참문어
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
참문어(Octopus vulgaris)는 외투막 길이가 최대 25cm, 팔은 최대 1m까지 자라는 문어 종이다. 전 세계 열대, 아열대, 온대 해역에 널리 분포하며, 게, 가재, 이매패류 등을 먹고, 주변 환경에 맞춰 색을 바꾸는 특징이 있다. 5억 개의 뉴런을 가진 거대한 신경계와 뇌를 가지고 있어 지능적이며, 1~2년 정도 산다. 아가미를 통해 호흡하며, 세 개의 심장을 가지고 있다.
더 읽어볼만한 페이지
- 문어과 - 흡혈오징어
흡혈오징어는 피를 빨지 않고 심해 유기물을 먹는 두족류로, 문어와 오징어의 특징을 가지며, 큰 눈과 독특한 방어 전략을 사용하고, 오징어와 문어의 공통 조상 형태로 여겨진다. - 문어과 - 낙지속
낙지속은 문어과에 속하는 속으로, 팔뚝문어, 붉은돗문어 등 다양한 종을 포함하며, 한국에서는 긴다리문어와 동아시아 문어가 발견된다. - 조르주 퀴비에가 명명한 분류군 - 망둑어과
망둑어과는 전 세계 다양한 환경에 서식하며 좌우 배지느러미가 합쳐진 흡반과 독특한 번식 전략이 특징인 농어목의 저서어류로, 식용이나 관상어로 이용되기도 하며 일부 종은 보존 노력이 필요하다. - 조르주 퀴비에가 명명한 분류군 - 놀래기과
놀래기과는 베라과에 속하는 어류로, 전 세계 열대 및 온대 해역에 분포하며 돌출 가능한 입과 턱니, 두꺼운 입술이 특징이고 육식성이며, 한국에서는 식용, 낚시 대상, 청소 물고기로 이용되기도 한다.
| 참문어 - [생물]에 관한 문서 | |
|---|---|
| 일반 정보 | |
 | |
| 학명 | Octopus vulgaris |
| 명명자 | Cuvier, 1797 |
| 이명 | Octopus octopodia (Tryon, 1879) Octopus vulgaris (Lamarck, 1798) Octopus rugosus (Bosc, 1792) |
| 보존 상태 | LC (IUCN 3.1) |
| 분포 | Common octopus worldwide distribution |
| 한국어 이름 | 참문어 |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 연체동물문 |
| 강 | 두족강 |
| 아강 | 초형아강 |
| 목 | 문어목 |
| 과 | 문어과 |
| 속 | 낙지속 |
| 시간 척도 | |
| 화석 범위 | 플라이스토세 - 현재 |
| 추가 정보 | |
| IUCN 평가 | https://www.iucnredlist.org/species/162571/918906 |
| 위키 공용 | Octopus vulgaris |
| 위키 생물종 | Octopus vulgaris |
| 라틴어 | Octopus vulgaris |
2. 특징
참문어는 주로 해 질 녘에 사냥하며, 게, 가재, 이매패류(바지락)를 포함해 잡을 수 있는 거의 모든 것을 먹는다. 주변 환경에 맞춰 색깔을 바꾸고, 방심한 먹이를 덮치는 능력을 지녔다. 부리로 껍데기를 깨고 독으로 먹이를 제압한다.[7]
5억 개 가량의 뉴런이 있는 발달된 신경계와 뇌를 가졌으며, 이는 개와 비슷한 수준이다. 밝기, 미로, 사람을 구별하고 병뚜껑을 열거나 바닷가재 덫을 습격하는 등 높은 지능을 보인다.[8][9][10] 해양 식물, 조류, 조개껍데기, 돌을 모아 "정원"을 가꾸는 모습은 비틀즈의 노래 "Octopus' Garden" (1969년)에 영감을 주었다. 영국 동물(과학적 절차)법 1986에 의해 보호받는 최초의 무척추동물이며,[11] 훈련 실험을 통해 물체의 밝기, 크기, 모양, 수평/수직 방향 구별 능력이 입증되었다.
짝짓기 시 식인 풍습이 나타나기도 한다.[6] 아프리카 북서부 해안에서 대규모 저인망 어업으로 연간 2만 톤 이상 포획된다.[3]
2. 1. 형태
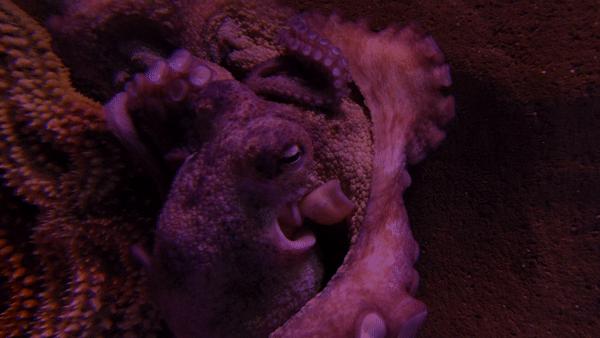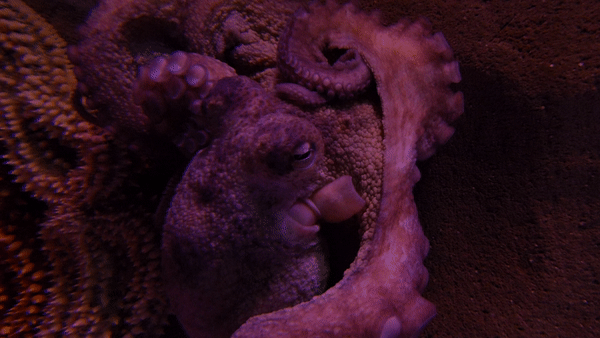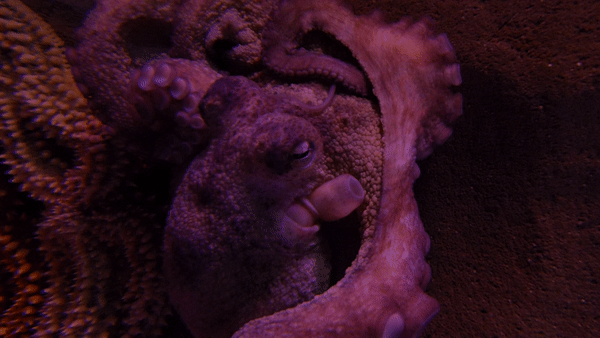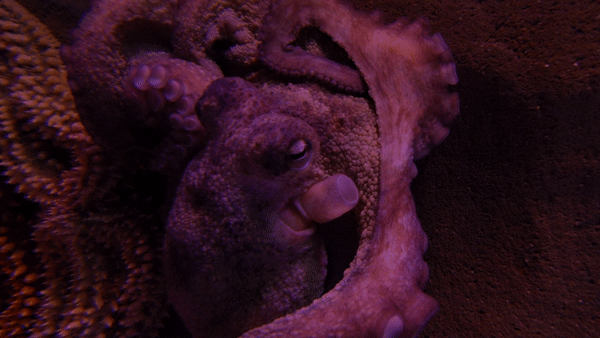



참문어(Octopus vulgarisla)는 외투막 길이가 25cm까지 자라며, 팔은 최대 1m까지 자란다.[3] 1~2년 동안 살며 최대 9.1kg까지 나갈 수 있다.[4][5]
2. 2. 생태
참문어(''Octopus vulgaris'')는 외투막 길이가 25cm, 팔은 최대 1m까지 자란다.[3] 1~2년 동안 살며 최대 약 9.07kg까지 나갈 수 있다.[4][5] 짝짓기는 식인 풍습이 될 수 있다.[6] 아프리카 북서부 해안에서 대규모 저인망 어업으로 잡히며, 연간 20000톤 이상이 수확된다.[3]
참문어는 해질녘에 사냥한다. 게, 가재, 이매패류(바지락)를 선호하지만, 잡을 수 있는 거의 모든 것을 먹는다. 주변 환경에 맞춰 색상을 변경할 수 있으며, 길을 잃은 조심성 없는 먹이를 덮칠 수 있다. 부리를 사용하여 껍질이 있는 연체동물의 껍질을 깨뜨릴 수 있다. 또한 먹이를 제압하기 위한 독을 가지고 있다.[7]
신경계와 뇌가 발달했으며, 약 5억 개의 뉴런을 가지고 있어 개와 비슷한 수준이다. 밝기 구별, 미로 탐색, 사람 인식, 병뚜껑 열기, 바닷가재 덫 습격 등을 할 수 있을 정도로 지능적이다.[8][9][10] 해양 식물, 조류, 조개껍데기, 돌을 수집하여 "정원"을 유지하는 모습이 관찰되었으며, 이는 1969년 비틀즈의 노래 "Octopus' Garden"의 영감이 되었을 수 있다. 영국 동물(과학적 절차)법 1986에 의해 보호받는 최초의 무척추동물이었다.[11] 훈련 실험을 통해 물체의 밝기, 크기, 모양, 수평 또는 수직 방향을 구별할 수 있음이 밝혀졌다.
2. 3. 지능
참문어는 거대한 신경계와 뇌를 가지고 진화했다. 참문어 개체는 약 5억 개의 뉴런을 가지고 있는데, 이는 개와 거의 비슷한 수준이다.[8] 참문어는 밝기를 구별하고, 미로를 탐색하며, 개별 사람을 인식하고, 병뚜껑을 풀거나 바닷가재 덫을 습격하는 방법을 배울 수 있을 정도로 지능적이다.[9][10] 또한 다양한 해양 식물과 조류, 조개껍데기 및 돌을 수집하여 "정원"을 유지하는 모습이 관찰되었으며, 이러한 행동은 1969년 비틀즈의 노래 옥토퍼스 가든에 영감을 주었을 수 있다. 참문어는 영국 동물(과학적 절차)법 1986에 의해 보호받는 최초의 무척추동물이었다.[11] 훈련 실험을 통해 참문어는 물체의 밝기, 크기, 모양, 수평 또는 수직 방향을 구별할 수 있음이 밝혀졌다.3. 생리학
참문어는 아가미를 통해 호흡하며, 얇은 피부를 통해서도 산소를 흡수한다. 아가미는 아가미 신경절과 접힌 새엽으로 구성되어 있으며, 물이 새엽 위로 이동하면서 산소가 혈액으로 확산된다.[22] 얇은 피부는 휴식 시 산소 흡수의 약 41%를 차지할 정도로 중요하다.[18] 물의 온도가 20°C일 때 최대 65%의 산소를 섭취할 수 있으며,[22] 헤모시아닌은 산소 섭취를 돕는 호흡 색소이다.[17]
참문어의 순환계는 세 개의 심장으로 구성된다. 몸 전체에 혈액을 공급하는 주 심장과 아가미에서 혈액 순환을 돕는 두 개의 아가미 심장이 있다.[26] 혈액은 구리가 풍부한 헤모시아닌으로 구성되어 산소를 운반하며,[32] 보어 효과에 따라 이산화탄소 농도와 혈액 pH에 따라 산소 방출 및 섭취가 조절된다.
참문어는 주변 해수와 등장성을 유지하며, 삼투 조절을 하지 않는 순응자이다.[36] 그러나 이온 농도는 해수와 차이가 있어 나트륨, 칼슘, 염화물 등의 농도를 낮게 유지하는데,[36] 이는 이온 조절 작용이 존재함을 시사한다.[36] 신장 시스템은 암모니아성 질소 화합물 및 기타 대사성 폐기물을 배설한다.[36]
참문어는 변온동물이자 외온동물로서 주변 온도에 따라 체온이 변한다.[40] 온도 변화에 따라 호흡 속도와 산소 소비량이 달라지며, 18°C에서 24°C 사이가 최적 온도이다.[41] 온도 변화는 혈림프 단백질 농도 변화를 유발하며, 헤모시아닌의 산소 친화도에 영향을 미친다.[42]
3. 1. 서식지 및 환경 요구
참문어는 전 세계 열대, 아열대 및 온대 해역에 널리 퍼져 산다.[12][13][14] 비교적 얕고 바위가 많은 연안 해역의 바닥을 좋아하며, 수심은 대개 200m를 넘지 않는다.[14] 전 세계 서식지의 염분은 대략 30g/L에서 45g/L 사이에서 발견되지만, 선호하는 염도는 약 36g/L이다.[15] 다양한 온도에 노출되지만, 선호하는 온도는 약 15°C에서 16°C 범위이다.[15] 특히 더운 계절에는 따뜻한 물층을 피하기 위해 평소보다 더 깊은 곳에서 발견될 수 있다.[16] 물속에서 수직으로 이동하면서 다양한 압력과 온도에 노출되어 물속의 산소 농도에 영향을 받는다.[15] 이는 헨리의 법칙을 통해 이해할 수 있는데, 헨리의 법칙은 물질 내 기체의 농도는 압력과 용해도에 비례하며, 용해도는 온도에 의해 영향을 받는다고 설명한다.주로 문어는 외부 물에 최소한의 신체 부위만 노출되는 은신처에 위치한다.[18] 이동할 때는 대부분 해저나 해저를 따라 이동하며, 이 경우에도 문어의 아랫면은 여전히 가려져 있다.[18] 이러한 기어가는 방식은 신진대사 요구량을 크게 증가시켜, 휴식 상태의 문어에게 필요한 양의 약 2.4배에 달하는 산소 섭취량을 필요로 한다.[19] 이러한 증가된 요구량은 문어 심장의 1회 박출량 증가로 충족된다.[20]
문어는 때때로 물속에서 헤엄쳐 다니며 몸을 완전히 노출한다.[15] 이때, 외투강에서 훨씬 높은 압력을 생성하여 물속을 추진하는 제트 메커니즘을 사용한다.[20] 참문어의 심장과 아가미가 외투 내에 위치하기 때문에, 이 높은 압력은 심장으로 혈액을 되돌리는 다양한 혈관을 압박하고 제약한다.[20] 궁극적으로 이는 순환 문제를 야기하며, 문어는 최대 운동 시의 신진대사 요구량을 균형 있게 맞출 수 있는 산소 섭취량을 얻을 수 없기 때문에 지속 가능한 이동 방식이 아니다.[20]
3. 2. 호흡
문어는 아가미를 호흡 표면으로 사용한다. 아가미는 아가미 신경절과 접힌 새엽으로 구성되어 있다. 물은 아가미와 새엽 위로 천천히 이동하여 외투강으로 들어가 문어의 깔때기 밖으로 나간다.[22]문어 아가미의 구조는 많은 양의 산소 섭취를 가능하게 하며, 20°C 물에서 최대 65%까지 가능하다.[22] 문어의 얇은 피부는 생체 외 연구에서 산소 섭취의 많은 부분을 차지하며, 휴식 시 피부를 통한 모든 산소 흡수의 약 41%를 차지한다고 한다.[18] 문어가 운동할 때에는 피부를 통한 총 산소 흡수량은 33%로 감소하고, 먹이를 먹고 나서 움츠리고 있을 때에는 3%까지 떨어질 수 있다.[18] 문어의 호흡 색소인 헤모시아닌은 산소 섭취를 증가시킨다.[17] 문어는 물 속의 산소 농도가 약 3.5kPa[22] 또는 31.6% 포화도(표준 편차 8.3%)까지 떨어져도 일정한 산소 섭취량을 유지할 수 있다.[17] 바닷물의 산소 포화도가 약 1~10%로 떨어지면, ''Octopus vulgaris''에게 치명적일 수 있다.[17] 높은 산소 추출 비율은 에너지 절약과 저산소 농도 환경에서 사는 데 이점을 제공한다.[21]
물은 문어의 외투강으로 펌핑되어 내부 아가미와 접촉한다. 물은 정맥에서 돌아오는 혈액보다 산소 농도가 높으므로 산소가 혈액으로 확산된다. 문어의 조직과 근육은 크레브스 회로에서 포도당을 분해할 때 산소를 사용하고 이산화탄소를 방출한다. 보어 효과는 정맥혈의 산소 농도가 동맥혈보다 낮고 산소가 혈류로 확산되는 이유를 설명한다. 피크의 확산 법칙은 문어의 아가미에 혈관이 풍부한 많은 작은 주름이 포함되어 있는 이유를 설명한다. 이는 표면적을 증가시켜 확산 속도도 증가시킨다. 아가미 상피의 주름을 덮는 모세 혈관은 매우 얇은 조직 장벽(10 μm)을 가지고 있어 산소가 혈액으로 빠르고 쉽게 확산될 수 있다.[23] 물 속의 산소 분압이 낮은 상황에서는 혈액으로의 산소 확산이 감소하며,[24] 헨리의 법칙이 이러한 현상을 설명한다. 이 법칙은 ''O. vulgaris''가 물 속의 산소 농도가 변화함에 따라 외투강을 통해 순환하는 물의 양을 변경해야 하는 이유를 설명한다.[22]
아가미는 혈액보다 많은 산소를 운반하는 물과 직접 접촉하고 있으며, 이 물은 문어의 외투강으로 들어왔다. 아가미 모세 혈관은 매우 작고 풍부하여 물이 접촉할 수 있는 표면적을 증가시켜 혈액으로의 산소 확산을 향상시킨다. 일부 증거에 따르면 아가미의 새엽과 새엽 내의 혈관이 수축하여 모세 혈관을 통해 혈액을 추진하는 데 도움이 된다고 한다.[25]
3. 3. 순환
참문어는 세 개의 심장을 가지고 있다. 하나는 몸 전체에 산소가 풍부한 혈액을 보내는 주 심장이고, 다른 두 개는 각각의 아가미 옆에 있는 작은 아가미 심장이다.[26] 혈액 순환은 아가미에서 산소가 풍부한 혈액을 전신 심장의 심방으로 보낸 후, 심실을 거쳐 몸의 나머지 부분으로 펌프질하는 방식으로 이루어진다. 몸에서 산소가 고갈된 혈액은 아가미 심장으로 이동하여 아가미를 통해 혈액을 펌프질하여 산소를 공급받고, 다시 순환 과정을 시작하기 위해 전신 심방으로 돌아간다.[26] 전신 심장에서는 세 개의 대동맥이 나오는데, 두 개의 작은 동맥(복부 대동맥과 생식선 대동맥)과 하나의 주요 동맥인 등쪽 대동맥이 몸 대부분에 혈액을 공급한다.[27] 또한 참문어는 소화관 주변과 눈 뒤에 큰 혈액 동맥을 가지고 있어, 생리적 스트레스 시 예비 저장고 역할을 한다.[28]참문어의 심박수는 운동 시 크게 변하지 않지만, 산소 부족, 갑작스러운 자극, 또는 제트 추진 중 외투 압력에 의해 전신 심장의 일시적인 심장 마비가 유발될 수 있다.[29] 운동에 대한 유일한 보상은 전신 심장의 일회 박출량을 최대 3배까지 증가시키는 것이다.[29] 이는 거의 모든 빠른 움직임으로 인해 산소 부족을 겪게 됨을 의미한다.[29][30] 그러나 참문어는 아가미의 수용체를 통해 각 호흡 시 물에서 흡수하는 산소량을 조절할 수 있어,[22] 다양한 산소 압력에서 산소 섭취량을 일정하게 유지할 수 있다.[29] 세 심장의 박동 리듬은 일반적으로 두 아가미 심장이 함께 박동한 후 전신 심장이 박동하는 위상과 일치한다.[26] 프랭크-스탈링 법칙은 수축력과 일회 박출량을 통해 심장 기능에 기여하며, 혈관의 총 부피를 유지하고 심장이 제대로 기능하도록 시스템 내에서 비교적 일정하게 유지해야 한다.[31]
참문어의 혈액은 구리가 풍부한 헤모시아닌으로 구성되어 있는데, 이는 척추동물의 철이 풍부한 헤모글로빈보다 효율성이 떨어진다.[32] 동맥 내의 산소화된 헤모시아닌은 이산화탄소(CO|CO2영어)에 결합하고, 정맥 내 혈액이 산소가 고갈될 때 방출된다. 이산화탄소가 혈액으로 방출되면 탄산 형성에 의해 혈액이 산성화된다.[33] 보어 효과는 이산화 탄소 농도가 혈액 pH와 산소 방출 또는 섭취에 영향을 미친다고 설명한다. 크렙스 회로는 혈액에서 산소를 사용하여 활성 조직이나 근육에서 포도당을 분해하고 이산화 탄소를 노폐물로 방출하며, 이는 더 많은 산소 방출로 이어진다. 조직이나 근육으로 방출된 산소는 산소가 고갈된 혈액을 생성하고, 이 혈액은 정맥을 통해 아가미로 되돌아간다. 참문어의 두 아가미 심장은 정맥에서 아가미 모세 혈관을 통해 혈액을 펌프질한다. 새로 산소화된 혈액은 아가미 모세 혈관에서 전신 심장으로 배출되고, 여기서 신체 전체로 다시 펌프질된다.[26]
참문어 몸의 혈액량은 체중의 약 3.5%이지만,[28] 혈액의 산소 운반 능력은 약 4 부피 퍼센트에 불과하다.[29] 이는 산소 부족에 대한 민감성에 기여한다. Shadwick과 Nilsson[30]은 참문어 순환계가 "높은 생리적 수행에 근본적으로 적합하지 않다"라고 결론지었다. 결합제는 혈액 세포가 아닌 혈장에 존재하므로, 참문어가 경험할 수 있는 산소 섭취량에 제한이 있다. 혈류 내의 헤모시아닌을 증가시키면 유체가 너무 점성이 되어 근원성[34] 심장이 펌프질할 수 없게 된다.[31] 푸아죄유의 법칙은 혈압과 혈관 저항의 차이를 통해 전체 순환계에서 대량 유체의 흐름 속도를 설명한다.[31]
척추동물의 혈관과 마찬가지로 참문어 혈관은 매우 탄력적이며 생리적 압력에서 70%의 복원력을 갖는다. 이는 주로 참문어 동맥 탄성 중합체라는 탄성 섬유로 만들어지며, 과도한 신장을 방지하면서 혈관이 형태를 유지하도록 돕기 위해 더 뻣뻣한 콜라겐 섬유가 고압에서 동원된다.[35] Shadwick과 Nilsson[30]은 모든 참문어 혈관이 혈액을 몸 전체로 이동시키는 데 도움이 되도록 평활근 수축을 사용할 수 있다는 이론을 세웠는데, 이는 부속 압력과 함께 물속에서 생활하는 상황에서 타당하다.
참문어 대동맥의 탄성과 수축성은 심장에서 혈류의 맥박이 혈관 길이를 따라 이동하면서 혈류의 맥박적인 특성을 완화시키는 역할을 하며, 정맥은 에너지 저장 능력 역할을 한다.[30] 전신 심장의 일회 박출량은 정맥을 통한 입력 혈압과 대동맥을 통한 출력 역압의 차이와 반비례하여 변화한다.
3. 4. 삼투 조절
참문어를 포함한 두족류의 혈림프, 심낭액, 소변은 서로 등장성이며 주변 해수와도 같다.[36] 두족류는 삼투 조절을 하지 않는 것으로 여겨지는데, 이는 이들이 환경의 삼투압에 맞춰 적응하는 순응자임을 나타낸다. 삼투압 구배가 없기 때문에 유기체에서 해수로, 또는 해수에서 유기체로 물의 순 이동이 없다.[36] 문어는 평균 최소 염도 요구량이 27g/L이며, 환경에 상당량의 담수를 유입하는 모든 교란은 치명적일 수 있다.[37]그러나 이온 측면에서 해수와 두족류 내 이온 농도 사이에는 불일치가 있는 것으로 보인다.[36] 일반적으로 두족류는 해수와 대조적으로 나트륨, 칼슘 및 염화물의 저이온 농도를 유지한다. 황산염과 칼륨도 저이온 상태로 존재하며, 두족류의 배설계를 제외하고 소변은 고이온 상태이다.[36] 이러한 이온들은 자유롭게 확산되며, 유기체 내에서 저이온 농도로 존재하기 때문에 해수에서 유기체로 이동한다.[36] 유기체가 저이온 농도를 유지할 수 있다는 사실은 두족류 내에 이온 조절의 한 형태가 존재하고, 칼륨 및 황산염과 같은 특정 이온을 능동적으로 배설하여 항상성을 유지한다는 것을 시사한다.[36]
''O. vulgaris''는 포유류와는 매우 다른 연체동물 스타일의 신장 시스템을 가지고 있다. 이 시스템은 각 아가미 심장의 부속 기관을 중심으로 구축되었으며, 이는 본질적으로 심낭의 연장선이다.[36] 이 길고 섬모가 있는 관은 혈액을 한 쌍의 신장낭으로 여과하는 동시에 포도당과 아미노산을 혈류로 능동적으로 재흡수한다.[36] 신장낭은 소변의 이온 농도를 능동적으로 조절하고 질소질 화합물 및 기타 대사성 폐기물을 소변에 능동적으로 추가한다.[36] 여과 및 재흡수가 완료되면 소변은 각 신장낭에서 하나씩 한 쌍의 신장 유두를 통해 ''O. vulgaris''의 외투강으로 비워진다.[36]
온도와 신체 크기는 ''O. vulgaris''의 산소 소비에 직접적인 영향을 미치며, 이는 신진대사율을 변화시킨다.[19] 산소 소비가 감소하면 신진대사율이 느려지면서 암모니아 배설량도 감소한다.[19] ''O. vulgaris''는 혈액, 심낭액, 소변 및 신장액의 네 가지 체액을 가지고 있다. 소변과 신장액은 칼륨과 황산염 농도가 높지만 염화물 농도는 낮다. 소변은 칼슘 농도가 낮으며, 이는 능동적으로 제거되었음을 시사한다. 신장액은 혈액과 유사한 칼슘 농도를 갖는다. 염화물 농도는 혈액에서 높고 나트륨은 다양하다. 심낭액은 문어가 삼투 조절을 하지 않고 순응한다는 것을 뒷받침하는 해수와 유사한 나트륨, 칼륨, 염소 및 칼슘 농도를 갖는다. 그러나 황산염 농도는 더 낮다.[36] 심낭관에는 심낭액으로 알려진 혈액의 초여과물이 들어 있으며, 여과 속도는 근육과 신경이 풍부한 아가미 심장에 의해 부분적으로 제어된다.[36] 신장 부속물은 질소질 및 기타 폐기물을 혈액에서 신장낭으로 이동시키지만 부피를 추가하지 않는다. 신장액은 소변이나 혈액보다 암모니아 농도가 높으므로 신장낭은 암모니아를 신장 부속물에서 끌어내기 위해 산성으로 유지된다. 암모니아는 농도 기울기를 따라 소변이나 혈액으로 확산되며, 여기서 아가미 심장을 통해 펌핑되어 아가미로 확산된다.[36] ''O. vulgaris''에 의한 암모니아 배설은 이들을 암모니아배설성 유기체로 만든다. 암모니아 외에도 ''O. vulgaris''에 의해 소량의 요소, 요산, 퓨린 및 일부 유리 아미노산과 같은 몇 가지 다른 질소질 폐기물이 배설되는 것으로 밝혀졌다.[36]
신장낭 내에서 이온 조절을 담당하는 두 개의 인식되고 특정한 세포가 있다. 두 종류의 세포는 lacuna 형성 세포와 신장 세뇨관에 전형적인 상피 세포이다. 상피 세포는 섬모가 있고, 원통형이며 세 가지 뚜렷한 영역(정점, 중간 세포질, 기저 판)을 가진 분극화된 세포이다. 중간 세포질 영역은 미토콘드리아 및 활면 및 조면 소포체와 같은 여러 세포 소기관이 집중되어 있어 가장 활동적이다. 활동의 증가는 기저 판의 맞물린 미로가 경골어류의 미토콘드리아가 풍부한 세포와 유사한 교차 전류 활동을 생성하기 때문이다. lacuna 형성 세포는 기저 판과의 접촉을 특징으로 하지만 관련 상피 세포의 정점 가장자리에 도달하지 않으며 아가미 심장 상피에 위치한다. 모양은 매우 다양하며 때때로 상피 세포보다 전자 밀도가 높아 이온 농도를 조절하는 "확산 신장"으로 보인다.[38]
''O. vulgaris''의 적응 중 하나는 신장에 대한 직접적인 제어이다.[36] 이들은 원할 때 여과의 대부분을 수행하는 오른쪽 또는 왼쪽 신장 사이를 전환할 수 있으며, 스트레스나 운동으로 혈압이 증가할 때 여과 속도가 증가하지 않도록 조절할 수 있다.[36] 일부 문어 종은 생식소 공간에서 아가미 심낭으로 이어지는 관을 가지고 있다.[36] Wells[36]는 이 관이 혈관이 풍부하고 신경이 분포되어 있어 임신한 암컷의 난낭액에서 중요한 대사 산물을 신장 부속물로 유도하여 재흡수할 수 있다고 이론화했다.
3. 5. 체온 조절
참문어(O. vulgarisla)는 해양 생물로서 계절, 지리적 위치, 수심 등 여러 요인에 따라 다양한 온도를 경험한다.[39] 예를 들어, 나폴리 주변에 서식하는 문어는 여름에는 25°C, 겨울에는 15°C의 온도를 경험할 수 있다.[39] 이러한 온도 변화는 매우 점진적으로 일어나기 때문에 급격한 체온 조절은 필요하지 않다.참문어는 변온동물이자 광온성 외온동물로서 주변 온도에 순응한다.[40] 즉, 주변 환경과 체온 사이에 큰 차이가 없으며, 빠르게 평형을 이룬다. 더 따뜻한 곳으로 이동하면 주변 물에서 열을 얻고, 더 차가운 곳으로 이동하면 열을 잃는 방식이다.
''O. vulgaris''는 다양한 환경 온도에 적응하기 위해 행동을 변화시킨다. 문어의 호흡 속도는 온도에 민감하여 온도가 높아질수록 호흡이 빨라진다.[41] 산소 소비량은 16°C에서 28°C 사이에서 증가하여 28°C에서 최대가 되고, 32°C에서 감소하기 시작한다.[41] 신진대사와 산소 소비에 가장 적합한 온도는 18°C에서 24°C 사이이다.[41] 온도 변화는 혈림프 단백질 농도 변화를 유발하는데, 온도가 상승하면 단백질 농도가 증가하여 헤모시아닌의 협동성은 증가하고 친화도는 감소한다.[42] 반대로 온도가 감소하면 호흡 색소 협동성은 감소하고 친화도는 증가한다.[42] 온도 변화에 따라 P50이 약간 증가하여 산소 압력이 모세혈관에서 높게 유지되도록 돕고, 산소 소비량이 많은 기간 동안 미토콘드리아로의 산소 확산을 증가시킨다.[42] 온도 증가는 효소 활성을 높이지만, 헤모시아닌 친화도 감소는 효소 활성이 일정하게 유지되고 항상성을 유지하도록 돕는다. 가장 높은 혈림프 단백질 농도는 32°C에서 나타나며, 이보다 높은 온도에서는 감소한다.[41] 혈액 내 산소 친화도는 pH 7.4에서 만큼 감소한다.[42] 문어의 내열성은 산소 소비 능력에 의해 제한되며, 극단적인 온도에서 충분한 산소를 공급받지 못하면 치명적일 수 있다.[41] ''O. vulgaris''는 산소의 일정한 압력에서 호흡 색소에 결합된 산소의 양을 나타내는 pH와 무관한 정맥 예비량을 가지고 있어, 온도와 관련된 광범위한 pH를 견딜 수 있다.[42]
''O. vulgaris''는 온도 순응자로서,[43] 열 생산 또는 열 교환을 위한 특정 기관이나 구조가 없다. 다른 동물들과 마찬가지로 음식 소화와 같은 일반적인 신진대사 과정에서 열을 생성하지만,[39] 체온을 특정 범위로 유지하기 위한 특별한 방법은 없다. 문어가 선호하는 온도는 적응된 온도를 직접적으로 반영한다.[43] 13°C에서 28°C의 주변 온도 범위를 허용하며,[43] 최대 신진대사 효율을 위한 최적 온도는 약 20°C이다.[40]
외온성 동물인 참문어는 온도 변화에 큰 영향을 받는다. 모든 종은 기초대사율로 기능할 수 있는 열 선호를 갖는다.[43] 낮은 신진대사율은 빠른 성장을 가능하게 하므로, 물이 선호하는 구역에 가까워질 때 짝짓기를 한다. 온도가 상승하면 ''O. vulgaris''의 산소 소비량이 증가한다.[19] 이는 신진대사율과 직접 관련되는데, 포도당과 같은 분자 분해에는 크렙스 회로에서처럼 산소가 필요하기 때문이다. 암모니아 배설량은 온도가 상승함에 따라 감소한다.[19] 암모니아 배설량 감소는 온도 증가에 따라 더 많은 에너지를 소비해야 하는 문어의 신진대사와 관련이 있다. 참문어는 신진대사율 증가로 인해 배설했을 과도한 용질을 사용하기 위해 암모니아 배설량을 줄인다. 문어는 죽음을 피하기 위해 조절을 시작해야 하는 임계값에 도달할 때까지 내부 온도를 조절하지 않는다.[19] 온도 상승에 따른 신진대사율 증가는 문어가 선호하는 온도 구역 내에 머물기 위해 얕거나 깊은 곳으로 이동하기 때문일 가능성이 높다.
4. 생식
참문어의 산란기는 12월부터 9월까지이며, 봄에 가장 활발하다.
참조
[1]
논문
Phylogeography of the insular populations of common octopus, Octopus vulgaris Cuvier, 1797, in the Atlantic Macaronesia
2020-03-19
[2]
간행물
"''Octopus vulgaris''"
2022-03-18
[3]
서적
"''Cephalopods: A World Guide''"
ConchBooks
[4]
웹사이트
Common Octopus
https://www.national[...]
National Geographic
2011-05-10
[5]
논문
Age Validation in Octopus vulgaris Beaks Across the Full Ontogenetic Range: Beaks as Recorders of Life Events in Octopuses
https://bioone.org/j[...]
[6]
논문
Cannibalistic behavior of octopus (Octopus vulgaris) in the wild.
2014-11
[7]
논문
Isolation and characterization of novel tachykinins from the posterior salivary gland of the common octopus Octopus vulgaris
2003-01
[8]
Youtube
Octopus opens jar
https://www.youtube.[...]
YouTube
2010-09-16
[9]
Youtube
Octopus in prawn trap
https://www.youtube.[...]
YouTube
2007-02-12
[10]
서적
Other Minds: The Octopus, the Sea, and the Deep Origins of Consciousness
https://us.macmillan[...]
Farrar, Straus and Giroux
[11]
웹사이트
The Animals (Scientific Procedures) Act(Amendment) Order 1993
http://www.legislati[...]
1993-08-23
[12]
웹사이트
FAO Fisheries & Aquaculture – Aquatic species
http://www.fao.org/f[...]
[13]
웹사이트
Octopus vulgaris
https://animaldivers[...]
[14]
문서
Distribution and abundance of ''Octopus vulgaris'' Cuvier 1797, (Cephalopoda: Octopoda) in the Mediterranean Sea. Scientia Marina, '''66'''(S2): 157–166.
[15]
문서
Essential habits for pre-recruit Octopus vulgaris along the Portuguese coast. Fisheries Research, '''152''': 74–85.
[16]
문서
Abundance of ''Octopus vulgaris'' on soft sediment. Scientia Marina, 68, 553–560.
[17]
문서
Suitable dissolved oxygen levels for common octopus (''Octopus vulgaris'' cuvier, 1797) at different weights and temperature: analysis of respiratory behavior. Aquaculture. 244: 303–314.
[18]
문서
Cutaneous respiration in ''Octopus vulgaris'']. The Journal of Experimental Biology, '''199''': 2477–2483
http://jeb.biologist[...]
[19]
문서
Oxygen consumption and ammonia excretion of Octopus vulgaris (Cephalopoda) in relation to body mass and temperature. Marine Biology, 146, 725–732.
[20]
문서
Blood flow and pressure changes in exercising octopuses (Octopus vulgaris). The Journal of Experimental Biology, 131, 175–187
[21]
문서
Evolution of the gills in the octopodiformes. Bulletin of marine science. 71(2): 1003–1017
[22]
문서
The control of ventilatory and cardiac responses to changes in ambient oxygen tension and oxygen demand in Octopus. The Journal of Experimental Biology, 198, 1717–1727
[23]
문서
The morphometrics of cephalopod gills. Journal of the Marine Biological Association of the United Kingdom, 74(3), 687–706
1994-08
[24]
문서
Temperature-dependent oxygen extraction from the ventilatory current and the costs of ventilation in the cephalopod Sepia officinalis. Journal of Comparative Physiology B, 176, 607–621
[25]
문서
The performance of the octopus circulatory system: A triumph of engineering over design. Experientia, 43, 487–499
[26]
문서
Nervous control of the heartbeat in Octopus]. The Journal of Experimental Biology, 85, 111–128.
http://jeb.biologist[...]
[27]
문서
The role of venous pressure in regulation of output from the heart of the octopus, ''Eledone cirrhosa'' (Lam.)]. The Journal of Experimental Biology, 93, 243–255
http://jeb.biologist[...]
[28]
문서
Circulation time, blood reserves and extracellular space in a cephalopod. The Journal of Experimental Biology, 113, 461–464.
[29]
문서
The heartbeat of ''Octopus vulgaris'']. The Journal of Experimental Biology, 78, 87–104
http://jeb.biologist[...]
[30]
문서
The importance of vascular elasticity in the circulatory system of the cephalopod ''Octopus vulgaris'']. The Journal of Experimental Biology, 152, 471–484
http://jeb.biologist[...]
[31]
서적
"Animal Physiology"
Sinauer Associates
[32]
웹사이트
Octopus Fact Sheet
http://www.worldanim[...]
World Animal Foundation
[33]
논문
Coordination of metabolism, acid-base regulation and haemocyanin function in cephalopods
https://epic.awi.de/[...]
1995-01
[34]
문서
Excitation-contraction coupling
[35]
문서
Mechanical properties of the octopus aorta]. The Journal of Experimental Biology, 114, 259–284
http://jeb.biologist[...]
[36]
서적
Octopus: Physiology and behaviour of an advanced invertebrate
Cambridge: University Printing House
1978
[37]
논문
Aquaculture potential of the common octopus (Octopus vulgaris Cuvier, 1797): a review
2004-09
[38]
논문
The fine structure of the renopericardial cavity of the cephalopod ocotopus dofleine martini
1975
[39]
서적
Animal Physiology
Sunderland: SinauerAssociates
2012
[40]
논문
Effect of temperature on specific dynamic action in the common octopus, Octopus vulgaris (Cephalopoda)
2005
[41]
논문
The respiration, excretion and biochemical response of the juvenile common Chinese cuttlefish, Sepiella maindroni at different temperatures
2013-07
[42]
논문
Temperature effects on hemocyanin oxygen binding in an antarctic cephalopod
2001
[43]
논문
Thermopreference, tolerance and metabolic rate of early stages juvenile Octopus maya acclimated to different temperatures
2013
[44]
논문
Reproductive cycle and energy allocation of Octopus vulgaris in Galician waters, NE Atlantic
https://doi.org/10.1[...]
2007
[45]
웹사이트
생물종정보 :: 참문어 Octopus vulgaris
http://www.nifs.go.k[...]
[46]
서적
Cephalopods: A World Guide
ConchBooks
2000
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com