포도당신생합성
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
포도당신생합성은 젖산, 글리세롤, 아미노산 등 전구체로부터 포도당을 생성하는 대사 경로이다. 주요 전구체로는 젖산, 글리세롤, 알라닌, 글루타민 등이 있으며, 반추동물에서는 프로피온산이 주요 기질로 사용된다. 포유류에서는 간, 콩팥, 장, 뇌 등에서 일어나며, 호르몬, 기질 가용성, 알로스테릭 조절을 통해 조절된다. 글루카곤은 포도당신생합성을 촉진하고, 인슐린은 억제하며, 인슐린 저항성은 포도당신생합성을 증가시켜 고혈당증을 유발할 수 있다. 포도당신생합성은 가장 오래된 대사 경로 중 하나로 간주되며, 초기 생명체에서 해당 작용과 함께 기원했을 가능성이 있다.
더 읽어볼만한 페이지
- 당생물학 - 펩티도글리칸
펩티도글리칸은 세균 세포벽의 골격 구조를 이루는 주요 성분으로, N-아세틸글루코사민과 N-아세틸무라민산이 반복된 다당사슬에 펩티드 곁사슬이 붙은 형태이며, 세균의 형태 유지, 삼투압 저항성 제공, 면역 반응 유도, 항생제 표적 등의 역할을 한다. - 당생물학 - 글리코젠
글리코젠은 포도당 잔기가 α(1→4) 결합으로 연결된 분지형 생체고분자이며, 간과 근육에 저장되어 에너지원으로 사용되고, 혈당 조절 및 근육 수축에 기여하며, 대사 장애는 다양한 질환과 관련된다. - 대사경로 - 스테로이드
스테로이드는 4개의 융합된 고리 구조를 가지는 유기 화합물로, 콜레스테롤, 스테로이드 호르몬, 담즙산 등 생물학적 활성 분자를 포함하며, 메발론산 경로를 통해 생합성되어 다양한 생리적 기능을 수행하고 의학적으로 사용되지만 오남용 시 부작용을 초래할 수 있다. - 대사경로 - 시트르산 회로
시트르산 회로는 아세틸-CoA를 이산화탄소로 산화시켜 NADH와 FADH₂를 생성, ATP 생산 및 생합성에 기여하며, 탄수화물, 지방, 단백질 대사를 연결하는 중심적인 대사 경로이다. - 당뇨병 - 인슐린 저항성
인슐린 저항성은 인슐린이 제 기능을 못하여 혈당 수치가 상승하는 상태로, 고인슐린혈증을 유발하고 2형 당뇨병과 같은 대사 질환으로 이어질 수 있으며, 유전적 요인, 비만, 운동 부족 등이 주요 원인이고 생활 습관 개선과 약물 치료로 관리한다. - 당뇨병 - 인슐린
인슐린은 췌장에서 생성되는 단백질 호르몬으로 혈당 조절, 단백질 합성, 세포 성장에 관여하며, 당뇨병 치료에 사용되고 지속적인 연구가 진행되고 있다.
| 포도당신생합성 | |
|---|---|
| 개요 | |
| 명칭 | 포도당신생성 |
| 영어 명칭 | Gluconeogenesis |
| 설명 | 포도당이 아닌 탄수화물 전구체로부터 포도당을 합성하는 대사 경로 |
| 상세 정보 | |
| 기질 | 피루브산 젖산 글리세롤 아미노산 (글루코겐성) |
| 최종 산물 | 포도당 |
| 위치 | 간 신장 (덜 중요) |
| 관련 경로 | 해당과정 (역반응의 일부) 코리 회로 |
| 조절 | 호르몬 (글루카곤, 인슐린) 기질 농도 효소 활성 조절 |
| 중요성 | 혈당 유지, 특히 기아 상태에서 중요 |
| 임상적 중요성 | 제2형 당뇨병과 관련 |
2. 전구체
사람에서 포도당신생합성에 사용되는 주요 전구체는 젖산, 글리세롤(트라이글리세라이드 분자의 일부분), 포도당생성성 아미노산(특히 알라닌과 글루타민)이다. 이들은 전체 포도당신생합성의 90% 이상을 차지한다.[52] 그 외에 시트르산 회로의 모든 대사 중간생성물들(옥살로아세트산으로 전환될 수 있는)도 포도당신생합성을 위한 기질로 작용할 수 있다.[46]
반추동물에서는 프로피온산이 주요 포도당신생합성 기질이다.[53][54] 사람을 포함한 비반추동물에서 프로피온산은 홀수 지방산 및 가지사슬 지방산의 β 산화에서 생성되지만, 포도당신생합성에 대한 기여도는 비교적 미미하다.[55][56]
일반적으로 사람이 음식물에서 포도당신생합성 기질을 섭취해도 포도당신생합성이 증가하지는 않는다.[69]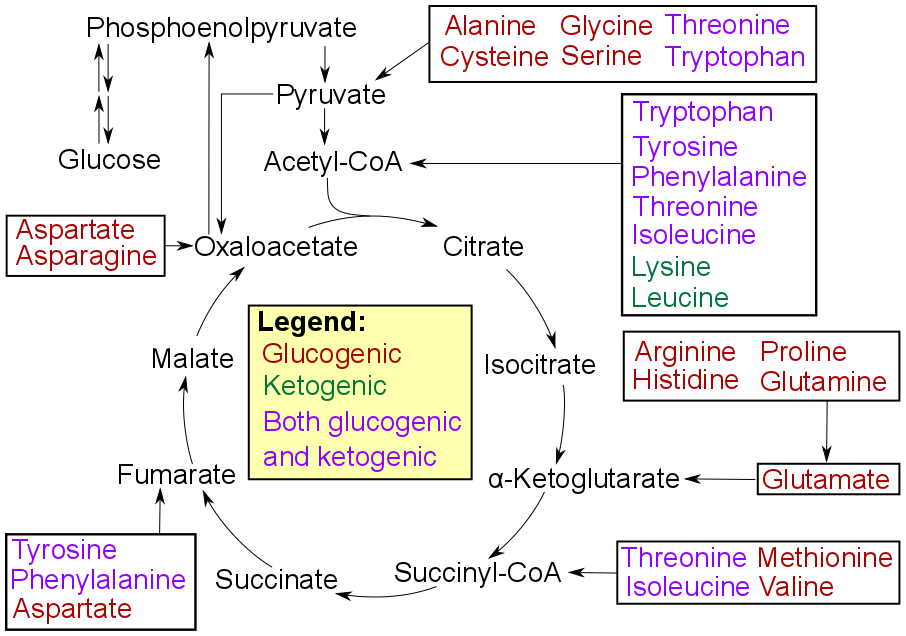
젖산, 글리세롤, 아미노산, 프로피온산 외에 예외적으로 지방산이 포도당신생합성에 기여하는 경우가 있다. 자세한 내용은 하위 섹션을 참고한다.
2. 1. 젖산
젖산은 근육 등에서 혐기성 해당과정의 결과로 생성되는 물질이다. 이렇게 생성된 젖산은 코리 회로를 통해 간으로 운반되어 젖산 탈수소 효소에 의해 피루브산으로 전환된다.[12] 피루브산은 포도당신생합성의 첫 번째 기질로 사용되어 포도당을 생성하는데 사용된다.[12]단식 기간이 길어질수록 코리 회로를 통한 젖산의 포도당 생성 기여도는 증가한다.[13] 예를 들어, 12시간, 20시간, 40시간 단식한 사람의 경우, 코리 회로를 통한 젖산의 포도당신생합성 기여도는 각각 41%, 71%, 92%였다.[13]
2. 2. 글리세롤
트라이글리세라이드의 일부인 글리세롤은 사람에게서 주요 포도당신생합성 전구체 중 하나이다.[7] 글리세롤은 간에서 글리세롤 3-인산으로, 이어서 다이하이드록시아세톤 인산으로 변환되어 포도당신생합성 경로에 진입한다.2. 3. 아미노산
대부분의 아미노산은 분해되어 시트르산 회로의 중간 대사물이나 피루브산으로 전환될 수 있으며, 이를 통해 포도당신생합성에 기여한다.[46] 특히 알라닌과 글루타민은 사람에서 주요 포도당신생합성성 전구체로, 전체 포도당신생합성의 90% 이상을 차지한다.[52] 다른 포도당생성성 아미노산들도 옥살로아세트산으로 전환되어 포도당신생합성의 기질로 작용할 수 있다.[1]아미노산의 아미노기 전이 또는 탈아미노화를 통해 탄소 골격이 직접(피루브산 또는 옥살로아세트산으로) 또는 시트르산 회로를 통해 간접적으로 포도당신생합성 경로로 들어간다.[12]
단백질 섭취 시 인슐린과 글루카곤이 동시에 분비되어 포도당신생합성이 활성화된다. 육식에 편중된 식단에서는 섭취 영양소가 단백질과 지방에 편중되어 당분 섭취가 부족하게 된다. 고양이와 같은 육식 동물은 포도당신생합성 효소 활성이 낮아 단백질에서 분해된 당 생성 아미노산을 포도당신생합성에 사용하지 않고 에너지원으로 이용한다. 반면, 개와 같은 잡식 동물이나 사람은 포도당신생합성 효소 활성이 높아 단백질에서 분해된 당 생성 아미노산으로부터 포도당신생합성을 수행하여 체내에서 필요한 당분을 생성한다.[41]
2. 4. 프로피온산
반추동물에서 프로피오네이트는 주요 포도당신생합성 기질이다.[8][9] 사람을 포함한 비반추 동물에서 프로피오네이트는 홀수 사슬 및 가지 사슬 지방산의 β 산화로부터 발생하며 포도당신생합성에 대한 (상대적으로 미미한) 기질이다.[10][11]프로피온산으로부터 포도당신생합성은 다음 경로를 거친다. 이 반응의 전반부는 홀수 사슬 지방산의 β 산화로부터 발생한 프로피온산의 대사 경로를 따른다.[44]
프로피온산 → 프로피오닐-CoA → S-메틸말로닐-CoA → R-메틸말로닐-CoA → 석시닐-CoA → 호박산 → 푸마르산 → 사과산 → 옥살아세트산 → 포스포엔올피루브산
2. 5. 지방산 (예외적인 경우)
사람은 글리옥실산 회로를 가지고 있지 않기 때문에, 지방산은 직접적으로 포도당으로 전환될 수 없다는 것이 일반적인 견해이다.[59] 짝수 지방산은 산화 과정을 통해 아세틸-CoA만을 생성하는데, 이 아세틸-CoA가 포도당신생합성에 참여하려면 글리옥실산 회로가 필요하지만 사람에게는 존재하지 않기 때문이다.[46]그러나 케톤증 상태에서는 지방산에서 유래한 아세틸-CoA가 아세톤을 포함한 케톤체를 생성하며, 생성된 아세톤의 최대 60%가 간에서 피루브산의 전구물질인 아세톨과 메틸글리옥살로 산화될 수 있다.[63][48] 이러한 과정을 통해 지방산에서 유래한 케톤체는 기아 상태에서 포도당신생합성의 최대 11%까지 기여할 수 있다. 더불어, 지방산의 이화작용은 포도당신생합성에 필요한 ATP 형태의 에너지를 생성한다.
3. 위치
포유류에서 포도당신생합성은 간[64], 콩팥[64], 장[65], 근육[66]에서 일어나는 것으로 알려져 왔으나, 최근에는 뇌의 별세포(별아교세포)[67]에서도 일어난다는 증거가 나타났다. 이들 기관은 다소 다른 포도당신생합성 전구체를 사용한다. 간은 주로 젖산, 글리세롤, 포도당생성성 아미노산(특히 알라닌)을 사용하고, 콩팥은 주로 젖산, 글루타민, 글리세롤을 사용한다.[68][52] 코리 회로의 젖산은 특히 콩팥에서 포도당신생합성을 위한 가장 큰 기질 공급원이다.[52] 간은 글리코젠 분해와 포도당신생합성을 모두 사용하여 포도당을 생성하지만, 콩팥은 포도당신생합성만 사용한다.[52] 장은 주로 글루타민과 글리세롤을 사용한다.[46]
대부분의 세포에서는 포도당 6-인산(글루코스-6-인산)의 인산기를 제거하여 유리 포도당을 생성할 수 있는 효소인 글루코스-6-포스파테이스가 없기 때문에 유리 포도당을 생성할 수 없다. 그러나 간세포와 소장, 콩팥 세포는 이 효소를 가지고 있어 포도당을 혈액으로 방출할 수 있다.
4. 경로
포도당신생합성은 11가지 효소 촉매 반응으로 구성된 대사 경로이다. 이 경로는 간이나 콩팥에서 시작되며, 사용되는 기질에 따라 세포의 미토콘드리아 또는 세포질에서 일어난다. 포도당신생합성을 구성하는 반응의 대부분은 해당과정의 역반응과 유사하다. 그러나 해당과정의 비가역적 반응 단계에서는 다른 효소를 사용하여 우회 경로를 거친다.[43]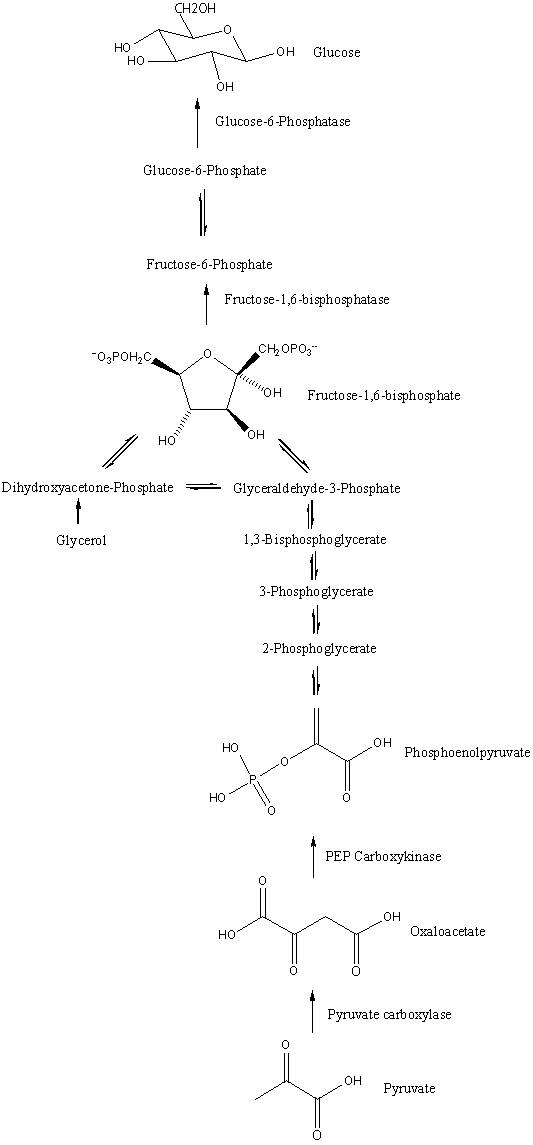
- 피루브산 → 옥살로아세트산: 포도당신생합성은 미토콘드리아에서 피루브산이 피루브산 카복실화효소에 의해 옥살로아세트산으로 전환되면서 시작된다. 이 반응에는 ATP 1분자가 필요하며, 아세틸-CoA 농도가 높으면 촉진되고, ADP와 포도당 농도가 높으면 억제된다.
- 옥살로아세트산 → 말산: 옥살로아세트산은 NADH를 사용하여 말산으로 환원되어 미토콘드리아 밖으로 운반된다.
- 말산 → 옥살로아세트산: 세포질로 이동한 말산은 NAD+를 사용하여 다시 옥살로아세트산으로 산화된다.
- 옥살로아세트산 → 포스포엔올피루브산: 옥살로아세트산은 포스포엔올피루브산 카복시키네이스에 의해 탈카복실화 및 인산화되어 포스포엔올피루브산으로 전환된다. 이 과정에서 GTP 1분자가 GDP로 가수분해된다.
- 포스포엔올피루브산 → 과당 6-인산: 이후 단계는 해당과정의 역반응과 동일하게 진행되지만, 과당 1,6-이중인산은 과당 1,6-이중인산가수분해효소에 의해 과당 6-인산으로 전환된다. 이 과정에서 물 1분자가 사용되고 인산 1분자가 방출된다. 해당과정에서는 포스포프럭토키네이스-1이 과당 6-인산과 ATP를 과당 1,6-이중인산과 ADP로 전환하는 반응을 수행한다. 과당 1,6-이중인산가수분해효소에 의한 반응은 포도당신생합성의 속도 제한 단계이다.
- 과당 6-인산 → 포도당 6-인산: 포도당 6-인산은 포스포헥소스 이성질화효소에 의해 과당 6-인산으로부터 생성된다. (해당 과정의 두 번째 단계의 역반응). 포도당 6-인산은 다른 대사 경로에 사용되거나 탈인산화되어 포도당으로 전환될 수 있다.
- 포도당 6-인산 → 포도당: 포도당신생합성의 마지막 단계는 소포체 내강에서 일어난다. 포도당 6-인산은 포도당 6-인산가수분해효소에 의해 가수분해되어 포도당과 무기 인산을 생성한다. 해당과정에서는 헥소키네이스가 포도당과 ATP를 포도당 6-인산과 ADP로 전환한다. 생성된 포도당은 포도당 운반체를 통해 세포질로 운반된다.
세포는 인산화된 형태(포도당 6-인산)를 세포 내에 가두고, 포도당은 세포 안팎으로 쉽게 확산될 수 있도록 하여 세포 내의 포도당 수준을 조절한다.
4. 1. 피루브산에서 포스포엔올피루브산으로의 전환
피루브산은 먼저 미토콘드리아 내에서 피루브산 카복실화효소에 의해 옥살로아세트산으로 전환된다. 이 반응에는 ATP 1분자가 소모되며, 아세틸-CoA에 의해 활성화되고 ADP와 포도당에 의해 억제된다.[1] 옥살로아세트산은 미토콘드리아 내막을 통과할 수 없으므로, 말산 또는 아스파르트산 형태로 미토콘드리아에서 세포질로 이동한다.[1]미토콘드리아에서 세포질로의 포스포에놀피루브산 수송은 말산-아스파트산 셔틀에 의해 조절된다. 옥살로아세트산은 미토콘드리아에서 말산 탈수소효소에 의해 말산으로 환원되며, 이 과정에서 NADH가 사용된다. 말산은 미토콘드리아 내막을 통과하여 세포질로 이동한 후, 세포질에서 다시 말산 탈수소효소에 의해 옥살로아세트산으로 산화된다. 이 과정에서 NAD+가 사용된다.[1]
세포질에 도달한 옥살로아세트산은 포스포엔올피루브산 카복시키네이스(PEPCK)에 의해 포스포엔올피루브산(PEP)으로 전환된다. 이 반응에서 GTP 1분자가 GDP로 가수분해된다.[1]
전체 반응 과정은 다음과 같다.[1]
- (미토콘드리아 내)
단계 1: 피루브산 + HCO3- + ATP → 옥살로아세트산 + ADP + Pi (피루브산 카복실화효소)
단계 2: 옥살로아세트산 + NADH + H+ L-말산 + NAD (말산 탈수소효소)
- (말산-아스파트산 셔틀에 의해 말산 형태로 미토콘드리아 내막, 외막을 통과하여 세포질에 도달)
단계 3: L-말산 + NAD 옥살로아세트산 + NADH + H+ (말산 탈수소효소)
단계 4: 옥살로아세트산 + GTP → 포스포엔올피루브산 + GDP + CO2 (포스포엔올피루브산 카복시키네이스)
이 반응의 깁스 자유 에너지 총합은 ⊿G'°=0.9 kj/mol이다.[1]
4. 2. 과당 1,6-이중인산에서 과당 6-인산으로의 전환
과당 1,6-이중인산가수분해효소는 과당 1,6-이중인산을 과당 6-인산으로 전환시키며, 이 과정에서 물 1분자를 사용하고 인산 1분자를 방출한다. 이 반응은 해당과정에서 포스포프럭토키네이스-1이 과당 6-인산과 ATP를 과당 1,6-이중인산과 ADP로 전환시키는 반응의 역반응이다. 이 단계는 포도당신생합성의 속도 제한 단계이기도 하다.해당과정에서는 포스포프럭토키네이스가 이 반응을 수행하지만, 이는 비가역적이므로 포도당신생합성에서는 과당 1,6-이중인산가수분해효소(FBPase)가 사용된다.[1] 이 효소는 Mg2+을 필요로 하며,[1] 과당 1,6-이중인산의 C-1 위치의 인산기를 가수분해한다.[1]
: 과당 1,6-이중인산 + H2O → 과당 6-인산 + Pi (⊿G'°=-16.3 kj/mol)
4. 3. 포도당 6-인산에서 포도당으로의 전환
포도당신생합성의 마지막 단계인 포도당 형성은 소포체의 내강에서 일어난다. 포도당 6-인산은 포도당 6-인산가수분해효소에 의해 가수분해되어 포도당과 무기 인산을 생성한다. 이는 헥소키네이스가 포도당과 ATP를 포도당 6-인산과 ADP로 전환하는 해당과정의 반응과 다르다. 포도당은 소포체 막에 위치한 포도당 운반체에 의해 세포질로 운반된다.[1]포도당 6-인산가수분해효소는 포도당 6-인산을 인산 에스터로 단순 가수분해를 촉진한다.[1]
: 포도당 6-인산 + H2O → 포도당 + Pi (⊿G'°=-13.8 kj/mol)[1]
5. 조절
포도당신생합성은 해당과정의 역반응과 유사하지만, 몇 가지 중요한 차이점이 존재한다. 해당과정의 헥소키네이스/글루코키네이스, 포스포프럭토키네이스-1, 피루브산 키네이스는 포도당신생합성에서 각각 포도당 6-인산가수분해효소, 과당 1,6-이중인산가수분해효소, 포스포엔올피루브산 카복시키네이스/피루브산 카복실화효소로 대체된다. 이 효소들은 서로 비슷한 분자에 의해 조절되지만, 정반대의 효과를 나타낸다.[73]
포도당신생합성에 관여하는 대부분의 효소는 세포질에서 발견되지만, 미토콘드리아의 피루브산 카복실화효소와 동물의 포스포엔올피루브산 카복시키네이스는 예외이다. 포스포엔올피루브산 카복시키네이스는 미토콘드리아와 세포질 모두에 존재하는 동질효소로 존재한다.[74]
5. 1. 호르몬 조절
글루카곤은 cAMP 조절 단백질 키네이스 A에 의한 효소 및 조절 단백질의 인산화를 유발하여 해당과정을 억제하고 포도당신생합성을 촉진한다.[75] 인슐린은 포도당신생합성을 억제함으로써 글루카곤에 길항적으로 작용한다.제2형 당뇨병은 과도한 글루카곤 분비와 인슐린 저항성을 특징으로 한다.[75] 인슐린은 더 이상 포스포엔올피루브산 카복시키네이스와 같은 효소의 유전자 발현을 억제할 수 없게 되어, 신체의 고혈당증을 유발한다.[76] 항당뇨병제인 메트포르민은 주로 포도당신생합성을 억제하여 혈당을 감소시키고, 인슐린 저항성으로 인한 인슐린의 포도당신생합성 억제 실패를 극복한다.[77]
단백질 섭취 시에는 인슐린과 글루카곤이 동시에 분비되지만, 포도당신생합성은 대폭 활성화된다. 건강한 사람에서는 인슐린이 다량 분비되어도 포도당신생합성은 크게 억제되지 않는다(간 글리코젠의 분해는 거의 완전히 멈춘다).[45] 반면, 인슐린이 고갈된 1형 당뇨병 환자의 경우에는 소량의 인슐린 주입으로 포도당신생합성이 크게 억제되었다는 보고가 있다.
5. 2. 기질 가용성 조절
포도당신생합성의 속도는 핵심 효소인 과당-1,6-이중인산가수분해효소의 작용에 의해 조절되며, 이 효소는 cAMP와 인산화를 통한 신호 전달을 통해서도 조절된다.[45]글루카곤은 단백질 키나아제 A를 통해 효소 및 조절 단백질의 인산화를 유발하여 해당과정을 억제하고 포도당신생합성을 자극한다. 인슐린은 포도당신생합성을 억제함으로써 글루카곤에 대응한다. 제2형 당뇨병은 과도한 글루카곤과 신체의 인슐린 저항성을 특징으로 한다. 항당뇨병 약물 메트포르민은 주로 포도당신생합성을 억제하여 혈당을 감소시킨다.[45]
연구에 따르면 간 포도당 생성의 부재는 공복 혈장 포도당 농도 조절에 큰 영향을 미치지 않는다. 글루카곤, 글루코코르티코이드, 산증에 의해 유도되는 포도당신생합성의 보상 유도가 신장과 창자에서 발생한다.[45]
건강한 사람에서는 인슐린이 다량 분비되어도 포도당신생합성은 크게 억제되지 않지만(간 글리코겐의 분해는 거의 완전히 멈춘다),[45] 단백질 섭취 시에는 인슐린과 글루카곤이 동시에 분비되어 포도당신생합성이 대폭 활성화된다. 인슐린이 고갈된 1형 당뇨병 환자의 경우에는 소량의 인슐린 주입으로 포도당신생합성이 크게 억제되었다는 보고가 있다.
5. 3. 알로스테릭 조절
아세틸-CoA와 시트르산은 포도당신생합성의 효소인 피루브산 카복실화효소 및 과당 1,6-이중인산가수분해효소를 활성화하는 반면, 해당과정의 효소인 피루브산 키네이스를 억제한다.[73] 이러한 상호 조절 시스템은 해당과정과 포도당신생합성이 서로 반대 작용을 하도록 조절하여, 포도당을 합성한 다음 분해만 하는 낭비 회로가 되는 것을 방지한다.6. 인슐린 저항성과 포도당신생합성
간에서 FOX 단백질 FOXO6은 일반적으로 공복 상태에서 포도당신생합성을 촉진하지만, 인슐린은 음식물 섭취 시 FOXO6을 차단한다.[79] 인슐린 저항성 상태에서 인슐린은 FOXO6을 차단하지 못하여 음식물을 섭취해도 계속 포도당신생합성을 일으켜 고혈당증을 초래한다.[79]
인슐린 저항성은 대사 증후군과 제2형 당뇨병의 일반적인 특징이다. 이러한 이유로 포도당신생합성은 포도당 생성을 억제하고 세포에 의한 포도당 흡수를 자극하는 항당뇨병제인 메트포르민과 같은 제2형 당뇨병에 대한 치료제들의 표적이다.[80]
7. 기원
포도당신생합성은 가장 오래된 동화 작용 경로 중 하나로 간주되며, 최후 보편 공통 조상에서 나타났을 가능성이 높다.[36] 라파엘 F. 세이(Rafael F. Say)와 게오르크 푹스(Georg Fuchs)는 2010년에 다음과 같이 말했다. "모든 고세균 그룹과 깊이 갈라진 세균 계통에는 FBP 알돌라아제와 FBP 포스파타아제 활성을 모두 가진 이중 기능의 과당 1,6-이중인산(FBP) 알돌라아제/포스파타아제가 포함되어 있다. 이 효소는 대부분의 다른 세균과 진핵생물에는 없으며, 중온성 해양 크렌고세균에서도 열 안정성을 보인다." 과당 1,6-이중인산 알돌라아제/포스파타아제가 조상 포도당신생합성 효소였으며, 해당 작용보다 앞섰을 것으로 제안되었다.[37]
8. 소모지 효과 (Somogyi effect)
클로드 베르나르가 밝힌 소모지 효과는 포도당신생합성이 급격하게 일어나 고혈당을 초래하는 현상이다.[42] 단식을 하면 당 부족을 보충하기 위해 근육이 분해되어 포도당신생합성이 일어나는데,[42] 이는 근육 감소 및 신진대사 감소로 이어진다.
참조
[1]
서적
Lehninger Principles of Biochemistry
https://archive.org/[...]
Worth Publishers
[2]
웹사이트
The Chemical Logic Behind Gluconeogenesis
http://www2.ufp.pt/~[...]
2009-09-08
[3]
논문
In silico evidence for gluconeogenesis from fatty acids in humans
2011-07
[4]
논문
Molecular signaling mechanisms of renal gluconeogenesis in nondiabetic and diabetic conditions
2019-06
[5]
서적
Harper's illustrated Biochemistry, 30th edition
McGraw Hill
[6]
서적
Biochemistry
Lippincott Williams & Wilkins
2004-08-01
[7]
논문
Renal gluconeogenesis: its importance in human glucose homeostasis
https://www.research[...]
2001-02
[8]
서적
Dukes' Physiology of Domestic Animals
Cornell Univ. Press
2004
[9]
서적
Nutritional Ecology of the Ruminant
Cornell Univ. Press
1994
[10]
서적
Harper's Illustrated Biochemistry
McGraw-Hill Publishing Company
2018
[11]
서적
Medical Biochemistry
Elsevier
2014
[12]
서적
Principles of Biochemistry with a Human Focus
Brooks/Cole, Thomson Learning
[13]
논문
Gluconeogenesis and the Cori cycle in 12-, 20-, and 40-h-fasted humans
http://ajpendo.physi[...]
2017-07-28
[14]
논문
Can sugars be produced from fatty acids? A test case for pathway analysis tools
2009-01
[15]
논문
Bifunctional glyoxylate cycle protein of Caenorhabditis elegans: a developmentally regulated protein of intestine and muscle
1995-06
[16]
논문
Evolution of glyoxylate cycle enzymes in Metazoa: evidence of multiple horizontal transfer events and pseudogene formation
2006-10
[17]
논문
Conversion of fatty acids to carbohydrate; application of isotopes to this problem and role of the Krebs cycle as a synthetic pathway
1957-04
[18]
논문
Plasma acetone metabolism in the fasting human
1979-04
[19]
서적
Vander's Human Physiology
https://archive.org/[...]
McGraw Hill
[20]
논문
A novel role for glucose 6-phosphatase in the small intestine in the control of glucose homeostasis
2004-10
[21]
논문
Gain of glucose-independent growth upon metastasis of breast cancer cells to the brain
2015-02
[22]
논문
Cerebral Gluconeogenesis and Diseases
[23]
논문
Role of the kidney in normal glucose homeostasis and in the hyperglycaemia of diabetes mellitus: therapeutic implications
2010-02
[24]
논문
Regulation of hepatic glucose production and the role of gluconeogenesis in humans: is the rate of gluconeogenesis constant?
2008-09
[25]
논문
Substrate utilization for hepatic gluconeogenesis is altered by increased glucose demand in ruminants
1999-07
[26]
논문
Insulin and glucagon regulation of gluconeogenesis in preruminating and ruminating bovine
1995-02
[27]
서적
Fundamentals of Biochemistry
https://archive.org/[...]
John Wiley & Sons Inc
[28]
간행물
From Glucose to Lactate and Transiting Intermediates Through Mitochondria, Bypassing Pyruvate Kinase: Considerations for Cells Exhibiting Dimeric PKM2 or Otherwise Inhibited Kinase Activity
https://www.frontier[...]
2020
[29]
논문
Factors that control the tissue-specific transcription of the gene for phosphoenolpyruvate carboxykinase-C
[30]
논문
Metformin and insulin suppress hepatic gluconeogenesis through phosphorylation of CREB binding protein
2009-05
[31]
논문
Insulin regulation of gluconeogenesis
2018-01
[32]
논문
Expression profile analysis of long non-coding RNAs involved in the metformin-inhibited gluconeogenesis of primary mouse hepatocytes
https://www.spandido[...]
2018-01
[33]
논문
Control of blood glucose in the absence of hepatic glucose production during prolonged fasting in mice: induction of renal and intestinal gluconeogenesis by glucagon
2011-12
[34]
논문
FoxO integration of insulin signaling with glucose and lipid metabolism
2017-05
[35]
논문
Mechanism by which metformin reduces glucose production in type 2 diabetes
2000-12
[36]
논문
Life as a guide to prebiotic nucleotide synthesis
2018-12
[37]
논문
Fructose 1,6-bisphosphate aldolase/phosphatase may be an ancestral gluconeogenic enzyme
2010-04
[38]
논문
Nonenzymatic Metabolic Reactions and Life's Origins
https://hal.archives[...]
2020-08
[39]
논문
Nonenzymatic gluconeogenesis-like formation of fructose 1,6-bisphosphate in ice
2017-07
[40]
논문
An appeal to magic? The discovery of a non-enzymatic metabolism and its role in the origins of life
2018-08-30
[41]
논문
Comparison of the activities of enzymes related to glycolysis and gluconeogenesis in the liver of dogs and cats
[42]
웹사이트
糖尿病シラバス
http://www.med.kyush[...]
九州大学大学院医学研究院 病態制御内科学(第三内科) 糖尿病研究室
2013-04-26
[43]
서적
"レーニンジャーの新生化学[上]第4版"
廣川書店
2006
[44]
간행물
糖新生
http://www.sc.fukuok[...]
[45]
뉴스
null
https://doi.org/10.1[...]
The Journal of Clinical Endocrinology & Metabolism
2019-07
[46]
서적
Lehninger Principles of Biochemistry
https://archive.org/[...]
Worth Publishers
[47]
웹인용
The Chemical Logic Behind Gluconeogenesis
http://www2.ufp.pt/~[...]
2009-09-08
[48]
논문
In silico evidence for gluconeogenesis from fatty acids in humans
2011-07
[49]
논문
Molecular signaling mechanisms of renal gluconeogenesis in nondiabetic and diabetic conditions
2019-06
[50]
서적
Harper's illustrated Biochemistry, 30th edition
McGraw Hill
[51]
서적
Biochemistry (Lippincott's Illustrated Reviews)
Lippincott Williams & Wilkins
2004-08-01
[52]
논문
Renal gluconeogenesis: its importance in human glucose homeostasis
https://www.research[...]
2001-02
[53]
서적
Dukes' Physiology of Domestic Animals
https://archive.org/[...]
Cornell Univ. Press
2004
[54]
서적
Nutritional Ecology of the Ruminant
Cornell Univ. Press
1994
[55]
서적
Harper's Illustrated Biochemistry
McGraw-Hill Publishing Company
2018
[56]
서적
Medical Biochemistry
Elsevier
2014
[57]
서적
Principles of Biochemistry with a Human Focus
https://archive.org/[...]
Brooks/Cole, Thomson Learning
[58]
논문
Gluconeogenesis and the Cori cycle in 12-, 20-, and 40-h-fasted humans
http://ajpendo.physi[...]
2023-04-13
[59]
논문
Can sugars be produced from fatty acids? A test case for pathway analysis tools
2009-01
[60]
논문
Bifunctional glyoxylate cycle protein of Caenorhabditis elegans: a developmentally regulated protein of intestine and muscle
1995-06
[61]
논문
Evolution of glyoxylate cycle enzymes in Metazoa: evidence of multiple horizontal transfer events and pseudogene formation
2006-10
[62]
논문
Conversion of fatty acids to carbohydrate; application of isotopes to this problem and role of the Krebs cycle as a synthetic pathway
1957-04
[63]
논문
Plasma acetone metabolism in the fasting human
1979-04
[64]
서적
Vander's Human Physiology
https://archive.org/[...]
McGraw Hill
[65]
논문
A novel role for glucose 6-phosphatase in the small intestine in the control of glucose homeostasis
2004-10
[66]
논문
Gain of Glucose-Independent Growth upon Metastasis of Breast Cancer Cells to the Brain
2015-02-01
[67]
논문
Cerebral Gluconeogenesis and Diseases
[68]
논문
Role of the kidney in normal glucose homeostasis and in the hyperglycaemia of diabetes mellitus: therapeutic implications
2010-02
[69]
논문
Regulation of hepatic glucose production and the role of gluconeogenesis in humans: is the rate of gluconeogenesis constant?
https://www.research[...]
2008-09
[70]
논문
Substrate utilization for hepatic gluconeogenesis is altered by increased glucose demand in ruminants
1999-07
[71]
저널
Insulin and glucagon regulation of gluconeogenesis in preruminating and ruminating bovine
1995-02
[72]
서적
Fundamentals of Biochemistry
https://archive.org/[...]
John Wiley & Sons Inc
[73]
문서
From Glucose to Lactate and Transiting Intermediates Through Mitochondria, Bypassing Pyruvate Kinase: Considerations for Cells Exhibiting Dimeric PKM2 or Otherwise Inhibited Kinase Activity
https://www.frontier[...]
2020
[74]
저널
Factors that control the tissue-specific transcription of the gene for phosphoenolpyruvate carboxykinase-C
[75]
저널
Metformin and insulin suppress hepatic gluconeogenesis through phosphorylation of CREB binding protein
2009-05
[76]
저널
Insulin regulation of gluconeogenesis
2018-01
[77]
저널
Expression profile analysis of long non-coding RNAs involved in the metformin-inhibited gluconeogenesis of primary mouse hepatocytes
https://www.spandido[...]
2018-01
[78]
저널
Control of blood glucose in the absence of hepatic glucose production during prolonged fasting in mice: induction of renal and intestinal gluconeogenesis by glucagon
2011-12
[79]
저널
FoxO integration of insulin signaling with glucose and lipid metabolism
2017-05
[80]
저널
Mechanism by which metformin reduces glucose production in type 2 diabetes
2000-12
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com