체세포 분열
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
체세포 분열은 세포가 증식하는 과정으로, 유전 물질을 정확하게 복제하여 두 개의 동일한 딸세포로 분열하는 현상이다. 18세기와 19세기에 걸쳐 발견되었으며, 동물, 식물, 원생생물 등 다양한 진핵생물에서 나타난다. 체세포 분열은 간기, 핵분열(전기, 전중기, 중기, 후기, 말기), 세포질분열의 단계를 거치며, 세포 주기 조절, DNA 복구 메커니즘, 그리고 다양한 오류 및 변이 과정을 포함한다. 체세포 분열은 핵막 유지 여부, 방추체 대칭성, 중심 방추 위치에 따라 분류되며, 조직병리학에서 종양의 공격성을 진단하는 지표로 활용되기도 한다.
더 읽어볼만한 페이지
- 세포 분열 - 감수분열
감수분열은 유성생식을 하는 생물에서 배우자를 형성하는 세포분열 과정으로, 염색체 수를 줄여 유전적 다양성을 확보하며, 감수분열 I과 II 두 단계로 구성되어 상동염색체 및 자매염색분체 분리, 독립적 분리 및 교차를 통해 유전적 다양성을 증가시키지만, 염색체 비분리 오류는 염색체 이상 질환을 유발할 수 있고 진핵생물 유성생식과 진화에 필수적인 과정이다. - 체세포 분열 - 염색분체
염색분체는 염색체를 구성하는 두 가닥 중 하나로, 동일한 염색체의 복제본인 자매 염색분체와 상동 염색체 쌍을 이루는 비자매 염색분체로 나뉘며, 자매 염색분체는 유사 분열 또는 감수 분열 II 후기에 분리되고 비자매 염색분체는 감수 분열 전기 I에 염색체 교차를 통해 유전 물질을 교환한다. - 체세포 분열 - 방추체
방추체는 세포 분열 시 염색체를 분리하는 세포 내 구조물로, 미세소관, 운동 단백질, 염색체로 구성되어 타원체 모양을 가지며 중심체 매개 모델과 염색질 매개 모델로 형성되고 유사 분열 키나아제에 의해 조절되며 세포 분열 방향을 결정한다. - 1835년 과학 - 세포 분열
세포 분열은 원핵생물과 진핵생물의 방식으로 나뉘며, 진핵생물의 경우 염색체 수 변화에 따라 체세포 분열과 감수 분열로 구분되고, 최근에는 DNA 복제 없는 비합성 분열도 발견되었으며, 세포 주기를 거쳐 DNA 손상 등에 의해 조절되는 세포 증식 과정이다. - 1835년 과학 - 1835년 5월 27일 일식
| 체세포 분열 | |
|---|---|
| 개요 | |
| 정의 | 세포 분열의 한 형태로, 모세포의 핵이 분열하여 두 개의 동일한 딸핵을 생성하는 과정임. |
| 과정 | 염색체 복제 및 분리 과정을 통해 유전 물질을 정확히 분배함. |
| 결과 | 유전적으로 동일한 두 개의 딸세포 생성. |
| 특징 | |
| 유사 분열 | 염색체를 분리하여 두 개의 새로운 동일한 핵을 생성하는 과정. |
| 세포 주기 | 일반적으로 세포 주기의 일부로, 세포 성장 및 DNA 복제와 함께 발생함. |
| 진핵 세포 | 진핵 세포에서만 일어남. 핵이 없는 원핵 세포는 이분법이라는 다른 과정으로 분열함. |
| 명칭 유래 | 그리스어 'μίτος (mitos, 실)'에서 유래. |
| 단계 | |
| 유사 분열 단계 | 전기 전중기 중기 후기 말기 |
| 세포질 분열 | 세포질 분열은 유사 분열과는 별개의 과정이지만, 유사 분열 후기에 시작되어 세포 분열을 완료함. |
| 기능 및 중요성 | |
| 성장 및 발달 | 다세포 유기체의 성장 및 발달에 필수적임. |
| 조직 복구 | 손상된 조직의 복구 및 재생에 관여함. |
| 무성 생식 | 일부 유기체에서 무성 생식의 주요 방식임. |
| 유전적 안정성 | 딸세포가 모세포와 동일한 유전 정보를 갖도록 보장함. |
| 조절 | |
| 세포 주기 조절 | 세포 주기의 여러 지점에서 조절되어 정확한 DNA 복제 및 염색체 분리를 보장함. |
| 유사 분열 검문소 | 오류가 있는 경우 유사 분열을 중단시키는 검문소 메커니즘이 존재함. |
| 이상 | |
| 염색체 수 이상 | 유사 분열 오류는 이수성과 같은 염색체 수 이상을 초래할 수 있으며, 이는 암을 포함한 다양한 질병과 관련이 있음. |
| 삼중 극 유사 분열 | 사람 세포와 배아에서 발생할 수 있으며, 병리학적 및 의학적 영향을 미침. |
| 기타 | |
| 유사 분열 지수 (Mitotic Index, MI) | 세포 집단에서 유사 분열을 겪고 있는 세포의 비율을 나타내는 지표임. |
2. 역사
세포 분열에 대한 초기 연구는 18세기와 19세기에 걸쳐 이루어졌다. 1882년 발터 플레밍은 그리스어 μίτος (mitos, 날실)에서 유래한 "체세포분열(mitosis)"이라는 용어를 만들었다.[84][85][86]
"체세포분열"이라는 용어 외에도 아우구스트 바이스만이 제안한 "균등 분열"이라는 용어가 있는데, 이는 체세포분열과 유사한 감수 2분열을 지칭할 때 더 일반적으로 사용된다.[92] 일부 학자들은 "체세포분열"을 핵분열과 세포질분열을 모두 포함하는 넓은 의미로 사용하기도 한다.[91]
원래 "mitosis"는 세포 분열 과정에서 나타나는 날실 모양의 방추섬유 때문에 붙여진 이름이다. 이 용어는 일본에서 "有絲分裂"(유사분열)로 번역되었고, 한국에서는 일본의 영향을 받아 "유사분열"로 사용하다가 이후 "체세포분열"로 변경되었다. "체세포분열"은 "mitosis"의 번역어로 권장되지만, 원래 용어의 어원을 충분히 살리지 못했다는 점에서 더 적절한 용어가 필요하다는 의견도 있다.
2. 1. 초기 발견
세포 분열은 18세기와 19세기에 걸쳐 여러 학자들에 의해 묘사되었다.[80] 1835년 독일의 식물학자 후고 폰 몰은 녹조류의 일종인 ''클라포라''에서 세포 분열을 통해 세포가 증식하는 과정을 관찰했다.[74][75][76] 1838년 독일의 식물학자 마티아스 야콥 슐라이덴은 식물 세포 내에서 새로운 세포가 형성되는 것이 세포 증식의 일반적인 법칙이라고 주장했지만, 이후 로베르트 레마크 등의 연구를 통해 후고 폰 몰의 모델이 지지를 받게 되었다.[77]동물 세포에서의 체세포분열은 1873년 개구리, 토끼, 고양이의 각막 세포에서 처음 발견되었으며, 1875년 폴란드의 조직학자 바츨라프 마이젤에 의해 최초로 묘사되었다.[78][79]
1873년 독일의 동물학자 오토 뷔칠리는 선형동물 관찰 결과를 발표했으며, 몇 년 후 이 관찰 결과를 바탕으로 체세포분열 과정을 발견하고 묘사했다. 오토 뷔칠리 외에도 슈나이더(Schneider), 폴(Fol) 등이 체세포분열 과정 발견에 기여했을 가능성이 있다.[80][81][82][83]
2. 2. 용어의 정립
1882년 발터 플레밍은 그리스어 μίτος (mitos, 날실)에서 유래한 "체세포분열(mitosis)"이라는 용어를 만들었다.[84][85][86] 1878년 슐라이허(Schleicher)가 도입한 "핵분열(karyokinesis)"[87][88], 아우구스트 바이스만이 제안한 "균등 분열(equational division)"[89] 등의 다른 용어도 존재했다.[90]일본에서는 "有絲分裂"(유사분열)로 번역되었고, 한국에서는 이를 그대로 읽어 '유사분열'로 사용하다가 '체세포분열'로 굳어졌다. '유사분열'은 실이 존재하는 분열이라는 뜻을 담고 있다.
3. 단계
체세포분열은 간기 이후, 전기, 전중기, 중기, 후기, 말기의 단계를 거친다.
- 전기(Prophase): 염색체가 응축되고, 핵막이 사라지며, 방추사가 형성되기 시작한다.
- 전중기(Prometaphase): 핵막이 완전히 사라지고, 방추사가 염색체의 동원체에 부착된다.
- 중기(Metaphase): 염색체가 세포 중앙의 적도판에 배열된다.
- 후기(Anaphase): 자매 염색분체가 분리되어 양쪽 극으로 이동한다.
- 말기(Telophase): 분리된 염색체 주위에 새로운 핵막이 형성되고, 염색체가 다시 풀어진다.
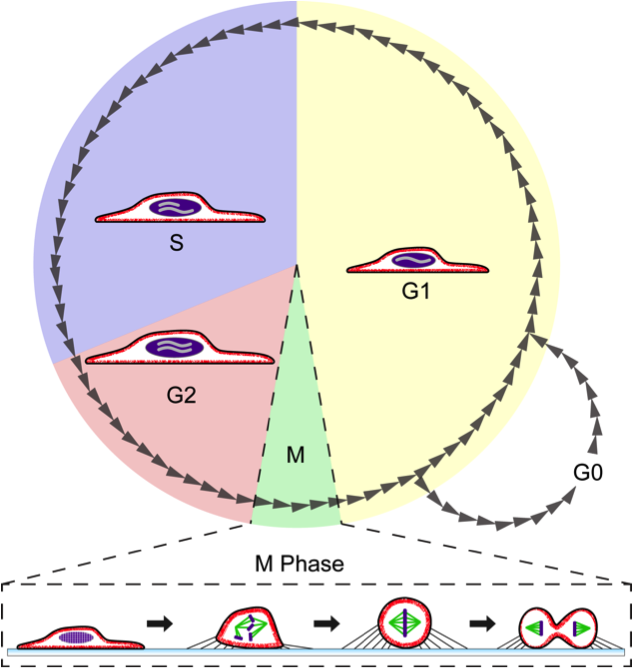
3. 1. 간기 (Interphase)
간기(Interphase)는 세포 주기에서 체세포분열기(M기)를 제외한 시기로, 세포가 성장하고 DNA를 복제하며 체세포분열을 준비하는 기간이다. 간기는 G1기, S기, G2기의 세 단계로 나뉜다.- G1기(Gap 1): 세포가 성장하며, 단백질과 세포 소기관을 합성한다.
- S기(Synthesis): DNA가 복제되어 염색체가 두 개의 자매 염색 분체로 구성된다.
- G2기(Gap 2): 체세포분열을 준비하며, 추가적인 성장과 단백질 합성이 일어난다.
간기 동안 DNA 이중 가닥 절단이 발생하면, 비상동 말단연결(non-homologous end joining; NHEJ) 또는 상동 재조합 복구(homologous recombinational repair; HRR) 과정을 통해 복구된다. NHEJ는 간기의 모든 시기에 작동하며, HRR은 S기와 G2기에 작동한다.
사이클린과 사이클린-의존성 카이네이스는 세포 주기를 조절하는 주요 단백질이며, 체크포인트를 통해 간기의 각 단계가 정확하게 진행되도록 한다. 간기 동안 세포의 DNA 손상이나 중요 과정의 미완료는 세포 주기를 중단시킬 수 있다. 이러한 조절 기작은 체세포분열의 성공적인 완료를 보장하고, 잠재적인 암세포 발생을 억제하는 데 중요한 역할을 한다.[96]
3. 1. 1. G0기 (휴지기)
G0영어기(휴지기)는 세포 주기를 벗어나 분열하지 않는 상태이다. 밀도 의존성 억제, 세포 분화 등에 의해 유도될 수 있다.3. 2. 체세포분열기 (M기)
체세포분열기는 핵분열(Mitosis)과 세포질분열(Cytokinesis)로 구성된다.
체세포분열과 세포질분열을 통해 모세포의 지놈이 두 개의 딸세포로 전달된다. 각 딸세포는 모세포와 유전적으로 동일해야 하기 때문에, 모세포는 체세포분열을 하기 전에 S기에서 염색체를 복제한다. 염색체 복제가 일어나면 두 개의 동일한 자매 염색분체가 생기며, 각 자매 염색분체는 중심절(동원체)에서 코히신(cohesin)이라는 단백질로 서로 묶여 있게 된다.
체세포분열이 시작되면 염색체는 응축하여 광학 현미경으로 보이게 된다. 동물과 같은 일부 진핵생물은 체세포분열이 진행되면 핵막을 작은 소포들로 해체시키고, 핵에서 리보솜을 만드는 핵소체도 사라진다. 미세소관은 세포의 반대편 끝으로 뻗어나가 중심절에 부착하고 염색체를 세포 중앙에 배열시킨다. 이후 미세관은 각 염색체의 자매 염색분체를 끌어당겨 복제된 염색체를 분리시킨다. 분리된 직후 자매 염색분체는 ''딸염색체''라고 부른다. 세포가 늘어나면서 각 딸염색체는 세포의 반대편 끝을 향하여 당겨지고, 후기 후반부에 최대로 응축한다. 말기에는 새로운 핵막이 분리된 딸염색체 주변에서 형성되고, 딸염색체들은 풀려서 간기 핵을 형성한다.
일반적으로 체세포분열 후기가 진행되고 나면 세포는 세포질분열 과정을 겪는다. 동물 세포의 경우 만들어지고 있는 두 개의 핵 사이로 세포막이 세포 안쪽으로 좁혀 들어가고, 식물 세포의 경우 세포판이 두 개의 핵 사이에서 형성된다. 세포질분열이 항상 일어나는 것은 아니며, 일부 다핵세포는 세포질분열 없이 체세포분열을 진행한다.

3. 2. 1. 전기 (Prophase)

전기는 G2 간기 후에 발생하며, 세포는 염색체 응축을 시작하고 세포 분열 방추체 형성을 시작하여 분열을 준비한다. 간기 동안 핵 내의 유전 물질은 느슨하게 포장된 크로마틴으로 구성된다. 전기가 시작될 때, 크로마틴 섬유는 광학 현미경으로 고배율에서 볼 수 있는 개별 염색체로 응축된다.[34][35][36] 이 단계에서 염색체는 길고 얇으며 실 모양이다. 각 염색체는 두 개의 염색분체를 가지고 있다. 두 개의 염색분체는 중심절에서 결합된다.
유전자 전사는 전기 동안 중단되며 후기 후기에서 초기 G1 단계까지 재개되지 않는다.[34][35][36] 핵소체도 전기 초기에 사라진다.[37]
동물 세포의 핵 근처에는 중심체라고 하는 구조가 있으며, 이 구조는 중심립 쌍과 느슨한 단백질 덩어리로 구성된다. 중심체는 세포의 미세 소관 조절 중심이다. 세포는 세포 분열 시 단일 중심체를 상속받으며, 새로운 세포 분열이 시작되기 전에 복제되어 중심체 쌍을 형성한다. 두 중심체는 튜불린을 중합하여 미세 소관 방추체 형성에 도움을 준다. 그런 다음 운동 단백질은 이 미세 소관을 따라 중심체를 세포의 반대쪽으로 밀어낸다. 중심체는 미세 소관 조립을 돕지만, 식물에는 없기 때문에 방추체 형성에 필수적이지 않으며, 동물 세포 분열에도 절대적으로 필요하지 않다.[38]
3. 2. 2. 전중기 (Prometaphase)
염색체 응축이 더욱 진행된다. 핵소체 소실이 시작된다. 떨어져 있던 두 개의 중심체는 방추체 극이 되고, 거기에서 뻗어 나온 미세소관이 염색체의 동원체에 결합한다. 이 미세소관은 동원체 미세소관이라고 불린다. 미세소관은 핵막 안으로 진입한다. 핵막 붕괴형 유사분열을 하는 생물에서는 핵막이 소실된다.[39]전기 후반에는 ''동원체 미세 소관''이 염색체 동원체를 찾고 부착하기 시작한다. 동원체는 후기 전기에 염색체 중심체에 형성되는 단백질성 미세 소관 결합 구조이다.[39] 다수의 ''극 미세 소관''은 반대쪽 중심체에서 해당하는 극 미세 소관을 찾아 상호 작용하여 유사 분열 방추사를 형성한다.
동원체의 구조와 기능은 완전히 이해되지 않았지만, 일종의 분자 모터를 포함하고 있다는 것은 알려져 있다. 미세 소관이 동원체와 연결되면, 모터가 활성화되어 ATP로부터 에너지를 사용하여 기원하는 중심체를 향해 관을 따라 "기어" 올라간다. 이러한 모터 활성은 미세 소관의 중합 및 탈중합과 결합되어 나중에 염색체의 두 자매 염색 분체를 분리하는 데 필요한 인장력을 제공한다.
3. 2. 3. 중기 (Metaphase)
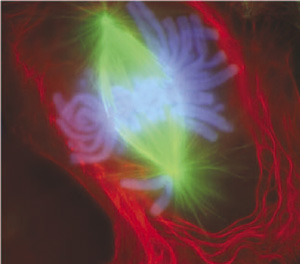
염색체가 세포의 적도면(세포 중앙)에 배열되며, 각 염색체에서 한 쌍의 자매 염색 분체가 식별 가능하게 된다. 이때 염색체의 각 1세트가 각각 올바르게 방추사에 결합되었는지 확인하는 방추사 점검 지점이 작동한다. 결합에 문제가 있는 경우 세포 주기가 중단된다.[93] 이 시기는 올바른 염색체 분배, 더 나아가 세포 분열을 위한 본질적인 단계로 여겨진다.
미세소관이 전중기에서 동원체에 위치하고 부착된 후, 두 개의 중심체는 염색체를 세포의 반대쪽 끝으로 당기기 시작한다. 그 결과 염색체는 두 개의 중심체 사이 중앙에 위치한 가상의 선인 중기판을 따라 정렬된다(세포의 대략 중앙선). 유사 분열의 끝에서 염색체의 공정한 분배를 보장하기 위해, ''중기 검문점''은 동원체가 유사 분열 방추에 적절하게 부착되어 있고 염색체가 중기판을 따라 정렬되어 있는지 확인한다.
3. 2. 4. 후기 (Anaphase)

후기 A 동안, 자매 염색분체를 함께 묶는 코헤신이 절단되어 두 개의 동일한 딸 염색체가 형성된다.[41] 동원체 미세 소관이 짧아지면서 새롭게 형성된 딸 염색체를 세포의 반대쪽 끝으로 당긴다. 후기 B 동안에는 극 미세 소관이 서로 밀어내 세포가 길어진다. 후기 말에는 염색체가 최대 응축 수준에 도달하여 염색체 분리와 핵의 재형성을 돕는다.[40] 대부분의 동물 세포에서는 후기 A가 후기 B보다 먼저 일어나지만, 일부 척추동물 난세포에서는 반대 순서로 일어난다.[41]
자매 염색 분체를 연결하고 있던 동원체 부근의 코헤신 단백질이 절단되어 분리가 가능해진다. 각각의 자매 염색 분체는 분열 장치의 작용에 의해 양극으로 이동한다. 미세 소관의 작용에 의해 염색 분체가 이동함과 동시에 방추체 극이 멀어진다.
3. 2. 5. 말기 (Telophase)
말기는 전중기 및 전기의 현상들이 역전되는 단계이다. 말기에 극 미세소관들은 계속 길어져 세포를 더욱 길게 만든다. 핵막이 붕괴되었을 경우, 모세포의 오래된 핵막의 막 소포들을 사용하여 새로운 핵막이 형성된다. 새로운 막은 분리된 각 딸 염색체 집합체 주위에 형성되며 (막이 중심체를 둘러싸지는 않음) 핵소체가 다시 나타난다. 새로운 핵막으로 둘러싸인 두 세트의 염색체는 이제 "풀리거나" 탈응축되기 시작한다. 이로써 체세포분열은 완료된다. 각 딸핵은 동일한 염색체 집합체를 갖는다.[93] 세포 분열은 이 시점에 생물체에 따라 발생할 수도 있고, 그렇지 않을 수도 있다.
후기에 분배된 딸 염색체는 분산되어 염색질로 돌아간다. 핵막 붕괴형 분열을 하는 생물에서는 핵막도 재구성된다.
3. 2. 6. 세포질분열 (Cytokinesis)
세포질분열은 체세포분열 시기에 포함되지는 않지만, 체세포분열기(M기)의 마지막 단계로서 세포 분열을 완료한다. 동물 세포와 식물 세포 모두 세포 분열은 골지체에서 유래한 소포에 의해 일어나며, 이 소포들은 미세소관을 따라 세포 가운데로 이동한다.
동물 세포에서는 수축 고리를 가지는 분열고랑(cleavage furrow)이 중기판이 있던 부위에 집게처럼 만들어져 각 분리된 핵을 나눈다.
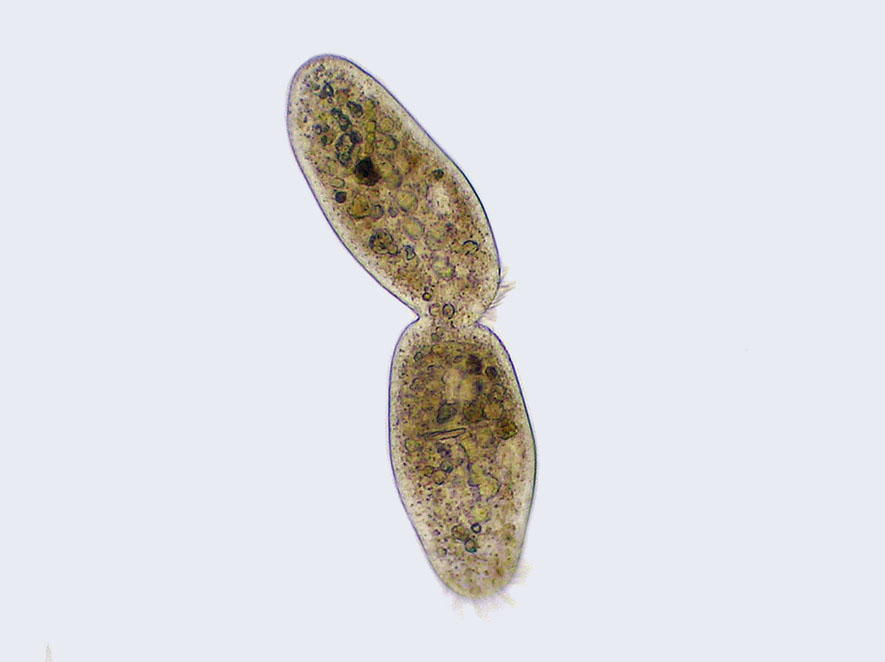
식물 세포에서는 골지체 구조가 합쳐져 격벽형성체(phragmoplast) 중심에서 세포판을 형성하고, 이것이 발달하여 세포벽이 되어 두 개의 딸핵을 분리한다. 격벽형성체는 일반적으로 고등 식물에서 나타나는 미세관 구조이고, 일부 녹조류는 해조체(phycoplast)를 이용하여 세포질분열 때 미세관을 배열한다.[65] 각 딸세포는 모세포와 완전히 똑같은 사본을 가진다. 세포질분열이 끝나면 체세포분열기(M기)도 함께 끝난다.
여러 세포들은 체세포분열과 세포질분열이 각각 따로 일어나면서 하나의 세포가 여러 개의 핵을 가지기도 한다. 이런 현상은 진균, 점균류, 다핵 조류에서 두드러지게 관찰할 수 있다. 심지어 일부 동물도 세포질분열과 체세포분열이 독립적으로 일어나기도 하는데, 초파리의 배아 발생 때 나타나는 특정 단계가 대표적인 예시다.
4. 체세포분열의 다양성
생물 종에 따라 체세포분열 과정에 차이가 있다. 진핵생물 세포의 체세포분열은 유사한 패턴을 따르지만, 세 가지 주요 세부 사항에서 변형이 있다. 핵막 유지 여부, 방추체 대칭성, 중심 방추 위치(닫힌 가쪽체세포분열의 경우)에 따라 다양한 형태로 나타난다.
박테리아와 고세균은 핵이 없으므로 핵 분열은 진핵생물 세포에서만 일어난다. 세균과 고세균은 다른 유형의 분열을 겪는다.[44][45] 각 진핵생물 상위 분류군 내에서, 단세포 Excavata를 제외하고는 개방형 유사 분열과 폐쇄형 유사 분열을 모두 찾을 수 있으며, Excavata는 전적으로 폐쇄형 유사 분열을 보인다.[46]
4. 1. 핵막 유지 여부에 따른 분류
진핵생물 세포에서의 체세포분열 과정은 비슷하게 진행되지만, 구체적인 과정에서는 3가지 변이가 존재한다. 핵막이 온전하게 유지되는지, 분해되는지에 따라 "닫힌(closed)" 체세포분열과 "열린(open)" 체세포분열로 구별할 수 있고, 핵막이 부분적으로만 분해되는 중간 형태를 "반열린(semiopen)" 체세포분열이라고 한다.아래는 진핵생물에서 발생하는 체세포분열 형태이다.[107]
| 유형 | 특징 | 예시 |
|---|---|---|
| 닫힌 핵속 가쪽체세포분열 | 핵막 유지, 중심 방추 핵 속, 가쪽체세포분열 | 유공충, 일부 담녹조강(Prasinophyceae), 옥시모나드목(Oxymonad), 아세토스포레아강(Ascetosporea), 많은 진균들(호상균류, 난균류, 접합균류, 자낭균문), 일부 방산충(스푸멜라리아(Spumellaria)와 아칸싸리아(Acantharia)) |
| 닫힌 핵바깥 가쪽체세포분열 | 핵막 유지, 중심 방추 핵 바깥, 가쪽체세포분열 | 트리코모나디다목(Trichomonadidia), 와편모충류 |
| 닫힌 정상체세포분열 | 핵막 유지, 정상체세포분열 | 규조류, 섬모충, 일부 미포자충류, 단세포 효모, 일부 다세포 진균 |
| 반열린 가쪽체세포분열 | 핵막 부분적 붕괴, 가쪽체세포분열 | 대부분의 정단복합체충류 |
| 반열린 정상체세포분열 | 핵막 부분적 붕괴, 정상체세포분열 | 일부 아메바(엽상근족충류), 일부 녹색 편모충(라피도파이트강(Raphidophyta)이나 좁쌀공말) |
| 열린 정상체세포분열 | 핵막 붕괴, 정상체세포분열 | 포유류와 다른 후생동물, 육지 식물, 일부 원생생물 |
4. 2. 방추체 대칭성에 따른 분류
방추장치의 대칭성에 따라 중기에서 나타나는 방추장치가 거의 중심에서 축의 방향으로 대칭인 경우를 "정상체세포분열(orthomitosis)"이라고 하고, 방추 장치 모양이 기이하게 좌우대칭인 경우를 "가쪽체세포분열(pleuromitosis)"이라고 한다.4. 3. 중심 방추 위치에 따른 분류 (닫힌 가쪽체세포분열의 경우)
닫힌 가쪽체세포분열의 경우, 중심 방추의 위치에 따라 핵바깥과 핵속으로 나뉜다. 중심 방추가 세포질에 위치하면 "핵바깥"이라 하고, 중심 방추가 핵에 위치하면 "핵속"이라고 한다.5. 체세포분열의 오류 및 변이
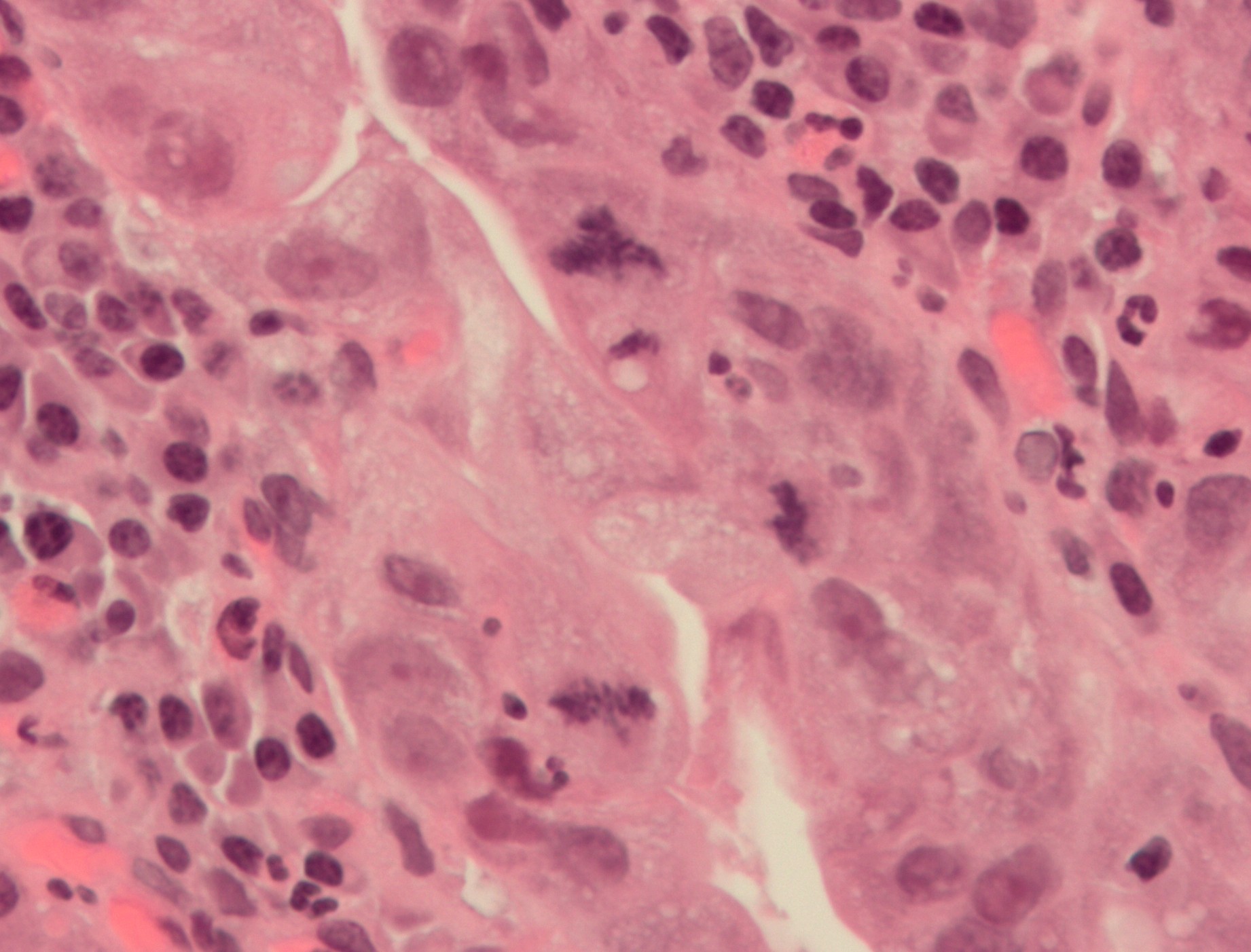
체세포분열 과정에서 오류가 발생하면 염색체 수에 이상이 있는 세포가 생길 수 있다. 이러한 오류는 특히 사람의 초기 배아 발생 과정에서 나타날 수 있으며, 암과도 관련이 있다.[108][110] 체세포분열의 각 단계에는 정상적인 분열을 조절하는 체크포인트가 존재하지만, 드물게 실수가 발생할 수 있다.[109]
체세포분열 과정에서 발생할 수 있는 주요 오류는 다음과 같다:
- '''비분리''': 후기 때 자매 염색분체가 분리되지 않는 현상.
- '''후기 지연''': 후기 동안 염색체의 이동이 지연되는 현상.
- '''속재복제''': 염색체가 복제되었지만 세포가 분열하지 않는 현상.
- '''속체세포분열''': 세포가 염색체를 복제한 뒤 체세포분열에 들어가지만, 완료하지 못하고 다시 염색체를 복제하는 과정.
- '''무사분열''': 섬모충과 동물의 태반 조직에서 일어나는 현상으로, 부모의 대립유전자를 무작위로 배분.
이 외에도 세포질분열 없이 핵분열만 일어나 다핵세포를 만들기도 한다.
5. 1. 비분리 (Nondisjunction)
'''비분리'''(非分離, Nondisjunction)는 세포 분열 후기 때 자매 염색분체가 제대로 분리되지 않아, 딸세포에 염색체 수 이상이 생기는 현상이다.[111] 비분리가 일어나면 하나의 딸세포는 비분리된 염색체의 자매 염색분체를 모두 물려받고, 다른 딸세포는 염색체를 아예 물려받지 못한다. 그 결과, 염색체를 많이 받은 딸세포는 특정 염색체를 3개 가지게 되어 세염색체증(trisomy)이 되고, 염색체를 받지 못한 딸세포는 해당 염색체가 1개뿐인 홑염색체증(monosomy)이 된다. 때로는 비분리된 세포가 세포질분열에 실패하여 이핵 세포(binucleated cell)를 형성하기도 한다.[112] 이러한 체세포 분열 오류는 염색체 수가 정상이 아닌 이수성 세포를 만들어 암 발생과 관련되기도 한다.5. 2. 후기 지연 (Anaphase lag)
후기 동안 염색체의 이동이 지연되면 ''후기 지연''이 일어난다.[111] 이는 방추체가 염색체에 제대로 부착하지 않았을 때 나타난다. 지연이 일어나는 염색체는 두 딸핵에 들어가지 못하고 소실된다. 그러므로 딸세포 중 하나는 그 염색체에 대해 홑염색체증이 된다.5. 3. 속재복제 (Endoreduplication)
속재복제(endoreduplication)는 염색체가 복제된 후 세포 분열이 일어나지 않는 현상이다. 이는 다배수체 세포를 만들며, 염색체가 반복해서 복제되면 다섬유 염색체(polytene chromosome)가 만들어진다.[111][113] 속재복제는 많은 종에서 정상적인 발생 과정의 일부로 나타난다.[113]5. 4. 속체세포분열 (Endomitosis)
속체세포분열(endomitosis)은 속재복제의 한 변이로, 세포가 S기 때 염색체를 복제한 뒤 체세포분열에 들어가지만 성숙하지 않은 상태로 체세포분열을 끝내는 과정이다.[114] 복제된 염색체는 2개의 새로운 딸핵으로 나뉘지 않고 원래의 핵에 그대로 남는다.[114] 이후 세포는 다시 G1기와 S기로 들어가 염색체를 또 복제한다.[114] 이 과정은 여러 번 반복될 수 있으며, 복제와 속체세포분열 과정이 끝날 때마다 염색체 수가 늘어난다. 혈소판을 만드는 거대핵세포는 세포 분화 과정에서 속체세포분열을 거친다.[115]5. 5. 무사분열 (Amitosis)
섬모충과 동물의 태반 조직에서 일어나는 무사분열은 부모의 대립유전자를 무작위로 배분한다.[111]5. 6. 세포질분열 없는 핵분열
세포질분열 없이 핵분열만 일어나면 다핵세포 (공세포)를 만든다.[51]6. 체세포분열 관련 세포 과정
동물 조직에서 대부분의 세포는 체세포분열을 할 동안 구 모양에 가깝게 동그랗게 변한다. 상피와 표피에서 효율적인 라운딩 과정은 방추체를 적절하게 배열하고 딸세포를 올바르게 위치하는 것과 관련이 있다. 라운딩이 억제된 세포에서는 방추 결함, 주요 극 쪼개짐 결함, 염색체를 효율적으로 잡지 못하는 현상 등이 나타난다. 따라서 체세포분열 세포 라운딩은 정확한 체세포분열에 필수적이다.
체세포분열 세포를 G1기에 X선으로 피폭시키면, 세포는 두 상동 염색체 사이의 재조합으로 DNA를 수선한다.[118] 반면 G2기에 피폭시키면 두 자매 염색분체 사이의 재조합을 우선적으로 사용한다.[118] 재조합 관련 효소 유전자에 돌연변이가 발생하면, 여러 DNA 손상 물질에 의해 세포가 죽을 수 있는 민감성이 높아진다.[119][120][121] 이는 체세포분열 재조합이 잠재적으로 치명적일 수 있는 DNA 손상을 복구하는 적응 과정임을 시사한다.
6. 1. 세포 라운딩 (Cell rounding)
동물 조직에서 대부분의 세포는 체세포분열을 하는 동안 구 모양에 가깝게 동그랗게 변한다. 상피와 표피에서 효율적인 라운딩 과정은 방추체를 적절하게 배열하고 딸세포를 올바르게 위치하는 것과 관련이 있다. 또한, 연구자들은 라운딩이 억제된 세포에서는 방추 결함과 주요 극 쪼개짐에서 결함이 일어나고, 염색체를 효율적으로 잡지 못하는 것을 발견했다. 그러므로 체세포분열 세포 라운딩은 정확하게 체세포분열을 하기 위해 필요하다.라운딩을 가능하게 하는 힘은 F-액틴과 마이오신이 수축하는 균질의 세포 겉질로 재배열하는 과정을 통해 발생한다. 그 첫 번째 과정은 세포 주변을 굳게 하는 것이고, 두 번째 과정은 사이기보다 10배 높게 세포 내부의 정수압을 유발하는 것이다. 세포 내부의 높아진 압력은 세포 외부로 힘이 작용하게 하여 주변의 세포나 세포바깥 바탕질을 밀어내면서 세포를 둥글게 만든다. 이 점은 세포끼리 얽매여 있는 조직 시나리오에 특히 중요하다. 압력이 높아지는 것은 포민-매개 F-액틴 핵생성(formin-mediated F-actin nucleation)과 Rho 카이네이스-매개 마이오신 II 수축(Rho kinase-mediated myosin II contraction)에 의존적이다. 두 과정 모두 상류의 신호전달 경로인 RhoA와 ECT2의 지배를 Cdk1의 활성을 거쳐 받는다. 체세포분열에서 라운딩은 그만큼 중요하기 때문에 체세포분열 세포 겉질에서의 분자 성분과 역학은 활발한 연구 분야이다.
6. 2. 체세포 재조합 (Mitotic recombination)
체세포분열을 할 세포를 G1기에 X선으로 피폭시키면, 세포는 두 상동 염색체 사이에서 나타나는 재조합으로 DNA를 수선한다.[118] 반면 G2기에 피폭시키면 두 자매 염색분체 사이에서 나타나는 재조합을 우선적으로 사용한다.[118] 재조합에 관여하는 효소의 유전자에 돌연변이가 일어나면, 여러 DNA 손상 물질에 의해 세포가 죽을 수 있는 민감성이 높아진다.[119][120][121] 이는 체세포분열 재조합이 잠재적으로 치명적일 수 있는 DNA 손상을 복구시키는 적응 과정을 암시한다.세포 주기의 G1기에 X선을 조사한 체세포 분열 세포는 상동 재조합을 통해 재조합 유발 DNA 손상을 주로 복구한다.[59] G2기에 조사된 체세포 분열 세포는 자매 염색 분체 교환을 통해 이러한 손상을 우선적으로 복구한다.[59] 재조합에 사용되는 효소를 암호화하는 유전자의 돌연변이는 세포가 다양한 DNA 손상제에 의해 사멸될 가능성에 대한 민감도를 증가시킨다.[60][61][62] 이러한 연구 결과는 체세포 분열 재조합이 잠재적으로 치명적인 손상을 포함한 DNA 손상을 복구하기 위한 적응임을 시사한다.
7. 진화
진핵생물의 체세포분열 과정에 등장하는 액틴이나 튜뷸린 같은 분자들은 원핵생물에도 대응하는 상동 분자가 존재한다. 체세포분열이 진핵생물의 보편적 특징이라는 점에서 진핵생물 계통수의 기저에서 발생했을 것으로 추정된다. 또한 체세포분열은 감수분열보다 덜 복잡하기 때문에 감수분열이 체세포분열 이후에 발생했을 수 있다.[122] 그러나 감수분열을 포함하는 유성 생식 또한 진핵생물의 원시적 특징이다.[123] 따라서 감수분열과 체세포분열은 조상 원핵생물 과정에서 병행적으로 진화했을 수 있다.
박테리아의 세포 분열에서 DNA가 복제되고 나면, 2개의 원형 염색체는 세포막의 특별한 부위에 부착된다. 반면에 진핵생물의 체세포분열에는 여러 선형 염색체가 존재하고, 염색체의 방추부착점에는 방추의 미세관이 부착한다. 체세포분열의 여러 형태를 살펴보면 닫힌 핵속 가쪽체세포분열(closed intranuclear pleuromitosis)이 박테리아의 분열과 비슷한 가장 원시적인 유형으로 보인다.
8. 진단 표지자
조직병리학에서 체세포 분열률(체세포 분열 수 또는 체세포 분열 지수)은 다양한 유형의 조직 샘플에서 종양의 공격성, 즉 악성도를 구체화하고 진단하기 위한 중요한 지표이다. 예를 들어, 유방암 분류에서 체세포 분열 수를 일상적으로 정량화한다.[56] 체세포 분열은 체세포 분열 활동이 가장 높은 영역에서 계산해야 한다. 그러나 체세포 분열 활동이 매우 높은 종양에서는 이러한 영역을 시각적으로 식별하는 것이 어렵다.[57]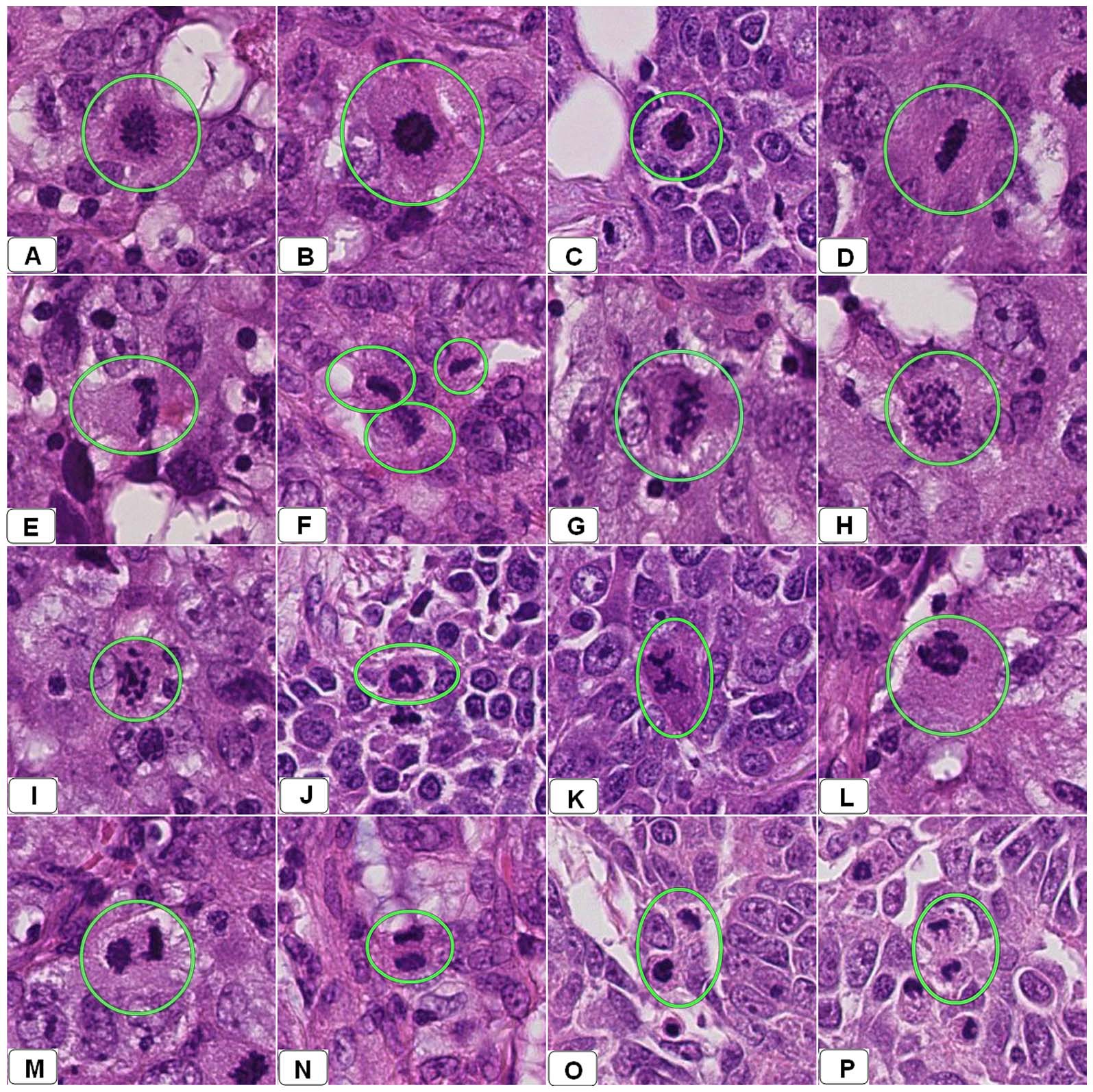
또한, 비정형 체세포 분열 형태의 발견은 진단 및 예후 지표로 모두 사용될 수 있다. 예를 들어, '지연형 체세포 분열'(체세포 분열 구조 영역에서 부착되지 않은 응축된 염색질)은 고위험 인간 유두종 바이러스 감염 관련 자궁경부암을 나타낸다. 체세포 분열 수의 재현성과 정확성을 향상시키기 위해 딥 러닝 기반 알고리즘을 사용한 자동 이미지 분석이 제안되었으나,[58] 이러한 알고리즘을 일상적인 진단에 사용하기 전에는 추가적인 연구가 필요하다.
참조
[1]
웹사이트
Cell division and growth
https://www.britanni[...]
ENCYCLOPÆDIA BRITANNICA
2018-11-04
[2]
웹사이트
Mitosis
http://biology.clc.u[...]
2014-01-14
[3]
웹사이트
Mitosis - an overview {{!}} ScienceDirect Topics
https://www.scienced[...]
2020-11-24
[4]
웹사이트
Cell Division: Stages of Mitosis {{!}} Learn Science at Scitable
http://www.nature.co[...]
2015-11-16
[5]
논문
Image-based analysis of living mammalian cells using label-free 3D refractive index maps reveals new organelle dynamics and dry mass flux
2019-12
[6]
논문
Tripolar mitosis in human cells and embryos: occurrence, pathophysiology and medical implications
2015-01
[7]
논문
On the road to cancer: aneuploidy and the mitotic checkpoint
2005-10
[8]
논문
The cell biology of open and closed mitosis
2013
[9]
서적
Cell Biology
https://books.google[...]
APH Publishing
[10]
간행물
Ueber die Vermehrung der Pflanzenzellen durch Theilung
https://books.google[...]
Tübingen
1835
[11]
문서
NDB|17|690|691|Mohl, Hugo von|Karl Mägdefrau|118830538
[12]
뉴스
"Notes and memoranda: The late professor von Mohl"
https://books.google[...]
Quarterly Journal of Microscopical Science
1875
[13]
웹사이트
150 Years of cell division
https://www.derm101.[...]
Dermatopathology: Practical & Conceptual
2002
[14]
논문
Kilka słów o doktorze Wacławie Mayzlu i jego odkryciu
http://ptbk.mol.uj.e[...]
[15]
서적
Dzieje nauki polskiej
Wydawnictwo Interpress
[16]
웹사이트
Human Anatomy & Physiology I: A Chronology of the Description of Mitosis
http://facstaff.cbu.[...]
Christian Brothers University
2018-05-02
[17]
간행물
Beiträge zur Kenntnis der freilebenden Nematoden
https://www.biodiver[...]
Nova Acta der Kaiserlich Leopoldinisch-Carolinischen Deutschen Akademie der Naturforscher
1873
[18]
간행물
Studien über die ersten Entwicklungsvorgänge der Eizelle, die Zelleilung und die Conjugation der Infusorien
https://www.biodiver[...]
Abh.d. Senckenb. Naturf. Ges. Frankfurt a. M.
1876
[19]
논문
Otto Bütschli (1848–1920) Where we will genuflect?
http://protistology.[...]
[20]
서적
Introduction To Cytology
https://archive.org/[...]
McGraw Hill Book Company Inc.
[21]
웹사이트
mitosis
http://www.etymonlin[...]
2019-11-12
[22]
문서
LSJ|mi/tos|μίτος|ref
[23]
논문
Caryoneme alternative to chromosome and a new caryological nomenclature.
http://www.caryologi[...]
2009
[24]
논문
Die Knorpelzelltheilung
https://www.biodiver[...]
1878
[25]
웹사이트
Karyokinesis
http://www.biologica[...]
2018-05-02
[26]
논문
Embryological questions: 12. Have the ''Polygonum'' and ''Allium'' types been rightly established?
1987
[27]
서적
Botany: an Introduction to Plant Biology
https://books.google[...]
Saunders College Publishing
1991
[28]
서적
Meiosis and Fertilization
https://www.ncbi.nlm[...]
2000
[29]
서적
The Human Genome
https://www.ncbi.nlm[...]
Wiley-Liss
2002
[30]
웹사이트
Mitosis
https://www.biologyo[...]
2020-04-28
[31]
논문
Regulation of repair pathway choice at two-ended DNA double-strand breaks
[32]
논문
Injection of anticentromere antibodies in interphase disrupts events required for chromosome movement at mitosis
1990-10-01
[33]
서적
Biology of Plants
https://archive.org/[...]
W. H. Freeman and Co.
[34]
논문
Sequential entry of components of the gene expression machinery into daughter nuclei
2003-03
[35]
논문
Mitotic bookmarking by transcription factors
2013-04
[36]
논문
Synthesis of RNA and protein during mitosis in mammalian tissue culture cells
1962-03
[37]
서적
The Nucleolus
Springer Science & Business Media
2011
[38]
논문
Flies without centrioles
2006-06
[39]
논문
Molecular architecture of the kinetochore-microtubule interface
2008-01
[40]
웹사이트
Chromosome condensation through mitosis
https://www.scienced[...]
2020-10-04
[41]
논문
Anaphase B precedes anaphase A in the mouse egg
http://discovery.ucl[...]
2019-09-17
[42]
서적
The Cell: A Molecular Approach. 2nd edition.
Sinauer Associates
[43]
논문
Measurement of Red Cell Lifespan and Aging
2012-08-27
[44]
웹사이트
Archaea
https://eol.org/docs[...]
2011-08-23
[45]
웹사이트
Binary Fission and other Forms of Reproduction in Bacteria
https://cals.cornell[...]
[46]
논문
The cell biology of open and closed mitosis
[47]
간행물
Key Transitions in Animal Evolution.
https://books.google[...]
CRC Press
2010
[48]
논문
Molecular origin of mitotic aneuploidies in preimplantation embryos
2012-12
[49]
논문
Mitotic checkpoints: from yeast to cancer
http://www.sciencedi[...]
2001-02-01
[50]
논문
Short- and long-term effects of chromosome mis-segregation and aneuploidy
2015-08
[51]
서적
International Review Of Cytology: A Survey of Cell Biology
Academic Press
2006
[52]
논문
Chromosome nondisjunction yields tetraploid rather than aneuploid cells in human cell lines
2005-10
[53]
논문
Endoreplication cell cycles: more for less
2001-05
[54]
논문
Endoreplication: polyploidy with purpose
2009-11
[55]
논문
Endomitosis of human megakaryocytes are due to abortive mitosis
1998-05
[56]
웹사이트
Infiltrating Ductal Carcinoma of the Breast (Carcinoma of No Special Type)
http://surgpathcrite[...]
2019-10-02
[57]
논문
Computerized Calculation of Mitotic Count Distribution in Canine Cutaneous Mast Cell Tumor Sections: Mitotic Count Is Area Dependent
https://boris.unibe.[...]
2020-03
[58]
논문
Computer-assisted mitotic count using a deep learning–based algorithm improves interobserver reproducibility and accuracy
[59]
논문
Sister chromatids are preferred over homologs as substrates for recombinational repair in Saccharomyces cerevisiae
1992-10
[60]
논문
rad51 leads to Fanconi anemia-like symptoms in zebrafish
2017-05
[61]
논문
p53 is linked directly to homologous recombination processes via RAD51/RecA protein interaction
1996-04
[62]
논문
Rad51-deficient vertebrate cells accumulate chromosomal breaks prior to cell death
1998-01
[63]
논문
The evolution of meiosis from mitosis
2009-01
[64]
문서
Bernstein, H., Bernstein, C. Evolutionary origin and adaptive function of meiosis. In "Meiosis", Intech Publ (Carol Bernstein and Harris Bernstein editors), Chapter 3: 41-75 (2013).
[65]
웹인용
Cell division and growth
https://www.britanni[...]
ENCYCLOPÆDIA BRITANNICA
2018-11-04
[66]
웹인용
4.1: Meiosis
https://bio.libretex[...]
2021-05-29
[67]
웹인용
Explain why mitosis is called equational and meiosis class 11 biology CBSE
https://www.vedantu.[...]
2021-05-29
[68]
웹인용
Mitosis
http://biology.clc.u[...]
2019-11-12
[69]
웹인용
Mitosis - an overview {{!}} ScienceDirect Topics
https://www.scienced[...]
2020-11-24
[70]
웹인용
Cell Division: Stages of Mitosis {{!}} Learn Science at Scitable
http://www.nature.co[...]
2015-11-16
[71]
저널
Image-based analysis of living mammalian cells using label-free 3D refractive index maps reveals new organelle dynamics and dry mass flux
2019-12
[72]
저널
Tripolar mitosis in human cells and embryos: occurrence, pathophysiology and medical implications
2015-01
[73]
저널
On the road to cancer: aneuploidy and the mitotic checkpoint
2005-10
[74]
학위논문
Ueber die Vermehrung der Pflanzenzellen durch Theilung
https://books.google[...]
Tübingen
1835
[75]
NDB
Mohl, Hugo von
[76]
간행물
Notes and memoranda: The late professor von Mohl
https://books.google[...]
1875
[77]
간행물
150 Years of cell division
https://www.derm101.[...]
2002
[78]
저널
Kilka słów o doktorze Wacławie Mayzlu i jego odkryciu
http://ptbk.mol.uj.e[...]
[79]
서적
Dzieje nauki polskiej
Wydawnictwo Interpress
[80]
웹사이트
Human Anatomy & Physiology I: A Chronology of the Description of Mitosis
http://facstaff.cbu.[...]
2018-05-02
[81]
저널
Beiträge zur Kenntnis der freilebenden Nematoden
https://www.biodiver[...]
1873
[82]
저널
Studien über die ersten Entwicklungsvorgänge der Eizelle, die Zelleilung und die Conjugation der Infusorien
https://www.biodiver[...]
1876
[83]
저널
Otto Bütschli (1848–1920) Where we will genuflect?
http://protistology.[...]
2014-08-06
[84]
서적
Introduction To Cytology
https://archive.org/[...]
McGraw Hill Book Company Inc.
[85]
웹사이트
mitosis
http://www.etymonlin[...]
2019-11-12
[86]
LSJ
μίτος
[87]
저널
Die Knorpelzelltheilung
https://www.biodiver[...]
1878
[88]
웹인용
Karyokinesis
http://www.biologica[...]
2018-05-02
[89]
저널
Embryological questions: 12. Have the Polygonum and Allium types been rightly established?
1987
[90]
저널
Caryoneme alternative to chromosome and a new caryological nomenclature.
http://www.caryologi[...]
2009
[91]
서적
Botany: an Introduction to Plant Biology
https://books.google[...]
Saunders College Publishing
1991
[92]
저널
Meiosis and Fertilization
https://www.ncbi.nlm[...]
2000
[93]
서적
The Human Genome
https://www.ncbi.nlm[...]
Wiley-Liss
2002
[94]
웹인용
Mitosis
https://www.biologyo[...]
[95]
저널
Regulation of repair pathway choice at two-ended DNA double-strand breaks
[96]
저널
Injection of anticentromere antibodies in interphase disrupts events required for chromosome movement at mitosis
1990-10-01
[97]
서적
Biology of Plants
https://archive.org/[...]
W. H. Freeman and Co.
[98]
저널
Sequential entry of components of the gene expression machinery into daughter nuclei
2003-03
[99]
저널
Mitotic bookmarking by transcription factors
2013-04
[100]
저널
Synthesis of RNA and protein during mitosis in mammalian tissue culture cells
1962-03
[101]
서적
The Nucleolus
Springer Science & Business Media
2011
[102]
저널
Flies without centrioles
2006-06
[103]
저널
Molecular architecture of the kinetochore-microtubule interface
2008-01
[104]
웹인용
Chromosome condensation through mitosis
https://www.scienced[...]
2020-10-04
[105]
저널
Anaphase B precedes anaphase A in the mouse egg
http://discovery.ucl[...]
2019-09-17
[106]
저널
The cell biology of open and closed mitosis
[107]
서적
Key Transitions in Animal Evolution.
https://books.google[...]
CRC Press
2010
[108]
저널
Molecular origin of mitotic aneuploidies in preimplantation embryos
2012-12
[109]
저널
Mitotic checkpoints: from yeast to cancer
http://www.sciencedi[...]
2001-02-01
[110]
저널
Short- and long-term effects of chromosome mis-segregation and aneuploidy
2015-08
[111]
서적
International Review Of Cytology: A Survey of Cell Biology
Academic Press
2006
[112]
저널
Chromosome nondisjunction yields tetraploid rather than aneuploid cells in human cell lines
2005-10
[113]
저널
Endoreplication cell cycles: more for less
2001-05
[114]
저널
Endoreplication: polyploidy with purpose
2009-11
[115]
저널
Endomitosis of human megakaryocytes are due to abortive mitosis
https://archive.org/[...]
1998-05
[116]
웹인용
Infiltrating Ductal Carcinoma of the Breast (Carcinoma of No Special Type)
http://surgpathcrite[...]
2019-10-02
[117]
저널
Computerized Calculation of Mitotic Count Distribution in Canine Cutaneous Mast Cell Tumor Sections: Mitotic Count Is Area Dependent
https://boris.unibe.[...]
2020-03
[118]
저널
Sister chromatids are preferred over homologs as substrates for recombinational repair in Saccharomyces cerevisiae
1992-10
[119]
저널
rad51 leads to Fanconi anemia-like symptoms in zebrafish
2017-05
[120]
저널
p53 is linked directly to homologous recombination processes via RAD51/RecA protein interaction
1996-04
[121]
저널
Rad51-deficient vertebrate cells accumulate chromosomal breaks prior to cell death
1998-01
[122]
저널
The evolution of meiosis from mitosis
2009-01
[123]
문서
Evolutionary origin and adaptive function of meiosis
2013
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com