인산 가수 분해 효소
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
인산 가수 분해 효소는 포스포모노에스터의 가수분해를 촉매하여 기질에서 인산염 부분을 제거하는 효소이다. 인산 가수 분해 효소는 기질의 특정 부위를 탈인산화할 수 있으며, 이러한 기질 인식을 조절하는 메커니즘은 "인산가수분해효소 코드"로 불린다. 인산 가수 분해 효소는 단백질, 뉴클레오타이드 등 다양한 기질에 작용하며, 세포 내 신호 전달, 포도당 신생합성 등 다양한 생화학적 과정에 관여한다. IUBMB는 인산 가수 분해 효소를 기질 특이성 및 서열 상동성에 따라 104가지 효소군으로 분류한다.
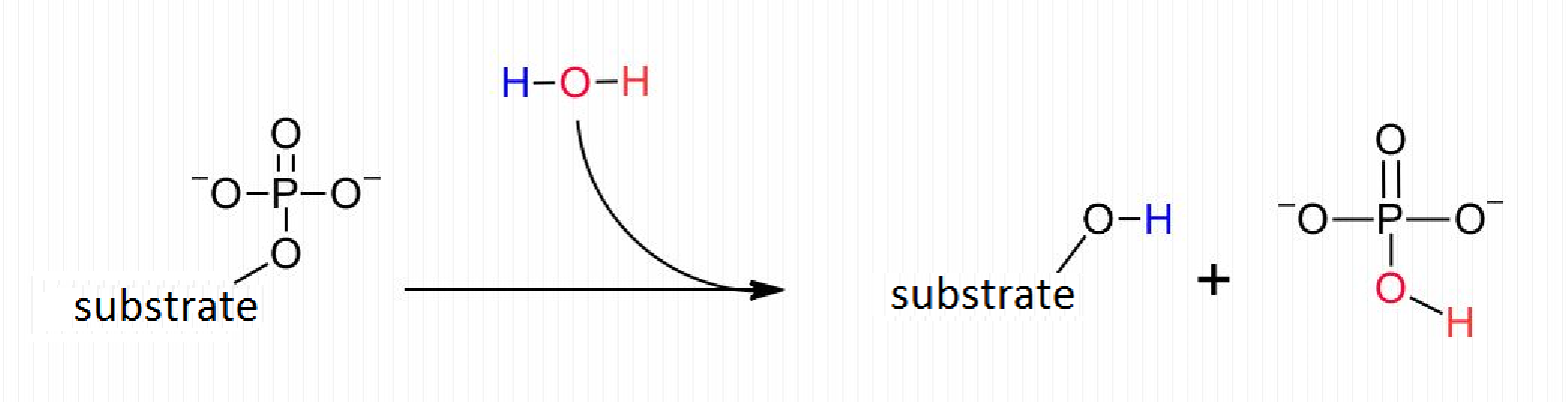
인산가수분해효소는 인산모노에스터의 가수분해 반응을 촉매하여 기질로부터 인산기 부분을 제거하는 효소이다.[38][4][21] 이 반응 과정에서 물(H2O) 분자가 관여하여 분해되는데, 이때 생성된 수산기(–OH)는 인산 이온에 결합하고, 수소 이온(H+)은 다른 생성물의 수산기를 양성자화한다. 결과적으로 인산모노에스터가 분해되어 인산 이온과 자유 수산기를 가진 분자가 생성된다.[38][4][21]
키네이스와는 대조적으로 인산기를 제거하는 효소인 인산가수분해효소는, 키네이스에 비해 더 다양한 기질을 인식하고 반응을 촉매하는 경향을 보인다.[38][4][21] 인산가수분해효소는 종류가 매우 다양하고 세포 내에서 중요한 기능을 수행하지만, 아직 모든 종류나 각각의 정확한 기질이 밝혀진 것은 아니다.[37][3][20][38][4][21] 특정 세포나 조직 등 생체 내에서 발현되는 인산가수분해효소 전체의 집합을 포스파톰(phosphatome)이라고 부르며, 이에 대한 완전한 이해는 아직 부족한 상태이다.[37][3][20]
2. 생화학
인산가수분해효소는 기질 특이성이 높아, 기질 분자의 특정 부위에 있는 인산기만을 선택적으로 제거(탈인산화)할 수 있다.[40][6][23] 기질을 인식하고 결합하는 구체적인 메커니즘에 대해서는 하위의 '인산가수분해효소 코드' 및 '도킹 상호작용' 섹션에서 더 자세히 다룬다.
2. 1. 인산가수분해효소 코드
인산가수분해효소는 매우 특이적으로 기질의 특정 부위를 탈인산화시킬 수 있다. 인산가수분해효소가 기질을 인식하는 메커니즘과 규칙, 즉 "인산가수분해효소 코드"를 밝히는 연구는 아직 진행 중이다. 하지만 최근 9종의 진핵생물 유전체에 암호화된 모든 단백질 인산가수분해효소를 비교 분석한 연구 결과가 나오기도 했다.[40][6][23]
연구에 따르면, 기질 결합에는 "도킹 상호작용"이 중요한 역할을 하는 것으로 밝혀졌다.[37][3][20] 인산가수분해효소는 기질에 있는 다양한 모티프(단백질의 특정 구조 단위)를 인식하고 상호작용한다. 이 모티프들은 효소의 활성 부위가 아닌 다른 결합 부위(도킹 부위)에 상대적으로 약한 친화력으로 결합한다. 각각의 도킹 상호작용은 약하지만, 여러 상호작용이 동시에 일어나면서 누적 효과를 만들어내어 결과적으로 특정 기질에만 결합하는 결합 특이성을 갖게 된다.[41][7][24] 또한, 이러한 도킹 상호작용은 알로스테릭 조절을 통해 인산가수분해효소의 촉매 활성에도 영향을 미칠 수 있다.[42][8][25]
2. 2. 도킹 상호작용
인산가수분해효소는 매우 특이적으로 기질의 특정 부위를 탈인산화시킬 수 있다. 이러한 기질 인식을 조절하는 메커니즘과 규칙, 즉 "인산가수분해효소 코드"를 밝히려는 연구가 진행 중이며, 9종의 진핵생물 유전체에 암호화된 모든 단백질 인산가수분해효소에 대한 비교 분석이 이루어졌다.[40][6][23]
연구에 따르면 "도킹 상호작용"이 기질 결합에 중요한 역할을 한다.[37][3][20] 인산가수분해효소는 기질에 있는 다양한 모티프(2차 구조의 요소)를 인식하고 상호작용한다. 이 모티프들은 효소의 활성 부위가 아닌 다른 도킹 부위에 낮은 친화도로 결합한다. 각각의 개별 도킹 상호작용은 약하지만, 여러 상호작용이 동시에 일어나면서 결합 특이성에 누적적인 효과를 부여한다.[41][7][24] 또한, 도킹 상호작용은 인산가수분해효소를 알로스테릭 조절하여 촉매 활성에 영향을 미칠 수도 있다.[42][8][25]
3. 기능
3. 1. 단백질 인산가수분해효소
단백질 인산가수분해효소는 효소의 일종으로, 단백질 기질의 아미노산 잔기에 붙어 있는 인산기를 제거하여 탈인산화시키는 역할을 한다. 이는 단백질을 인산화하여 신호 전달 분자로 만드는 단백질 키네이스와는 반대되는 작용이다. 인산가수분해효소에 의한 인산기 제거는 세포 내 신호전달 시스템이 다음 신호를 받아들일 수 있도록 초기 상태로 되돌리는 데 필수적이다.
단백질 키네이스와 인산가수분해효소의 연계 작용은 세포의 기능을 조절하는 네트워크에서 중요한 부분을 차지한다.[51][17][34] 단백질의 인산화와 탈인산화는 번역 후 변형 중 가장 흔하게 일어나는 방식 중 하나로, 특정 시점에서는 세포 내 전체 단백질의 최대 30%까지 인산화된 상태일 것으로 추정된다.[43][44][9][10][26][27]
주요 단백질 인산가수분해효소로는 단백질 인산가수분해효소 2(PP2A)와 PP2B가 있다. PP2A는 DNA 복제, 물질대사, 전사, 발생과 같은 다양한 세포 조절 과정에 관여한다. PP2B는 칼시뉴린이라는 이름으로도 알려져 있으며, 특히 T 세포의 증식 과정에 중요한 역할을 한다. 이러한 기능 때문에 칼시뉴린은 면역계의 활동을 억제하려는 면역 억제제 개발의 주요 표적이 되고 있다.[51][17][34]
단백질 인산가수분해효소는 키네이스와 비교했을 때 종류는 더 적지만, 다양한 기질을 인식하고 반응을 촉매하는 경향이 있다. 예를 들어, 사람의 경우 세린/트레오닌 특이적 단백질 키네이스의 종류가 세린/트레오닌 인산가수분해효소보다 약 10배 더 많다.[21] 이러한 불균형은 아직 인산가수분해효소에 대한 연구가 부족하여, 세포나 조직 내에서 어떤 인산가수분해효소들이 존재하는지(인산가수분해효소체, phosphatome) 완전히 파악되지 않았기 때문일 수 있다.[20] 아직 발견되지 않은 인산가수분해효소가 많으며, 발견된 것들 중에서도 어떤 기질에 작용하는지 밝혀지지 않은 경우가 많다. 그러나 연구가 진행된 일부 키네이스-인산가수분해효소 쌍에서는, 인산가수분해효소가 상대 키네이스보다 구조적, 기능적으로 더 다양한 특징을 보이는 것으로 나타났다. 이는 인산가수분해효소 자체의 불안정성 때문일 수도 있다고 여겨진다.[21]
3. 2. 뉴클레오타이드가수분해효소
뉴클레오타이드가수분해효소는 뉴클레오타이드의 가수분해를 촉매하여 뉴클레오사이드와 인산 이온을 생성하는 효소이다.[45][11][28] 이 효소는 뉴클레오타이드와 뉴클레오사이드의 균형을 유지하는 데 기여하여 세포의 항상성 유지에 필수적이다.[46][12][29]
일부 뉴클레오타이드가수분해효소는 세포 외부에서 작용하며, 세포로 운반되어 뉴클레오타이드 회수 경로를 통해 뉴클레오타이드를 재생성하는 데 사용될 수 있는 뉴클레오사이드를 생성한다.[47][13][30] 세포 내부에서는 스트레스 조건 하에서 에너지 수준을 유지하는 데 도움을 줄 수 있다. 예를 들어, 산소나 영양분이 부족한 세포는 세포의 주요 에너지원인 ATP와 같은 뉴클레오사이드 삼인산의 수준을 높이기 위해 더 많은 뉴클레오타이드를 분해할 수 있다.[48][14][31]
3. 3. 포도당신생합성에서의 역할
인산가수분해효소는 포도당신생합성의 대사 중간생성물과 같은 탄수화물에도 작용할 수 있다. 포도당신생합성은 비탄수화물 전구체로부터 포도당을 생성하는 생합성 경로이다. 많은 조직이 포도당만을 에너지원으로 사용하기 때문에 이 경로는 필수적이다.[51][17][34] 포도당신생합성 과정에서 중요한 역할을 하는 인산가수분해효소로는 포도당 6-인산가수분해효소와 과당 1,6-이중인산가수분해효소가 있다. 이 두 효소는 포도당신생합성의 비가역적인 단계를 촉매하며,[49][50][15][16][32][33] 각각 6탄소 당인산 중간체에서 인산기를 제거하는 역할을 수행한다.
4. 분류
국제 생화학·분자생물학 연합(IUBMB)의 효소 위원회는 인산가수분해효소를 기질 특이성과 촉매 도메인에서의 sequence homology|서열 상동성eng을 기준으로 104개의 뚜렷한 효소 계열(또는 군)로 분류한다.[37][3][20] 이렇게 100개가 넘는 계열로 분류됨에도 불구하고, 모든 인산가수분해효소는 기본적으로 동일한 일반적인 가수분해 반응을 촉매한다.[35][1][18]
하지만 인산가수분해효소의 분류는 명확하게 구분되기 어려운 측면이 있다. 생체 외(in vitro) 실험에서는 인산가수분해효소가 여러 종류의 기질을 인식하는 것처럼 보이지만, 생체 내(in vivo)에서는 특정 기질에 대해 매우 높은 특이성을 나타내는 경우가 많다.[37][3][20] 또한, 특정 기질(예: 단백질)을 기준으로 정의된 단백질 인산가수분해효소가 다른 종류의 기질(예: 비단백질)의 탈인산화를 촉매하기도 한다.[38][4][21] 예를 들어, 단백질 티로신 인산가수분해효소 중 일부는 티로신 잔기뿐만 아니라 세린 잔기도 탈인산화시킬 수 있다. 이처럼 하나의 인산가수분해효소가 여러 계열의 특성을 동시에 나타낼 수 있어 분류 체계와 실제 효소의 작용 방식 사이에 차이가 있을 수 있다.[51][17][34]
참조
[1]
웹사이트
ENZYME: 3.1.3.-
http://enzyme.expasy[...]
2017-02-21
[2]
논문
HuPho: the human phosphatase portal
https://art.torverga[...]
2013-01-01
[3]
논문
The human phosphatase interactome: An intricate family portrait
2012-08-14
[4]
논문
Elucidating Human Phosphatase-Substrate Networks
2013-05-14
[5]
논문
Phosphoproteomic Analysis Reveals Interconnected System-Wide Responses to Perturbations of Kinases and Phosphatases in Yeast
2010-12-21
[6]
논문
Genomics and evolution of protein phosphatases
2017-04-11
[7]
논문
Cracking the Phosphatase Code: Docking Interactions Determine Substrate Specificity
2009-12-08
[8]
논문
Docking interactions in protein kinase and phosphatase networks
2006-12-01
[9]
논문
The origins of protein phosphorylation
2002-05-01
[10]
논문
Protein tyrosine phosphatases: from genes, to function, to disease
[11]
웹사이트
ENZYME entry 3.1.3.31
http://enzyme.expasy[...]
2017-03-21
[12]
논문
Interrelations between substrate cycles and de novo synthesis of pyrimidine deoxyribonucleoside triphosphates in 3T6 cells
[13]
논문
Cellular function and molecular structure of ecto-nucleotidases
2012-09-01
[14]
논문
The 5'-nucleotidases as regulators of nucleotide and drug metabolism
2005-07-01
[15]
웹사이트
ENZYME entry 3.1.3.9
http://enzyme.expasy[...]
2017-03-21
[16]
웹사이트
ENZYME entry 3.1.3.11
http://enzyme.expasy[...]
2017-03-21
[17]
서적
Fundamentals of biochemistry : life at the molecular level
Wiley
2013-01-01
[18]
웹사이트
ENZYME: 3.1.3.-
http://enzyme.expasy[...]
2017-02-21
[19]
논문
HuPho: the human phosphatase portal
http://onlinelibrary[...]
2013-01-01
[20]
논문
The human phosphatase interactome: An intricate family portrait
2012-08-14
[21]
논문
Elucidating Human Phosphatase-Substrate Networks
http://stke.sciencem[...]
2013-05-14
[22]
논문
Phosphoproteomic Analysis Reveals Interconnected System-Wide Responses to Perturbations of Kinases and Phosphatases in Yeast
http://stke.sciencem[...]
2010-12-21
[23]
논문
Genomics and evolution of protein phosphatases
http://stke.sciencem[...]
2017-04-11
[24]
논문
Cracking the Phosphatase Code: Docking Interactions Determine Substrate Specificity
http://stke.sciencem[...]
2009-12-08
[25]
논문
Docking interactions in protein kinase and phosphatase networks
http://www.sciencedi[...]
2006-12-01
[26]
논문
The origins of protein phosphorylation
2002-05-01
[27]
논문
Protein tyrosine phosphatases: from genes, to function, to disease
http://www.nature.co[...]
[28]
웹사이트
ENZYME entry 3.1.3.31
http://enzyme.expasy[...]
2017-03-21
[29]
논문
Interrelations between substrate cycles and de novo synthesis of pyrimidine deoxyribonucleoside triphosphates in 3T6 cells
[30]
논문
Cellular function and molecular structure of ecto-nucleotidases
https://link.springe[...]
2012-09-01
[31]
논문
The 5'-nucleotidases as regulators of nucleotide and drug metabolism
2005-07-01
[32]
웹사이트
ENZYME entry 3.1.3.9
http://enzyme.expasy[...]
2017-03-21
[33]
웹사이트
ENZYME entry 3.1.3.11
http://enzyme.expasy[...]
2017-03-21
[34]
서적
Fundamentals of biochemistry : life at the molecular level
https://www.worldcat[...]
Wiley
2013-01-01
[35]
웹인용
ENZYME: 3.1.3.-
http://enzyme.expasy[...]
2017-02-21
[36]
저널
HuPho: the human phosphatase portal
https://art.torverga[...]
2013-01-01
[37]
저널
The human phosphatase interactome: An intricate family portrait
2012-08-14
[38]
저널
Elucidating Human Phosphatase-Substrate Networks
2013-05-14
[39]
저널
Phosphoproteomic Analysis Reveals Interconnected System-Wide Responses to Perturbations of Kinases and Phosphatases in Yeast
2010-12-21
[40]
저널
Genomics and evolution of protein phosphatases
2017-04-11
[41]
저널
Cracking the Phosphatase Code: Docking Interactions Determine Substrate Specificity
2009-12-08
[42]
저널
Docking interactions in protein kinase and phosphatase networks
2006-12-01
[43]
저널
The origins of protein phosphorylation
2002-05-01
[44]
저널
Protein tyrosine phosphatases: from genes, to function, to disease
[45]
웹인용
ENZYME entry 3.1.3.31
http://enzyme.expasy[...]
2017-03-21
[46]
저널
Interrelations between substrate cycles and de novo synthesis of pyrimidine deoxyribonucleoside triphosphates in 3T6 cells
[47]
저널
Cellular function and molecular structure of ecto-nucleotidases
2012-09-01
[48]
저널
The 5'-nucleotidases as regulators of nucleotide and drug metabolism
2005-07-01
[49]
웹인용
ENZYME entry 3.1.3.9
http://enzyme.expasy[...]
2017-03-21
[50]
웹인용
ENZYME entry 3.1.3.11
http://enzyme.expasy[...]
2017-03-21
[51]
서적
Fundamentals of biochemistry : life at the molecular level
Wiley
2013-01-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com