크로노사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
크로노사우루스는 플리오사우루스과에 속하는 대형 해양 파충류이다. 몸길이는 9~10.5m에 달하며, 2.85m에 달하는 큰 두개골과 날카로운 원뿔형 이빨을 가지고 있었다. 어류, 두족류, 다른 해양 파충류를 잡아먹는 최상위 포식자였으며, 동족을 공격하거나 엘라스모사우루스과인 에로망가사우루스를 공격하기도 했다. 크로노사우루스는 브라키오케니우스아과에 속하며, 쥐라기 이후 살아남은 탈라소포네아의 한 계통으로 여겨진다. 백악기 전기 호주의 에로망가 해에 서식했으며, 다양한 해양 생물과 공룡, 익룡 등과 공존했다.
더 읽어볼만한 페이지
| 크로노사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| |
| 학명 | Kronosaurus |
| 명명자 | 롱먼, 1924년 |
| 모식종 | Kronosaurus queenslandicus |
| 모식종 명명자 | 롱먼, 1924년 |
| 이명 | Eiectus longmani ? (Noè & Gómez-Pérez, 2022) |
| 분류 | |
| 속 | 크로노사우루스속 |
| 생물학적 특징 | |
| 생존 시기 | 압트기-알비기 후기 (~1억 2500만 년 전 ~ 9960만 년 전) |
| 추정 길이 | 13m (43 ft) |
| 최대 추정 길이 | 9m-10m |
| 최소 추정 길이 | 5m |
| 추정 무는 힘 | 15,000~27,000 뉴턴 |
| 학문적 분류 | |
| 속하는 목 | 수장룡목 (Plesiosauria) |
| 속하는 아목 | 플리오사우루스아목 (Pliosauroidea) |
| 속하는 과 | 플리오사우루스과 (Pliosauridae) |
2. 특징
크로노사우루스는 플리오사우루스과에 속하는 거대한 해양 파충류였다. 두개골은 몸 전체 길이의 약 1/4을 차지할 정도로 컸으며, 길게 늘어진 삼각형 모양의 주둥이를 가졌다. 턱에는 길이 25~30cm에 달하는 날카로운 원뿔형 이빨이 여러 개 있었고, 이빨에는 뚜렷한 용골(칼날)이 없었으며 거친 능선이 있었다.[19][18] 위 내용물, 이빨 형태, 턱 구조 등을 바탕으로 어류, 두족류, 다른 해양 파충류(수장룡, 바다거북 등)를 잡아먹는 최상위 포식자였을 것으로 추정된다.
성격이 난폭하여 다른 수장룡을 공격하기도 했으며, 종종 동족끼리도 싸웠을 가능성이 있다. 짧은 꼬리와 네 개의 지느러미를 가지고 있었고, 뒷지느러미가 앞지느러미보다 더 컸다.
2. 1. 크기
크로노사우루스는 지금까지 확인된 가장 큰 플리오사우루스 중 하나이지만, 정확한 크기에 대해서는 여러 추정치가 제시되었다. 알프레드 로머는 최대 몸길이를 12.8m로 추정했지만[55], 2003년 벤자민 키어(Benjamin Kear)는 다른 플리오사우루스류 화석과의 비교를 통해 최대 10m 정도였을 것으로 추정했다[53].1930년 초, 롱먼은 앞다리뼈에 대한 설명을 통해 ''크로노사우루스''가 후기 쥐라기에 살았던 북미 플리오사우루스과인 거대한 ''메갈네우사우루스''의 크기를 능가했을 것이라고 생각했다.[5][4] 하버드 원정대가 화석을 수집한 후, ''크로노사우루스''의 최대 크기는 표본 MCZ 1285를 기반으로 일반적으로 로 설정되었다.[12][13][10] 그러나 하버드 골격 복원에 오류가 있어, 맥헨리는 이 표본의 크기를 길이 로, 무게를 11ton로 더 작게 제시했다.[10] 2009년 맥헨리의 논문이 출판되기 전에도, 고생물학자 벤자민 P. 커와 해양 생물학자 리처드 엘리스는 2003년에 각각 출판된 저서에서 9m(커에 따르면 에서 엘리스에 따르면 )에 이르는 비슷한 추정치를 제시했다. 2024년, 루이즈 제이커번 자오는 MCZ 1285의 측정을 로 수정했다.
다른 표본들에 대한 신체 추정치도 제시되었는데, 일부는 제한적인 화석으로만 알려져 있다. 모식 표본인 QM F1609는 매우 단편적이지만, 길이가 이고 체질량이 1.9ton일 것으로 추정된다. 제안된 신형식 표본인 QM F18827은 길이가 이고 체질량이 6.7ton에 달했을 것이다. 알려진 가장 완전한 표본인 QM F10113은 길이가 이고 체질량이 5.7ton로 약간 더 작았을 것이다. 투르부크 형성에서 발견된 가장 큰 ''크로노사우루스'' 표본인 QM F2446과 QM F2454는 하버드 골격과 거의 동일한 크기였을 것이다. 각각 이 두 표본은 길이가 에 달했고 체질량은 9.9ton[24]에서 15.5ton로 추정되었다.
2. 2. 두개골
''K. 퀸즐란디쿠스''(QM F1609)의 모식 표본은 부분적인 하악 결합부만으로 구성되어 있어 자세한 설명은 어렵다. 그러나 이 분류군에 속하는 더 완전한 화석 두개골은 고유한 특징을 보여준다.[18] 알려진 ''크로노사우루스'' 표본의 두개골 크기는 다양하다. 모식 표본은 부분적이고 조각났지만, 총 1.31m 길이로 측정되었을 두개골에서 나왔다. 후보 신형 표본 QM F10113과 QM F18827은 각각 1.87m에서 1.98m에 달하는 두개골 길이를 가지고 있다. 하버드 골격의 두개골은 2.85m 길이로 추정된다.[31] 이 세 표본의 두개골 측정치는 알려진 어떤 수각류 공룡의 두개골보다 크다.[33] 주둥이와 하악의 돌기는 길고 좁은 형태이며, 일반적으로 아치형으로 보이고 상대적으로 길쭉하며 뚜렷한 중앙 및 등쪽 능선을 가진다. 눈구멍은 뒤쪽을 향하고 있으며, 두개골 앞쪽 절반의 측면에 위치한다. 측두와(temporal fossae)(두개골 뒤쪽의 열린 부분)은 매우 크지만, 앞쪽의 내익상판 사이 구멍은 없다.''크로노사우루스''의 고유한 특징 중 하나는 전상악골(앞쪽 위쪽 치아를 지탱하는 뼈)에 4개의 견치형 치아가 있다는 것이다.[18] 전두골(눈구멍을 경계하는 뼈)은 후전두골과 전두개골 사이의 연결로 인해 눈구멍 가장자리에 닿지 않는다. 전두골은 또한 두정골과 전상악골의 얼굴 돌기 사이의 연결로 인해 두개골 천장의 중간 부분에 닿지 않는다. 전두개골은 크고 눈구멍의 앞쪽 중간 부분과 콧구멍의 뒤쪽 경계에 접촉한다. 누골(눈구멍의 아래쪽 앞쪽 가장자리를 경계하는 뼈)은 작은 표본에서는 나타나지만 성체에서는 융합되는 경향이 있다. 중앙 등쪽 능선의 등쪽 표면은 전상악골과 비골(외부 콧구멍을 경계하는 뼈)에 의해 형성되며, 성체에서는 융합된다. 설골은 튼튼하다.
''크로노사우루스''의 하악 결합부는 길쭉하고 주걱 모양이며, 근연종인 ''브라키오케니우스'' 및 ''메가세팔로사우루스''와 마찬가지로 최대 6쌍의 치아를 포함한다.[18] 각 치골(하악의 치아를 지탱하는 뼈)은 최대 26개의 치아를 가지고 있다. 하악의 관절와(턱 관절의 소켓)는 콩팥 모양이며 위쪽과 안쪽으로 각을 이룬다.[18] ''크로노사우루스'' 치아의 주요 자가형질은 원뿔 모양이며, 대략적인 능선이 있고 뚜렷한 용골이 없다는 것이다.[19][18] ''크로노사우루스''의 치아는 이형치성으로, 다양한 모양의 치아를 가지고 있다. 더 큰 치아는 견치형이며 턱 앞쪽에 위치하고, 더 작은 치아는 더 날카롭게 휘어져 있고 튼튼하며 뒤쪽에 위치한다.[18]
2. 3. 골격
McHenry의 분석에 따르면, ''크로노사우루스''는 최소 35개의 척추뼈를 가졌을 것으로 추정되며, 여기에는 13개의 목뼈와 5개의 등뼈가 포함된다.[32] ''플리오사우루스''와는 달리, 목뼈 중심체(척추뼈 몸통)가 등뼈보다 더 넓다.[32] 그러나 앞쪽 등뼈는 폭보다 높다. 관절돌기는 앞쪽 등뼈와 꼬리뼈에서 육안으로 보이지 않았을 것이다. 흉부 부위의 갈비뼈는 튼튼하며, 단일두(single-headed)였을 것이다. ''크로노사우루스''의 꼬리는 알려져 있지 않지만, 꼬리뼈 끝은 다른 플레시오사우루스처럼 작은 꼬리 지느러미를 지지했을 것이다.[45][46] 부리돌기와 두덩뼈는 앞뒤로 길쭉하다. ''크로노사우루스''의 뒷다리는 앞다리보다 길며, 넙다리뼈가 위팔뼈보다 더 길고 튼튼하다. 이는 ''크로노사우루스''의 가장 큰 개체들이 5m을 초과하는 날개 길이를 형성했을 뒷지느러미를 가졌을 것임을 시사한다.
3. 분류

크로노사우루스는 1899년 오스트레일리아 퀸즐랜드주 휴겐든 근처에서 발견된 화석을 바탕으로, 1924년 앨버트 헤버 롱맨이 *Kronosaurus queenslandicus*라는 학명으로 명명했다. 속명은 그리스 신화의 티탄족인 크로노스에서 유래했으며, 종명은 화석 발견지인 퀸즐랜드를 의미한다.[2][3]
모식 표본(QM F1609)은 6개의 원뿔형 이빨이 있는 부분적인 하악 이음부로, 불완전성 때문에 한때 의문명(nomen dubium)으로 간주되었다.[2] 1962년 새뮤얼 폴 웰스는 크로노사우루스를 ''무효명''으로 보았다.[34]
이후 오스트레일리아 각지에서 크로노사우루스 추정 화석들이 발견되었고, 2009년 맥헨리는 툴레북 형성에서 발견된 두 부분 골격(QM F10113, QM F18827)을 신모식표본 후보로 제안했다.[15][16][17][18] 그러나 2022년 에이엑투스 롱가니라는 새로운 분류군으로 재분류하는 논문이 나왔고,[20] 2023년에는 이에 대한 비판과 함께 QM F18827을 신모식표본으로 권장하는 논문이 발표되었다.[22]
콜롬비아에서 발견된 화석은 *Kronosaurus boyacensis*로 명명되었으나,[25] 현재는 몬키라사우루스(''Monquirasaurus'')라는 별개의 속으로 재분류되었다.[26]
3. 1. 계통 발생

크로노사우루스는 플리오사우루스과에 속하며, 그 안에서도 브라키오케니우스아과에 속하는 것으로 보인다. 브라키오케니우스아과는 브라키오케니우스, 메가케팔로사우루스와 가까운 관계를 가지는 분지군이다.[37] 이 아과는 "전형적인" 짧은 목을 가진 플리오사우루스과를 포함하며, 쥐라기 이후 살아남은 탈라소포네아의 한 계통으로 여겨진다.[37]
아래의 계통도는 Madzia 외.(2018)에서 수정된 것이다.[40]
브라키오케니우스아과는 백악기에서 유래된 플리오사우루스과의 대다수를 포함한다. 하지만, 쥐라기 이후 살아남은 탈라소포네아의 유일한 계통은 아닐 수도 있다. 브라키오케니우스아과와는 다른 특징을 보이는 하부 백악기 플리오사우루스 이빨은 적어도 다른 한 계통이 쥐라기-백악기 경계를 넘었음을 시사한다.[18][40][41]
브라키오케니우스아과 구성원들은 다양하며, 모든 구성원을 연결하는 단 하나의 공유파생형질은 쥐라기의 일부 플리오사우루스에서 보이는 완전하거나 다소 삼면체 모양의 이빨 대신 다소 원형의 이빨을 가지고 있다는 것이다. ''메가케팔로사우루스''와 같은 대부분의 브라키오케니우스류는 더 작은 먹이에 적응된 두개골 특징(예: 길쭉한 주둥이, 날씬한 주둥이, 일정한 크기의 이빨)을 공유한다. 그러나 ''크로노사우루스''는 이러한 특징을 공유하지 않고, 모양이 다른 이빨을 가진 이 그룹의 몇 안 되는 대표자 중 하나이다.[41] 따라서 이러한 유형의 치아는 ''크로노사우루스''가 다른 대부분의 이 그룹의 대표와 달리 큰 먹이를 사냥하는 데 특화된 속임을 나타낸다.[41][18]
4. 연구사
1899년, 호주 퀸즐랜드에서 크로노사우루스의 첫 번째 화석(부분적인 아래턱)이 발견되었다. 이 화석은 퀸즐랜드 박물관의 찰스 드 비스에 의해 플레시오사우루스와 어룡을 포함하는 Enaliosauria의 대표적인 표본으로 생각되었다. 하지만, 이 표본의 독특한 치열은 드 비스가 생각을 바꾸게 만들었다.[2]
1924년, 앨버트 헤버 롱맨은 앤드루 크롬비가 발견한 화석을 바탕으로 *Kronosaurus queenslandicus*라는 학명을 부여했다. 속명은 그리스 신화의 티탄인 크로노스와 고대 그리스어 σαῦρος (''saûros'', "도마뱀")에서 유래하여 "크로노스의 도마뱀"을 의미한다. 롱맨은 크로노스가 자신의 자녀를 삼킨 이야기에서 영감을 받아 이 동물의 인상적인 크기와 잠재적 맹렬함을 나타내고자 했다.[9] 종명 ''queenslandicus''는 호주 퀸즐랜드주에서 유래되었으며, 모식표본이 발견되었을 가능성이 가장 높은 곳이다.[9]
1931~1932년, 하버드 대학교 비교동물학 박물관 탐험대는 호주에서 크로노사우루스의 거의 완전한 골격(MCZ 1285)을 발견했다. 이 표본은 이후 크로노사우루스 연구의 중요한 기준이 되었으나, 과도한 복원으로 인해 '플라스터사우루스'라는 별명이 붙기도 했다.[24]
1977년, 콜롬비아에서 또 다른 크로노사우루스 화석이 발견되었고, 1992년에 *Kronosaurus boyacensis*로 명명되었으나,[26] 2022년에 몬키라사우루스로 재분류되었다.[20]
2009년, 콜린 맥헨리는 크로노사우루스에 대한 포괄적인 연구를 발표하여 크기, 해부학적 특징, 분류, 고생태 등에 대한 이해를 넓혔다.
2021년, 일부 연구자들은 하버드 표본을 비롯한 일부 화석을 에이엑투스 롱가니(*Eiectus longani*)라는 새로운 속으로 재분류할 것을 제안했으나,[20] 2023년 연구에서 이 제안은 국제동물명명규약(ICZN)에 위배된다는 비판을 받았다.[22]
5. 고생물학
크로노사우루스는 강력한 턱과 최대 30cm에 달하는 날카로운 이빨을 가진 최상위 포식자였다. 위 내용물 분석과 다른 수장룡을 공격한 흔적을 통해 바다거북, 수장룡(특히 엘라스모사우루스과), 어류, 두족류 등 다양한 해양 동물을 먹이로 삼았음을 알 수 있다.[23] 엘라스모사우루스과는 긴 목과 작은 머리를 가지고 있어 크로노사우루스의 공격에 취약했을 것으로 보인다.
크로노사우루스는 동족 간의 경쟁이나 식인 행위를 했을 가능성도 제기된다. 크로노사우루스의 물기 힘은 매우 강력하여 티라노사우루스와 비교되기도 한다.
5. 1. 서식 환경
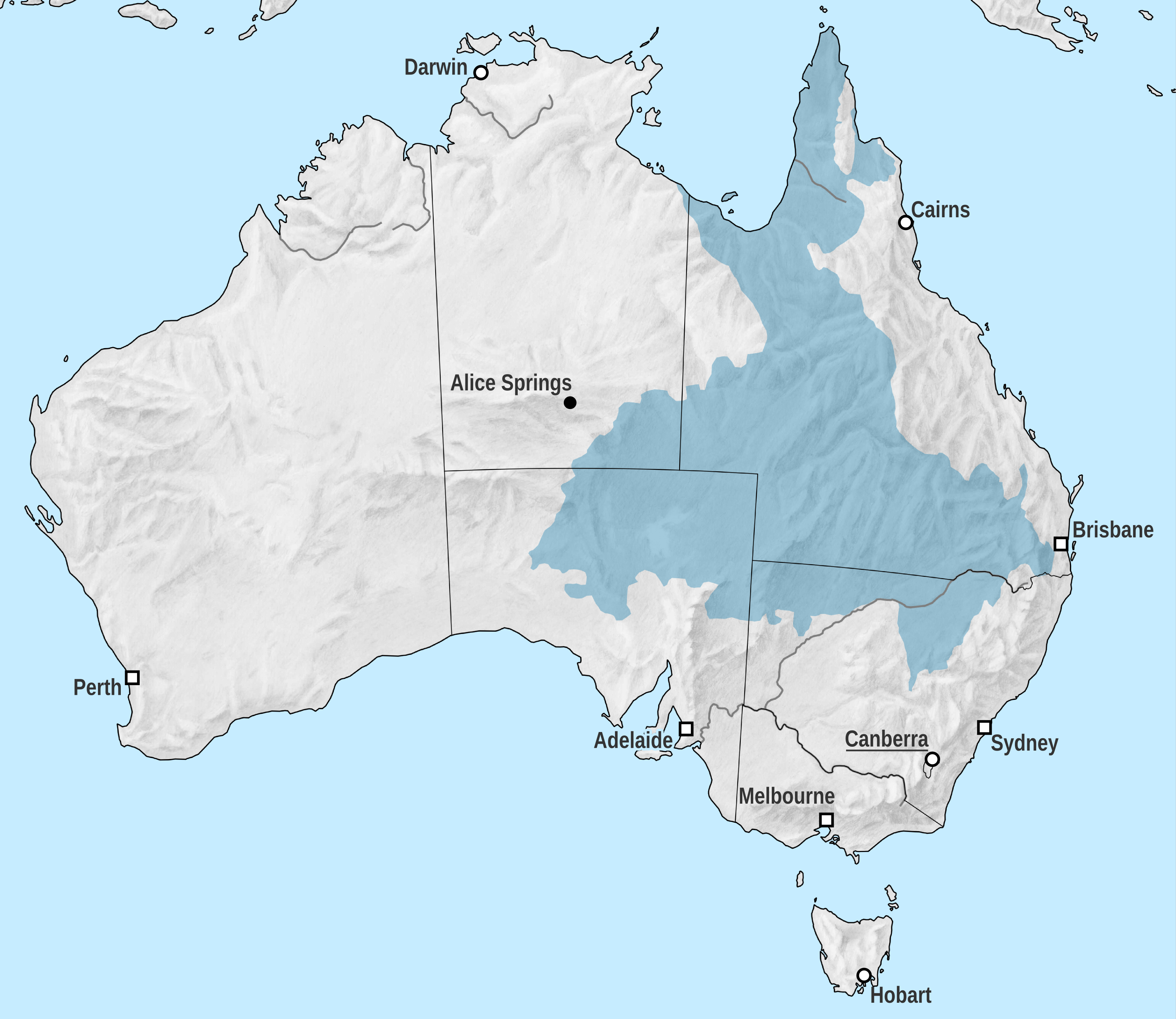
''크로노사우루스'' 화석은 모두 그레이트 아르테시안 분지(GAB)에서 발견되었다.[48] 백악기 전기, 이 지역은 에로망가 해라는 내해에 잠겨 있었다.[48][49] 퇴적 기록에 따르면 이 바다는 비교적 얕고 진흙이 많으며 정체되어 있었다.[48] 에로망가 해의 온도는 특히 춥고 어는점에 가까웠으며,[48][50] 일부 지역에서는 계절적 얼음이 형성되었을 수도 있다.[51] 알비안기 해수 온도는 앱트기보다 더 따뜻했을 것으로 보인다.[52]
GAB의 후기 앱트기~후기 알비안기 화석 기록에는 연체동물을 중심으로 많은 무척추동물이 알려져 있다. 자유 유영 생물에는 암모나이트, 총석기, 오징어 등 두족류가 포함된다. 저서 생물대는 주로 이매패류가 지배하며, 복족류와 굴족류는 다양성이 덜하다. 극피동물의 유경류, 십각 갑각류, 완족류, 다모류 환형동물, 유리 해면 등 다른 무척추동물도 알려져 있다.[52] 에로망가 해의 어류 다양성은 지질 시대에 따라 다른데, 알비안기 지층에서는 많지 않지만 앱트기 기록, 특히 상부 앱트기에서 풍부하다. ''Australopachycormus'', ''Richmondichthys'', ''Flindersichthys'', ''Cooyoo'', ''Pachyrhizondontus'' 등 조기어류와, ''Ceratodus'', ''Neoceratodus'' 등 폐어 (육기어류)가 알려져 있다. 연골어류에는 ''Archaeolamna'', ''Carcharias'', ''Cretolamna'', ''Cretoxyrhina'', ''Edaphodon'', ''Echinorhinus'', ''Leptostyrax'', ''Microcorax'', ''Notorynchus'', ''Pseudocorax'', ''Pristiophorus'', ''Scapanorhynchus'', 여러 종의 오레크톨로보형류 및 팔레오스피나키드가 있다. 이 물고기들은 다양한 크기로 표면, 중간 수심, 저서 등 다양한 생태적 지위를 채웠으며, 일부는 상당히 컸다. 이들은 무척추동물 포식자, 어식 동물, 그리고 ''Cretoxyrhina''의 경우 대형 최상위 포식자였다.

에로망가 해는 해양 파충류가 다양하기로 유명하다.[17] 확인된 바다거북에는 프로토스테기드 ''Cratochelone'', ''Bouliachelys'', ''Notochelone''이 있으며,[17] ''Notochelone''은 내해에서 가장 다양하다. 퀸즐랜드에서는 여러 어룡 화석이 발견되었고, 역사적으로 여러 속으로 분류되었다. 현재 이 화석들은 화석 기록상 가장 젊은 어룡 중 하나인 ''Platypterygius australis''에 속하는 것으로 알려져 있다.[17] 이 종의 다른 화석은 GAB의 다른 지층, 특히 불독 셰일에서 발견되었지만, 명확한 진단을 내리기에는 너무 단편적이다. 여러 플레시오사우루스가 확인되었지만, 대부분 너무 단편적이거나 진단 불가능하여 특정 속이나 종으로 분류할 수 없다.[17] ''크로노사우루스''는 호주에서 층서적으로 가장 광범위한 플레시오사우루스이며,[17] 제안된 속 ''Eiectus''를 제외하면 이 나라에서 알려진 유일한 대형 플리오사우르과이다.[20] 유일하게 알려진 크립토클리두스과는 ''Opallionectes''이다.[19][17] 엘라스모사우루스과에는 ''Eromangasaurus''와 수많은 중간 대표자가 포함된다.[19][17] Leptocleidia 분기군 (Leptocleididae 및 Polycotylidae 포함)의 일부 대표자도 알려져 있다. 렙토클레이드에는 ''Leptocleidus'', ''Umoonasaurus'' 및 불확실한 속의 표본들이 있다.[17] 폴리코틸리드는 아직 설명되지 않았거나 미기재된 표본으로만 알려져 있으며, 가장 주목할 만한 것은 리치몬드 표본이다.[19][17]
에로망가 해 화석 기록에서는 다양한 고룡 그룹도 확인되었다. 용각류 ''Austrosaurus'', 곡룡류 ''Minmi'', 조각류 ''Muttaburrasaurus'' 등, 에로망가 해에서 익사했을 가능성이 있는 공룡들의 단편적인 잔해도 알려져 있다. 공룡 외에도 많은 익룡 화석이 알려져 있는데, 이들은 현대 바닷새와 유사한 포식자였을 것이다. 그러나 익룡 화석은 종종 단편적이라, 명명된 분류군은 소수에 불과하다.[49] ''Aussiedraco'', ''Mythunga'', ''Thapunngaka'' 등이 있다.
5. 2. 경쟁
최상위 포식자였음에도 불구하고, ''크로노사우루스''는 때때로 다른 동시대 포식자들에게 공격을 받기도 했다. 실제로, KK F0630으로 분류된 턱뼈는 크고 덜 성숙한 개체 또는 작은 성체 표본일 가능성이 있는데, 이는 크레톡시리나이과에 속하는 람니형 상어에 의해 만들어진 물린 자국을 보여준다. 이러한 유형의 부상은 드문 일이 아닌데, 이 과에 속하는 여러 상어들이 ''크로노사우루스''가 알려진 다양한 지질 형성에서 확인되었기 때문이다. 물린 자국 주위에 비정상적으로 융기된 골성장이 있는 홈은 이 표본이 생전에 치유되었음을 나타낸다.[18]참조
[1]
웹사이트
"''Kronosaurus''"
http://www.paleofile[...]
[2]
논문
A new gigantic marine reptile from the Queensland Cretaceous, ''Kronosaurus queenslandicus'' new genus and species
https://www.biodiver[...]
[3]
서적
A Time for a Museum : The History of the Queensland Museum 1862-1986
https://archive.org/[...]
Memoirs of the Queensland Museum
[4]
웹사이트
Ben Creisler's Plesiosaur Pronunciation Guide
http://oceansofkansa[...]
2021-06-26
[5]
논문
"''Kronosaurus queenslandicus'' : A Gigantic Cretaceous Pliosaur"
https://www.biodiver[...]
[6]
논문
Restoration of ''Kronosaurus queenslandicus''
https://www.biodiver[...]
[7]
논문
Palæontological notes
https://www.biodiver[...]
[8]
논문
The Harvard Museum Expedition to Australia
[9]
서적
About the Exhibits
https://archive.org/[...]
Museum of Comparative Zoology
[10]
서적
The Rarest of the Rare : Stories Behind the Treasures at the Harvard Museum of Natural History
https://archive.org/[...]
HarperResource
[11]
논문
On the skull of ''Kronosaurus queenslandicus'' Longman
https://archive.org/[...]
[12]
논문
A mounted skeleton of the giant plesiosaur ''Kronosaurus''.
https://www.biodiver[...]
[13]
논문
"''Kronosaurus queenslandicus'': Ancient Monarch of the Seas"
http://oceansofkansa[...]
[14]
논문
A Giant Plesiosaur
[15]
논문
Marine reptiles from the Lower Cretaceous (Aptian) deposits of White Cliffs, southeastern Australia: implications of a high latitude, cold water assemblage
https://doc.rero.ch/[...]
[16]
논문
Plesiosaur remains from Cretaceous high-latitude non-marine deposits in southeastern Australia
[17]
논문
Cretaceous marine amniotes of Australia: perspectives on a decade of new research
[18]
논문
The mandible of ''Kronosaurus queenslandicus'' Longman, 1924 (Pliosauridae, Brachaucheniinae), from the Lower Cretaceous of northwest Queensland, Australia
https://www.research[...]
[19]
논문
Marine reptiles from the Lower Cretaceous of South Australia: elements of a high-latitude cold-water assemblage
[20]
논문
Giant pliosaurids (Sauropterygia; Plesiosauria) from the Lower Cretaceous peri-Gondwanan seas of Colombia and Australia
[21]
웹사이트
Article 75. Neotypes
https://code.iczn.or[...]
[22]
논문
Anatomy and relationships of the bizarre Early Cretaceous pliosaurid ''Luskhan itilensis''
[23]
논문
An elasmosaur bitten by a pliosaur
https://www.research[...]
[24]
논문
Applications of computed tomography to fossil conservation and education
[25]
논문
Noticia preliminar sobre el hallazgo de un presunto ''Kronosaurus'' (Reptilia: Dolichorhynchopidae) en el Aptiano superior de Villa de Leiva, Colombia
[26]
논문
Ein großwüchsiger Pliosauride (Reptilia: Plesiosauria) aus der Unterkreide (oberes Aptium) von Kolumbien
[27]
논문
A new large Pliosaurid from the Barremian (Lower Cretaceous) of Sáchica, Boyacá, Colombia
[28]
논문
Anatomy and functional morphology of the largest marine reptile known, ''Mosasaurus hoffmanni'' (Mosasauridae, Reptilia) from the Upper Cretaceous, Upper Maastrichtian of The Netherlands
https://royalsociety[...]
[29]
논문
A giant mosasaur (Reptilia, Squamata) with an unusually twisted dentition from the Argille Scagliose Complex (late Campanian) of Northern Italy
https://www.disva.un[...]
[30]
논문
The last giants: New evidence for giant Late Triassic (Rhaetian) ichthyosaurs from the UK
[31]
논문
A Giant Pliosaurid Skull from the Late Jurassic of England
[32]
논문
A new species of ''Pliosaurus'' (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway
https://njg.geologi.[...]
[33]
웹사이트
Sauropterygia: Pliosauroidea: Pliosauridae
http://palaeos.com/v[...]
[34]
논문
A review of the short-necked plesiosaurs from the Cretaceous of the Western Interior, North America
https://www.research[...]
[35]
서적
The osteology of the reptiles
https://www.biodiver[...]
Harvard University Press
[36]
논문
The skull of the giant predatory pliosaur ''Rhomaleosaurus cramptoni'': implications for plesiosaur phylogenetics
https://www.research[...]
[37]
논문
Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition
[38]
논문
Peculiar macrophagous adaptations in a new Cretaceous pliosaurid
[39]
논문
Plasticity and Convergence in the Evolution of Short-Necked Plesiosaurs
[40]
논문
Morphological and phylogenetic aspects of the dentition of ''Megacephalosaurus eulerti'', a pliosaurid from the Turonian of Kansas, USA, with remarks on the cranial anatomy of the taxon
https://www.research[...]
[41]
논문
Increased pliosaurid dental disparity across the Jurassic-Cretaceous transition
[42]
논문
Bone histology of aquatic reptiles: What does it tell us about secondary adaptation to an aquatic life?
[43]
논문
Quantitative histological models suggest endothermy in plesiosaurs
[44]
논문
Hematological convergence between Mesozoic marine reptiles (Sauropterygia) and extant aquatic amniotes elucidates diving adaptations in plesiosaurs
[45]
논문
Swimming Capabilities of Mesozoic Marine Reptiles: Implications for Method of Predation
[46]
논문
Morphology of the caudal vertebrae in ''Rhomaleosaurus zetlandicus'' and a review of the evidence for a tail fin in Plesiosauria
https://plesiosauria[...]
[47]
논문
Functional anatomy and feeding biomechanics of a giant Upper Jurassic pliosaur (Reptilia: Sauropterygia) from Weymouth Bay, Dorset, UK
[48]
논문
Opalisation of the Great Artesian Basin (central Australia): an Australian story with a Martian twist
https://www.research[...]
[49]
웹사이트
Mineralogical and Paleontological Treasures from the Australian Outback
https://www.research[...]
[50]
논문
Glendonites as a paleoenvironmental tool: implications for early Cretaceous high latitude climates in Australia
https://www.research[...]
[51]
논문
Glaciations at high-latitude Southern Australia during the Early Cretaceous
[52]
서적
Stratigraphy and Palaeontology: Essays in honour of Dorothy Hill
https://archive.org/[...]
Australian National University Press
[53]
간행물
Cretaceous marine reptiles of Australia: a review of taxonomy and distribution.
[54]
서적
小学館の図鑑NEO 大むかしの生物
小学館
[55]
간행물
A mounted skeleton of the giant plesiosaur ''Kronosaurus''.
[56]
서적
恐竜博物図鑑
新樹社
[57]
논문
Giant pliosaurids (Sauropterygia; Plesiosauria) from the Lower Cretaceous peri-Gondwanan seas of Colombia and Australia
https://ui.adsabs.ha[...]
2022-04-01
[58]
논문
Anatomy and relationships of the bizarre Early Cretaceous pliosaurid ''Luskhan itilensis''
https://doi.org/10.1[...]
2023-03-15
[59]
논문
An annotated checklist of Australian Mesozoic tetrapods
https://www.tandfonl[...]
2023-04-03
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com