할키에리아류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
할키에리아류는 캄브리아기에 살았던 멸종된 동물 분류군으로, 몸은 골편으로 덮여 있고 양쪽 끝에 껍데기가 있는 것이 특징이다. 1989년 그린란드의 시리우스 파세트에서 할키에리아 에반겔리스타의 완전한 화석이 발견되어 연구가 시작되었으며, 초기에는 작은껍질화석의 일부로 여겨졌다. 할키에리아류의 진화적 관계는 연체동물, 환형동물, 완족동물 등과의 관련성을 중심으로 논쟁이 있으며, 특히 위왁시아와의 관계가 주목받는다. 멸종 시기는 캄브리아기 보토마절 말 대량 멸종으로 추정되나, 오스트레일리아에서 아우스트랄로할키에리아가 발견되어 멸종 시기에 대한 연구가 진행 중이다.
더 읽어볼만한 페이지
- 캄브리아기의 무척추동물 - 필석
필석은 캄브리아기부터 석탄기 지층에서 화석으로 발견되는 고생대 초기의 멸종된 해양 동물 군체로, 반삭동물로 분류되며, 지질 시대 구분에 중요한 표준 화석으로 활용되어 오르도비스기와 실루리아기 지층 연구에 기여한다. - 캄브리아기의 무척추동물 - 위왁시아
위왁시아는 캄브리아기에 번성한 2.5~5cm 크기의 해양 동물로, 경판과 가시로 덮인 몸과 달팽이 유사한 발을 가졌으며, 최근 연구에서 연체동물과의 연관성이 제기되고 전 세계에서 화석이 발견된다. - 캄브리아기의 동물 - 오파비니아
오파비니아는 캄브리아기에 살았던 멸종된 해양 절지동물로, 5개의 눈, 코, 지느러미를 가진 몸통과 꼬리를 가지며, 절지동물 줄기군으로 분류되어 캄브리아기 대폭발 연구에 영향을 미쳤다. - 캄브리아기의 동물 - 아노말로카리스
캄브리아기에 번성한 라디오돈타류에 속하는 멸종된 속인 아노말로카리스는, 기묘한 새우라는 뜻의 학명처럼 부분적인 화석으로 인해 갑각류로 오해받았으나, 캄브리아기 해양 생태계의 중요한 포식자로서 당시 생물 진화 연구에 중요한 자료를 제공한다. - 버지스 셰일 화석 - 피카이아
피카이아는 캄브리아기 버제스 셰일에서 발견된 몸길이 약 4cm의 멸종된 척삭동물로, 척삭과 근육 덩어리를 가지고 있어 초기 척삭동물의 진화 연구에 중요한 위치를 차지한다. - 버지스 셰일 화석 - 오파비니아
오파비니아는 캄브리아기에 살았던 멸종된 해양 절지동물로, 5개의 눈, 코, 지느러미를 가진 몸통과 꼬리를 가지며, 절지동물 줄기군으로 분류되어 캄브리아기 대폭발 연구에 영향을 미쳤다.
| 할키에리아류 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| |
| 학명 | Halkieria |
| 명명자 | Poulsen, 1967 |
| 모식종 | Halkieria obliqua |
| 모식종 명명자 | Poulsen, 1967 |
| 시대 | 캄브리아기 전기 ~ 중기 (플로이안절) (5억 3,300만 년 전 ~ 5억 1,100만 년 전) |
| 분류 | |
| 계 | 동물계 Animalia |
| 문 | 연체동물문(?) Mollusca 또는 완족동물문(?) Brachiopod |
| 강 | 하르왁시이다 Halwaxiida |
| 과 | 하르키에리아과 Halkieriidae |
| 종 | |
| 종 목록 | H. anabarica H. evangelista H. obliqua |
2. 발견사
경판이라고 불리는 덮개는 오랫동안 작은껍질화석의 구성 요소로 알려져 왔으며, 상세한 분석을 통해 이들 중 일부가 동일한 동물에 속하며 어떻게 함께 맞춰지는지 보여주었다. 모든 단단한 부분이 함께 연결된 ''Halkieria evangelista''의 첫 번째 연결 표본은 1989년 그린란드의 시리우스 파세트 라거슈테트에서 수집되었으며, 1990년 사이먼 컨웨이 모리스와 존 S. 필에 의해 기술되었다.[3] ''H. evangelista''는 할키에리아류를 비롯해 그와 비슷한 껍질 및 골편을 감정하고 재구성하는 과정에 있어 모델로 사용되고 있다.[60][58] 종명인 에반겔리스타는 이 종이 하부 캄브리아기 지층에서 발견되는 화석들을 설명하는 데 큰 도움을 준다는 의미를 담고 있다.[59]
할키에리아류에 속한다고 확신할 수 있는 파편들은 중국의 신장 위구르 자치구[23]와 오스트레일리아의 조지나 분지에서 발견되었으며,[25] 가능한 할키에리아류의 껍질은 캐나다의 버제스 셰일에서 발견되었다.[17]
2. 1. 초기 발견
골편이라고 불리는 보호판은 작은껍질화석을 구성하는 요소들로 알려져 있었고, 세부적인 연구에 의해 이들 중 일부는 하나의 동물에 속한 것으로 원래 어떤 모습이었는지도 밝혀지게 되었다. 단단한 껍질 부분을 포함해 할키에리아 에반겔리스타의 전체 모습을 보여주는 표본이 1989년에 그린란드에 위치한 시리우스 파세트 라거슈태테에서 발견되었으며 1990년에 사이먼 콘웨이 모리스와 존 S. 필에 의해 보고되었다.[57] ''H. 에반겔리스타''(''H. evangelista'')는 할키에리아류를 비롯해 그와 비슷한 껍질 및 골편을 감정하고 재구성하는 과정에 있어 모델로 사용되고 있다.[58][60] 종명인 에반겔리스타는 이 종이 하부 캄브리아기 지층에서 발견되는 화석들을 설명하는 데 큰 도움을 준다는 의미를 담고 있다.[59]할키에리아 종의 화석은 1967년 Poulsen에 의해 덴마크 보른홀름 섬에서 발견된 인편으로 보고되었으며, "할키에리아 오블리쿠아 ''Halkieria obliqua''"로 명명되었다.[49]
2. 2. 전신 화석 발견과 학계 보고
골편이라고 불리는 보호판은 작은껍질화석을 구성하는 요소들로 알려져 있었고, 세부적인 연구에 의해 이들 중 일부는 하나의 동물에 속한 것으로 원래 어떤 모습이었는지도 밝혀지게 되었다. 단단한 껍질 부분을 포함해 할키에리아 에반겔리스타의 전체 모습을 보여주는 표본이 1989년에 그린란드에 위치한 시리우스 파세트 라거슈태테에서 발견되었으며 1990년에 사이먼 콘웨이 모리스와 존 S. 필에 의해 보고되었다.[57] H. 에반겔리스타(''H. evangelista'')는 할키에리아류를 비롯해 그와 비슷한 껍질 및 골편을 감정하고 재구성하는 과정에 있어 모델로 사용되고 있다.[60][58] 종명인 에반겔리스타는 이 종이 하부 캄브리아기 지층에서 발견되는 화석들을 설명하는 데 큰 도움을 준다는 의미를 담고 있다.[59]1967년 Poulsen에 의해 덴마크 보른홀름 섬에서 할키에리아 종의 화석이 인편으로 보고되었으며, "할키에리아 오블리쿠아 ''Halkieria obliqua''"로 명명되었다[49]。이후 인편은 세계 각지에서 발견되었지만, 그 소유자의 전체 모습이 밝혀진 것은 1989년 시리우스 파세트에서 발견된 화석을 바탕으로, 1990년 Simon Conway Morris와 John S. Peel이 동종 ''H. evangelista''를 보고한[50] 것에 기인한다.
2. 3. 추가 발견과 연구
골편이라고 불리는 보호판은 작은껍질화석을 구성하는 요소들로 알려져 있었고, 세부적인 연구에 의해 이들 중 일부는 하나의 동물에 속한 것으로 원래 어떤 모습이었는지도 밝혀지게 되었다. 단단한 껍질 부분을 포함해 할키에리아 에반겔리스타(''Halkieria evangelista'')의 전체 모습을 보여주는 표본이 1989년에 그린란드에 위치한 시리우스 파세트 라거슈태테에서 발견되었으며 1990년에 사이먼 콘웨이 모리스와 존 S. 필에 의해 보고되었다.[57] H. 에반겔리스타(''H. evangelista'')는 할키에리아류를 비롯해 그와 비슷한 껍질 및 골편을 감정하고 재구성하는 과정에 있어 모델로 사용되고 있다.[60][58] 종명인 에반겔리스타는 이 종이 하부 캄브리아기 지층에서 발견되는 화석들을 설명하는 데 큰 도움을 준다는 의미를 담고 있다.[59]할키에리아류 경판의 가장 오래된 것으로 알려진 발생은 시베리아의 네마키트-달디니안절 상부의 ''Purella antiqua Zone''에서 발견된다.[24] 캄브리아기 시대의 보토미안절 말에 일어난 대멸종으로 할키에리아류를 포함한 대부분의 소형 쉘이 사라졌다고 생각되었지만, 2004년에는 ''Australohalkieria''로 분류된 할키에리아류 화석이 오스트레일리아의 조지나 분지의 중기 캄브리아기 암석에서 보고되었다. 이 분지군이 다른 할키에리아류 분지군이 멸종한 동안 살아남은 이유는 알려져 있지 않다.[25] 보토미안절 말 멸종에서 살아남은 것으로 알려진 유일한 고배류 또한 남아메리카, 아프리카, 인도, 오스트레일리아 및 남극 대륙을 포함하는 고대 초대륙인 곤드와나에서 발생한다는 점도 중요하다.[26][27][25]
할키에리아류와 다른 소형 쉘 화석은 일반적으로, 항상 그런 것은 아니지만, 인산염으로 보존되며, 이는 원래의 광물 조성일 수도 있고 아닐 수도 있다. 인산염 피복에 의한 보존은 초기 캄브리아기 동안에만 흔했던 것으로 보이며, 굴을 파는 동물의 해저 교란의 증가로 인해 시간이 지남에 따라 희귀해졌다. 따라서 할키에리아류와 다른 소형 쉘 화석은 가장 오래된 화석보다 더 일찍 살았고 가장 최근의 화석보다 더 늦게 살았을 수 있다.[28][29][30] 고생물학자들은 이러한 종류의 불확실성을 시뇨르-립스 효과라고 부른다.[31]
할키에리아 종의 화석은 1967년 Poulsen에 의해 덴마크 보른홀름 섬에서 발견된 인편으로 보고되었으며, "할키에리아 오블리쿠아 ''Halkieria obliqua''"로 명명되었다.[49] 인편은 세계 각지에서 발견되었지만, 그 소유자의 전체 모습이 밝혀진 것은 1989년 시리우스 파세트에서 발견된 화석을 바탕으로, 1990년 Simon Conway Morris와 John S. Peel이 동종 ''H. evangelista''를 보고한[50] 것에 기인한다.
3. 형태 및 구조
할키에리아류는 몸 전체가 골편이라고 불리는 작은 갑옷 조각들로 덮여 있고, 몸 앞뒤 양 끝에 껍데기를 가진 것이 특징이다.
아우스트랄로할키에리아는 골편만이 발견되었는데, 할키에리아와 매우 유사하여 같은 종류의 "체강골편"으로 분류된다.[60] 체강골편은 원래 유기질 조직으로 채워진 공간을 광물화된 껍질이 둘러싸고 있는 형태이며, 바깥쪽에 물질을 추가하며 성장하지는 않는다.[61]
할키에리아류는 공통적으로 세 종류의 골편을 가진다.[60]
- 손바닥형 골편: 납작하고 단풍나무 잎 모양을 하고 있으며, 세 가지 골편 중 가장 작다.
- 칼날형 골편: 납작하고 칼날 모양을 하고 있다.
- 낫형 골편: 칼날형과 비슷한 크기지만 가시 형태이며, 원통이 눌린 것 같은 모양이다.
할키에리아와 아우스트랄로할키에리아의 손바닥형 및 칼날형 골편에는 뚜렷한 이랑이 있으며, 몸에 붙어있기 위해 바닥 쪽이 90도로 꺾인 것을 제외하면 거의 평평하다. 낫형 골편에는 이랑이 잘 보이지 않으며, 몸에서 45~90도 각도로 뻗어 있다.[60]
할키에리아의 골편 속은 비어 있거나, (일부 표본에서는) 측면에 가는 구멍이 나 있다.[63] 할키에리아가 성장하면 껍데기는 바깥쪽 가장자리에 새로운 물질이 추가되며 커지지만, 골편은 같은 크기를 유지한다. 비교적 완전한 표본에서 칼날형 골편은 특정 패턴을 보이는데, 성장하면서 오래되고 작은 골편은 떨어져 나가고 더 큰 골편으로 대체되는 것으로 보인다. 골편은 아래쪽에서 분비되어 만들어지며, 골편과 피부 사이에는 얇은 이랑 구조가 관찰된다.[63][64]
오스트레일리아에서 발견된 아우스트랄로할키에리아 수페르스테스는 보토마절 말 멸종에서 살아남은 종으로, "남쪽에서 살아남은 할키에리아"라는 뜻이다. 이 종의 골편은 위쪽이 볼록하고 아래쪽은 오목하며, 서로 겹쳐져 있다. 내부 공간은 할키에리아보다 복잡하며, 중앙 관이 납작해져 있고 양쪽 관은 연결되지 않은 것으로 보인다. 골편 측면 역시 다른 미세구조를 가진다.[60] A. 수페르스테스 골편 중앙 관 위쪽은 납작하고 표면 바로 밑에 있어 움푹 들어간 자국이 생기는데, 이는 후각 역할을 했을 수 있다. 골편이 인산염으로 덮여 있는 것은 유기질 피부로 덮여 있었기 때문일 수 있다.[60]
A. 수페르스테스의 골편은 좌우 대칭을 이루는 두 종류로 구성되며, 크기는 매우 작다. 초기 캄브리아기 할키에리아류는 낫형보다 손바닥형 및 칼날형 골편이 훨씬 많지만, A. 수페르스테스는 낫형 골편이 더 많고 손바닥형은 드물다. 이는 사망 후 골편 소실, 골편체 구성 차이, 혹은 종 크기 차이 때문일 수 있다.[60]
시포고누키테스류는 할키에리아류보다 단순한 골편체를 가지며, 골편 내부 구조 역시 단순하다.[65] 시포고누키테스, 다바샤니테스, 로포키테스, 마이카넬라는 모두 시포고누키테스와 유사하며,[65] 드레파노키테스 골편은 길이와 너비 비율로 구분된다.[65] 마이카넬라 껍질은 시포고누키테스 골편이 석회화된 기질과 합쳐진 형태이며, 어린 개체는 골편과 합쳐지지 않은 것으로 보인다.[66] 시포고누키테스 골편 내부는 단순하며 측면 공간은 없다.[63]
니넬라로 대표되는 니넬라류는 하부 캄브리아기 지층에서 발견되며, 한 종류의 골편만 있는 단순한 골편체를 가진다. 골편은 갈고리 모양으로, 할키에리아류나 시포고누키테스류와 유사하며, 석회암 성분으로 속이 비어 있고 표면에 이랑 구조가 있다.[65]
히포파란기테스[67] 골편은 중앙에 넓은 빈 공간과 껍질 전체에 작은 기공을 가지는데, 이는 다른 할키에리아류 측면 공간이나 다판류 지상감각기에 해당할 수 있다.[63] 히포파란기테스는 챈슬로리드 골편과 유사하여, 할키에리아류와 챈슬로리드를 실로스클레리토포라로 묶는 증거로 제시되기도 한다.[65]
사키테스는 가시 모양 골편으로 이루어진 속으로, 이전에는 할키에리아류와 가깝다고 생각되었으나,[69] 최근에는 챈슬로리드와 근연관계라는 주장이 제기되었다.[70] 시노사키테스는 골편만 발견된 할키에리아류로, 중앙 관과 수직 방 모양 공간이 특징이다.[63][69] 이 골편은 오스트레일리아에서 발견된 탐베톨레피스와 동일하며, 좌우대칭 몸을 가졌을 것으로 추정된다. 할키에리아처럼 손바닥형, 칼날형, 낫형으로 구분된다.[63]
오이코제테테스[71]는 버제스 셰일에서 발견된 두 종류 모자 형태 껍질로만 알려져 있으며, 할키에리아류 앞, 뒤 껍질로 추정된다.[58] 껍질은 석회질 성분이었을 것으로 보인다.[72]
하부 캄브리아기 지층에서 발견되는 오크루라누스(혹은 에오할로비아)는 오이코제테테스 껍질과 동일하며, 할키에리아류 몸에 붙어 있었을 것으로 추정되지만,[73] 중간 판을 보면 팔레오로리카타와 유사했을 수도 있다.[74]
할키에리아는 앞뒤가 원형인 직사각형 모양이며, 몸 앞뒤에 "껍데기"를 하나씩 갖는 것이 특징이다. 콘웨이 모리스 등은 이 "껍데기"가 합쳐져 완족동물이 탄생했다는 가설을 제창했지만,[51] 할키에리아보다 2000만 년 이상 후 지층에서 껍데기가 하나뿐인 오르스로잔클루스가 발견되어 가능성은 낮아졌다. 인편 형태 외에 치설을 가진 화석도 발견되어,[52] 위왁시아와 근연 관계이며, ''Halwaxiida'' 강(한국어 명칭 미정)을 이룬다고 여겨진다. 치설 때문에 오돈토그리푸스와 관계도 지적된다.[53][54]
3. 1. 몸체
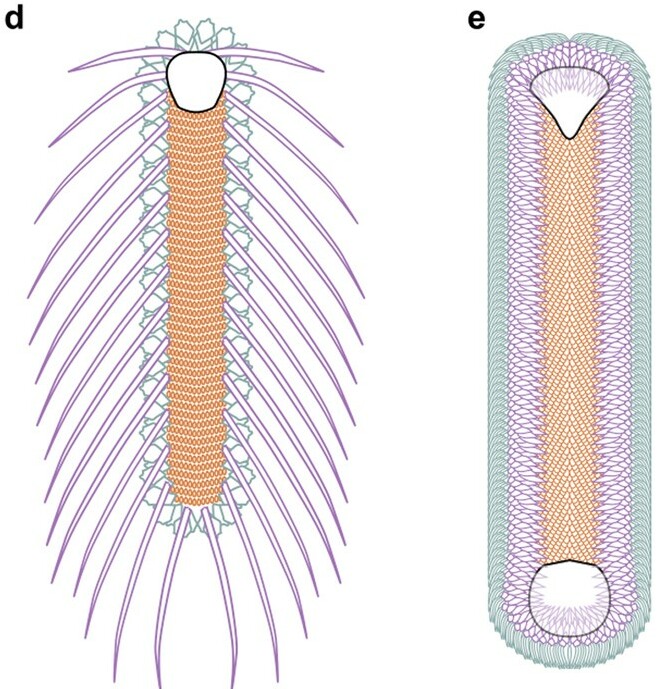
할키에리아는 사슬갑옷을 입은 민달팽이처럼 생겼다. 몸길이는 1.5cm에서 8cm 정도이고 좌우대칭이며 위에서 아래로 눌린 형태이다. 아래쪽에는 골편이 없고, 앞뒤 양쪽에는 판 모양의 껍질이 있는데 이 껍질에는 나무의 나이테와 유사한 성장선이 뚜렷하게 보인다. 위쪽면은 타일처럼 서로 겹쳐진 약 2,000개 정도의 골편으로 덮여 있으며, 골편 형태에 따라 세 부위로 나뉜다.[62]- "손바닥형" 골편: 단풍나무 잎처럼 생겼으며 위쪽 면의 앞에서 뒤까지, 두 개의 판 모양 껍질 사이를 채우고 있다.
- "칼날형" 골편: 칼날처럼 생겼으며 손바닥형 골편 부위의 양 측면에 위치, 위쪽 면의 중앙부를 향해 놓여 있다.
- "낫형" 골편: 낫 같이 생겼으며 바깥쪽 가장자리에 위치한다.
골편의 가운데 공간은 비어 있으며 (최소한 일부 표본에서는) 측면에 가느다란 구멍이 나있다.[63]
할키에리아가 성장하면 판 모양 껍질도 바깥쪽 가장자리에 새로운 물질이 추가되면서 성장한다.[59] 각각의 골편은 같은 크기를 유지한다. 비교적 완전한 형태를 갖춘 표본에서 칼날형 골편은 특정한 패턴을 보이는데, 할키에리아가 성장하면서 오래되고 작은 골편은 떨어져 나가고 더 큰 골편으로 대체되는 것으로 보인다. 골편은 아래쪽에서부터 분비되어 만들어진다.[63] 골편과 피부 사이에는 얇은 이랑 구조의 흔적이 있다.[64]
판 모양 껍질과 골편은 원래 탄산칼슘 성분이었던 것으로 보인다.[59] 보존 상태를 고려할 때 껍질이 유기질이었을 가능성도 제기되었으나, 석회화되지 않은 유기체 화석은 보통 얇은 막 형태로 발견되는 데 비해 할키에리아 화석은 삼엽충이나 히올리트처럼 3차원 형태를 가지고 있어 가능성은 낮아 보인다. 몇몇 표본에서는 평평한 판이 휘어져 있는데, 이는 생물이 매몰될 당시 골편과 연결된 근육이 보존되었음을 의미한다.[64]
할키에리아의 발은 말랑말랑하고 근육으로 되어 있었을 것이다. 헤엄치기에 적합하지 않고 바닥을 파고 들어가는 기관도 없기 때문에 해저 표면에 살며 발의 근육을 물결처럼 움직여 "걸어다녔"던 것으로 보인다. 뒤로 뻗은 낫형 골편은 접지력을 높여 미끄러지는 것을 방지했을 수 있다. 일부 표본은 공벌레처럼 몸 일부가 말려있는데, 이 자세에서는 칼날형 골편이 바깥쪽으로 뻗어 포식자를 물리치는 역할을 했을 가능성이 있다. 몸 양쪽 끝 껍질의 기능은 골편만으로도 충분히 몸을 보호할 수 있었을 것으로 보여 알기 어렵다. 앞쪽 껍질 안쪽에는 내부기관 부착 자리로 생각되는 자국이 있다. 한 표본에서는 화석화 전에 뒤쪽 껍질이 45° 회전되었는데, 뒤쪽 껍질 아래 빈 공간에 아가미가 있었을 가능성이 있다.[59]
일부 화석 뒤쪽에서는 내장 흔적이 발견되었다.[64] 어떤 표본에서는 일부를 치설[59]( 연체동물의 특징인 이빨 달린 키틴질 혀)로 해석했으나, 이 표본에서 "골편체"(몸을 둘러싼 골편 전체)가 접혀 있어 치설로 해석된 것이 원래 위치에서 이동한 낫형 골편이었을 가능성이 있다.[64]
3. 2. 골편 (Sclerite)
아우스트랄로할키에리아에서는 갑옷 같은 골편들만이 발견되었으며, 그 분석은 이들이 할키에리아와 유사했을 것이라는 가정에 기반한다. 하지만 골편이 매우 유사하여 이러한 가정은 상당히 안전해 보인다.[60] 두 속 모두 골편은 "체강골편"이라고 불리는 유형인데, 원래 유기 조직으로 채워졌던 공간 주위에 광물화된 껍질이 있으며 바깥쪽에 물질을 추가하여 성장했다는 증거는 없다.[61] 두 속 모두 세 가지 다른 모양의 골편을 가지고 있다. 일반적으로 가장 작은 "손바닥형"은 평평하고 단풍나무 잎과 유사하며, "칼날형"은 평평하지만 칼날 모양이다. 그리고 "낫형"은 칼날 모양과 크기가 거의 같지만 가시 모양이며 약간 찌그러진 원통과 같다. 할키에리아와 아우스트랄로할키에리아 모두에서 손바닥형과 칼날형 골편은 두드러진 이랑을 가지고 있으며, 몸에 꼭 맞게 고정되었음을 나타내는 기저부의 90° 굴곡을 제외하고는 비교적 평평하다. 낫형은 대부분 이랑이 없고 약 45°에서 90° 사이의 각도로 몸에서 돌출된 것으로 보인다.[60]할키에리아는 사슬갑옷을 입은 민달팽이처럼 생겼다. 몸길이는 1.5cm에서 8cm 정도이고 좌우대칭이며 위에서 아래로 눌린 형태로 아래쪽에는 골편이 없다. 앞뒤 양쪽에는 판 모양의 껍질이 있는데 이 껍질에는 나무의 나이테와 유사한 성장선이 뚜렷하게 보인다. 위쪽면의 그 외 부분은 서로 타일처럼 겹쳐진 약 2,000개 정도의 골편이 덮고 있으며 서로 다른 형태의 골편들이 차지하고 있는 세 부위로 나뉜다.[62] "손바닥형" 골편은 단풍나무 잎처럼 생겨서 위쪽 면의 앞에서 뒤까지, 두 개의 판 모양 껍질 사이를 채우고 있다. 칼날처럼 생긴 "칼날형" 골편들은 손바닥형 골편이 있는 부위의 양 측면에 위치하며 위쪽 면의 중앙부를 향해 놓여 있다. 낫 같이 생긴 "낫형" 골편들은 바깥쪽 가장자리에 위치하고 있다. 골편의 가운데 공간은 비어 있으며 (최소한 일부 표본에서는) 측면에 가느다란 구멍이 나 있다.[63] 할키에리아가 성장하게 되면 판 모양의 껍질도 바깥쪽 가장자리에 새로운 물질이 추가되면서 성장하게 된다.[59] 각각의 골편은 같은 크기를 유지한다. 비교적 완전한 형태를 갖추고 있는 표본 모두에서 칼날형 골편은 특정한 패턴을 보여주는데, 할키에리아가 성장하면서 오래되고 작은 골편들은 떨어져 나가고 더 큰 골편으로 대체되는 것으로 보인다. 골편들은 아래쪽에서부터 분비되어 만들어진다.[63] 골편과 피부 사이에는 얇은 이랑 구조의 흔적을 찾아볼 수 있다.[64]
판 모양의 껍질과 골편은 원래 탄산칼슘 성분이었던 것으로 보인다.[59] 보존 상태를 고려해 볼 때 원래는 이 껍질들이 모두 유기질이었을 가능성도 제기되었으나 석회화되지 않은 유기체의 화석이 보통 얇은 막 형태로 발견되는 데 비해 할키에리아 화석은 삼엽충이나 히올리트처럼 3차원적인 형태를 가지고 있는 것으로 보아 그럴 가능성은 낮아 보인다. 실제로 몇몇 표본에서는 평평한 판이 휘어져 있는 모습도 발견되었는데, 이것은 생물이 매몰될 당시 골편과 연결되어 있던 근육이 아직 그대로 보존되어 있었음을 의미한다.[64]
할키에리아의 발은 말랑말랑했고, 아마도 근육으로 되어 있었을 것이다. 할키에리아는 헤엄치기에 적합하지 않았고 바닥을 파고 들어가기 위해 적응된 기관도 가지고 있지 않기 때문에 해저 표면에 살며 발의 근육을 물결처럼 움직여 "걸어다녔"던 것으로 보인다. 뒤로 뻗어 있는 낫형 골편은 접지력을 높여 뒤로 미끄러지는 것을 방지했을 수 있다. 일부 표본은 공벌레처럼 몸 일부가 말려있는 모습으로 발견되는데 이 자세에서는 칼날형 골편이 바깥쪽으로 뻗게 되어 포식자를 물리치는 역할을 했을 가능성이 있다. 몸의 양쪽 끝에 있는 모자처럼 생긴 껍질의 기능이 무엇이었는지 알기는 쉽지 않은데, 골편들만으로도 충분히 몸을 보호할 수 있었을 것으로 보이기 때문이다. 앞쪽 껍질의 안쪽은 내부기관이 부착되는 자리였을 것으로 생각되는 자국이 있다. 한 표본에서는 화석화가 일어나기 전에 뒤쪽 껍질이 45° 회전되어 있었던 것으로 보이는데, 뒤쪽 껍질 아래의 빈 공간에 아가미가 있었을 가능성이 있다.[59]
일부 화석의 뒤쪽에서는 내장의 흔적이 발견되었다.[64] 어떤 표본에서는 그 일부를 치설[59] 즉 연체동물의 중요한 특징인 이빨이 달린 키틴질 혀로 해석한 경우가 있었으나, 이 표본에서 "골편체", 즉 몸을 둘러싸고 있는 골편 전체가 접혀 있었기 때문에 치설이라고 해석된 것이 원래 있던 곳에서 다른 위치로 이동한 낫형 골편들이었을 가능성이 있다.[64]
오스트레일리아에서 발견된 것 중 가장 온전하고 풍부한 아우스트랄로할키에리아 수페르스테스의 학명은 "남쪽에서 살아남은 할키에리아"라는 의미를 가지고 있는데, 이것은 해당 종이 보토마절 말의 멸종사건에서 살아남은 할키에리아류이기 때문이다. 이 종으로 분류된 골편은 위쪽 표면으로는 볼록하고 아래쪽은 오목하다. 또 골편이 있는 평면 상에서 휘어 있으며 서로 겹쳐져 있는데, 각각의 골편에서 오목한 부분은 다음 골편의 볼록한 부분에 덮여 있다. 아우스트랄로할키에리아 내부의 빈 공간은 할키에리아의 단순한 관 모양보다 더 복잡한 형태를 가지는데 골편 높이의 절반 정도를 올라가면 원통 모양의 관이 한 쌍의 가느다란 관으로 나뉘고, 가운데 부분은 납작해 져서 양쪽의 관은 연결되어 있지 않은 것으로 보인다. 골편의 측면 역시 다른 미세구조를 가지고 있다.[60]
A. 수페르스테스의 골편 중앙 관의 위쪽은 납작한 형태로 표면 바로 밑에 위치하는데 이때문에 위쪽 표면의 끝쪽에는 움푹 들어간 자국이 생긴다. 이 부분은 광물화되지 않은 상태로 남아 있어서 이것이 물 속의 화학물질을 받아들여 후각의 역할을 했을 수도 있다. A. 수페르스테스 골편이 인산염 성분으로 덮여 있는 것은 원래 유기질 피부로 덮여 있었기 때문일 가능성이 있다. 선인장처럼 생긴 챈슬로리드(chancelloriid)의 골편 역시 유기질로 덮여 있는 경우가 있다. 만일 할키에리아류가 초기 연체동물이었다면 골편의 표면은 현생 연체동물 일부에서 볼 수 있는 각피층과 유사했을지도 모른다.[60]
A. 수페르스테스의 골편은 서로 거울상을 하고 있는 두 종류로 이루어지는데, 이 두 종류 골편이 발견되는 양이 비슷한 것으로 보아 A. 수페르스테스는 좌우대칭의 몸을 가지고 있었던 것으로 보인다. 골편은 모두 매우 작은 크기로, 손바닥형은 250μm에서 650μm, 칼날형은 300μm에서 1000μm 정도의 길이를 가진다. 낫형은 다시 두 그룹으로 나뉘는데 밑부분에 완만한 S-곡선을 가지고 있는 것은 400μm에서 1000μm의 길이로 바닥쪽이 살짝 비틀려 있는 경우가 많으며, 밑부분이 45°에서 90° 휘어져 있는 것은 400μm에서 500μm 정도 길이이다.[60]
초기 캄브리아기 할키에리아류의 골편체에는 낫형 골편보다 손바닥형 및 칼날형 골편이 훨씬 더 많이 포함되어 있다. 반면, A. 수페르스테스의 낫형 골편은 칼날형이나 손바닥형보다 훨씬 더 많이 발견되며 손바닥형 골편은 드물다. 어쩌면 사망 이후 어떤 과정에 의해 다수의 손바닥형과 일부 칼날형 골편이 제거되었을 수도 있지만, A. 수페르스테스의 골편체 중 해저 가까이에 있는 바닥 부분이 상대적으로 측면이나 위쪽 부분보다 더 큰 면적을 차지하고 있었을 가능성이 높다. 또 A. 수페르스테스 골편은 초기 캄브리아기 할키에리아류의 골편에 비해 1/3 정도 크기밖에 되지 않는다. 대부분의 초기 캄브리아기 할키에리아류와 더 큰 생물들을 포함하는 조지나 어셈블리지의 화석들이 모두 동일한 방식, 즉 인산염화가 일어나는 방식으로 보존되었기 때문에 보존 방식에서의 차이가 이런 차이를 만들어냈을 가능성은 적다. A. 수페르스테스의 크기가 작은 이유에 대한 설명으로는 조지나 어셈블리지의 할키에리아류가 어린 것이었을 가능성, 이들의 골편체가 초기 캄브리아기 할키에리아류보다 더 많은 수의 골편들로 구성되어 있었을 가능성, 혹은 A. 수페르스테스의 크기가 원래 작았을 가능성 등이 꼽힌다.[60]
조지나 분지에서는 할키에리아류의 껍질로 분류될 수 있는 화석이 발견되지 않았다. 할키에리아의 껍질도 매우 드물게 발견되기 때문에 이것이 아우스트랄로할키에리아가 껍질을 가지고 있지 않았다는 의미는 아니다.[60]
시포고누키테스류는 두 가지 형태의 골편, 그리고 껍질을 가지고 있어서 할키에리아나 그와 가까운 할키에리아류보다는 단순한 골편체를 가지고 있으며 이들 골편의 내부 역시 할키에리아보다 단순한 구조를 가지고 있다.[65]
여기에 해당하는 속들인 시포고누키테스(''Siphogonuchites''), 다바샤니테스(''Dabashanites''), 로포키테스(''Lopochites''), 그리고 마이카넬라(''Maikhanella'')는 모두 시포고누키테스와 유사한 형태를 하고 있다.[65] 드레파노키테스(''Drepanochites'')의 골편은 길이와 너비 등의 비율로 다른 종류들과 구분된다.[65]
마이카넬라의 껍질은 시포고누키테스의 골편이 석회화된 기질(matrix)과 합쳐진 형태를 하고 있다. 어린 개체의 껍질은 골편과 합쳐지지 않은 것으로 보인다.[66]
시포고누키테스 골편 내부의 빈 공간은 단순한 형태로 측면에 연결된 공간은 발견되지 않는다.[63]
니넬라(''Ninella'')로 대표되는 니넬라류는 하부 캄브리아기 지층에서 발견되며 (골편들의 형태에 변이는 있고, 거울상을 보이는 두 종류가 발견되지만) 형태적으로는 한 종류의 골편만이 존재하는 단순한 골편체를 가지고 있다. 니넬라류의 골편은 갈고리 형태로, 할키에리아류나 시포고누키테스류의 골편과 유사하게 생겼다. 석회암 성분으로 속이 비었으며 표면에는 솟아 올라온 이랑 구조가 있다.[65]
히포파란기테스[67]의 골편은 중앙에 넓은 빈 공간을 가지고 있으며 껍질 전체에 걸쳐 작은 기공이 있는데 이것은 다른 할키에리아류의 측면 공간 (그리고 어쩌면 다판류의 지상감각기 asthete)에 해당하는 것이다.[63]
히포파란기테스 속은 형태 상으로 챈슬로리드의 골편과 유사한데, 할키에리아류와 챈슬로리드를 묶어 실로스클레리토포라(Coeloscleritophora)로 분류하는 관점을 지지하는 증거로 제시되기도 한다.[65]
사키테스는 가시 모양의 골편으로 이루어진 속이다. 사키테스의 표본들 중 많은 수가 지금은 다른 할키에리아류로 분류되었다.[68]
사키테스는 할키에리아류와 가까운 관계라고 생각되어 왔으나,[69] 최근에는 챈슬로리드와 근연관계라는 주장이 제기되었다.[70]
시노사키테스는 골편만 발견된 할키에리아류이다. 이 골편 내부에는 중앙에 위치한 관과 거의 수직으로 나있는 방 모양의 공간이 있으며 이 공간은 좁은 관을 통해 중앙으로 이어진다.[63][69] 방 모양의 공간은 아우스탈로할키에리아에서 발견되는 긴 관과 비슷하게 40μm 정도의 지름을 가지며 이런 공간들이 골편의 긴 방향이 아니라 그와 수직으로 나있다는 것은 길이 1mm에서 2mm 정도 되는 시노사키테스의 골편이 다른 할키에리아류의 골편보다 크다는 의미이다.[63]
이 골편은 오스트레일리아에서 처음 발견된 탐베톨레피스(''Thambetolepis'')와 동일한 것으로 밝혀졌다. 골편이 서로 거울상으로 나타나는 두 종류로 구성되는 것으로 보아 좌우대칭의 몸을 가지고 있었던 것으로 생각된다. 할키에리아와 마찬가지로 시노사키테스의 골편은 손바닥형, 칼날형, 그리고 낫형으로 구분할 수 있다.[63]
"경화판"이라고 불리는 덮개는 오랫동안 소형 쉘 화석의 구성 요소로 알려져 왔으며, 상세한 분석을 통해 이들 중 일부가 동일한 동물에 속하며 어떻게 함께 맞춰지는지 보여주었다. 모든 단단한 부분이 함께 연결된 ''Halkieria evangelista''의 첫 번째 연결 표본은 1989년 그린란드의 시리우스 파셋 라거슈테트에서 수집되었으며, 1990년 사이먼 컨웨이 모리스와 존 S. 필에 의해 기술되었다.[3] ''H. evangelista''는 다른 유사한 껍질과 경화판을 할키에리아류로 식별하고 재구성하는 모델로 사용된다;[25][17] 그 종명 ''evangelista''는 초기 캄브리아기 화석 기록을 설명하는 능력을 반영한다.[6]
3. 3. 껍데기 (Shell)
할키에리아는 사슬갑옷을 입은 민달팽이처럼 생겼으며, 몸길이는 1.5cm에서 8cm 정도이다. 좌우대칭이며 위에서 아래로 눌린 형태이고, 아래쪽에는 골편이 없다. 앞뒤 양쪽에는 판 모양의 껍데기가 있는데, 이 껍질에는 나무의 나이테와 유사한 성장선이 뚜렷하게 나타난다. 위쪽면의 나머지 부분은 서로 타일처럼 겹쳐진 약 2,000개 정도의 골편으로 덮여 있으며, 서로 다른 형태의 골편들이 차지하는 세 부위로 나뉜다.[62] 판 모양의 껍질과 골편은 원래 탄산칼슘 성분이었던 것으로 보인다.[59]몸 양쪽 끝에 있는 모자 모양 껍데기의 기능은 정확히 알기 어려운데, 골편만으로도 충분히 몸을 보호할 수 있었을 것으로 보이기 때문이다. 앞쪽 껍질 안쪽에는 내부 기관이 부착되는 자리로 추정되는 자국이 있다. 한 표본에서는 화석화가 일어나기 전에 뒤쪽 껍질이 45° 회전되어 있었는데, 뒤쪽 껍질 아래의 빈 공간에 아가미가 있었을 가능성이 제기된다.[59]
오이코제테테스[71]는 버제스 셰일에서 발견된 두 종류의 모자 형태 껍질로만 알려져 있으며, 연대는 5억 5백만 년 전으로 확인되었다. 이 두 종류의 껍질은 할키에리아류의 앞쪽 껍질과 뒤쪽 껍질로 추정된다.[58] 오이코제테테스가 살아있을 때 그 껍질은 석회질 성분이었던 것으로 보인다(교결 과정에서 원래의 광물은 다른 광물, 예를 들면 규산으로 치환되기도 한다).[72]
하부 캄브리아기 지층에서 발견되는 오크루라누스(''Ocruranus'' 혹은 에오할로비아''Eohalobia'')는 오이코제테테스의 껍질과 동일한 것으로 생각되며, 할키에리아류 형태의 몸에 붙어 있었던 것으로 보이지만,[73] 중간 판(intermediate valve)을 보면 팔레오로리카타(Palaeoloricata)와 유사한 형태의 몸을 가지고 있었을 수도 있다.[74]
할키에리아는 앞뒤가 원형인 직사각형(약제 캡슐과 같은 형태)이며, 전신이 인편으로 덮여 있다. 가장 큰 특징은 몸의 앞뒤에 하나씩 "껍데기"를 갖는다는 것이다.
3. 4. 기타
할키에리아는 사슬갑옷을 입은 민달팽이처럼 생겼다. 몸길이는 1.5cm에서 8cm 정도이고 좌우대칭이며 위에서 아래로 눌린 형태이다. 아래쪽에는 골편이 없고, 앞뒤 양쪽에는 판 모양의 껍질이 있는데 이 껍질에는 나무의 나이테와 유사한 성장선이 뚜렷하게 보인다. 위쪽면의 그 외 부분은 서로 타일처럼 겹쳐진 약 2,000개 정도의 골편으로 덮여 있으며, 서로 다른 형태의 골편들이 차지하고 있는 세 부위로 나뉜다.[62]
- "손바닥형" 골편: 단풍나무 잎처럼 생겼으며 위쪽 면의 앞에서 뒤까지, 두 개의 판 모양 껍질 사이를 채우고 있다.
- "칼날형" 골편: 칼날처럼 생겼으며 손바닥형 골편이 있는 부위의 양 측면에 위치하며 위쪽 면의 중앙부를 향해 놓여 있다.
- "낫형" 골편: 낫 같이 생겼으며 바깥쪽 가장자리에 위치하고 있다.
골편의 가운데 공간은 비어 있으며 (최소한 일부 표본에서는) 측면에 가느다란 구멍이 나있다.[63] 할키에리아가 성장하면 판 모양의 껍질도 바깥쪽 가장자리에 새로운 물질이 추가되면서 성장한다.[59] 각각의 골편은 같은 크기를 유지한다. 비교적 완전한 형태를 갖추고 있는 표본 모두에서 칼날형 골편은 특정한 패턴을 보여주는데, 할키에리아가 성장하면서 오래되고 작은 골편들은 떨어져 나가고 더 큰 골편으로 대체되는 것으로 보인다. 골편들은 아래쪽에서부터 분비되어 만들어진다.[63] 골편과 피부 사이에는 얇은 이랑 구조의 흔적을 찾아볼 수 있다.[64]
판 모양의 껍질과 골편은 원래 탄산칼슘 성분이었던 것으로 보인다.[59] 보존 상태를 고려해 볼 때 원래는 이 껍질들이 모두 유기질이었을 가능성도 제기되었으나, 석회화되지 않은 유기체의 화석이 보통 얇은 막 형태로 발견되는 데 비해 할키에리아 화석은 삼엽충이나 히올리트처럼 3차원적인 형태를 가지고 있는 것으로 보아 그럴 가능성은 낮아 보인다. 실제로 몇몇 표본에서는 평평한 판이 휘어져 있는 모습도 발견되었는데, 이것은 생물이 매몰될 당시 골편과 연결되어 있던 근육이 아직 그대로 보존되어 있었음을 의미한다.[64]
할키에리아의 발은 말랑말랑했고, 아마도 근육으로 되어 있었을 것이다. 헤엄치기에 적합하지 않았고 바닥을 파고 들어가기 위해 적응된 기관도 가지고 있지 않기 때문에 해저 표면에 살며 발의 근육을 물결처럼 움직여 "걸어다녔"던 것으로 보인다. 뒤로 뻗어 있는 낫형 골편은 접지력을 높여 뒤로 미끄러지는 것을 방지했을 수 있다. 일부 표본은 공벌레처럼 몸 일부가 말려있는 모습으로 발견되는데, 이 자세에서는 칼날형 골편이 바깥쪽으로 뻗게 되어 포식자를 물리치는 역할을 했을 가능성이 있다. 몸의 양쪽 끝에 있는 모자처럼 생긴 껍질의 기능이 무엇이었는지 알기는 쉽지 않은데, 골편들만으로도 충분히 몸을 보호할 수 있었을 것으로 보이기 때문이다. 앞쪽 껍질의 안쪽은 내부기관이 부착되는 자리였을 것으로 생각되는 자국이 있다. 한 표본에서는 화석화가 일어나기 전에 뒤쪽 껍질이 45° 회전되어 있었던 것으로 보이는데, 뒤쪽 껍질 아래의 빈 공간에 아가미가 있었을 가능성이 있다.[59]
일부 화석의 뒤쪽에서는 내장의 흔적이 발견되었다.[64] 어떤 표본에서는 그 일부를 치설[59], 즉 연체동물의 중요한 특징인 이빨이 달린 키틴질 혀로 해석한 경우가 있었으나, 이 표본에서 "골편체"(몸을 둘러싸고 있는 골편 전체)가 접혀 있었기 때문에 치설이라고 해석된 것이 원래 있던 곳에서 다른 위치로 이동한 낫형 골편들이었을 가능성이 있다.[64]
히포파란기테스[67]의 골편은 중앙에 넓은 빈 공간을 가지고 있으며 껍질 전체에 걸쳐 작은 기공이 있는데, 이것은 다른 할키에리아류의 측면 공간(그리고 어쩌면 다판류의 지상감각기 asthete)에 해당하는 것이다.[63] 히포파란기테스 속은 형태 상으로 챈슬로리드의 골편과 유사한데, 할키에리아류와 챈슬로리드를 묶어 실로스클레리토포라(Coeloscleritophora)로 분류하는 관점을 지지하는 증거로 제시되기도 한다.[65] 로마술카키테스는 골편으로만 알려진 속이다.[65]
할키에리아는 앞뒤가 원형인 직사각형(약제 캡슐과 같은 형태)이며, 전신이 인편으로 덮여 있다. 특징적인 점은 몸의 앞뒤에 하나씩 "껍데기"를 갖는다는 것이다. Conway Morris 등은 이 "껍데기"가 합쳐져 완족동물이 탄생했다는 가설을 제창하고 있다.[51] 그러나 할키에리아보다 2000만 년 이상 지난 지층에서, "껍데기"를 1개만 가진 오르스로잔클루스(Orthrozanclus)가 발견되면서, 앞서 언급한 가설의 가능성은 낮아졌다.
또한, 인편을 가진 형태에 더해, 치설을 가진 화석도 발견[52]되어, 위왁시아와 근연 관계에 있으며, 단계통의 ''Halwaxiida'' 강(한국어 명칭 미정)을 이룬다고 여겨진다. 치설을 가진 점으로 미루어 오돈토그리푸스와의 관계도 지적되고 있다.[53][54]
4. 생태
할키에리아류 화석은 대부분 골편(갑옷판) 형태로 발견되며, 보존 상태는 화석이 발견된 지역의 환경에 따라 다르다. 할키에리아류를 포함한 작은 껍질 화석들은 대개 인산염으로 보존되어 있는데, 이들의 원래 광물 성분은 인산염일 수도 있고 아닐 수도 있다. 초기 캄브리아기에는 인산염 피복 보존 방식이 흔했지만, 시간이 지나면서 해저 바닥을 파고 들어가는 동물들의 교란 작용이 증가함에 따라 드물어졌다.[79][80][81]
고생물학자들은 화석 기록의 불완전성 때문에 생물의 실제 생존 시기보다 화석 발견 시기가 늦어질 수 있다는 시그노어-립스효과를 고려해야 한다.[82] 따라서 할키에리아류는 화석으로 알려진 것보다 더 이전부터 존재하고 더 늦게까지 살아남았을 가능성이 있다.
4. 1. 서식지
거의 완벽한 할키에리아 에반겔리스타 표본은 그린란드의 시리우스 파세트 라거슈태테에서 발견되었다.[57] 할키에리아류 화석으로 확실하게 분류될 수 있는 것은 중국 신장 지방과[75] 오스트레일리아 조지나 분지에서,[60] 껍질일 수도 있는 화석은 캐나다 버제스 셰일에서 발견되었다.[58] 할키에리아류 갑옷판, 즉 골편으로 보이는 것들은 작은껍질화석의 일부로 여러 곳에서 발견되었다.[61]할키에리아류 골편 중 가장 오래된 것은 할키에리아 롱가(''Halkieria longa'')인데, 시베리아 상부 네마키트-달디니아절 지층의 푸렐라 안티쿠아 존에서 발견되었다.[76] 캄브리아기 보토마절 말 대량멸종으로 할키에리아류를 포함, 대부분 작은껍질화석들이 멸종했으리라 생각되었으나 2004년 오스트레일리아 조지나 분지의 중기 캄브리아기 지층에서 할키에리아류로 분류되는 아우스트랄로할키에리아가 발견되었다. 다른 할키에리아류 분지군이 멸종하는 가운데 어떻게 아우스트랄로할키에리아가 포함된 분지군이 살아남을 수 있었는지는 알려지지 않았다.[60] 보토마절 말 멸종에서 살아남은 유일한 고배류 화석 역시 남아메리카, 아프리카, 인도, 오스트레일리아, 남극을 포함하는 초대륙 곤드와나에서 발견되는 것을 감안하면 이것이 중요한 함의를 가질 수도 있다.[77][78][60]
할키에리아류와 그 외 작은껍질화석들은 항상은 아니지만 대개는 인산염으로 보존되어 있는데, 이들의 원래 광물 성분은 인산염일 수도 있고 아닐 수도 있다. 인산염으로 덮여 보존되는 방식은 초기 캄브리아기에는 흔했으나 시간이 지남에 따라 해저 바닥을 파고 들어가는 동물들의 교란 작용이 증가함에 따라 드물어졌다. 할키에리아류와 그 외 작은껍질화석들은 화석으로 알려진 것보다 더 이전부터 존재하기 시작해 더 늦게까지 살아남았을 수도 있다.[79][80][81] 고생물학자들은 이런 종류의 불확실성을 시그노어-립스효과라고 부른다.[82]
4. 2. 이동 방식
조지나 분지에서 발견된 다른 경화편들은 ''Australohalkieria superstes''와 구분될 정도로 충분히 다르지만, 분류할 수 있을 만큼 충분한 세부 정보를 제공할 만큼 충분하지는 않다. 한 종류는 두 갈래로 갈라진 끝을 가지고 있다는 점에서 ''A.superstes''의 경화편과 매우 유사하지만, 중간 관이 평평하지 않다. 다른 종류는 납작한 중앙 관과 종 방향 관이 없으며, 이는 초기 캄브리아기 ''할키에리아''와 ''Australohalkieria''와 구별되는 또 다른 중기 캄브리아기 할키에리아류 속을 나타낼 수 있다.[25]''로마술카키테스(Lomasulcachites)''는 경화판만 알려진 또 다른 속이다.[9]
''Sinosachites''는 경화기만으로 알려진 '할키에리아류'의 한 속이다. 이 경화기는 중앙 관에 수직에 가까운 내부 챔버를 가지고 있으며, 좁은 채널로 연결되어 있다.[15][13] 챔버는 ''Australohalkieria''의 세로 관과 동일한 직경(40μm)을 가지며, 그 수가 더 많고 세로가 아닌 가로로 배열되어 있는 것은 길이가 약 1mm–2mm인 ''Sinosachites'' 경화기의 더 큰 크기를 반영한다.[15]
이 경화기는 원래 호주에서 기술된 ''Thambetolepis''와 동의어이다. 좌우 경화기가 존재하므로, 이 동물은 양측 대칭이었으며, ''할키에리아''와 마찬가지로 손바닥 모양, 칼 모양 및 낫 모양의 경화기 형태가 존재한다.[15]
4. 3. 섭식
버제스 셰일에서 발견된 두 종류의 뚜껑 모양 껍데기로만 알려진 ''오이코제테테스''(Oikozetetes)[16]는 약 5억 5백만년 전 시기로 추정된다. 이 두 종류는 할키에리아류의 앞면과 뒷면 껍데기로 여겨진다.[17] 이들은 생존 당시에는 아마도 석회질이었을 것이다(비록 속성 작용으로 인해 원래의 광물이 실리카와 같은 다른 물질로 대체되는 경우도 있지만).[20] 또한 ''할키에리아''(Halkieria)와 같이 생물 광물화된 경화판으로 구성된 갑옷 덮개를 지녔을 것으로 생각된다. 이러한 경화판은 껍데기와 직접적인 연관성을 가지는 경우는 절대 발견되지 않지만, 이러한 사실을 설명할 수 있는 많은 생물층적 과정이 존재한다.[18]하부 캄브리아기 분류군 ''오크루라누스''(Ocruranus) (=''에오할로비아''(Eohalobia))는 ''오이코제테테스''(Oikozetetes)의 껍데기와 동일하다고 추정되며,[18] 겉보기에는 할키에리아류와 유사한 몸체에 속했지만,[21] 중간 껍데기는 고생물 로리카류와 유사한 형태를 시사한다.[22] ''로마술카키테스''(Lomasulcachites)는 경화판만 알려진 또 다른 속이다.[9]
5. 분류 및 진화
오스트레일리아에서 발견된 아우스트랄로할키에리아 수페르스테스(Australohalkieria superstes|아우스트랄로할키에리아 수페르스테스영어)는 보토마절 말의 멸종 사건에서 살아남은 할키에리아류이다. 이 종의 골편은 윗면은 볼록하고 아랫면은 오목하며, 서로 겹쳐져 있다. 골편 내부에는 복잡한 빈 공간이 있는데, 원통 모양의 관이 한 쌍의 가는 관으로 나뉘고, 가운데 부분은 납작해져 있다. 골편 윗부분에는 움푹 들어간 자국이 있는데, 이는 후각 기관 역할을 했을 가능성이 있다. 골편 표면은 선인장의 골편처럼 유기질로 덮여 있었을 가능성이 있으며, 이는 현생 연체동물의 각피층과 유사했을 수 있다.[60]
아우스트랄로할키에리아 수페르스테스의 골편은 좌우 대칭을 이루는 두 종류로 구성되어 있으며, 크기는 매우 작다. 초기 캄브리아기 할키에리아류와 비교하면 낫형 골편이 더 많고 손바닥형 골편은 드물며, 크기도 1/3 정도 작다. 이는 사망 후 골편이 유실되었거나, 아우스트랄로할키에리아 수페르스테스의 골편체가 더 많은 골편으로 구성되었거나, 원래 크기가 작았을 가능성 등이 원인으로 제시된다. 조지나 분지에서는 할키에리아류의 껍질 화석이 발견되지 않았지만, 할키에리아의 껍질도 드물게 발견되므로 아우스트랄로할키에리아가 껍질을 가지지 않았다는 의미는 아니다.[60]
시포고누키테스류(Siphogonuchitids)는 두 가지 형태의 골편과 껍질을 가지며, 할키에리아류보다 단순한 골편체와 내부 구조를 가진다.[65] 시포고누키테스(Siphogonuchites|시포고누키테스영어), 다바샤니테스(Dabashanites|다바샤니테스영어), 로포키테스(Lopochites|로포키테스영어), 마이카넬라(Maikhanella|마이카넬라영어)는 모두 유사한 형태를 가지며,[65] 드레파노키테스(Drepanochites|드레파노키테스영어)는 골편의 비율로 구분된다.[65] 마이카넬라의 껍질은 시포고누키테스의 골편이 석회화된 기질과 합쳐진 형태이며, 어린 개체는 골편과 합쳐지지 않은 것으로 보인다.[66] 시포고누키테스 골편 내부의 빈 공간은 단순하며 측면에 연결된 공간은 없다.[63]
사키테스(Sachites|사키테스영어)는 가시 모양 골편으로 이루어진 속으로, 많은 표본이 다른 할키에리아류로 분류되었다.[68] 사키테스는 할키에리아류와 가깝다고 생각되었으나,[69] 최근에는 챈슬로리드와 근연관계라는 주장이 제기되었다.[70]
시노사키테스(Sinosachites|시노사키테스영어)는 골편만 발견된 할키에리아류로, 골편 내부에는 중앙 관과 수직으로 나 있는 방 모양의 공간이 있으며, 이 공간은 좁은 관을 통해 중앙으로 이어진다.[63][69] 시노사키테스의 골편은 오스트레일리아에서 처음 발견된 탐베톨레피스(Thambetolepis|탐베톨레피스영어)와 동일하며, 좌우 대칭의 몸을 가졌던 것으로 보인다.[63]
할키에리아류의 진화적 관계는 복잡하고 논쟁이 진행 중이다. 주요 논쟁은 할키에리아류와 ''위왁시아''의 관계, 그리고 연체동물, 환형동물, 완족동물과의 관계에 대한 것이다. 챈슬로리아류와의 관계 또한 중요한 문제이다.
- 1995년 콘웨이 모리스와 필은 분자 계통학 연구를 바탕으로 한 분기도를 발표했다.[6]
- 2003년, 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지하며, 완족동물이 몸길이가 짧고 껍질이 큰 할키에리아류 계통에서 발생했을 수 있다고 제안했다.[36]
- 빈터와 닐슨(2005)은 ''Halkieria''가 관생 그룹 연체동물이라고 제안했다. ''Halkieria''의 경화는 현대의 무판강 껍질 없는 연체동물, 일부 현대 다판강 연체동물의 경화, 오르도비스기 다판강 ''Echinochiton''의 경화와 유사했다. ''Halkieria''의 껍질은 껍질강 연체동물의 껍질과 더 유사했다.[37]
- 캐런, 쉘테마, 샨더 및 럿킨(2006)도 ''Halkieria''를 관생 그룹 연체동물로 해석했으며, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다.[38]
- 2006년, 콘웨이 모리스는 빈터와 닐슨(2005)의 ''Halkieria''를 관생 그룹 연체동물로 분류한 것을 비판했다.[40]
- 버터필드(2006)는 ''Wiwaxia''와 ''Odontogriphus''가 줄기 그룹 연체동물이 아니라 줄기 그룹 다모류라고 주장했다.[41]
- 콘웨이 모리스와 캐런(2007)은 ''Orthrozanclus reburrus''에 대한 첫 번째 설명을 발표했다. 이는 할키에리아류와 유사하게 동심원 띠의 경화를 가지고 있었지만, 광물화되지 않은 2개의 경화만 있었고, 앞쪽으로 추정되는 껍질 하나는 ''Halkieria''의 앞쪽 껍질과 모양이 유사했다. 또한 ''Wiwaxia''와 매우 유사한 긴 가시가 있었다.
- 할키에리아류 ''Sinosachites''의 경화 내부에 있는 내부 공동의 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 친화성을 강화했다.[15]
- 포터(2008)는 ''할키에리아'' 경화기가 칸첼로리아류 경화기와 매우 유사하다는 1980년대 초의 아이디어를 되살렸다. 칸첼로리아류는 위쪽이 열린 자루 모양의 방사상 대칭 생물이었다.[45]
5. 1. 할키에리아류 (Halkieriids)
할키에리아류는 캄브리아기 초기에서 중기까지 살았던 생물군으로, 몸은 여러 종류의 작은 골편(경화)으로 덮여 있고, 앞뒤 양 끝에는 껍데기를 가진 것이 특징이다.오스트레일리아에서 발견된 아우스트랄로할키에리아 수페르스테스(Australohalkieria superstes영어)는 "남쪽에서 살아남은 할키에리아"라는 뜻으로, 보토마절 말의 멸종 사건에서 살아남은 종이다. 이 종의 골편은 위쪽은 볼록하고 아래쪽은 오목하며, 서로 겹쳐져 있다. 골편 내부에는 복잡한 빈 공간이 있는데, 원통 모양의 관이 한 쌍의 가는 관으로 나뉘고, 가운데 부분은 납작해져 있다. 골편 윗부분은 납작하고 움푹 들어간 자국이 있는데, 이는 후각 기관의 역할을 했을 가능성이 있다. 골편 표면은 선인장의 골편처럼 유기질로 덮여 있었을 가능성이 있으며, 이는 현생 연체동물의 각피층과 유사했을 수 있다.[60]
아우스트랄로할키에리아 수페르스테스의 골편은 좌우 대칭을 이루는 두 종류로 구성되어 있으며, 크기는 매우 작다. 손바닥형은 250µm에서 650µm, 칼날형은 300µm에서 1000µm, 낫형은 400µm에서 1000µm이거나 400µm에서 500µm이다. 초기 캄브리아기 할키에리아류와 비교하면 낫형 골편이 더 많고 손바닥형 골편은 드물며, 크기도 1/3 정도 작다. 이는 사망 후 골편이 유실되었거나, 아우스트랄로할키에리아 수페르스테스의 골편체가 더 많은 골편으로 구성되었거나, 원래 크기가 작았을 가능성 등이 원인으로 제시된다. 조지나 분지에서는 할키에리아류의 껍질 화석이 발견되지 않았지만, 할키에리아의 껍질도 드물게 발견되므로 아우스트랄로할키에리아가 껍질을 가지지 않았다는 의미는 아니다.[60]
시포고누키테스류(Siphogonuchitids)는 두 가지 형태의 골편과 껍질을 가지며, 할키에리아류보다 단순한 골편체와 내부 구조를 가진다.[65] 시포고누키테스(Siphogonuchites영어), 다바샤니테스(Dabashanites영어), 로포키테스(Lopochites영어), 마이카넬라(Maikhanella영어)는 모두 유사한 형태를 가지며,[65] 드레파노키테스(Drepanochites영어)는 골편의 비율로 구분된다.[65] 마이카넬라의 껍질은 시포고누키테스의 골편이 석회화된 기질과 합쳐진 형태이며, 어린 개체는 골편과 합쳐지지 않은 것으로 보인다.[66] 시포고누키테스 골편 내부의 빈 공간은 단순하며 측면에 연결된 공간은 없다.[63]
니넬라(Ninella영어)로 대표되는 니넬라류(Ninellids)는 하부 캄브리아기 지층에서 발견되며, 한 종류의 골편만으로 구성된 단순한 골편체를 가진다. 골편은 갈고리 형태로, 석회암 성분이며 속이 비어 있고 표면에 이랑 구조가 있다.[65]
히포파란기테스(Hippopharangites영어)[67]의 골편은 중앙에 넓은 빈 공간과 껍질 전체에 작은 기공을 가지는데, 이는 다른 할키에리아류의 측면 공간이나 다판류의 지상감각기에 해당할 수 있다.[63] 히포파란기테스 속은 챈슬로리드의 골편과 유사하여, 할키에리아류와 챈슬로리드를 실로스클레리토포라(Coeloscleritophora)로 분류하는 증거로 제시되기도 한다.[65]
로마술카키테스(Lomasulcachites영어)는 골편으로만 알려진 속이다.[65] 사키테스(Sachites영어)는 가시 모양 골편으로 이루어진 속으로, 많은 표본이 다른 할키에리아류로 분류되었다.[68] 사키테스는 할키에리아류와 가깝다고 생각되었으나,[69] 최근에는 챈슬로리드와 근연관계라는 주장이 제기되었다.[70]
시노사키테스(Sinosachites영어)는 골편만 발견된 할키에리아류로, 골편 내부에는 중앙 관과 수직으로 나 있는 방 모양의 공간이 있으며, 이 공간은 좁은 관을 통해 중앙으로 이어진다.[63][69] 방 모양의 공간은 아우스트랄로할키에리아의 긴 관과 비슷하게 40µm 정도의 지름을 가지며, 시노사키테스의 골편은 다른 할키에리아류보다 크다.[63] 시노사키테스의 골편은 오스트레일리아에서 처음 발견된 탐베톨레피스(Thambetolepis영어)와 동일하며, 좌우 대칭의 몸을 가졌던 것으로 보인다. 할키에리아와 마찬가지로 손바닥형, 칼날형, 낫형으로 구분할 수 있다.[63]
오이코제테테스(Oikozetetes영어)[71]는 버제스 셰일에서 발견된 두 종류의 모자 형태 껍질로만 알려져 있으며, 5억 5백만 년 전으로 연대가 확인되었다. 두 종류의 껍질은 할키에리아류의 앞쪽 껍질과 뒤쪽 껍질로 추정된다.[58] 오이코제테테스의 껍질은 석회질 성분이었으며,[72] 할키에리아처럼 갑옷에 해당하는 골편을 가졌을 것으로 보이지만, 껍질과 함께 발견된 적은 없다.[72]
하부 캄브리아기 지층에서 발견되는 오크루라누스(Ocruranus영어) 또는 에오할로비아(Eohalobia영어)는 오이코제테테스의 껍질과 동일하며 할키에리아류 형태의 몸에 붙어 있었을 것으로 보이지만,[73] 중간 판을 보면 팔레오로리카타(Palaeoloricata)와 유사한 형태였을 수도 있다.[74]
"경화판"이라고 불리는 덮개는 소형 쉘 화석의 구성 요소로 알려져 왔으며, 일부는 동일한 동물에 속하며 함께 맞춰진다. ''Halkieria evangelista''의 첫 번째 연결 표본은 1989년 그린란드의 시리우스 파셋 라거슈테트에서 수집되었으며, 1990년 사이먼 컨웨이 모리스와 존 S. 필에 의해 기술되었다.[3] ''H. evangelista''는 다른 유사한 껍질과 경화판을 할키에리아류로 식별하고 재구성하는 모델로 사용되며,[25][17] 그 종명 evangelistala는 초기 캄브리아기 화석 기록을 설명하는 능력을 반영한다.[6]
Siphogonuchitids는 껍데기뿐만 아니라 두 가지 경화판 형태를 가지고 있어, 경화판의 내부 구조가 더 단순하다는 점과 일치하여, ''Halkieria''와 그 족속들보다 더 단순한 경화판체를 가지고 있었을 수 있다.[9]
''Siphogonuchites'', ''Dabashanites'', ''Lopochites'', 그리고 ''Maikhanella'' 속은 모두 ''Siphogonuchites'' 동물체의 구성 요소를 나타내는 것으로 보인다.[9]
''Drepanochites''의 경화판은 종횡비에 따라 구별할 수 있다.[9]
''Maikhanella''는 석회화된 기질과 함께 융합된 ''Siphogonuchites'' 경화판으로 형성된 껍데기이다. 유생 껍데기는 경화판을 포함하지 않는 것으로 보인다.[10]
''Siphogonuchites'' 경화판의 중심 공동은 측면 방이 부착되어 있지 않은 단순한 구조이다.[15]
닌넬리드(ninellids)는 전형적으로 ''닌넬라''(Ninella)로 대표되며, 하부 캄브리아기에 존재했던 분류군으로, 단 하나의 경화자(sclerite) 유형만을 가진 훨씬 단순한 경화피(scleritome)를 가지고 있었다(경화자의 형태적 변이가 관찰되며, 좌우 대칭의 경화자가 존재한다). 이들의 경화자는 갈고리 모양 또는 국자 모양이며, 할키에리아류(halkieriid) 또는 시포노구누키티드(siphonogunuchitid)의 경화자와 매우 유사했다; 이들은 속이 비어 있고 석회질이었으며, 융기된 윗면을 가지고 있었다.[9]
''히포파랑기테스''(Hippopharangites)[11]는 넓은 중앙 공동과 껍질 벽을 통해 열리는 작은 구멍을 가진 경화기를 가지고 있으며, 이는 다른 할키에리아류의 측면 챔버(그리고 키톤의 미세감각관?)와 동일하다.[15]
이 속은 칸셀로리아류 경화기의 형태와 가장 유사하며, 따라서 할키에리아류와 칸셀로리아류를 Coeloscleritophora로 통합하는 것을 지지하는 데 사용된다.[9]
할키에리아류(속)의 거의 모든 종은 분리된 비늘 모양의 경화기 발견만으로 알려져 있다.
| 학명 | 명명자 | 비고 |
|---|---|---|
| Halkieria alata | Duan, 1984 | |
| Halkieria amorpha | Meshkova,1974 | |
| Halkieria bisulcata | Qian et Yin, 1984 | |
| Halkieria costulata | Meshkova, 1974 | |
| Halkieria curvativa | Mambetov in Missarzhevsky and Mambetov, 1981 | |
| Halkieria deplanatiformis | Mambetov in Missarzhevsky and Mambetov, 1981 | |
| Halkieria desquamata | Duan, 1984 | |
| Halkieria directa | Mostler, 1980 | |
| Halkieria elonga | Qian et Yin, 1984 | |
| Halkieria equilateralis | Qian et Yin, 1984 | |
| Halkieria folliformis | Duan, 1984 | |
| Halkieria fordi | Landing, 1991 | |
| Halkieria hexagona | Mostler, 1980 | |
| Halkieria lata | Mostler, 1980 | |
| Halkieria longa | Qian, 1977 | |
| Halkieria longispinosa | Mostler, 1980 | |
| Halkieria maidipingensis | Qian, 1977 | |
| Halkieria mina | Qian, Chen et Chen, 1979 | |
| Halkieria mira | Qian et Xiao, 1984 | |
| Halkieria obliqua | Poulsen, 1967 | |
| Halkieria operculus | Qian, 1984 | |
| Halkieria pennata | He, 1981 | ?=Halkieria sthenobasis Jiang in Luo et al., 1982 |
| Halkieria phylloidea | He, 1981 | |
| Halkieria praeinguis | Jiang in Luo et al., 1982 | |
| Halkieria projecta | Bokova, 1985 | |
| Halkieria sacciformis | Meshkova, 1969 | |
| Halkieria solida | Mostler, 1980 | |
| Halkieria sthenobasis | Jiang in Luo et al., 1982 | |
| Halkieria stonei | Landing, 1989 | |
| Halkieria symmetrica | Poulsen, 1967 | |
| Halkieria terastios | Qian, Chen et Chen, 1979 | |
| Halkieria uncostata | Qian et Yin, 1984 | |
| Halkieria undulata | Wang, 1994 | |
| Halkieria ventricosa | Mostler, 1980 | |
| Halkieria wangi | Demidenko, 2010 | |
| Halkieria zapfei | Mostler, 1980 |
현재, 완전한 경화피의 구조는 그린란드(시리우스 파셋 층)의 전기 캄브리아기에서 발견된 단일 종인 ''Halkieria evangelista''에 대해서만 알려져 있다.
할키에리아류의 진화적 관계는 복잡하고 논쟁이 진행 중이다. 주요 논쟁은 할키에리아류와 ''위왁시아''의 관계, 그리고 연체동물, 환형동물, 완족동물과의 관계에 대한 것이다. 챈슬로리아류와의 관계 또한 중요한 문제이다.
1995년, 콘웨이 모리스와 필은 분자 계통학 연구를 바탕으로 한 분기도를 발표했다.[6]
- 시포고노투키다는 초기 캄브리아기 암석에서 발견된 그룹으로, 나머지 모든 그룹의 "자매" 그룹이다.[6]
- 초기 할키에리아류는 연체동물의 "자매" 그룹이었다.
- ''Thambetolepis'' / ''Sinosachites''는 환형동물의 "고모 할머니", ''Wiwaxia''는 환형동물의 "고모"였다.
- ''Halkieria evangelista''는 연체동물과 완족동물의 "자매" 그룹이었다.
2003년, 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지하며, 완족동물이 몸길이가 짧고 껍질이 큰 할키에리아류 계통에서 발생했을 수 있다고 제안했다.[36]
빈터와 닐슨(2005)은 ''Halkieria''가 관생 그룹 연체동물이라고 제안했다. ''Halkieria''의 경화는 현대의 무판강 껍질 없는 연체동물, 일부 현대 다판강 연체동물의 경화, 오르도비스기 다판강 ''Echinochiton''의 경화와 유사했다. ''Halkieria''의 껍질은 껍질강 연체동물의 껍질과 더 유사했다.[37]
캐런, 쉘테마, 샨더 및 럿킨(2006)도 ''Halkieria''를 관생 그룹 연체동물로 해석했으며, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다.[38]
2006년, 콘웨이 모리스는 빈터와 닐슨(2005)의 ''Halkieria''를 관생 그룹 연체동물로 분류한 것을 비판했다.[40]
버터필드(2006)는 ''Wiwaxia''와 ''Odontogriphus''가 줄기 그룹 연체동물이 아니라 줄기 그룹 다모류라고 주장했다.[41]
콘웨이 모리스와 캐런(2007)은 ''Orthrozanclus reburrus''에 대한 첫 번째 설명을 발표했다. 이는 할키에리아류와 유사하게 동심원 띠의 경화를 가지고 있었지만, 광물화되지 않은 2개의 경화만 있었고, 앞쪽으로 추정되는 껍질 하나는 ''Halkieria''의 앞쪽 껍질과 모양이 유사했다. 또한 ''Wiwaxia''와 매우 유사한 긴 가시가 있었다.
할키에리아류 ''Sinosachites''의 경화 내부에 있는 내부 공동의 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 친화성을 강화했다.[15]
5. 2. 연체동물과의 관계
할키에리아류의 진화적 관계는 복잡하고 논쟁의 여지가 많다. 특히 위왁시아 및 주요 로포트로코조아 문인 연체동물, 환형동물, 완족동물과의 관계가 주된 논쟁 대상이다. 찬셀로리아류와의 관계 또한 중요한 문제이다.1995년 콘웨이 모리스와 필은 화석 특징과 분자 계통학 연구를 바탕으로 한 분기도를 제시했다.[6]
- 시포고노투키다는 초기 캄브리아기 암석에서 발견된 그룹으로, 나머지 그룹의 "자매" 그룹이다.[6]
- 초기 할키에리아류는 연체동물의 "자매" 그룹으로, 근육질 발을 가졌을 것으로 추정된다.[6]
- ''Thambetolepis''는 환형동물의 "고모 할머니", ''Wiwaxia''는 환형동물의 "고모"로, 두 그룹 모두 3개의 동심원 구역으로 나뉜 경화를 가진다는 점이 유사하다. ''Wiwaxia''와 환형동물의 관계는 ''Wiwaxia''의 경화와 다모류 환형동물의 강모 사이의 유사성에 기반한다.[6][34]
- ''Halkieria evangelista''는 완족동물의 "자매" 그룹으로, 완족동물은 할키에리아류와 유사한 유충 단계를 거치며, 일부 화석 껍질은 완족동물과 유사한 미세 구조를 가진다.[6][35]
2003년 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지하며, 완족동물이 몸길이가 짧고 껍질이 큰 할키에리아류 계통에서 발생하여 스스로 접었을 수 있다고 제안했다.[36]
반면 빈터와 닐슨(2005)은 ''Halkieria''가 관생 그룹 연체동물이라고 주장했다. 이들은 ''Halkieria''의 경화가 현대 무판강 연체동물, 일부 현대 다판강 연체동물의 경화와 유사하다고 보았다. ''Halkieria''의 껍질은 껍질강 연체동물의 껍질과 더 유사하며, 완족동물과 환형동물의 강모는 서로 유사하지만 ''Halkieria''의 경화와는 다르다고 주장했다.[37]
캐런, 쉘테마, 샨더 및 럿킨(2006)도 ''Halkieria''를 관생 그룹 연체동물로, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다. 이들은 ''Halkieria''가 탄산 칼슘 갑옷을 가졌고, ''Wiwaxia''와 ''Odontogriphus''는 연체동물 치설을 가졌다고 보았다.[38][39]
2006년 콘웨이 모리스는 빈터와 닐슨(2005)의 주장을 비판하며, 할키에리아류가 연체동물과 완족동물 모두의 조상에 가깝다는 이전 결론을 반복했다.[40]
버터필드(2006)는 ''Wiwaxia''와 ''Odontogriphus''가 줄기 그룹 다모류라고 주장하며, 이들의 섭식 장치가 연체동물 치설과 다르다고 보았다. 또한 ''Odontogriphus'' 화석의 분절 증거를 제시하며, ''Wiwaxia''의 경화와 다모류 강모의 유사성을 강조했다.[41]
콘웨이 모리스와 캐런(2007)은 ''Orthrozanclus reburrus''를 발표하며, "할왁시드"가 유효한 분류군이라고 주장했다. 이들은 연체동물, 환형동물, 완족동물을 포함하는 로포트로코아 진화에 대한 두 가지 분기도를 제시했다.[33]
할키에리아류 ''Sinosachites'' 경화 내부의 공동 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 관계를 강화한다.[15]
포터(2008)는 ''할키에리아'' 경화기가 칸첼로리아류 경화기와 매우 유사하다는 아이디어를 되살렸다. 칸첼로리아류는 방사상 대칭 생물로, 처음에는 해면동물로 분류되었으나, 이후 연구에서 상피동물의 구성원일 가능성이 제기되었다.[45]
할키에리아류와 칸첼로리아류의 공동 경화기는 여러 면에서 유사하지만, 신체 구조의 차이로 인해 이들의 관계는 불분명하다. 이 딜레마는 다양한 방식으로 해석될 수 있다.[45]
할키에리아는 앞뒤가 원형인 직사각형이며, 전신이 인편으로 덮여 있고, 앞뒤에 "껍데기"를 가진다. 콘웨이 모리스 등은 이 "껍데기"가 합쳐져 완족동물이 탄생했다는 가설을 제시했으나,[51] 이후 "껍데기"가 1개뿐인 오르스로잔클루스가 발견되면서 이 가설의 가능성은 낮아졌다.
또한, 인편 외에 치설을 가진 화석도 발견되어[52] 위왁시아와 근연 관계이며, ''Halwaxiida'' 강을 이룬다고 여겨진다. 오돈토그리푸스와의 관계도 지적된다.[53][54]
5. 3. 완족동물과의 관계
할키에리아류의 진화적 관계는 복잡하고 논쟁의 여지가 많은 주제이다. 논쟁의 중심에는 할키에리아류와 위왁시아 사이의 관계, 그리고 연체동물, 환형동물, 완족동물이라는 세 개의 주요 로포트로코조아 문과의 관계가 있다. 캄브리아기의 더 원시적인 그룹인 찬셀로리아류와의 관계 또한 중요한 문제이다.1995년 콘웨이 모리스와 필은 화석 특징과 DNA 및 RNA 분자 계통학 연구를 바탕으로 다음과 같은 분기도를 발표했다.[6]
- 시포고노투키다는 초기 캄브리아기 암석에서 발견된 그룹으로, 다른 모든 그룹의 "자매" 그룹이다.[6][33]
- 초기 할키에리아류는 연체동물의 "자매" 그룹으로, 비교적 가까운 공통 조상의 후손이다.[6]
- ''Thambetolepis'' / ''Sinosachites''는 환형동물의 "고모 할머니"였고, ''Wiwaxia''는 환형동물의 "고모"였다. 이들은 두 그룹 모두 3개의 동심원 구역으로 나뉜 경화를 가졌다는 점을 근거로 들었다.[6]
- ''Canadia''는 버제스 셰일 화석으로, ''Wiwaxia'' 경화와 다모류 환형동물의 강모 사이의 유사성을 기반으로 한다.[6][34]
- 그린란드의 시리우스 파셋 화석 산지에서 발견된 ''Halkieria evangelista''는 완족동물의 "자매" 그룹이었다. 완족동물은 환형동물의 강모와 유사하며, ''Wiwaxia'' 경화와도 유사하다.[6]
2003년, 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지하며, 완족동물이 몸길이가 짧고 껍질이 큰 할키에리아류 계통에서 발생했을 수 있다고 제안했다.[36]
2005년, 빈터와 닐슨은 ''Halkieria''가 관생 그룹 연체동물이라고 제안했다. 이들은 ''Halkieria'' 경화가 현대 무판강 무판류 껍질 없는 연체동물, 일부 현대 다판강 연체동물, 오르도비스기 다판강 ''Echinochiton''의 경화와 유사하다고 주장했다. 또한 ''Halkieria'' 껍질은 껍질강 연체동물의 껍질과 더 유사하며, 완족동물과 환형동물의 강모는 서로 유사하지만 ''Halkieria'' 경화와는 유사하지 않다고 했다.[37]
2006년, 캐런, 쉘테마, 샨더 및 럿킨도 ''Halkieria''를 관생 그룹 연체동물로 해석하고, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다.[38] 이들은 ''Halkieria''와 관생 그룹 연체동물 모두 탄산 칼슘으로 광물화된 갑옷을 가졌다는 점을 근거로 들었다.[39]
같은 해 콘웨이 모리스는 빈터와 닐슨(2005)의 ''Halkieria'' 분류를 비판하며, 무판류와 다판류의 가시 성장이 할키에리아류 경화와 유사하지 않다고 지적했다. 그는 할키에리아류가 연체동물과 완족동물 모두의 조상에 가깝다는 이전 결론을 반복했다.[40]
버터필드(2006)는 ''Wiwaxia''와 ''Odontogriphus''가 밀접하게 관련되어 있지만, 줄기 그룹 연체동물이 아니라 줄기 그룹 다모류라고 주장했다. 그는 ''Wiwaxia'' 경화와 다모류 강모 사이의 내부 구조 유사성을 강조했다.[41]
콘웨이 모리스와 캐런(2007)은 ''Orthrozanclus reburrus''를 발표하며, "할왁시드"가 유효한 분류군이며, 로포트로코아 진화에 대한 대안적 가설을 제시했다.[33]
할키에리아류 ''Sinosachites'' 경화 내부의 공동 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 친화성을 강화했다.[15]
할키에리아는 앞뒤가 원형인 직사각형이며, 전신이 인편으로 덮여 있고, 몸 앞뒤에 "껍데기"를 갖는다. 콘웨이 모리스 등은 이 "껍데기"가 합쳐져 완족동물이 탄생했다는 가설을 제창했다.[51] 그러나 ''할키에리아''보다 2000만 년 이상 지난 지층에서 껍질이 하나뿐인 오르스로잔클루스가 발견되어 이 가설의 가능성은 낮아졌다.
인편 외에도 치설을 가진 화석도 발견되어, 위왁시아와 근연 관계이며, ''Halwaxiida'' 강(한국어 명칭 미정)을 이룬다고 여겨진다. 오돈토그리푸스와의 관계도 지적된다.[53][54]
5. 4. 환형동물과의 관계
할키에리아류의 진화적 관계는 복잡하고 논쟁의 여지가 많은 주제이다. 특히 할키에리아류와 위왁시아 사이의 관계, 그리고 연체동물, 환형동물, 완족동물이라는 세 개의 주요 로포트로코조아 문과의 관계에 대한 논쟁이 주를 이룬다. 겉보기에는 더 원시적인 캄브리아기 그룹인 찬셀로리아류와의 관계 또한 중요한 문제이다.1995년 콘웨이 모리스와 필은 화석 특징과 1990년대 초 분자 계통학 연구를 바탕으로 한 분기도를 발표했다.[6] 이 분기도에서는 다음과 같은 관계를 제시했다.
- 시포고노투키다는 초기 캄브리아기 암석에서 발견된 그룹으로, 나머지 모든 그룹의 "자매" 그룹이다.[6]
- 초기 할키에리아류는 연체동물의 "자매" 그룹이다.[6]
- ''Thambetolepis'' / ''Sinosachites''는 환형동물의 "고모 할머니"였고, ''Wiwaxia''는 환형동물의 "고모"였다. 이들은 두 그룹 모두 3개의 동심원 구역으로 나뉜 경화를 가지고 있다는 점을 근거로 들었다.[6]
- ''Canadia''는 버제스 셰일 화석으로, 일반적으로 다모류로 여겨진다.[6][34]
- 시리우스 파셋 화석 산지에서 발견된 ''Halkieria evangelista''는 완족동물의 "자매" 그룹이었다.[6]
2003년, 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지했다.[36]
2005년, 빈터와 닐슨은 ''Halkieria''가 관생 그룹 연체동물이라고 제안했다. 이들은 ''Halkieria'''s 경화가 현대 무판강 무판류 껍질 없는 연체동물, 일부 현대 다판강 연체동물의 경화, 오르도비스기 다판강 ''Echinochiton''의 경화와 유사하다고 주장했다. 또한 ''Halkieria'''s 껍질은 껍질강 연체동물의 껍질과 더 유사하다고 보았다.[37]
2006년, 캐런, 쉘테마, 샨더 및 럿킨도 ''Halkieria''를 관생 그룹 연체동물로 해석하고, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다.[38]
같은 해 콘웨이 모리스는 빈터와 닐슨(2005)의 ''Halkieria'' 분류를 비판하며, 할키에리아류가 연체동물과 완족동물 모두의 조상에 가깝다는 이전 결론을 반복했다.[40]
2006년 버터필드는 ''Wiwaxia''와 ''Odontogriphus''가 줄기 그룹 다모류라고 주장했다.[41]
2007년 콘웨이 모리스와 캐런은 ''Orthrozanclus reburrus''에 대한 설명을 발표하며 "할왁시드"가 유효한 분류군이라는 증거로 간주하고, 로포트로코아 진화에 대한 대안적 가설을 제시하는 두 개의 분기도를 발표했다.[33]
- 첫 번째 분기도는 ''Kimberella''와 ''Odontogriphus''를 초기 원시 연체동물로, ''Wiwaxia'', 시포고노투키다, ''Orthrozanclus'' 및 ''Halkieria''를 연체동물 계통의 측면 분지로 보았다.
- 두 번째 분기도는 ''Kimberella''와 ''Odontogriphus''를 초기 원시 로포트로코아로, 시포고노투키다, ''Halkieria'', ''Orthrozanclus'' 및 ''Wiwaxia''를 연체동물보다 환형동물과 완족동물의 공통 조상에 더 가까운 그룹으로 보았다.
할키에리아류 ''Sinosachites'' 경화 내부의 공동 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 친화성을 강화했다.[15]
2008년 포터는 ''할키에리아''의 경화기가 칸첼로리아류의 경화기와 매우 유사하다는 아이디어를 되살렸다.[45]
하지만, 칸첼로리아류와 할키에리아류의 신체 구조 차이로 인해 이들의 관계는 여전히 불확실하다.
5. 5. 할왁시아류 (Halwaxiids)
할키에리아류의 진화적 관계는 아직 논쟁 중인 복잡한 주제이다. 이러한 논쟁의 대부분은 할키에리아류와 ''위왁시아'' 사이의 관계, 그리고 연체동물, 환형동물, 완족동물의 3개 주요 로포트로코조아 문과의 관계에 대한 것이다. 겉보기에는 훨씬 더 원시적인 캄브리아기 그룹인 찬셀로리아류와의 관계에 대한 질문 역시 중요하다.[6]1995년, 콘웨이 모리스와 필은 화석의 특징과 1990년대 초 분자 계통학 연구를 바탕으로 한 분기도를 발표했다.[6]
- 시포고노투키다는 초기 캄브리아기 암석에서 발견된 그룹으로, 나머지 모든 그룹의 "자매" 그룹이었다.[6] 이들은 단편 조각으로만 알려져 있다.[33]
- 초기 할키에리아류는 연체동물의 "자매" 그룹이었다. 그들은 이러한 관계가 대부분의 연구자들이 할키에리아류가 가졌을 것으로 추정하는 근육질 발에 의해 뒷받침된다고 말했다.[6]
- 또 다른 할키에리아류 속인 ''Thambetolepis'' / ''Sinosachites''는 환형동물의 "고모 할머니"였고, ''Wiwaxia''는 환형동물의 "고모"였다. 할키에리아류와 ''Wiwaxia''의 밀접한 관계에 대한 그들의 주장은 두 그룹 모두 3개의 동심원 구역으로 나뉜 경화를 가지고 있다는 데 근거했다. ''Wiwaxia''와 환형동물의 밀접한 관계는 버터필드(1990)가 ''Wiwaxia'' 경화와 다모류 환형동물의 강모 사이에서 발견한 유사성에 근거했다. ''Canadia''는 일반적으로 다모류로 여겨지는 버제스 셰일 화석이다.[6][34]
- 콘웨이 모리스가 그린란드의 시리우스 파셋 화석 산지에서 발견한 ''Halkieria evangelista''는 현대 형태가 이매패 껍질을 가지고 있지만, 근육질 줄기와 독특한 섭식 장치인 촉수엽을 가지고 있다는 점에서 연체동물과 다른 동물인 완족동물의 "자매" 그룹이었다. 완족동물은 환형동물의 강모와 유사하고, 따라서 ''Wiwaxia'' 경화와 유사하며, 따라서 할키에리아류 경화와 유사한 강모를 가지고 있다.[6] 완족동물은 할키에리아류를 닮은 유충 단계를 거치고, 할키에리아류에 속한다고 생각되는 일부 고립된 화석 껍질이 완족동물과 유사한 미세 구조를 가지고 있기 때문에 완족동물과의 친화성이 타당해 보였다.[35]
2003년, 코헨, 홀머, 루터는 할키에리아류-완족동물 관계를 지지하며, 완족동물이 몸길이가 짧고 껍질이 큰 할키에리아류 계통에서 발생하여 스스로 접고, 결국에는 뒤쪽에서 줄기를 키웠을 수 있다고 제안했다.[36]
빈터와 닐슨(2005)은 그 대신 ''Halkieria''가 관생 그룹 연체동물이라고 제안했다. 그들은 ''Halkieria'' 경화가 현대의 무판강 무판류 껍질 없는 연체동물, 일부 현대 다판강 연체동물의 경화, 그리고 오르도비스기 다판강 ''Echinochiton''의 경화와 유사했다고 주장했다. ''Halkieria'' 껍질은 껍질강 연체동물의 껍질과 더 유사한데, 이 두 그룹의 껍질 모두 다판강 껍질 판에서 보이는 운하와 구멍의 흔적이 없기 때문이다. 완족동물과 환형동물의 강모는 서로 유사하지만 ''Halkieria'' 경화와는 유사하지 않다.[37]
캐런, 쉘테마, 샨더 및 럿킨(2006)도 ''Halkieria''를 관생 그룹 연체동물로 해석했으며, ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 해석했다.[38] ''Halkieria''를 관생 그룹 연체동물로 간주하는 주된 이유는 둘 다 탄산 칼슘으로 광물화된 갑옷을 가지고 있었기 때문이다. 그들은 ''Wiwaxia''와 ''Odontogriphus''를 줄기 그룹 연체동물로 취급했는데, 그 이유는 그들이 보기에는 둘 다 독특한 연체동물 치설, 즉 키틴질의 이빨이 있는 "혀"를 가지고 있었기 때문이다.[39]
2006년에 콘웨이 모리스는 빈터와 닐슨(2005)의 ''Halkieria''를 관생 그룹 연체동물로 분류한 것을 비판했는데, 무판류와 다판류의 가시 성장이 복잡한 할키에리아류 경화에 대해 추론된 성장 방식과 유사하지 않다는 점을 근거로 들었다. 특히, 그는 다양한 연체동물의 속이 빈 가시는 복잡한 내부 채널이 있는 할키에리아류 경화와 전혀 같지 않다고 말했다. 콘웨이 모리스는 할키에리아류가 연체동물과 완족동물 모두의 조상에 가깝다는 이전의 결론을 반복했다.[40]
버터필드(2006)는 ''Wiwaxia''와 ''Odontogriphus''가 밀접한 관련이 있다는 것을 받아들였지만, 그들이 줄기 그룹 연체동물이 아니라 줄기 그룹 다모류라고 주장했다. 그의 견해에 따르면, 이들 유기체의 섭식 장치는 2개 또는 최대 4개의 이빨 열로 구성되어 있어, 여러 이빨 열을 가진 "벨트형" 연체동물 치설의 기능을 수행할 수 없었다. ''Wiwaxia''와 ''Odontogriphus'' 이빨 열의 서로 다른 이빨 열 또한 눈에 띄게 다른 모양을 가지고 있는 반면, 연체동물 치설의 이빨은 동일한 "공장" 세포 그룹에 의해 차례로 생성되므로 거의 동일하다. 그는 또한 ''Odontogriphus'' 화석의 중간 부분을 가로지르는 선을 외부 분절의 증거로 간주했는데, 그 이유는 선이 균등하게 간격을 두고 있으며 신체의 긴 축에 정확히 직각으로 뻗어 있기 때문이다. 이전 논문에서와 마찬가지로 버터필드는 ''Wiwaxia'' 경화와 다모류의 강모 사이의 내부 구조의 유사성과 다모류가 등쪽을 덮는 강모를 형성하는 유일한 현대 유기체라는 사실을 강조했다.[41]
콘웨이 모리스와 캐런(2007)은 ''Orthrozanclus reburrus''에 대한 첫 번째 설명을 발표했다. 이는 할키에리아류와 유사하게 동심원 띠의 경화를 가지고 있었지만, 광물화되지 않은 2개의 경화만 있었고, 앞쪽으로 추정되는 껍질 하나는 ''Halkieria'' 앞쪽 껍질과 모양이 유사했다. 또한 ''Wiwaxia''와 매우 유사한 긴 가시가 있었다. 콘웨이 모리스와 캐런은 이 생물을 "할왁시드"가 유효한 분류군이라는 증거로 간주했다.[33] 그들은 연체동물, 환형동물 및 완족동물을 포함하는 계통인 로포트로코아의 진화에 대한 대안적 가설을 나타내는 ''두 개의'' 분기도를 발표했다:[33]
- 이것은 더 가능성이 높지만, 유기체의 특성이 약간이라도 변경되면 무너진다:[33]
- ''Kimberella''와 ''Odontogriphus''는 경화나 어떤 종류의 광물화된 갑옷도 없는 초기 원시 연체동물이다.
- ''Wiwaxia'', 시포고노투키다, ''Orthrozanclus'' 및 ''Halkieria''는 연체동물 계통수의 측면 분지에서 형성되었으며, 그 순서대로 분기되었다. 이것은 다음을 의미한다. ''Wiwaxia''가 경화를 처음 갖게 되었고, 경화는 광물화되지 않았다. 시포고노투키다는 광물화된 경화를 처음 갖게 되었지만, 경화체는 더 단순했다. 할키에리아류는 더 복잡한 경화체를 개발한 다음, ''Orthrozanclus''에서 경화체가 다시 광물화되지 않고 뒷껍질이 사라지거나 화석에서 보이지 않을 정도로 작아졌다. 이 가설은 시포고노투키다가 더 이른 시기에 나타나고 다른 세 그룹보다 더 단순한 경화체를 가지고 있다는 어려움에 직면한다.[33]
- 환형동물과 완족동물은 연체동물을 포함하지 않는 계통수의 다른 주요 분기에서 진화했다.
- 대안적 견해는 다음과 같다.
- ''Kimberella''와 ''Odontogriphus''는 초기 원시 로포트로코아이다.
- 시포고노투키다, ''Halkieria'', ''Orthrozanclus'' 및 ''Wiwaxia''는 연체동물보다 환형동물과 완족동물의 공통 조상에 더 가까운 그룹을 형성한다. 시포고노투키다는 ''두 가지'' 유형의 광물화된 경화와 융합된 경화로 만들어진 "껍질"을 갖게 되면서 이 그룹의 첫 번째로 구별된다. 할키에리아류는 세 가지 유형의 경화와 두 개의 일체형 껍질을 가지고 있었다. ''Orthrozanclus''에서 경화는 광물화되지 않았고, ''Wiwaxia''에서는 껍질이 손실되었다.[33]
할키에리아류 ''Sinosachites''의 경화 내부에 있는 내부 공동의 네트워크는 다판류의 미적 운하와 유사하여 연체동물과의 친화성을 강화했다.[15] 할키에리아는 앞뒤가 원형인 직사각형이며, 전신이 인편으로 덮여 있다. 특징적인 점은 몸의 앞뒤에 하나씩 "껍데기"를 갖는다는 것이다. Conway Morris 등은 이 "껍데기"가 합쳐져 완족동물이 탄생했다는 가설을 제창하고 있다.[51] 그러나 할키에리아보다 2000만 년 이상 지난 지층에서, "껍데기"를 1개만 가진 오르스로잔클루스가 발견되면서, 앞서 언급한 가설의 가능성은 낮아졌다.
또한, 인편을 가진 형태에 더해, 치설을 가진 화석도 발견[52]되어, 위왁시아와 근연 관계에 있으며, 단계통의 ''Halwaxiida'' 강(한국어 명칭 미정)을 이룬다고 여겨진다. 치설을 가진 점으로 미루어 오돈토그리푸스와의 관계도 지적되고 있다.[53][54]
6. 멸종
''Halkieria evangelista''의 유일하게 온전한 표본은 그린란드의 시리우스 파셋 라거슈테테에서 발견되었다.[3] 할키에리아류에 속한다고 확신할 수 있는 파편들은 중국의 신장 위구르 자치구[23]와 오스트레일리아의 조지나 분지에서 발견되었으며,[25] 가능한 할키에리아류의 껍질은 캐나다의 버제스 셰일에서 발견되었다.[17] "경판"이라고 불리는 할키에리아류와 유사한 갑피는 소형 쉘 화석의 일부로 다른 많은 장소에서 발견되었다.[4]
할키에리아류 경판의 가장 오래된 것으로 알려진 발생은 시베리아의 네마키트-달디니안절 상부의 ''Purella antiqua Zone''에서 발견된다.[24] 캄브리아기 시대의 보토미안절 말에 일어난 대멸종으로 할키에리아류를 포함한 대부분의 소형 쉘이 사라졌다고 생각되었지만, 2004년에는 ''Australohalkieria''로 분류된 할키에리아류 화석이 오스트레일리아의 조지나 분지의 중기 캄브리아기 암석에서 보고되었다. 이 분지군이 다른 할키에리아류 분지군이 멸종한 동안 살아남은 이유는 알려져 있지 않다.[25] 보토미안절 말 멸종에서 살아남은 것으로 알려진 유일한 고배류 또한 남아메리카, 아프리카, 인도, 오스트레일리아 및 남극 대륙을 포함하는 고대 초대륙인 곤드와나에서 발생한다는 점도 중요하다.[26][27][25]
할키에리아류와 다른 소형 쉘 화석은 일반적으로, 항상 그런 것은 아니지만, 인산염으로 보존되며, 이는 원래의 광물 조성일 수도 있고 아닐 수도 있다. 인산염 피복에 의한 보존은 초기 캄브리아기 동안에만 흔했던 것으로 보이며, 굴을 파는 동물의 해저 교란의 증가로 인해 시간이 지남에 따라 희귀해졌다. 따라서 할키에리아류와 다른 소형 쉘 화석은 가장 오래된 화석보다 더 일찍 살았고 가장 최근의 화석보다 더 늦게 살았을 수 있다[28][29][30] — 고생물학자들은 이러한 종류의 불확실성을 시뇨르-립스 효과라고 부른다.[31]
참조
[1]
간행물
Fossils from the Lower Cambrian of Bornholm
[2]
논문
Redescription of the Lower Cambrian ''Halkieria obliqua ''Poulsen
[3]
웹사이트
Showdown on the Burgess Shale
http://www.stephenja[...]
2008-07-31
[4]
논문
Early skeletal fossils
[5]
서적
Palaeobiology II
Wiley-Blackwell
2009-11-12
[6]
논문
Articulated Halkieriids from the Lower Cambrian of North Greenland and their Role in Early Protostome Evolution
[7]
논문
The Early Cambrian ''Halkieria'' is a mollusc
http://www.jakobvint[...]
2008-07-31
[8]
논문
Halkieriids in Middle Cambrian Phosphatic Limestones from Australia
2004-05
[9]
논문
Lower Cambrian coeloscleritophorans (''Ninella, Siphogonuchites'') from Xinjiang and Shaanxi, China
[10]
논문
The cap-shaped Cambrian fossil ''Maikhanella ''and the relationship between coeloscleritophorans and molluscs
[11]
논문
Skeletal Microstructure Indicates Chancelloriids and Halkieriids Are Closely Related
[12]
논문
Lower Cambrian Halkieriids and Other Coeloscleritophorans from Aksu-Wushi, Xinjiang, China
1997-01
[13]
논문
''Thambetolepis delicata'' gen. et sp. nov., an enigmatic fossil from the Early Cambrian of South Australia
[14]
논문
Small Shelly Fauna from the Upper Lower Cambrian Bastion and Ella Island Formations, North-East Greenland
[15]
논문
The Canal System in Sclerites of Lower Cambrian ''Sinosachites'' (Halkieriidae: Sachitida): Significance for the Molluscan Affinities of the Sachitids
[16]
웹사이트
"''Oikozetetes seilacheri''"
http://burgess-shale[...]
Virtual Museum of Canada
2011
[17]
논문
Enigmatic shells, possibly halkieriid, from the Middle Cambrian Burgess Shale, British Columbia
[18]
논문
"''Oikozetetes ''from the early Cambrian of South Australia: implications for halkieriid affinities and functional morphology"
[19]
논문
Enigmatic shells, possibly halkieriid, from the Middle Cambrian Burgess Shale, British Columbia
[20]
논문
"''Oikozetetes ''from the early Cambrian of South Australia: implications for halkieriid affinities and functional morphology"
[21]
논문
The Ocruranus-Eohalobia group of small shelly fossils from the Lower Cambrian of Yunnan
[22]
논문
New data on the enigmatic Ocruranus–Eohalobia group of Early Cambrian small skeletal fossils
[23]
논문
Halkieriids in Middle Cambrian Phosphatic Limestones from Australia
2004-05
[24]
문서
"The Upper Precambrian." In: "State of level of scrutiny of Precambrian and Phanerozoic stratigraphy of the Russia. The goals of the further studies." Decisions of the Interdepartmental Stratigraphical Committee and its constant Commissions
St.-Petersburg: VSEGEI
[25]
논문
Halkieriids in Middle Cambrian phosphatic limestones from Australia
[26]
논문
Halkieriids in Middle Cambrian Phosphatic Limestones from Australia
2004-05
[27]
논문
Halkieriids in Middle Cambrian Phosphatic Limestones from Australia
2004-05
[28]
간행물
The Rise and Fall of the Ediacaran Biota
http://www.paleo.pan[...]
[29]
논문
Closing the Phosphatization Window: Testing for the Influence of Taphonomic Megabias on the Pattern of Small Shelly Fossil Decline
http://www.bioone.or[...]
2008-07-30
[30]
논문
Evolution of 'small shelly fossils' assemblages of the early Paleozoic
http://www.paleo.pan[...]
2008-08-01
[31]
서적
Geological Society of America Special Publication
[32]
논문
Zooproblematica and mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China
[33]
논문
Halwaxiids and the Early Evolution of the Lophotrochozoans
http://www.sciencema[...]
2007-03
[34]
논문
A reassessment of the enigmatic Burgess Shale fossil ''Wiwaxia corrugata'' (Matthew) and its relationship to the polychaete ''Canadia spinosa''. Walcott
[35]
논문
Articulated Palaeozoic fossil with 17 plates greatly expands disparity of early chitons
[36]
논문
The brachiopod fold: a neglected body plan hypothesis
http://eprints.gla.a[...]
2008-08-07
[37]
논문
The Early Cambrian Halkieria is a mollusc
http://www.jakobvint[...]
2008-08-07
[38]
서적
Epigenetic Mechanisms of the Cambrian Explosion
https://books.google[...]
Elsevier Science
2019
[39]
논문
A soft-bodied mollusc with radula from the Middle Cambrian Burgess Shale
2006-07-13
[40]
논문
Darwin's dilemma: the realities of the Cambrian 'explosion'
2006-06
[41]
논문
Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale
http://www3.intersci[...]
2008-08-06
[42]
논문
Burgess Shale-type preservation of both non-mineralizing and "shelly" Cambrian organisms from the Mackenzie Mountains, northwestern Canada
[43]
논문
New Well-preserved Scleritomes of Chancelloridae from the Early Cambrian Yuanshan Formation (Chengjiang, China) and the Middle Cambrian Wheeler Shale (Utah, USA) and paleobiological implications
http://jpaleontol.ge[...]
2002-07
[44]
간행물
walcott2009
[45]
논문
Skeletal microstructure indicates Chancelloriids and Halkieriids are closely related
http://www.geol.ucsb[...]
2008-08-07
[46]
논문
The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism
[47]
서적
Evolving form and function: fossils and development
Peabody Museum of Natural History, Yale University
[48]
문서
動物界より上位の階級(ドメイン (分類学)|ドメイン:真核生物、等)は省略する。
[49]
논문
Redescription of the Lower Cambrian Halkieria obliqua Poulsen
[50]
뉴스
Articulated halkieriids from the Lower Cambrian of north Greenland
https://www.nature.c[...]
1990-06
[51]
서적
カンブリア紀の怪物たち
講談社
[52]
논문
Articulated Halkieriids from the Lower Cambrian of North Greenland and their Role in Early Protostome Evolution
[53]
논문
Halwaxiids and the Early Evolution of the Lophotrochozoans
2007-03
[54]
논문
A soft-bodied mollusc with radula from the Middle Cambrian Burgess Shale
2008-08-07
[55]
논문
Zooproblematica and mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China
http://www.springerl[...]
[56]
논문
Redescription of the Lower Cambrian ''Halkieria obliqua ''Poulsen
[57]
논문
Articulated halkieriids from the Lower Cambrian of north Greenland
1990-06
[58]
논문
Enigmatic shells, possibly halkieriid, from the Middle Cambrian Burgess Shale, British Columbia
https://semanticscho[...]
[59]
논문
Articulated Halkieriids from the Lower Cambrian of North Greenland and their Role in Early Protostome Evolution
[60]
논문
Halkieriids in Middle Cambrian phosphatic limestones from Australia
[61]
논문
Neoproterozoic- Cambrian Biological Revolutions
https://semanticscho[...]
[62]
서적
Palaeobiology II
Wiley-Blackwell
2009-11-12
[63]
논문
The Canal System in Sclerites of Lower Cambrian ''Sinosachites'' (Halkieriidae: Sachitida): Significance for the Molluscan Affinities of the Sachitids
[64]
논문
The Early Cambrian ''Halkieria'' is a mollusc7
http://www.jakobvint[...]
2008-07-31
[65]
논문
Lower Cambrian coeloscleritophorans (''Ninella, Siphogonuchites'') from Xinjiang and Shaanxi, China
[66]
논문
The cap-shaped Cambrian fossil ''Maikhanella ''and the relationship between coeloscleritophorans and molluscs
[67]
논문
Skeletal Microstructure Indicates Chancelloriids and Halkieriids Are Closely Related
https://semanticscho[...]
[68]
논문
Lower Cambrian Halkieriids and Other Coeloscleritophorans from Aksu-Wushi, Xinjiang, China
1997-01
[69]
논문
''Thambetolepis delicata'' gen. et sp. nov., an enigmatic fossil from the Early Cambrian of South Australia
[70]
논문
Small Shelly Fauna from the Upper Lower Cambrian Bastion and Ella Island Formations, North-East Greenland
[71]
종
92
[72]
저널
"''Oikozetetes ''from the early Cambrian of South Australia: implications for halkieriid affinities and functional morphology"
[73]
저널
The Ocruranus-Eohalobia group of small shelly fossils from the Lower Cambrian of Yunnan
[74]
저널
New data on the enigmatic Ocruranus–Eohalobia group of Early Cambrian small skeletal fossils
[75]
저널
Lower Cambrian halkieriids and other coeloscleritophorans from Aksu-Wushi, Xinjiang, China
[76]
간행물
"The Upper Precambrian." In: "State of level of scrutiny of Precambrian and Phanerozoic stratigraphy of the Russia. The goals of the further studies."
St.-Petersburg: VSEGEI
[77]
저널
Upper Cambrian Archaeocyatha from Antarctica
[78]
저널
A new post-early Cambrian archaeocyath from Antarctica
[79]
서적
The Rise and Fall of the Ediacaran Biota
http://www.paleo.pan[...]
[80]
저널
Closing the Phosphatization Window: Testing for the Influence of Taphonomic Megabias on the Pattern of Small Shelly Fossil Decline
http://www.bioone.or[...]
2008-07-30
[81]
저널
Evolution of 'small shelly fossils' assemblages of the early Paleozoic
http://www.paleo.pan[...]
2008-08-01
[82]
서적
Geological Society of America Special Publication
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com