2025. 2. 26. 오후 8:34:20
꼬리 없는 ‘가짜 전갈’이 나타났다...신종 전갈붙이 3종 발견
출처: 한겨레 ( 한국 / 한국어 )
거미강은 절지동물에 속하며, 머리가슴과 배로 나뉘고 다리가 8개인 특징을 가진다. 협각과 촉지를 가지고 있으며, 서폐 또는 기관을 통해 호흡한다. 대부분 육식성이지만, 잡식성 또는 균류를 먹는 경우도 있다. 거미, 전갈, 진드기 등을 포함하며, 다양한 분류군으로 나뉜다. 거미강은 고생대부터 화석으로 발견되며, 멸종된 분류군도 존재한다.
| 거미강 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Arachnida |
| 명명자 | 라마르크, 1801 |
| 영어 이름 | arachnid |
| 어원 | 그리스어 ἀράχνη (arákhnē, 거미) |
| 한국어 이름 | 거미강 거미형강 거미류 |
| 생존 시기 | 실루리아기 전기 - 현재 |
| 분류 | |
| 하위 분류 | 본문 참조 |
| 목 | 리키눌레이목 (Ricinulei) 검미목 (Xiphosura) - 투구게 각궁목 (Eurypterida) - 바다 전갈 (멸종) 가위거미목 (Opiliones) - 가위거미 기생거미목 (Solifugae) - 낙타거미 응애목 (Acariformes) - 다양한 진드기 후기문절목 (Parasitiformes) - 다양한 진드기 및 참진드기 †팔랑기오타르비다목 (Phalangiotarbida) (멸종) 꼬리좀사마귀목 (Palpigradi) - 꼬리좀사마귀 |
거미강은 곤충류, 갑각류, 다지류와는 달리 촉각이 없고 입의 구조가 다르다.[65] 몸은 전체(머리가슴)와 후체(배) 두 부분으로 나뉘며, 다리는 4쌍(8개)이다. 더듬이와 턱이 없고, 입 주변에는 협각과 촉지라는 부속지가 있다.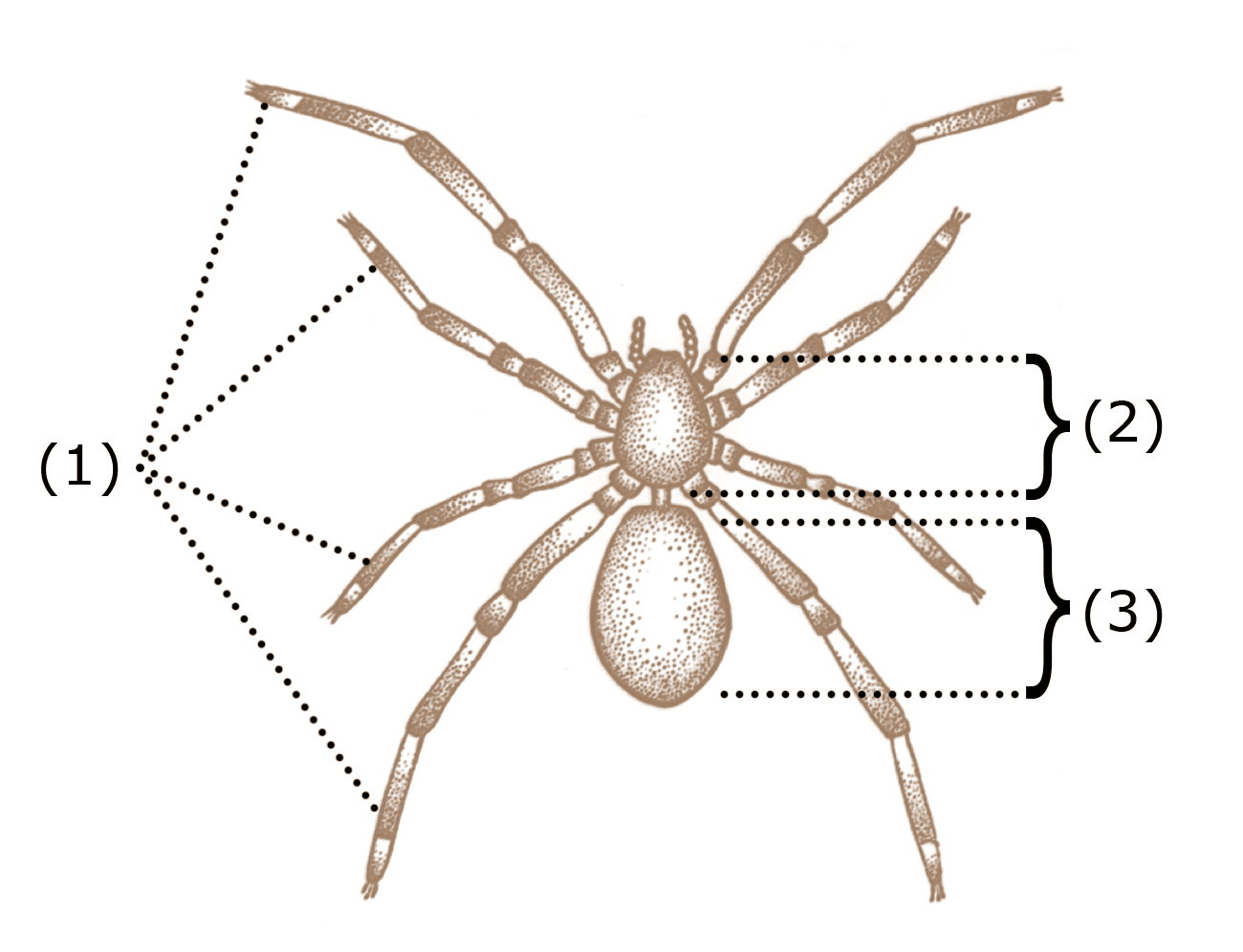
대부분의 거미강은 곤충이 6개의 다리를 가진 것과 달리 8개의 다리를 가지고 있다. 거미류는 먹이를 먹고, 자신을 방어하며, 감각을 느끼기 위해 적응된 두 쌍의 부속지를 가지고 있는데, 첫 번째 쌍인 협각은 먹이를 먹고 방어하는 데 사용된다. 다음 쌍인 촉지는 먹이를 먹거나, 이동하거나, 번식하는 기능을 한다.[6] 전갈, 유사전갈 등은 촉지가 집게발로 끝나지만, 채찍거미 등은 포식성이며 먹이를 잡는 데 사용된다.[6] 낙타거미의 촉지는 다리와 매우 유사하여 다리가 10개인 것처럼 보이기도 한다.
거미류는 촉각이나 날개가 없다는 점에서도 곤충과 구별된다. 몸은 머리가슴과 배라고도 불리는 두 개의 태그마로 구성되어 있다. 머리가슴은 보통 하나의 비분절 등갑으로 덮여 있고, 배는 더 원시적인 형태에서는 분절되어 있지만, 많은 그룹에서 분절 간에 다양한 정도의 융합이 발생한다. 전형적으로는 전배와 후배로 나뉘지만, 이는 전갈에서만 명확하게 보이며, 진드기와 같은 일부 목에서는 복부 부분이 완전히 융합된다.[11]
모든 절지동물과 마찬가지로, 거미류는 외골격을 가지고 있으며, 연골과 유사한 조직의 내부 구조인 내골격도 가지고 있다. 내골격은 일부 긴다리에서는 석회화되기도 한다.[16]
거미강은 책폐 또는 기관을 통해 호흡한다. 거미와 채찍거미는 혈림프의 압력을 이용하여 유압식으로 다리를 뻗는다.[17] 전갈과 일부 거미는 혈액에 헤모시아닌을 포함하고 있어 푸른색을 띤다. 효율적인 기관계를 가진 거미강은 혈액 내에서 산소를 운반할 필요가 없어 순환계가 축소될 수 있다.[34]
배설 기관으로는 앞가슴 옆을 따라 최대 4쌍의 콕사선과 창자를 비우는 1~2쌍의 말피기관이 있다. 많은 거미강은 이 중 한 종류의 배설선만 가지고 있지만, 일부는 두 종류 모두 가지고 있기도 하다. 거미강의 주요 질소성 폐기물은 구아닌이다.[34]
대부분의 거미강은 육식성이며, 곤충과 다른 작은 동물의 소화된 시체를 먹는다. 소화 효소를 먹이에 분비하여 액체 상태로 만들어 전구강으로 빨아들여 섭취하는 체외 소화를 한다.
거미강은 대부분 육상에서 생활하지만, 물거미, 물진드기와 같이 거미나 진드기류 중에서는 수생 종도 있다.[63] 진드기는 기생충이거나 질병을 옮기는 매개체 역할을 하기도 하고, 작은 동물, 곰팡이, 식물 즙, 썩은 물질 등을 먹기도 한다.[22] 긴다리거미는 썩어가는 식물과 동물, 배설물, 동물, 버섯을 먹는 등 잡식성이며 포식자이자 분해자이다.[23][24][25]

거미강은 다양한 목(Order)으로 분류되며, 학계에서는 거미강의 계통 발생과 분류에 대해 지속적으로 연구하고 있다. 최근 분자 계통학 연구 결과, 진드기아강은 측계통군으로 밝혀져, 응애류와 기생진드기류 두 분류군으로 나뉜다.[4]
거미류의 하위 분류는 일반적으로 목으로 취급된다. 역사적으로 진드기와 참진드기는 하나의 목, 즉 응애목으로 취급되었으나, 분자 계통학적 연구에 따르면 두 그룹은 단일 분기도를 형성하지 않으며, 형태학적 유사성은 수렴 진화의 결과이다. 따라서 현재는 두 개의 개별 분류군, 즉 응애류와 기생진드기류(목 또는 상목으로 분류)로 취급된다.
현생 거미강은 11개의 목으로 나뉘며, 멸종된 4개의 목을 포함하여 총 15개의 목이 알려져 있다. 거미강의 내부 계통에 대해서는 다양한 가설이 존재하며, 특히 분자계통분석 결과는 형태학적 분석 결과와 상충되는 경우가 많아 논란이 계속되고 있다.[56][118][122][123]
거미, 채찍거미, 채찍전갈, 분열거미 등으로 구성된 사폐류는 형태학적, 분자계통학적 분석 모두에서 안정적인 계통군으로 인정받고 있다.[56][117][118][132] 전갈은 과거 여러 계통적 위치가 제안되었으나, 최근에는 사폐류와의 유연 관계가 유력하게 지지받고 있다.[56][117][135]


거미강은 절지동물 중 협각류에 속한다. 협각류는 바다거미류, 투구게류, 그리고 거미류로 구성된다.[65][117][111] 2010년경부터 형태학적, 분자적 증거를 바탕으로 절지동물의 주요 하위 분류군 간의 계통 발생적 관계에 대한 합의가 이루어졌다.
현존하는 협각류는 바다거미와 투구게, 그리고 육상 거미류로 나뉜다. 2019년 연구에 따르면, 투구게류는 거미류 내부에 깊숙이 위치하는 것으로 나타났다. 이는 거미류의 육상화 과정이 복잡했음을 시사한다.[46][47]
거미류 내의 계통 관계는 밝혀내기 어려웠으며, 여러 연구에서 서로 다른 결과가 나왔다. 특히 응애목, 진드기목, 의벌목과 같이 진화 속도가 빠른 분류군에서 계통 발생 정보에 충돌이 있었다. 2014년 연구에서는 가장 느리게 진화하는 유전자들을 기반으로 분석하여, 거미류 내 일부 분류군의 단일 계통을 확인했다.
사폐류는 거미목, 주름거미목, 분주목을 포함하며, 이들의 계통 관계는 비교적 명확하게 밝혀졌다.[56][117][118][132] 전갈목은 유전자 중복[138][139], 서폐 구조[140] 등 여러 증거를 통해 사폐류와의 유연관계가 확인되어, 폐거미류를 구성한다.[56][117][135]
투구게류와 바다전갈류는 과거 절구류라는 분류군으로 묶였으나, 20세기 후반 이후 바다전갈류는 거미류와 단계통군을 이루는 것으로 보는 견해가 주류를 이루었다.[143][144][134][145][146][117]
협각류의 공통 조상은 수생이었을 것으로 추정되며, 거미류의 육상 생활은 이후 진화 과정에서 획득된 형질로 보인다.[75][120][123]

거미류는 음식물로 이용되는 종류가 거의 없기 때문에 인간과 별로 관계가 없다고 생각되기 쉽다. 그러나 기생성 진드기류나 독샘을 가지고 있어 가끔 인간에게 피해를 주는 전갈이나 거미류는 인간과 직접적인 관계가 있다.[8] 농작물의 해충이나 병원체를 매개하는 진드기류나, 반대로 농작물의 해충을 잡아먹거나 그것에 기생하는 거미류·진드기류 등은 인간과 깊은 관계를 가지고 있다.[8]
[1]
논문
A plurality of morphological characters need not equate with phylogenetic accuracy: A rare genomic change refutes the placement of Solifugae and Pseudoscorpiones in Haplocnemata
https://onlinelibrar[...]
2024-07-01
[2]
서적
Assembling the Tree of Life
https://archive.org/[...]
Oxford University Press
2004-01-01
[3]
서적
Foraging-Inspired Optimisation Algorithms
Springer International Publishing
[4]
논문
Grand challenges in research on arachnid diversity, conservation, and biogeography
2023-01-01
[5]
백과사전
Arachnid
1989-01-01
[6]
서적
Invertebrate Zoology: A Tree of Life Approach
https://books.google[...]
CRC Press
2021-07-08
[7]
서적
Giftige und gefährliche Spinnentiere
Westarp Wissenschaften
1993-01-01
[8]
논문
Morphological support for a clade comprising two vermiform mite lineages: Eriophyoidea (Acariformes) and Nematalycidae (Acariformes)
[9]
서적
Fundamentals of Applied Acarology
https://books.google[...]
Springer
2016-12-14
[10]
서적
The Tarantula Keeper's Guide
Barron's
2009-01-01
[11]
서적
Invertebrate Zoology: A Functional Evolutionary Approach
https://archive.org/[...]
Thomson Learning
2007-01-01
[12]
서적
The Colonisation of Land: Origins and Adaptations of Terrestrial Animals
https://books.google[...]
Cambridge University Press
1983-12-15
[13]
서적
Harvestmen: The Biology of Opiliones
https://books.google[...]
Harvard University Press
2007-02-28
[14]
논문
Homeosis in a scorpion supports a telopodal origin of pectines and components of the book lungs
[15]
논문
Fossil evidence for the origin of spider spinnerets
https://www.nature.c[...]
[16]
논문
Natural calcification of the prosomatic endosternite in the Phalangiidae (Arachnida:Opiliones)
1978-01-01
[17]
논문
Mechanics of Cuticular Elastic Energy Storage in Leg Joints Lacking Extensor Muscles in Arachnids
2003-02-15
[18]
논문
Evolution of locomotion in arachnida: The hydraulic pressure pump of the giant whipscorpion, ''Mastigoproctus giganteus'' (Uropygi)
2005-02-06
[19]
논문
Muscle Firing Patterns in Two Arachnids Using Different Methods of Propulsive Leg Extension
http://jeb.biologist[...]
1992-01-01
[20]
논문
Elastic energy storage in the pedipedal joints of scorpions and sun-spiders (Arachnida, Scorpiones, Solifugae)
https://www.biodiver[...]
2004-01-01
[21]
논문
Early Terrestrial Animals, Evolution, and Uncertainty
2011-09-01
[22]
논문
Feeding habits and multifunctional classification of soil-associated consumers from protists to vertebrates
[23]
논문
Diet, predators, and defensive behaviors of New Zealand harvestmen (Opiliones: Neopilionidae)
[24]
웹사이트
Common harvestman
https://www.wildlife[...]
[25]
웹사이트
How do harvestmen hunt?
https://www.discover[...]
[26]
서적
Harvestmen: the Biology of Opiliones
Harvard University Press
2007-01-01
[27]
웹사이트
Rare Vegetarian Spider Discovered
https://www.livescie[...]
[28]
논문
Transcriptomic Analysis of Pseudoscorpion Venom Reveals a Unique Cocktail Dominated by Enzymes and Protease Inhibitors
[29]
논문
From father to son: Transgenerational effect of tetracycline on sperm viability
[30]
논문
The Enzymatic Core of Scorpion Venoms
[31]
논문
Characterisation of protein families in spider digestive fluids and their role in extra-oral digestion
[32]
논문
Molecular basis of anticoagulant and anticomplement activity of the tick salivary protein Salp14 and its homologs
[33]
웹사이트
Tick Paralysis
https://www.ncbi.nlm[...]
[34]
서적
Invertebrate Zoology
Holt-Saunders International
1982-01-01
[35]
논문
Sexual dimorphism in the Arachnid orders
[36]
논문
Sexual dimorphism in the Arachnid orders
[37]
서적
Wilderness Medicine E-Book: Expert Consult Premium Edition - Enhanced Online Features
https://books.google[...]
Elsevier Health Sciences
2011-10-31
[38]
논문
The good fathers: efficiency of male care and the protective role of foster parents in a Neotropical arachnid
2019-04-00
[39]
논문
Mating system and exclusive postzygotic paternal care in a Neotropical harvestman (Arachnida: Opiliones)
2010-03-00
[40]
논문
A critical appraisal of the placement of Xiphosura (Chelicerata) with account of known sources of phylogenetic error
[41]
논문
The genome of a daddy-long-legs (Opiliones) illuminates the evolution of arachnid appendages
2021-00-00
[42]
논문
The house spider genome reveals an ancient whole-genome duplication during arachnid evolution
2017-07-00
[43]
논문
Spectacular alterations in the female reproductive system during the ovarian cycle and adaptations for matrotrophy in chernetid pseudoscorpions (Pseudoscorpiones: Chernetidae)
[44]
논문
Taxonomic sampling and rare genomic changes overcome long-branch attraction in the phylogenetic placement of Pseudoscorpions
https://academic.oup[...]
2021-06-00
[45]
논문
Chromosome-level assembly of the horseshoe crab genome provides insights into its genome evolution
2020-00-00
[46]
논문
Ordered phylogenomic subsampling enables diagnosis of systematic errors in the placement of the enigmatic arachnid order Palpigradi
2019-12-18
[47]
논문
Comprehensive species sampling and sophisticated algorithmic approaches refute the monophyly of Arachnida
https://academic.oup[...]
2022-02-03
[48]
논문
Cretaceous arachnid ''Chimerarachne yingi'' gen. et sp. nov. illuminates spider origins
https://www.research[...]
[49]
논문
The phylogeny of fossil whip spiders
[50]
논문
Almost a spider: A 305 million-year-old fossil arachnid and spider origins
[51]
논문
Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders
[52]
논문
A phylogenetic analysis of the arachnid orders based on morphological characters
[53]
논문
Fossil evidence for the origin of spider spinnerets, and a proposed arachnid order
2008-00-00
[54]
뉴스
'Extraordinary' fossil sheds light on origins of spiders
https://www.bbc.co.u[...]
2018-06-09
[55]
문서
クモガタ類の画像
[56]
간행물
なぜダニ類はクモガタ類の中で最も種数が多いのか?
[57]
서적
動物分類表
丸善
[58]
서적
第1章 系統と分類
東京大学出版会
[59]
서적
動物分類名
丸善
[60]
서적
クモ形類(クモ綱)総論
平凡社
[61]
서적
岩波 生物学辞典 第5版
岩波書店
[62]
문서
内田監修 (1966)
[63]
서적
クモガタ類
平凡社
[64]
문서
クモ類の呼称
[65]
논문
Segmentation and tagmosis in Chelicerata
https://www.academia[...]
[66]
문서
クモガタ類の分類
[67]
문서
クモガタ類の分類
[68]
논문
Expression of homeobox genes shows chelicerate arthropods retain their deutocerebral segment
https://www.ncbi.nlm[...]
1998-09-01
[69]
논문
Larval development and morphogenesis of the sea spider Pycnogonum litorale (Ström, 1762) and the tagmosis of the body of Pantopoda
http://www.sciencedi[...]
2003-12-01
[70]
논문
Arthropod Segmentation and Tagmosis
http://link.springer[...]
Springer Berlin Heidelberg
2022-04-25
[71]
문서
コヨリムシ・ヒヨケムシ・一部の胸板ダニ類・ザトウムシ(痕跡的)・ヤイトムシ
[72]
문서
あり:コヨリムシ・ムカシザトウムシ・コスリイムシ・ウデムシ・サソリモドキ・ヤイトムシ、なし:サソリ、ワレイタムシ、クモ
[73]
문서
ヒヨケムシ、カニムシ、ザトウムシ、クツコムシ
[74]
문서
触肢や脚の内突起は「下顎」や「顎葉」といい、クモの鋏角は「上顎」とも呼ばれるが、どれも大顎類の顎(大顎・小顎)とは別器官である。
[75]
논문
The evolution of feeding within Euchelicerata: data from the fossil groups Eurypterida and Trigonotarbida illustrate possible evolutionary pathways
https://peerj.com/ar[...]
2020-08-13
[76]
문서
ダニ・コヨリムシ・ザトウムシ・クツコムシ・ヒヨケムシ・カニムシ・サソリ
[77]
문서
ワレイタムシ・ウララネイダ類・クモ・コスリイムシ・ウデムシ・サソリモドキ・ヤイトムシ
[78]
문서
ヒヨケムシ・一部のクモ(ジグモ類・アゴダチグモ・アシナガグモ類の雄・アリグモの雄など)・一部のザトウムシ(サスマタアゴザトウムシなど)
[79]
논문
Cheliceral morphology in Solifugae (Arachnida) : primary homology, terminology, and character survey. (Bulletin of the American Museum of Natural History, no. 394)
https://hdl.handle.n[...]
2015-06-22
[80]
웹사이트
クモとは
https://kotobank.jp/[...]
2022-06-23
[81]
논문
岡山産オウギツチカニムシ ''Allochthonius(Allochthonius)opticus'' の再記載 (カニムシ目,ツチカニムシ科)
https://www.nat.muse[...]
[82]
문서
クツコムシ・カニムシ・サソリ・一部のワレイタムシ(パレオカリヌスなど)・サソリモドキ
[83]
문서
一部のザトウムシ(アカザトウムシなど)・ウデムシ・ヤイトムシ
[84]
문서
6節:一部のカニムシの脚(蹠節を欠く)、8節:ケダニの脚・クツコムシとヒヨケムシの第3-4脚(基部が1節多い)。
[85]
논문
Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications
https://doi.org/10.1[...]
1989-09-01
[86]
문서
ウデムシ・サソリモドキ・ヤイトムシ・ヒヨケムシ(全て第1脚)
[87]
문서
フシダニなど
[88]
논문
Miniaturisation in Chelicerata
https://linkinghub.e[...]
2019-01
[89]
문서
あり:サソリ・ザトウムシ・カニムシ、なし:クモ・ウデムシ・サソリモドキ・ヤイトムシ・ヒヨケムシ・クツコムシ・ダニ・コヨリムシ
[90]
논문
Mechanics of cuticular elastic energy storage in leg joints lacking extensor muscles in arachnids
https://doi.org/10.1[...]
2003-02-15
[91]
문서
ウデムシ・サソリモドキ・ヤイトムシ:第2-4脚3節、コスリイムシ:番目により4-6節、ヒヨケムシ:種類や番目により1-7節、クツコムシ:番目により4-5節、ザトウムシ:種類により1-70節以上、コヨリムシ:第4脚2節
[92]
서적
V. The Arachnida
https://www.degruyte[...]
Cornell University Press
2019-05-20
[93]
논문
The first Mesozoic Solifugae (Arachnida), from the Cretaceous of Brazil, and a redescription of the Palaeozoic solifuge
https://www.palass.o[...]
1996
[94]
문서
サソリの触肢・カニムシの触肢・一部のヒヨケムシの第1脚
[95]
문서
サソリ・ザトウムシ
[96]
논문
The epistomo-labral plate and lateral lips in solifuges, pseudoscorpions and mites
https://www.research[...]
2000
[97]
문서
ヒヨケムシ、カニムシ、アシナガダニ
[98]
논문
Comparative anatomy of the rostrosoma of Solifugae, Pseudoscorpiones and Acari
https://doi.org/10.1[...]
2022-03-01
[99]
웹사이트
The jumping spider mouth ( Araneae : Salticidae ) 1
https://www.semantic[...]
2011
[100]
논문
Lateral eye evolution in the arachnids
https://bioone.org/j[...]
2016-07
[101]
논문
REINTERPRETATION OF THE SILURIAN SCORPION PROSCORPIUS OSBORNI (WHITFIELD): INTEGRATING DATA FROM PALAEOZOIC AND RECENT SCORPIONS: REINTERPRETATION OF PROSCORPIUS
https://doi.org/10.1[...]
2008-03
[102]
논문
A new stem-group Palaeozoic harvestman revealed through integration of phylogenetics and development
https://scholar.harv[...]
2014
[103]
논문
Redescription of the sole species of the enigmatic solifuge genus ''Dinorhax'' Simon, 1879 (Solifugae: Melanoblossiidae) in Southeast Asia
http://www.bioone.or[...]
2018-11
[104]
문서
ほとんどのクモ(後疣類)・アシナガダニ以外のダニ
[105]
문서
コヨリムシ・クツコムシ・ワレイタムシ・ウララネイダ類・基盤的な化石クモ(キメララクネなど)・ウデムシ・サソリモドキ・ヤイトムシ
[106]
문서
コヨリムシ・ウララネイダ類・基盤的なクモ(キメララクネなど)・サソリモドキ・ヤイトムシ
[107]
문서
ダニ・ザトウムシ・クツコムシ・カニムシ
[108]
논문
Muscular anatomy of the giant whipscorpion Mastigoproctus giganteus (Lucas) (Arachnida: Uropygi) and its evolutionary significance
https://academic.oup[...]
1993-08-01
[109]
문서
書肺:サソリ、ワレイタムシ、クモ(クモ下目を除く)、ウデムシ、サソリモドキ、ヤイトムシ
気管系:カニムシ、ザトウムシ、クツコムシ、ヒヨケムシ、ダニ(一部を除く)
[110]
간행물
ダニとその生物学
https://doi.org/10.1[...]
日本農芸化学会
1968
[111]
논문
The Phylogeny and Evolutionary History of Arthropods
https://www.cell.com[...]
2019-06-17
[112]
논문
Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders
https://peerj.com/ar[...]
2014-11-13
[113]
논문
Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata
https://onlinelibrar[...]
1979
[114]
문서
「クモ亜綱」とも言い、コヨリムシ目・サソリ目・クモ目・ウデムシ目・サソリモドキ目・ヤイトムシ目からなる。
[115]
문서
「ダニ亜綱」とも言い、クツコムシ目・ダニ類・ザトウムシ目・カニムシ目・ヒヨケムシ目からなる。
[116]
문서
サソリ・ザトウムシ・ヒヨケムシ・カニムシからなる。
[117]
웹사이트
Current views on chelicerate phylogeny—A tribute to Peter Weygoldt
https://www.semantic[...]
2018-11-17
[118]
논문
Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal
https://academic.oup[...]
2014-08-08
[119]
논문
A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error
https://academic.oup[...]
2019
[120]
논문
Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion
https://www.cell.com[...]
2020-09-10
[121]
논문
What Is an “Arachnid”? Consensus, Consilience, and Confirmation Bias in the Phylogenetics of Chelicerata
https://www.mdpi.com[...]
2021-11
[122]
논문
Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida
https://www.nature.c[...]
2019-05-24
[123]
논문
Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata
http://www.sciencedi[...]
2020-11-01
[124]
논문
Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data
https://doi.org/10.1[...]
2002-02
[125]
논문
Evidence for Multiple Reversals of Asymmetric Mutational Constraints during the Evolution of the Mitochondrial Genome of Metazoa, and Consequences for Phylogenetic Inferences
https://academic.oup[...]
2005-04-01
[126]
논문
Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida
https://doi.org/10.1[...]
2022-02-01
[127]
논문
Major Revisions in Arthropod Phylogeny Through Improved Supermatrix, With Support for Two Possible Waves of Land Invasion by Chelicerates
https://www.research[...]
2020-01
[128]
문서
背甲の縁辺部折り返し構造/心域/肋部の欠如・前胃の退化/欠如・顎基の退化/欠如・可動な内葉の欠如・第7体節付属肢の欠如・陸上呼吸
[129]
문서
体外消化(ザトウムシと一部の胸穴ダニ類は体内消化)、マルピーギ管(コヨリムシ、カニムシ、ザトウムシと一部の胸板ダニ類はもたない)、5対以下の単眼に退化した側眼(それ以上に発達した側眼は化石種のサソリとワレイタムシに見られる)、スリット状感覚器(コヨリムシはもたない)
[130]
문서
前方/前腹側に向く口・背甲の縁辺部折り返し構造/心域/肋部の欠如
[131]
문서
背甲に覆われない第5付属肢の最初の肢節は第2肢節/転節(ヒヨケムシ、コヨリムシと胸板ダニ類に共通されるが、これらの群の該当体節はそもそも背甲に覆われていない)・脛節由来の apotele 屈筋(カブトガニ類とウミグモ類の諸説な肢節解釈により変わる)・第8体節の単一の生殖孔(胚発生ではカブトガニ類と同じく対になる)・後体第1節付属肢の欠如(ウミグモの第4脚は)・第後体第7節付属肢の欠如(サソリの該当体節には付属肢由来の書肺がある)
[132]
서적
動物学の百科事典
丸善出版
[133]
논문
The origins of tetrapulmonate book lungs and their significance for chelicerate phylogeny
https://www.academia[...]
1997-01-01
[134]
서적
The early history and phylogeny of the chelicerates
https://doi.org/10.1[...]
Springer Netherlands
2022-04-25
[135]
논문
Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks
https://doi.org/10.1[...]
2019-03-01
[136]
논문
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences
https://www.nature.c[...]
2010-02
[137]
논문
Taxonomic Sampling and Rare Genomic Changes Overcome Long-Branch Attraction in the Phylogenetic Placement of Pseudoscorpions
https://doi.org/10.1[...]
2021-06-01
[138]
논문
Homeobox Gene Duplication and Divergence in Arachnids
https://pubmed.ncbi.[...]
2018-09-01
[139]
논문
New genomic resources and toolkits for developmental study of whip spiders (Amblypygi) provide insights into arachnid genome evolution and antenniform leg patterning
https://www.biorxiv.[...]
2020-07-03
[140]
논문
The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): evidence for homology and a single terrestrialisation event of a common arachnid ancestor
https://pubmed.ncbi.[...]
2006
[141]
문서
[142]
논문
Sperm carriers in Silurian sea scorpions
https://doi.org/10.1[...]
2011-09-03
[143]
논문
Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata
https://onlinelibrar[...]
1979
[144]
논문
EVOLUTIONARY MORPHOLOGY AND PHYLOGENY OF ARAGHNIDA
https://onlinelibrar[...]
1990-03
[145]
논문
A phylogenetic analysis of the arachnid orders based on morphological characters
https://doi.org/10.1[...]
2007-06-01
[146]
논문
Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura
https://doi.org/10.1[...]
2013-01-01
[147]
논문
An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China
https://www.academia[...]
2015-11
[148]
논문
A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids
https://www.research[...]
2015-09-21
[149]
문서
[150]
문서
[151]
서적
16 Phylogeny of the Chelicerates: Morphological and molecular evidence
https://www.research[...]
De Gruyter
2014-01-29
[152]
문서
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com