural and molecular study of the microsporidian Toguebayea baccigeri n. gen., n. sp., a hyperparasite of the digenean trematode Bacciger israelensis (Faustulidae), a parasite of Boops boops (Teleostei, Sparidae)
EDP Sciences
[26]
논문
Latest Progress in Microsporidian Genome Research
[27]
논문
The Rad9-Rad1-Hus1 DNA Repair Clamp is Found in Microsporidia
2022-04-10
[28]
논문
Outline of Fungi and fungus-like taxa
http://www.mycospher[...]
[29]
논문
Microsporidia: a new taxonomic, evolutionary, and ecological synthesis
[30]
논문
Über die neue Krankheit der Seidenraupe und verwandte Organismen. pp. 760–61. In: Caspary, R. (ed.). Bericht über die Verhandlungen der 33. Versammlung deutscher Naturforscher und Aerzte, gehalten in Bonn von 18 bis 24 September 1857
https://archive.org/[...]
[31]
논문
Microsporidia: biology and evolution of highly reduced intracellular parasites
http://www3.botany.u[...]
[32]
논문
Kingdom protozoa and its 18 phyla
[33]
논문
Simplicity and Complexity of Microsporidian Genomes
2004-12
[34]
논문
Evidence from small-subunit ribosomal RNA sequences for a fungal origin of Microsporidia
2005-09
[35]
논문
Loss of the flagellum happened only once in the fungal lineage: phylogenetic structure of Kingdom Fungi inferred from RNA polymerase II subunit genes
[36]
저널
Assessing the microsporidia-fungi relationship: Combined phylogenetic analysis of eight genes
2006-06-00
[37]
저널
Microsporidia evolved from ancestral sexual fungi
2008-11-00
[38]
저널
Molecular phylogeny of the Microsporidia: ecological, ultrastructural and taxonomic considerations
2005-05-00
[39]
저널
Kingdom Protozoa and its 18 phyla
1993-00-00
[40]
서적
Protista 2: Handbook on zoology
https://books.google[...]
Nauka
2007-05-00
[41]
저널
Outline of Fungi and fungus-like taxa
http://www.mycospher[...]
[42]
저널
Outline of Fungi and fungus-like taxa – 2021
https://www.research[...]
[43]
저널
The Ediacaran emergence of bilaterians: congruence between the genetic and the geological fossil records
http://rstb.royalsoc[...]
2008-04-27
[44]
저널
Estimating the timing of early eukaryotic diversification with multigene molecular clocks
2011-08-16
[45]
저널
Novel Predators Reshape Holozoan Phylogeny and Reveal the Presence of a Two-Component Signaling System in the Ancestor of Animals
[46]
저널
High-level classification of the Fungi and a tool for evolutionary ecological analyses
2018-02-22
[47]
문서
BCG1 = basal clone group 1, BCG2 = basal clone group 2
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com