검미목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
검미목은 전절과 제1-6 체절이 융합된 합체절을 특징으로 하는 절지동물로, 1쌍의 겹눈, 1쌍의 단안, 그리고 섭식용 협각과 다리를 가지고 있다. 몸은 전체와 후체로 나뉘며, 후체에는 6쌍의 덮개판과 새엽이 존재한다. 다른 협각류와 달리 고체 음식을 섭취하며, 갯벌이나 해저에 서식하는 육식성 잡식성 생물이다. 현존하는 투구게류는 4종이 있으며, 멸종위기종으로 분류되어 보호받고 있다. 검미목은 진협각류의 초기 계통 발생 연구에 중요한 분류군으로 여겨지며, 분류학적으로 투구게아목으로 분류된다.
더 읽어볼만한 페이지
- 검미목 - 투구게
투구게는 절지동물문 협각아문에 속하는 해양 절지동물로, 말굽 모양의 등딱지와 긴 꼬리침이 특징이며, 혈액 채취와 서식지 파괴로 인해 멸종 위기에 놓여 있고 일부 지역에서는 문화적 중요성을 지닌다. - 검미목 - 대서양투구게
대서양투구게는 말굽 모양의 등딱지를 가진 종으로, 시각 생리학 연구와 세균성 내독소 검출에 활용되며, 국제 자연 보전 연맹에 의해 취약종으로 분류된다. - 피에르 앙드레 라트레유가 명명한 분류군 - 가재하목
가재하목은 십각목에 속하는 갑각류로, 앞 세 쌍의 보각에 집게발이 있으며 전 세계 바다와 민물에 분포하고, 가재상과, 남방가재상과, 붉은바다가재상과, 집게새우상과 등의 상과를 포함하며 일부 종은 식용으로 이용되고 한국에는 여러 담수 및 해산 가재류가 서식한다. - 피에르 앙드레 라트레유가 명명한 분류군 - 잎말이나방과
잎말이나방과는 작은 크기의 나방 무리로, 유충은 잎을 말거나 갉아먹으며 서식하고, 여름과일잎말이나방, 사과심식나방 등 농업 해충을 포함하여 천적이나 친환경 방제로 피해를 줄일 수 있다.
| 검미목 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| |
| 학명 | Xiphosura |
| 명명자 | 라트레이유, 1802 |
| 어원 | 그리스어 'ξίφος'(xiphos, 칼) + 'οὐρά'(oura, 꼬리) |
| 분류 | |
| 그룹 | †루나타스피스 †말디불라키아? †윌베라티아 †카시벨리누리다에 Xiphosurida †벨리누리다에 †팔레오리물리다에 †오스트로리물리다에 리물리다에 |
| 생물학적 분류 | |
| 계 | 동물계 Animalia |
| 문 | 절지동물문 Arthropoda |
| 아문 | 협각아문 Chelicerata |
| 아문 계급 없음 | 진협각류 Euchelicerata |
| 강 | 퇴구강 Merostomata |
| 목 | 카부토가니모쿠 (검미목) Xiphosura |
| 목 명명자 | Latreille, 1802 |
| 일본어 이름 | 카부토가니 (카부토가니루이), 겐비루이 |
| 영어 이름 | Horseshoe crab, King crab, Xiphosuran |
| 이명 | Merostomata Dana, 1852 |
| 하위 분류 | |
| 아목 | 하라후시카부토가니아모쿠 (쿄켄비아모쿠) "Synziphosurina" † (측계통군) 카부토가니아모쿠 Xiphosurida |
| 생존 시기 | |
| 시기 | 오르도비스기 – 현세 |
| 기간 | 약 4억 8천만 년 전 ~ 현재 |
2. 명칭
학명 '''Xiphosura'''는 그리스어 'Xiphos'(검)와 'uros'(꼬리)의 합성어로, 검과 같은 미절(꼬리 부분)에서 유래하여 이름 붙여졌다[30]. 한국어로는 흔히 투구를 닮은 등껍질 모양 때문에 '투구게'라고 불리며, 학명의 의미를 따른 '검미류'(劍尾類)라는 명칭도 사용된다.
검미목 동물은 협각류에 속하며, 이름과 달리 게나 다른 갑각류와는 유전적으로 거리가 멀다. 하지만 여러 언어에서 '게'(crab, 蟹 등)라는 이름이 포함된 호칭으로 불리는 경우가 많다.
- 일본어로는 '''카부토가니'''(カブトガニ)라고 부르며, 이는 일본에 서식하는 유일한 종인 ''Tachypleus tridentatus''의 표준 일본어 이름이기도 하다[31][32]. 한자로는 '''鱟''' 또는 '''兜蟹'''로 표기한다. 별명으로 '''검미류'''(剣尾類, 켄비루이)라고도 한다[33].
- 영어에서는 말굽 모양의 등갑 때문에 '''horseshoe crab'''이라고 부르는 것이 가장 일반적이다. 그 외에 '''king crab'''[30]이라는 이름도 있으며, 학명 Xiphosura에서 유래한 학술적 총칭으로 '''xiphosuran'''이라는 표현도 사용된다[97].
- 중국어에서는 '''鱟'''(간체자: 鲎, 병음: hòu, 鲎|호우zho)라고 하며, '''馬蹄蟹'''(馬蹄蟹|마티시에zho, 영어 'horseshoe crab'과 같은 의미)나 '''王蟹'''(王蟹|왕시에zho, 영어 'king crab'과 같은 의미)라고도 부른다. 암수가 짝지어 다니는 번식 행동 때문에 '''부처어'''(夫妻鱼|푸치위zho)나 '''원앙어'''(鴛鴦魚|위안양위zho)라는 별칭도 있다[34].
3. 형태
검미목의 몸은 크게 앞부분인 전체(prosoma)와 뒷부분인 후체(opisthosoma) 두 부분으로 나뉘며, 몸 끝에는 검처럼 생긴 뾰족한 미절(telson)이 달려 있다. 몸 전체는 단단한 갑각으로 덮여 있으며, 모든 다리와 같은 부속지는 몸의 아랫면(복면)에 위치한다. 현생종은 성체의 몸길이가 최대 60cm에 달하는 비교적 큰 절지동물이지만, 고생대에 살았던 화석종 중에는 몸길이가 1cm에서 3cm에 불과한 작은 종들도 많았다. 현존하는 종들은 겉모습이 서로 매우 비슷하지만[64], 멸종한 화석종, 특히 중생대의 종들은 형태가 비교적 다양했다.[97] 외골격은 질긴 큐티클로 이루어져 있으나, 단단한 결정성 생체 광물은 포함하지 않는다.[10] 전갈처럼 검미목의 외골격도 자외선을 비추면 형광을 내는 투명한 층을 가지고 있다.[11]
전체의 등면에는 반원형의 등갑이 덮고 있으며, 여기에는 최대 4개의 눈이 있다. 양 측면에는 한 쌍의 겹눈이 있고, 앞쪽 중앙에는 한두 개의 홑눈이 있다. 겹눈은 다른 절지동물에 비해 구조가 단순하여 움직임은 감지할 수 있지만 명확한 상을 맺지는 못하는 것으로 보인다. 홑눈 앞에는 화학물질을 감지하는 화학수용체로 기능하는 것으로 추정되는 기관이 있다.[9]
몸 아랫면에는 한 쌍의 집게 모양 협각과 다섯 쌍의 다리가 있다. 입은 다리 기저부 사이의 중앙에 위치한다.[8][9] 처음 네 쌍의 다리에는 악기부(gnathobase)라는 가시 구조가 있어 먹이를 잘게 부수는 데 사용된다. 마지막 다섯 번째 다리 쌍에는 집게나 가시 대신 아가미를 청소하거나 땅을 팔 때 흙을 밀어내는 구조가 있다. 다리 뒤쪽에는 기문(chilaria)이라는 작은 부속지가 있는데, 이는 후체의 첫 번째 체절이 퇴화하여 남은 흔적으로 여겨진다.[12][9]
후체는 현생종에서는 하나의 판으로 융합되어 있지만[13], 기본적으로는 부속지가 있는 앞부분(중체부)과 부속지가 없는 뒷부분(후체부)으로 나뉜다. 아랫면에는 생식과 관련된 구멍과 호흡을 담당하는 다섯 쌍의 판 모양 아가미(서새)가 있다.[9]
몸 끝의 긴 가시 모양 미절은 매우 유연하게 움직이며, 몸이 뒤집혔을 때 스스로 자세를 바로잡는 데 사용된다.[9]
3. 1. 전체
현생 검미목은 성체의 길이가 최대 60cm에 달하지만, 고생대 종은 종종 훨씬 작았으며, 어떤 종은 길이가 1cm에서 3cm에 불과했다.몸은 앞쪽의 전체(prosoma)와 뒤쪽의 후체(opisthosoma) 두 부분으로 구성되며, 말단에는 검과 유사한 미절(telson)을 가진다. 전체의 윗면은 반원형의 등갑으로 덮여 있으며, 아랫면에는 다섯 쌍의 다리와 한 쌍의 집게발 모양의 협각(chelicera)이 있다. 입은 전체의 중앙 아랫면에 위치하며, 다리 기저부 사이에 있으며, 입술이라고 불리는 입술 모양 구조 뒤에 있다.[8][9] 모든 부속지 (관절지)는 복부에 있으며, 폭넓은 갑각 형태의 몸으로 덮여 있다. 현존종은 모두 수십 cm에 달하는 대형 절지동물이지만, 멸종된 화석종까지 범위를 넓히면 수 cm 정도의 소형종이 더 많다. 현존종의 외관은 서로 매우 유사하지만[64], 화석종, 특히 중생대의 것은 비교적 다양했다.[97] 외골격은 질긴 큐티클로 구성되지만 결정성 생체 광물은 포함하지 않는다.[10] 전갈과 마찬가지로, 검미목은 자외선 형광을 나타내는 투명한 외피층을 가지고 있다.[11]
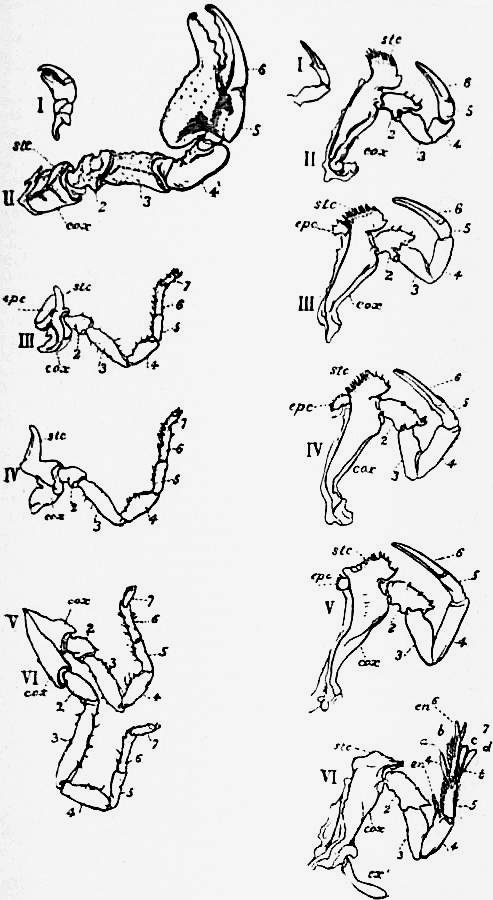
(I: 협각, II: 촉지, III-VI: 다리)
전체(Prosoma)전체(prosoma)는 전절과 제1-6 체절의 융합으로 만들어진 합체절이다.[97][42] 등쪽의 외골격인 등갑(carapace)은 넓은 반원형에서 초승달 형태로 발달하여 모든 복면 구조를 덮는 돔 모양의 갑각이 된다.[97] 등면의 좌우와 중앙에는 원칙적으로 각 1쌍의 겹눈(측안, lateral eye)과 단안(중안, median ocelli)이 있다. 현존종의 경우, 겹눈 바로 뒤와 복측의 입 앞 두 곳에서 각각 1쌍의 특수한 눈인 유충안(larval eye)이 확인되며, 복측의 것은 복안(ventral eye)이라고도 불린다.[35][36] 미국검미는 3개의 중안(일반적인 1쌍과 그 약간 뒤 중앙에 1개)을 가진다. 겹눈은 다른 절지동물의 눈보다 구조가 간단하며, 개별 겹눈이 조밀한 패턴으로 배열되어 있지 않다. 움직임을 감지할 수 있지만, 실제 이미지를 형성할 수는 없을 것으로 보인다.[9] 단안 앞에는 화학수용체 기능을 하는 것으로 보이는 추가 기관이 있다.[9] 할라후시검미류의 경우, 대부분의 종류는 눈이 퇴화하여 소실된 것으로 생각된다.[44]
검미류의 등갑은 눈 외에도 다양한 특징을 가진다. 겹눈 위쪽에는 안부 융기선(opthalmic ridge)[35]이 뻗어 있고, 후연 중앙에서 심엽(cardiac lobe)[35]이라는 솟아오른 영역이 앞으로 뻗어 있다. 양쪽 후측에서 튀어나온 부분은 후각(posterior angle)[35], 뺨가시(genal spine)[37], 또는 측극(lateral spine)[37]이라고 한다. 복면은 가장자리를 따라 접어 올린 외골격 구조(doublure)가 있으며, 이것과 등갑의 경계, 즉 등갑의 가장자리는 탈피 시 갈라지는 틈이기도 하다. 이러한 특징은 분류군에 따라 형태와 발달 정도가 다르며, 검미류의 계통 관계를 나타내는 데 중요하게 여겨지는 형질이다.[97][38]
전체 제1-6 체절에서 유래한 6쌍의 부속지는 모두 복면 중앙 부근에서 만나며, 입은 그 사이에 뒤쪽을 향해 열려 있다. 입과 입술 앞에 있는 제1쌍 부속지는 섭식용의 짧은 가위형 협각(chelicera)이다.[42][39] 다음 5쌍은 다리로, 6절 이상으로 나뉘며[40][41][46][39], 선단 2절이 가위를 이루는 경우가 많다. 그중 첫 번째 1쌍은 촉지(pedipalp)이지만, 거미류만큼 특화되지 않아 다른 다리와의 구분이 거의 없다.[41][39] 그러나 검미과의 수컷은 이 부속지 1쌍 또는 다음 다리와 함께 2쌍이 암컷을 붙잡기 위해 선단이 굵은 갈고리 모양의 구조(파지기, clasper)[35]로 특화되어 있다.[39]
다리의 처음 네 쌍은 집게발로 끝나고, 안쪽 면에 악기(gnathobase)라고 불리는 일련의 가시가 있다. 이 가시는 음식을 씹는 데 사용되어 입으로 전달되기 전에 찢는다.[12] 그러나 다섯 번째이자 마지막 다리 쌍에는 집게발이나 가시가 없으며, 대신 아가미를 청소하고 굴을 파는 동안 진흙을 치우는 구조가 있다. 현존종의 경우, 이 최종 다리 쌍은 가장 발달하여, 첫 번째 지절(기절, coxa)은 "노 모양 기관"(flabellum)이라는 잎 모양의 분기(외지 exopod 또는 외엽 exite로 여겨짐)를 상부에 가지며, 최종 지절의 뿌리 주위에 "주걱 모양 기관"(spatulate organ)[35]이라는 4장의 평평한 구조체를 가지고 있다.[42][39] 각각의 다리 기절에는 입을 둘러싼 악기(gnathobase)라는 섭식용 톱니 모양의 돌기(내엽, 내돌기)가 있다.[39] 다리 사이와 입 바로 뒤에는 "endostoma"라는, 복판(sternite)에서 유래한 것으로 보이는 눈에 띄지 않는 외골격이 1장 있다.[43][42] 다리 뒤에는 여섯 번째 부속지인 순상지(chilarium)가 있는데, 크기가 매우 작고 털과 가시로 덮여 있다.[12] 이들은 흡수된 첫 번째 후체절의 사지 잔재로 여겨진다.[9][97][42][39]
거미류와 마찬가지로, 검미류의 전체도 편의상 "두흉부"(cephalothorax)라고 불리는 경우가 있지만, 합체절적으로는 머리와 가슴/몸통의 융합으로 만들어진 두흉부가 아니라, 머리에만 해당하는 머리 융합절이다.[97] 다만 검미목의 경우, 등갑은 전체뿐만 아니라 (다른 절지동물의 몸통에 해당하는) 후체의 일부까지 포함하기 때문에, 등갑 부분이라면 두흉부라고도 할 수 있다.[43][97][42]
후체(Opisthosoma)후체(opisthosoma)는 편의상 "복부(abdomen)"라고도 하지만, 합체절적으로는 몸통에 해당한다.[97] 종류에 따라 9~11절(제7~15/17체절)을 포함하며, 많은 경우 전체보다 폭이 좁다. 후체부는 앞쪽의 중체부(mesosoma)와 편평한 부속지로 나뉘며, 뒤쪽에는 부속지가 없는 후체부(metasoma)가 있다. 현대 형태에서는 후체부 전체가 하나의 분절되지 않은 구조로 융합되어 있다.[13] 검미목의 경우, 후체 전방의 체절은 복잡한 유리와 융합이 진행되어 제1절(제7체절)과 제2절(제8체절)의 중앙까지 전체와 융합하여 등갑으로 덮여 있다. 유리된 후체 제2절의 가장자리는 "free lobe"라는 돌출부가 되어 후체 제3절(제9체절) 이후의 모든 체절과 융합하여 "thoracetron"이라는 1장의 갑각을 이룬다.[43][97] 다만 기반적인 화석군, 이른바 키불리류(Synziphosurina)의 후체는 모든 체절이 가동하는 등판(tergite)으로 나뉜다.[42][44] 등판은 삼엽형, 즉 중앙 1쌍의 종구로 정중선의 축부(axial region)와 좌우로 돌출된 늑부(tergopleura)로 나뉜다.[97] 다만 많은 검미목 종류는 그것이 불분명하며, 주로 한 쌍을 이루는 함몰부(내돌기 apodeme)나 가시(epimera)에 의해 나타난다.[97] 검미목 중에서 thoracetron의 양쪽에 후체 제3-8절(제9-14체절) 유래[43]의 가장자리 가시(marginal spine)[35]라고 불리는 6쌍의 가동 가시(movable spine)를 epimera 사이에 갖는 예가 많다.[94]
검미류의 후체 제1절(제7체절)은 대부분 외형상으로 인정할 수 없을 정도로 퇴화되어 있으며, 현존종의 경우 이 체절은 앞서 언급했듯이 전체의 등갑과 융합하고, 복면은 순상지(chilarium)라는, 최종 다리 사이에 배치되는 짧은 부속지를 갖는다.[97][42][39] 그러나 대부분의 화석종의 제7체절/부속지에 대한 자세한 정보는 불분명하며, 일부 키불리류에서는 독립적인 퇴화성 등판(microtergite)에 의해 나타난다.[97] 그중에서도 웨인벨기나(Weinbergina)는 이 부분이 전체와 거의 동일한 형태의 다리를 가졌을 것으로 생각된다.[45][46][42] 이러한 성질로 인해, 이 체절은 기능적으로 오히려 전체적이며, 전체의 일부로 다루어야 한다는 의견도 검토된다.[42]
다음 후체 제2-7절(제8-13체절)은 6쌍의 덮개판(operculum)이라는 평평한 부속지를 가지며, 기본적으로 폭이 넓은 외지와 손가락 모양의 짧은 내지(endopod)로 이루어진 이지형이다.[97][42] 양 부속지의 기부 중앙에는 "sternal lobe"라는, 복판에서 유래된 것으로 보이는 작고 뾰족한 구조물이 있다.[39] 첫 번째 1쌍은 좌우가 융합된 생식구개(genital operculum)라고 하며, 난자나 정자를 배출하는 1쌍의 생식공(gonopore)이 그 뒤쪽의 기부에 있다.[97][39] 다음 5쌍은 새엽(branchial appendage 또는 gill operculum)이며, 책의 페이지를 연상시키는 호흡용 서새(book gill)는 각 외지의 뒤쪽에서 매달려 있다.[97][39] 이러한 덮개판은 평소에는 뒤쪽으로 접히기 때문에, 새엽은 생식구개에 의해 덮여 보호된다.[39] 후체부의 아랫면에는 생식 구멍과 다섯 쌍의 판 모양 아가미(서새)가 있다.[9]
미절(Telson)후체의 말단에 해당하는 항문 바로 뒤에는, 1개의 가시 모양의 미절(telson)이 뻗어 있으며, 이것은 "caudal spine"이라고도 불린다. 단면은 삼각형(삼각기둥상) 혹은 원형(원주상)이며, 뿌리의 관절을 통해 상하와 좌우로 움직일 수 있다. 검미목의 미절은 종종 전체 길이의 절반 이상을 차지하지만, 키불리류의 미절은 그것에 비해 약간 짧다.[42] 이 척추는 매우 움직이며, 우연히 뒤집혔을 경우 동물을 똑바로 세우는 데 사용된다.[9]
3. 2. 후체
몸은 앞쪽의 전체부와 뒤쪽의 후체부(後體部, opisthosoma) 또는 복부로 나뉜다. 후체부는 다시 앞쪽의 중체부(mesosoma)와 뒤쪽의 후체부(metasoma)로 구분될 수 있으며, 중체부에는 편평한 부속지들이 있고 후체부에는 부속지가 없다. 현생 검미목에서는 후체부 전체가 하나의 분절되지 않은 구조로 융합되어 있다.[13]후체부의 아랫면에는 생식 구멍과 다섯 쌍의 판 모양 아가미가 있다.[9] 이 아가미는 서새(書鰓, book gill)라고도 불리며 호흡에 사용된다. 아가미는 덮개판(Operculum)이라는 판 모양 부속지 아래에 보호된다. 첫 번째 덮개판 쌍은 생식구개(Genital operculum)라고 하며, 생식 구멍과 관련이 있다.
다리 뒤에는 여섯 번째 부속지로 간주되는 기문(Chilaria)이 있는데, 크기가 매우 작고 털과 가시로 덮여 있다.[12] 이들은 흡수된 첫 번째 후체절의 사지 잔재로 여겨진다.[9]
후체부는 일반적으로 미절(Telson) 또는 미부(尾部)라고 불리는 길고 뾰족한 꼬리 가시로 끝난다. 이 꼬리 가시는 매우 움직이며, 동물이 우연히 뒤집혔을 경우 몸을 바로 세우는 데 사용된다.[9]
3. 3. 내부 구조
입은 단단해진 식도로 연결되며, 식도는 모이주머니와 모래주머니로 이어진다. 모래주머니에서 먹이를 잘게 부순 후, 소화할 수 없는 부분은 다시 입으로 뱉어내고 나머지는 위로 보낸다. 위에서는 소화 효소를 분비하며, 몸의 대부분을 차지하는 두 개의 큰 맹장이 위에 연결되어 있어 음식물로부터 영양소를 흡수한다. 소화관의 끝은 창자를 거쳐 단단해진 직장으로 이어지며, 직장은 꼬리 가시 바로 앞쪽의 항문으로 열린다.[9]검미류는 잘 발달된 순환계를 가지고 있다. 긴 관 모양의 심장은 여러 동맥을 통해 혈액을 몸 전체 조직으로 보낸다. 아가미 옆에는 두 개의 세로 방향 정맥동이 있다. 아가미에서 산소를 공급받은 혈액은 체강으로 흘러 들어가 다시 심장으로 돌아온다. 혈액에는 척추동물의 헤모글로빈과 유사한 기능을 하는 파란색의 구리 기반 색소인 헤모시아닌이 포함되어 있으며, 혈액 응고를 돕는 혈구도 들어 있다.[9]
배설계는 두 쌍의 체절샘으로 구성되며, 이는 방광에 연결되어 마지막 걷는 다리 밑부분 근처에서 몸 밖으로 열린다. 뇌는 비교적 크며, 다른 많은 절지동물처럼 식도를 둘러싸고 있다. 암수 모두 창자 옆에 하나의 생식선을 가지고 있으며, 이는 후체부 아래쪽에서 열린다.[9]
4. 생태
현존하는 말발굽게 종류는 갯벌이나 해저에 서식하는 저서생물인 바다 동물이다. 육식에 가까운 잡식성으로, 확보한 먹이는 다리 밑부분에 있는 턱기부의 움직임으로 으깨고[54], 협각을 이용해 입으로 가져간다[39]. 이매패류를 주식으로 하며[55], 고둥류, 다모류, 갑각류, 썩은 고기, 해조류 등 다양한 생물을 먹이로 삼는다[55][56]. 뒷다리의 주걱 모양 부분과 넓은 등껍질을 이용해 갯벌이나 모래사장의 진흙에 빠지지 않고 나아갈 수 있으며, 이는 갯벌 이동이나 모래 등 퇴적물 속에 숨기에 적합하다[57]. 몸 뒷부분의 아가미덮개를 움직여 뒤집힌 상태로 헤엄치기도 한다.
다 자란 개체의 성적 이형성은 뚜렷하다. 수컷은 암컷보다 약간 작으며, 짝짓기 시 암컷의 몸 뒷부분을 잡기 쉽도록 등껍질 앞쪽 가장자리 아랫부분이 약간 위로 파여 있다. 번식기가 되면 얕은 물가의 갯벌이나 모래사장에 모인다. 수컷은 갈고리 모양으로 변형된 첫 번째 다리를 이용해 암컷의 등에 올라타 몸 뒷부분을 잡는다. 암컷은 모래에 구멍을 파고 수백에서 수만 개의 알을 낳고, 수컷이 그 위에 정자를 뿌려 체외수정을 한다. 이후 암컷은 알을 모래로 덮는다.
알의 크기는 약 2mm에서 3mm이다. 알 속에서 배아는 4번 탈피한 후 유생으로 부화한다. 이 유생은 겉모습이 삼엽충과 비슷해 '삼엽충 유충'이라고도 불린다. 이 시기에는 아직 꼬리 가시(미부)가 없으며, 유생은 알 속의 영양분(난황)으로 살아가고(먹이를 먹지 않음) 물에 떠다니다가 바닥에 자리 잡고 탈피하면 처음으로 꼬리 가시가 생긴다. 여러 차례 탈피를 거치면서 유생은 아가미를 더 만들고 꼬리 가시를 길게 늘리며 점차 다 자란 모습으로 변해간다. 현존하는 투구게류는 성적으로 성숙하기까지 약 3년이 걸린다.
수명이 길고 다 자라면 탈피를 멈추기 때문에, 몸에는 삿갓조개나 고둥류 같은 부착생물이 많이 붙어 있는 경우가 많다[59].
멸종한 화석 종들은 현생종보다 더 다양했다. 민물에 살거나, 헤엄치거나 퇴적물 속에 숨는 대신 몸을 둥글게 마는 등 현생종과는 다른 생태나 행동을 했던 것으로 보이는 종류도 알려져 있다[60][45][57][94]. 특히 석탄기에 번성했던 벨리누루스과(''Bellinuridae'')는 모든 종류가 민물에 살았던 것으로 추정된다[44].
5. 현존종
현존하는 말굽게류는 말굽게(''Tachypleus tridentatus''), 남방말굽게(''Tachypleus gigas''), 둥근꼬리말굽게(''Carcinoscorpius rotundicauda''), 미국말굽게(''Limulus polyphemus'')의 4종만 알려져 있다.

어느 종이나 생태와 외모가 매우 비슷하며[64], 서식지와 몸길이의 차이점 외에는 다음과 같은 특징으로 구별할 수 있다.[26][64][65][66][67]
| 특징 | 말굽게 Tachypleus tridentatus | 남방말굽게 Tachypleus gigas | 둥근꼬리말굽게 Carcinoscorpius rotundicauda | 미국말굽게 Limulus polyphemus |
|---|---|---|---|---|
| 수컷의 등딱지 앞 가장자리 좌우의 움푹 들어간 곳 | 있음 | 없음 | 없음 | 없음 |
| 수컷의 갈고리발톱 모양의 다리 | 앞의 2쌍 | 앞의 2쌍 | 앞의 2쌍 | 앞의 1쌍만 |
| 생식구개내지 | 퇴화 | 퇴화 | 퇴화 | 발달 |
| 후체 앞 3쌍의 가동 가시 | 긺 | 긺 | 다소 짧음 | 긺 |
| 후체 말단 중앙의 가시 수(부동 가시) | 3개 | 1개 | 1개 | 1개 |
| 미절의 형태 | 삼각기둥 모양 | 삼각기둥 모양 | 원기둥 모양 | 삼각기둥 모양 |
| 미절의 길이 | 몸과 거의 일치 | 몸과 거의 일치 | 몸보다 긺 | 몸보다 짧음 |
| 염색체 수 (2n) | 26 | 28 | 32 | 52 |
| (기타 특유 형질) | 후체 배면의 미세한 가시가 많음 | 항문 주변의 가장자리에 미세 가시가 없음 | 수컷의 갈고리발톱은 가위 모양 | 몸은 전체적으로 약간 세로로 긺 |
각 종류의 자세한 내용은 각 해당 항목을 참조하라.
5. 1. 말굽게
투구게속( ''Tachypleus'' )에는 투구게가 속한다.5. 2. 남방말굽게
남아시아와 동남아시아의 해역에 서식한다. 몸길이는 수컷 25cm에서 40cm, 암컷 25cm에서 50cm 정도이다.5. 3. 둥근꼬리말굽게
둥근꼬리말굽게는 투구게과 (Limulidae)의 맹그로브투구게속 (''Carcinoscorpius'')에 속하는 종이다.5. 4. 미국말굽게
대서양투구게는 투구게과(Limulidae)의 대서양투구게속(''Limulus'')에 속한다.6. 분류

현존하는 검미목은 모두 투구게과(Limulidae)에 속하며, 맹그로브투구게속( ''Carcinoscorpius''), 대서양투구게속( ''Limulus'', 대서양투구게 포함), 투구게속( ''Tachypleus'', 투구게 포함)의 3개 속으로 분류된다.
가장 오래된 줄기 검미목으로 알려진 ''Lunataspis''는 약 4억 4천 5백만 년 전 오르도비스기 후기의 캐나다 지역 퇴적암에서 발견되었다.[16] 이후 실루리아기 동안에는 검미목 화석이 발견되지 않다가 데본기 후기에 다시 나타났다. 석탄기에는 Belinuridae과에 속하는 담수 검미목이 크게 번성했으며, 현생 투구게과(Limulidae)의 가장 오래된 구성원도 이 시기에 출현했을 가능성이 제기된다. 투구게과는 트라이아스기에 들어서면서 풍부하게 나타나기 시작했다. 또 다른 담수 검미목 그룹인 Austrolimulidae는 페름기와 트라이아스기에 번성했다.[17] 그러나 검미목은 역사적으로 종 다양성이 그리 높지 않았으며, 가장 다양했던 석탄기에도 알려진 화석 종은 50종 미만이다.[18] 현생 투구게류의 마지막 공통 조상은 분자 시계 연대 측정 결과 쥐라기-백악기 경계 시기에 존재했을 것으로 추정되지만,[19] 계통 발생 연구에 따라서는 화석 기록을 근거로 트라이아스기까지 거슬러 올라갈 수도 있다.[22]
상세한 분류 체계는 아래 하위 분류 섹션에서 다룬다. 검미목의 속 목록 문서에서도 관련 정보를 찾아볼 수 있다.
6. 1. 분류사
19세기에는 대략적인 형태의 유사성을 바탕으로 절지동물 중에서 투구게류가 특히 삼엽충과 가까운 관계에 있다고 생각되었다. 당시에는 바다전갈류와 함께 모두 갑각류로 취급되었다.[68][69] 그러나 20세기 이후 이루어진 재검토를 통해, 투구게류와 바다전갈류는 갑각류의 특징과는 거리가 멀고, 오히려 전갈과 같은 거미류와 공통적인 기본 신체 구조를 가지고 있다는 점이 밝혀졌다.[70][71] 그 결과, 삼엽충 및 갑각류와는 구별되어 투구게류와 바다전갈류는 바다거미류, 거미류 등과 함께 절지동물의 주요 그룹 중 하나인 '''협각류(Chelicerata)'''에 포함되게 되었다.[70]6. 2. 계통 관계
검미목(말발굽게류)은 겹눈, 악기(입 주변의 다리 기저부), 이지형 부속지, 발달한 후체 부속지와 서새(책 모양 아가미) 등 현생 거미강에서는 보이지 않는 진협각류의 조상 형질로 보이는 특징을 다수 가지고 있다.[41][36][39] 이 때문에 검미목은 진협각류의 초기 계통 발생과 공통 조상을 이해하는 데 중요한 정보를 제공하는 분류군으로 여겨진다.[41][36][39]일반적으로 검미목은 거미강보다 더 이른 시기에 분기한 기저 진협각류, 즉 진협각류 진화의 초기 단계에 위치하는 그룹으로 간주된다.[97] 이 두 그룹은 적어도 고생대 오르도비스기 무렵에 공통 조상으로부터 갈라졌을 것으로 추정된다.[72][82][81]
그러나 일부 분자 계통 분석 결과는 다른 가능성을 제시하기도 한다. 이 분석들에 따르면, 검미목은 단계통군이 아닌 거미강 내에서 분기했을 수 있으며, 특히 자라벌레류[73][78][74] 또는 서폐류(서폐(책허파)를 가진 거미강 그룹으로, 거미, 전갈, 우데무시, 사소리모도키 등을 포함)와 가까운 관계일 수 있다는 결과가 나왔다.[75] 이 가설에 기반하여, 검미목이 원래 육상 생활을 하던 거미강 조상으로부터 유래하여 이차적으로 물 속 생활에 적응했다거나[78][75], 또는 검미목은 계속 물에 살았고 거미강이 여러 차례 독립적으로 육상 생활에 적응했다는[78][79] 해석이 제기되었다. 만약 검미목이 서폐류와 실제로 가깝다면, 두 그룹에서 발견되는 게놈 중복 현상은 공통 조상으로부터 물려받은 공유 파생 형질일 수 있다.[75] 그렇지 않다면 이는 수렴 진화의 결과로 볼 수 있다.[77]
하지만 이러한 견해, 특히 검미목이 이차적으로 수생화했다는 주장에 대해서는 많은 반론이 제기되었다. 통설(검미목이 거미강보다 먼저 분기했다는 견해)을 지지하는 다른 분자 계통 분석 결과[80][81]와 분자 시계 분석[82]이 존재한다. 또한 고생물학적 증거들은 검미목의 이차적 수생화설을 지지하지 않는다.[79][81] 검미목(말발굽게류)이 가진 수생 적응 특징(악기, 서새 등)들은 다른 여러 화석 협각류에서도 널리 발견되므로, 이는 이차적으로 획득한 형질이라기보다는 진협각류의 원시적인 조상 형질을 그대로 유지한 결과일 가능성이 높다.[79][81] 설령 거미강이 검미목에 대해 단계통군이 아니라고 하더라도, 최대 절약법 원칙에 따르면 거미강이 여러 번 독립적으로 육상화했다는 설명이 더 설득력 있으며, 검미목의 수생적 특징은 조상으로부터 물려받은 원시적인 상태로 보는 것이 타당하다.[79][81]
6. 2. 1. 화석 협각류와의 관계성
현존하는 협각류 중에서는 검미목(투구게류)과 직접 비교할 만한 가까운 분류군은 없다. 하지만 멸종한 화석 협각류까지 범위를 넓히면, 바다전갈류, 카스마타스피스류, 그리고 넓은 의미의 검미목에 포함되었던 할라후시카부토가니류 등 다양한 그룹이 알려져 있으며, 이들과 검미목의 관계에 대한 많은 논의가 이루어져 왔다.과거에는 검미목과 이들 화석 그룹들이 여러 특징을 공유한다는 점에서 하나의 분류군으로 묶였다. 주요 공통 특징으로는 물에 서식하는 점(수생성), 잘 발달한 등갑, 서새(책 모양 아가미), 몸 뒷부분(후체)에 달린 부속지, 겹눈, 그리고 먹이를 씹는 데 사용되는 턱기저가 달린 다리 등이 있다.[83] 이러한 공통점을 바탕으로 이들은 거미류와 대비되는 절구류(Merostomata, 다리입류라고도 함[84])라는 분류군으로 함께 묶였다.[68][85][86][74]
그러나 20세기 후반부터 절구류가 실제로 하나의 단계통군(하나의 공통 조상에서 유래한 모든 후손을 포함하는 그룹)인지에 대한 의문이 제기되기 시작했다. 현재 학계의 주류 견해는 절구류 전체가 거미류에 대해 측계통군(공통 조상은 공유하지만 그 후손 일부가 제외된 그룹)이라는 것이다. 즉, 과거 절구류의 공통 특징으로 여겨졌던 형질들은 사실 진협각류 전체의 조상 형질(조상으로부터 물려받은 원시적인 특징)이라는 해석이다.[87][97][92][93]
이러한 견해에 따라, '절구류'라는 명칭은 계통학적 실체라기보다는 단순히 편의상 사용되는 용어가 되었다. 때로는 검미목 이외의 그룹을 제외하고 절구류(Merostomata)를 검미목(Xiphosura)의 동의어로 사용하기도 한다.[97] 또한, 검미목을 기존의 절지동물문 협각아문 절구강의 목에서 독립적인 강으로 승격시키는 분류 체계도 받아들여지고 있다.[97][27] 소수의 연구에서는 여전히 절구류의 단계통성을 지지하기도 하지만, 이 경우 절구류는 매우 기초적인 협각류 그룹으로 간주되거나[86], 오히려 거미류가 절구류 내의 여러 그룹에서 독립적으로 진화한 다계통군으로 해석된다.[74]
; 바다전갈류 (광익류, Eurypterida)
바다전갈류는 과거 검미목과 매우 가까운 관계로 여겨져 함께 절구류를 구성하는 대표적인 두 그룹으로 알려졌다.[86] 하지만 20세기 후반 이후, 특히 정자낭(정자 주머니) 전달에 적합한 생식기 구조 등을 근거로, 바다전갈류는 검미목보다는 거미류와 더 가까운 관계라는 견해가 유력해졌다. 이들은 거미류와 함께 'Sclerophorata'라는 그룹을 형성하는 것으로 본다.[88][89][87][97][42]
; 카스마타스피스류 (Chasmataspidida)
카스마타스피스류는 발견된 종에 따라 검미목 또는 바다전갈류와 매우 유사한 모습을 보인다. 과거에는 독특한 형태를 가진 검미목의 한 종류로 여겨지기도 했다.[90] 그러나 1990년대 후반 이후로는 검미목과는 별개의 독립적인 그룹으로 분류된다.[97] 한때 카스마타스피스류 자체가 다계통군일 수 있다는 가설도 제기되었다. 즉, 검미목과 유사한 카스마타스피스과는 검미목에 가깝고, 바다전갈류와 유사한 디플로아스피스과는 바다전갈류에 가깝다는 설이었다.[91] 하지만 이 가설은 확실한 증거가 부족하며, 특히 2010년대 이후의 연구들은 카스마타스피스류가 단계통군이며, 바다전갈류나 거미류와 더 가깝다는 설(이들과 함께 'Dekatriata' 그룹을 형성)을 지지하는 경향이 강하다.[97][92][93][94]
; 할라후시카부토가니류 (공검미류, Synziphosurina)
할라후시카부토가니류는 전통적으로 검미목에 포함되어 왔다.[127] 이들은 현생 투구게를 포함하는 더 진화된 검미목 그룹(카부토가니아목, Xiphosurida)의 자매군(가장 가까운 친척 그룹)[127]이거나, 카부토가니아목으로 이어지는 진화 과정상의 측계통군[96]으로 여겨졌다. 그러나 2013년 Lamsdell의 연구를 시작으로 할라후시카부토가니류의 특징들을 면밀히 재검토하고 다른 협각류와의 계통 관계를 재구성한 결과, 이들은 카부토가니아목과 단계통군을 이루지 않는다는 주장이 제기되었다. 오히려 할라후시카부토가니류는 진협각류 전체의 초기 단계나, 카스마타스피스류, 바다전갈류, 거미류를 포함하는 'Dekatriata' 그룹의 초기 단계에서 각각 갈라져 나온 다양한 그룹들의 집합체(즉, 측계통군 또는 다계통군)일 가능성이 시사되었다(주석[102][113][110] 참조).[97][92][93][94][44]
이러한 연구 결과에 따라, 과거 넓은 의미로 사용되던 '검미목'은 실제로는 'Prosomapoda'(오파콜루스와 다이바스테리움 제외 진협각류)에서 'Dekatriata' 그룹을 제외한 측계통군으로 해석된다. 따라서 현재 '협의의 검미목'은 할라후시카부토가니류를 제외하고, 카부토가니아목과 이들과 가까운 몇몇 속(예: 루나타스피스, ''Kasibelinurus'')만을 포함하는 단계통군으로 재정의되었다.[97] Lamsdell (2013)[97] 및 이를 따른 후속 연구들[92][93][94]에 기반한 협각류의 계통 관계를 해석하면, 진협각류는 초기 그룹인 오파콜루스와 다이바스테리움 등이 분기한 후 'Prosomapoda' 그룹으로 이어진다. 이 Prosomapoda 내에서 다양한 할라후시카부토가니류 그룹들이 먼저 갈라져 나왔고(측계통군), 이후 현재의 '협의의 검미목'(루나타스피스 등 일부 화석 속 + 카부토가니아목)과 'Planaterga' 그룹이 분기했다. Planaterga 그룹 내에서도 일부 할라후시카부토가니류가 분기한 후, 최종적으로 카스마타스피스류, 바다전갈류, 거미류 등을 포함하는 'Dekatriata' 그룹이 분기한 것으로 보인다. 즉, 과거 '넓은 의미의 검미목'(할라후시카부토가니류 포함)은 실제로는 Dekatriata 그룹을 제외한 진협각류의 초기 분기 그룹들을 포괄하는 측계통군에 해당한다.
6. 3. 하위 분류

검미목은 크게 광의의 검미목과 협의의 검미목으로 나눌 수 있다. 2020년 기준, 현존하는 4종과 화석 100종 이상의 광의의 검미목이 기록되어 있으며, 이 중 화석종 80종 이상이 협의의 검미목에 속한다.[98] 광의의 검미목은 측계통군으로 여겨진다.[97][93] 일부 기반 진협각류나 할아버지말발굽게류로 불리는 그룹들도 편의상 광의의 검미목에 포함되기도 한다.[92][93][114]
협의의 검미목은 거의 모든 종류가 검미아목(Xiphosurida)으로 분류되며, 이는 다시 2개의 하목(벨리누리나 Belinurina, 림룰리나 Limulina), 2개의 상과(고생검미상과 Paleolimuloidea, 검미상과 Limuloidea), 그리고 5개의 과로 세분된다.[98] 검미아목 외에 카시벨리누리과(Kasibelinuridae)가 있지만, 단계통군이 아니라는 지적이 있어 분류 체계에서 제외하는 경우도 있다.[94][114][44]
현생종은 모두 투구게과(Limulidae)에 속하며, 형태적 차이에 따라 2아과(대서양투구게아과 Limulinae, 투구게아과 Tachypleinae), 3속(''Limulus'', ''Tachypleus'', ''Carcinoscorpius''), 4종으로 구분된다.[26] 인공 교배 실험 결과, 대서양투구게( ''Limulus polyphemus'')와 아시아산 투구게류 사이에는 수정이 이루어지지 않았으나, 아시아산 3종 사이에서는 교잡 및 부화 사례가 확인되었다.[99] 아시아산 3종은 맹그로브투구게( ''Carcinoscorpius rotundicauda'', 맹그로브투구게속)와 나머지 2종(투구게속)으로 나뉜다. 투구게( ''Tachypleus tridentatus'')와 비교했을 때, 남방투구게( ''Tachypleus gigas'')보다 맹그로브투구게의 생식적 격리가 덜 진행되었을 가능성이 제기되어, 맹그로브투구게를 투구게속에 포함시키자는 주장도 있다.[100] 그러나 2012년 발표된 분자계통분석 결과는 투구게와 남방투구게가 하나의 계통군을 이루고 맹그로브투구게가 이들의 자매군이라는 결과를 보여주었다.[101]
아래는 검미목의 분류 체계 목록이다 (Lamsdell 2020[27] 및 Dunlop et al. 2020[98] 기준). 멸종한 분류군은 † 기호로 표시한다.
- '''목 검미목 (Xiphosura)'''
- * †''말디불라키아''(''Maldybulakia'') (데본기) (=''Lophodesmus''[106][97])
- * †''빌베라티아''(''Willwerathia'') (데본기)
- * †''Anderella'' [103](석탄기)
- * †''Borchgrevinkium'' [104](데본기)
- * †''Camanchia'' (실루르기)
- * †''Legrandella'' (데본기)
- * †베누스툴루스속 ''Venustulus'' [107][108](실루르기)
- * †'''웨인베르기나과 (Weinberginidae)''' (데본기)
- ** †웨인베르기나속 ''Weinbergina'' [45][46](데본기)
- * (Planaterga 그룹, Dekatriata 제외)[110][97]
- ** †''Bembicosoma'' (실루르기)
- ** †''Bunaia'' (실루르기)
- ** †'''프세우도니스쿠스과 (Pseudoniscidae)''' (실루르기)
†''Cyamocephalus'' [111][112](실루르기)
†''Pseudoniscus'' (=''Neolimulus'') (실루르기)
- ** †'''부노데스과 (Bunodidae)''' (실루르기)
†''Bunodes'' (=''Exapinurus'') (실루르기)
†리물로이데스속 ''Limuloides'' (=''Hemiaspis'') (실루르기)
†''Pasternakevia'' (실루르기)
- * '''협의의 검미목 (Xiphosura ''sensu stricto'')''' (오르도비스기 - 현세)[113][97]
- ** †''Pickettia'' (데본기)
- ** †''Patesia'' (데본기)
- ** †'''카시벨리누리과 (Kasibelinuridae)''' (오르도비스기~데본기): 검미아목 제외 시 측계통군[94][98]
†''Kasibelinurus'' (데본기)
†루나타스피스속 ''Lunataspis'' (오르도비스기): 알려진 가장 오래된 협의의 검미목[28]
(상과·과 미정)
* †''Albalimulus'' (석탄기)
* †''Sloveniolimulus'' (트라이아스기)
* †''Xiphosuroides'' [117](석탄기)
'''†하목 벨리누리나 (Belinurina)'''[118] (석탄기 - 페름기)
* †'''벨리누리과 (Belinuridae)''' (=''Euproopidae'', =''Liomesaspididae'') (석탄기 - 페름기)
** †''Anacontium'' (페름기)
** †''Andersoniella'' (석탄기)
** †''Alanops'' (석탄기)
** †''Belinurus'' (=''Bellinurus'', =''Steropsis'', =''Koenigiella'', =''Macrobelinurus'', =''Parabelinurus'') [119](석탄기)
** †유프로프스속 ''Euproops'' (=''Prestwichia'', =''Prestwichianella'') (석탄기)
** †''Liomesaspis'' (=''Palatinaspis'') (석탄기 - 페름기)
** †''Patesia'' [44](석탄기) (위의 Patesia와 별개일 수 있음)
** †''Prestwichianella'' (석탄기) (Euproops의 동의어일 수 있음)
** †''Pringlia'' [120](석탄기)
** †''Prolimulus'' (석탄기)
** †''Stilpnocephalus'' [121](석탄기)
'''하목 림룰리나 (Limulina)'''[118] (석탄기 - 현세)
* †''벨리누로옵시스''(''Bellinuroopsis'') (=''Neobelinuropsis'') (데본기)[122]
* †'''롤페이과 (Rolfeiidae)''' (석탄기)
** †''Rolfeia'' (석탄기)
* '''†상과 고생검미상과 (Paleolimuloidea)''' (석탄기 - 쥐라기)
** †'''고생검미과 (Paleolimulidae)'''[123] (=''Moravuridae'') (석탄기 - 쥐라기)
*** †''Moravurus'' (석탄기)
*** †''Norilimulus'' (석탄기)
*** †팔레오리물루스속 ''Paleolimulus''[124] (석탄기 - 페름기)
*** †''Xaniopyramis'' (석탄기 - 페름기)[122]
* '''상과 검미상과 (Limuloidea)'''[26][123] (석탄기 - 현세)
** †''발로이셀라''(''Valloisella'') (석탄기)
** †'''오스트랄리물루스과 (Austrolimulidae)''' (=''Dubbolimulidae'') (석탄기 - 트라이아스기)
*** †오스트랄리물루스속 ''Austrolimulus'' [60](트라이아스기)
*** †''Batracholimulus'' (트라이아스기)
*** †''Boeotiaspis'' (석탄기)
*** †''Dubbolimulus'' [120](트라이아스기)
*** †''Limulitella'' (=''Limulites'') [120](트라이아스기 - 쥐라기)
*** †''Panduralimulus'' [120](페름기)
*** †''Psammolimulus'' (트라이아스기) (Limulidae에도 속명 존재)
*** †''Shpineviolimulus'' [114][122](페름기)
*** †''Tasmaniolimulus'' [120](트라이아스기)[125]
*** †''Vaderlimulus'' (트라이아스기)
** '''투구게과 (Limulidae)''' (트라이아스기 - 현세): 현존하는 유일한 과.
*** †''Allolimulus'' (쥐라기)
*** †''Casterolimulus'' (백악기)
*** †''Crenatolimulus'' (쥐라기 - 백악기)[126]
*** †''Heterolimulus'' (트라이아스기)
*** †메솔리물루스속 ''Mesolimulus'' (트라이아스기 - 백악기)
*** †''Keuperlimulus'' (트라이아스기)
*** †''Psammolimulus'' (트라이아스기) (Austrolimulidae에도 속명 존재)
*** †''Tarracolimulus'' (트라이아스기)
*** †''Victalimulus'' (백악기)
*** †''Volanalimulus'' (트라이아스기)
*** †''Yunnanolimulus'' (트라이아스기)
*** '''아과 대서양투구게아과 (Limulinae)'''[26] (백악기 - 현세)
대서양투구게속 ''Limulus'' (백악기 - 현세)[126]
* 대서양투구게 ''Limulus polyphemus'' (현세)
*** '''아과 투구게아과 (Tachypleinae)''' (트라이아스기 - 현세): 생식덮개(genital operculum)가 없거나 퇴화됨[64]
투구게속 ''Tachypleus'' (트라이아스기 - 현세)
* 투구게 ''Tachypleus tridentatus'' (현세)
* 남방투구게 ''Tachypleus gigas'' (현세)
맹그로브투구게속 ''Carcinoscorpius'' (현세)
* 맹그로브투구게 ''Carcinoscorpius rotundicauda'' (현세)
6. 4. 투구게류로 오인된 화석
불완전한 화석으로 인해 투구게류 또는 그 근연종으로 잘못 알려진 고생물이 몇몇 존재한다.[98] 광갑류(Aglaspidida)가 대표적인 예이다. 20세기 후반까지 광갑류는 부속지가 절구류와 비슷할 것으로 추정되어 복원되었고, 이로 인해 투구게류나 그와 가까운 협각류로 오해받았다.[127][97] 그러나 이후 부속지를 다시 검토한 결과, 협각 대신 촉각이 있는 등 협각류의 특징과는 다른 점들이 밝혀졌다.[128] 결국 광갑류는 투구게류의 근연종은 물론 협각류조차 아니며, 오히려 삼엽충을 포함하는 별개의 계통(''Artiopoda''류)에 속하는 것으로 판명되었다.[129][130]광갑류 외에도 곤충(''Hypatocephala'', ''Permolimulinella'' 등), 삼엽충(''Hemiaspis''), 극피동물(''Lemoneites'') 중에서도 투구게류로 잘못 해석된 화석들이 발견되었다.[131][98] 몸의 뒷부분 일부만 알려진 ''Kiaeria''는 오랫동안 투구게류로 여겨졌으나, 2019년 Lamsdell의 재검토를 통해 오히려 카스마타스피스류일 가능성이 제기되었다.[132] 단편적인 화석만 발견되어 과거 투구게류로 기록되었던 ''Elleria'', ''Archeolimulus''와 ''Drabovaspis''는 2020년 Lamsdell의 재검토 결과 투구게류가 아닌 것으로 밝혀졌으며, 특히 ''Elleria''는 삼엽충의 꼬리 부분 화석일 가능성이 높은 것으로 나타났다.[27]
참조
[1]
OED
Xiphosuran
[2]
학술지
Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders
[3]
서적
Arthropod Phylogeny with Special Reference to Insects
John Wiley & Sons
[4]
학술지
Revised systematics of Palaeozoic 'horseshoe crabs' and the myth of monophyletic Xiphosura
2012-12-18
[5]
학술지
A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error
2019-02-14
[6]
학술지
Ordered phylogenomic subsampling enables diagnosis of systematic errors in the placement of the enigmatic arachnid order Palpigradi
2019-12-18
[7]
학술지
Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida
https://academic.oup[...]
2022-02-03
[8]
간행물
Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA
Marine Biology
1984
[9]
서적
Invertebrate Zoology
Holt-Saunders International
[10]
URL
Crystallographic Texture of the Arthropod Cuticle Using Synchrotron Wide Angle X-ray Diffraction
http://d-nb.info/987[...]
[11]
URL
Exocuticular hyaline layer of sea scorpions and horseshoe crabs suggests cuticular fluorescence is plesiomorphic in chelicerates
https://research.amn[...]
[12]
서적
Invertebrates
Sinauer Associates
[13]
학술지
Opisthosomal fusion and phylogeny of Palaeozoic Xiphosura
[14]
URL
Developmental ecology of the American horseshoe crab Limulus polyphemus
https://pdfs.semanti[...]
[15]
URL
Metamorphosis of Limulus Polyphemus Trilobite Larvae: Role of Chemical and Structural Cues, Competency, and The Cost of Delayed Metamorphosis
https://repository.l[...]
[16]
학술지
The oldest horseshoe crab: a new xiphosurid from Late Ordovician Konservat-Lagerstätten deposits, Manitoba, Canada
[17]
학술지
Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida
2020
[18]
학술지
Chromosome-level assembly of the horseshoe crab genome provides insights into its genome evolution
2020-05-08
[19]
학술지
Critical re‐evaluation of Limulidae uncovers limited Limulus diversity
http://dx.doi.org/10[...]
2021-03-02
[20]
간행물
A summary list of fossil spiders and their relatives
http://www.wsc.nmbe.[...]
World Spider Catalog. Natural History Museum Bern
2018
[21]
학술지
Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation
[22]
학술지
The phylogeny and systematics of Xiphosura
2020-12-04
[23]
문서
1と2:[[アメリカカブトガニ]] {{snamei||Limulus polyphemus}}(成体と幼生)、3と4:[[ユウプループス]] {{snamei||Euproops}}(=''Prestwichia'')、5:[[リムロイデス]] {{snamei||Limuloides limuloides}}(=''Hemiaspis limuloides'')、6:{{snamei||Pseudoniscus aculeatus}}
[24]
웹사이트
Xiphosurida カブトガニ/剣尾目 - Biological Information System for Marine Life
https://www.godac.ja[...]
2022-04-22
[25]
웹사이트
カブトガニ類とは
https://kotobank.jp/[...]
2022-04-22
[26]
서적
カブトガニの生物学〔増補版〕
制作同人社
[27]
학술지
The phylogeny and systematics of Xiphosura
https://peerj.com/ar[...]
2020-12-04
[28]
학술지
The oldest horseshoe crab: A new xiphosurid from Late Ordovician Konservat-Lagerstatten deposits, Manitoba, Canada.
https://doi.org/10.1[...]
2008-01-17
[29]
학술지
The Fezouata fossils of Morocco; an extraordinary record of marine life in the Early Ordovician
https://jgs.lyellcol[...]
2015-09-01
[30]
웹사이트
Limulus polyphemus
http://www.sms.si.ed[...]
2018-11-17
[31]
웹사이트
カブトガニとは - コトバンク
https://kotobank.jp/[...]
2018-11-17
[32]
웹사이트
Tachypleus tridentatus カブトガニ - Biological Information System for Marine Life
https://www.godac.ja[...]
2022-04-22
[33]
웹사이트
剣尾類(けんびるい)とは
https://kotobank.jp/[...]
2020-08-17
[34]
웹사이트
夫妻魚 - 水族大點名 - 國立海洋生物博物館
http://name.nmmba.go[...]
2018-11-17
[35]
서적
カブトガニの生物学〔増補版〕
制作同人社
[36]
논문
Opsin expression in ''Limulus'' eyes: a UV opsin is expressed in each eye type and co-expressed with a visible light-sensitive opsin in ventral larval eyes
http://jeb.biologist[...]
2014-09-01
[37]
서적
図解 世界の化石大百科
河出書房新社
2000
[38]
논문
A new method for quantifying heterochrony in evolutionary lineages
https://www.cambridg[...]
[39]
논문
A 3D anatomical atlas of appendage musculature in the chelicerate arthropod ''Limulus polyphemus''
https://journals.plo[...]
2018-02-14
[40]
문서
現生種の場合、前4対の脚は6節(基節 coxa・転節 trochanter・腿節 femur・膝節 patella・tibiotarsus・apotele/dactylopodite)、最終の脚は7節(基節・転節・腿節・膝節・脛節 tibia・跗節 tarsus・apotele)、[[ウェインベルギナ]]は全ての脚が8節をもつとされる。
[41]
논문
Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications
https://doi.org/10.1[...]
1989-09-01
[42]
논문
Segmentation and tagmosis in Chelicerata
https://doi.org/10.1[...]
[43]
논문
GROSS MUSCULAR ANATOMY OF LIMULUS POLYPHEMUS (XIPHOSURA, CHELICERATA) AND ITS BEARING ON EVOLUTION IN THE ARACHNIDA
https://www.semantic[...]
2001
[44]
논문
Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida
https://www.frontier[...]
2020
[45]
논문
''Weinbergina'', a xiphosuran arthropod from the devonian hunsrück slate
http://www.museunaci[...]
1981-12-01
[46]
논문
A new specimen of ''Weinbergina opitzi'' (Chelicerata: Xiphosura) from the Lower Devonian Hunsriick Slate, Germany
https://www.academia[...]
[47]
웹사이트
http://www.horseshoecrab.org/anat/gut.html
http://www.horseshoe[...]
2018-11-18
[48]
웹사이트
Atlantic Horseshoe Crab
http://tacbio.weebly[...]
2018-11-18
[49]
논문
An “ancient” complexity? Evolutionary morphology of the circulatory system in Xiphosura
http://www.sciencedi[...]
2015-08-01
[50]
서적
The American Horseshoe Crab
https://books.google[...]
Harvard University Press
2003
[51]
간행물
Xiphosura
https://www.research[...]
Oxford University Press
2022-03-19
[52]
웹사이트
Figure 4: Nervous systems of Chelicerata.
https://www.nature.c[...]
2018-11-18
[53]
서적
The Colonisation of Land: Origins and Adaptations of Terrestrial Animals
https://books.google[...]
Cambridge University Press
1983-12-15
[54]
논문
Central pattern generation underlying Limulus rhythmic behavior patterns
https://academic.oup[...]
2010-10-01
[55]
웹사이트
(PDF) Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA
https://www.research[...]
2018-11-15
[56]
웹사이트
Horseshoe Crabs, Limulus polyphemus
http://marinebio.org[...]
2018-11-15
[57]
논문
A New Three-Dimensionally Preserved Xiphosuran Chelicerate from the Montceau-Les-Mines Lagerstätte (Carboniferous, France)
https://onlinelibrar[...]
2002
[58]
웹사이트
Horseshoe crab videos, photos and facts - Limulus polyphemus
http://www.arkive.or[...]
2018-11-16
[59]
논문
Epibiotic community of the horseshoe crab Tachypleus gigas
http://drs.nio.org/d[...]
2000
[60]
뉴스
A NEW XIPHOSURAN FROM THE TRIASSIC SEDIMENTS AT BROOKVALE, NEW SOUTH WALES.
http://australianmus[...]
Records of the Australian Museum
1955
[61]
논문
Tetrodotoxin poisoning following ingestion of the toxic eggs of the horseshoe crab Carcinoscorpius rotundicauda, a case series from 1994 through 2006
https://www.ncbi.nlm[...]
2008-03
[62]
논문
カブトガニ''Carcinoscorpius rotundicauda''のフグ毒産生菌
https://doi.org/10.2[...]
1988
[63]
웹사이트
Tetrodotoxin (TTX) Determination of Horseshoe Crab Collected from Maludam, Sarawak
https://ir.unimas.my[...]
2018-11-17
[64]
논문
A collection of horseshoe crabs (Chelicerata: Xiphosura) in the National Museum, Prague (Czech Republic) and a review of their immunological importance
https://www.research[...]
2015-06-30
[65]
웹사이트
http://www.horseshoecrab.org/nh/species.html
http://www.horseshoe[...]
2018-11-15
[66]
웹사이트
Limulidae {{!}} Horseshoe crab monitoring site
http://horseshoecrab[...]
2018-11-15
[67]
서적
The American Horseshoe Crab
https://books.google[...]
Harvard University Press
2003
[68]
서적
A monograph of the British fossil Crustacea, belonging to the order Merostomata.
https://doi.org/10.5[...]
Printed for the Palæontographical Society,
1866
[69]
논문
I.—Notes on some British Palæozoic Crustacea Belonging to the order Merostomata
https://www.cambridg[...]
1872-10
[70]
웹사이트
Die Entwicklungsgeschichte der Scolopender
https://www.schweize[...]
2022-03-20
[71]
서적
The Eurypterida of New York
https://en.wikisourc[...]
New York State Education Department,
1912
[72]
논문
The colonization of land by animals: molecular phylogeny and divergence times among arthropods
http://www.biomedcen[...]
2004
[73]
논문
Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal
https://academic.oup[...]
2014-08-08
[74]
논문
Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida
https://doi.org/10.1[...]
2022-02-01
[75]
논문
Major Revisions in Arthropod Phylogeny Through Improved Supermatrix, With Support for Two Possible Waves of Land Invasion by Chelicerates
https://www.research[...]
2020-01
[76]
논문
Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data1
https://onlinelibrar[...]
2002
[77]
논문
Taxonomic Sampling and Rare Genomic Changes Overcome Long-Branch Attraction in the Phylogenetic Placement of Pseudoscorpions
https://doi.org/10.1[...]
2021-06-01
[78]
논문
A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error
https://academic.oup[...]
2019
[79]
논문
Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion
https://www.cell.com[...]
2020-09-10
[80]
논문
Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida
https://www.nature.c[...]
2019-05-24
[81]
논문
Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata
http://www.sciencedi[...]
2020-11-01
[82]
논문
A Cambrian–Ordovician Terrestrialization of Arachnids
https://www.frontier[...]
2020
[83]
서적
The Eurypterida of New York,
https://doi.org/10.5[...]
New York State Education Department,
1912
[84]
웹사이트
腿口類とは
https://kotobank.jp/[...]
2022-04-22
[85]
서적
The eurypterid Stoermeropterus conicus from the lower Silurian Pentland Hills, Scotland
https://www.research[...]
Palaeontographical Society, London
[86]
논문
Three-dimensional reconstruction and the phylogeny of extinct chelicerate orders
https://peerj.com/ar[...]
2014-11-13
[87]
논문
A phylogenetic analysis of the arachnid orders based on morphological characters
https://doi.org/10.1[...]
2007-06-01
[88]
논문
Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata
https://onlinelibrar[...]
2009-04-27
[89]
논문
EVOLUTIONARY MORPHOLOGY AND PHYLOGENY OF ARAGHNIDA
https://onlinelibrar[...]
1990-03
[90]
서적
Chelicerate palaeobiology and evolution
http://worldcat.org/[...]
Royal Society of Edinburgh
2004
[91]
논문
A redescription of ''Chasmataspis laurencii'' Caster & Brooks (Chelicerata: Chasmataspidida) from the Middle Ordovician of Tennessee, USA, with remarks on chasmataspid phylogeny
http://www.museunaci[...]
[92]
논문
An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China
https://www.academia[...]
2015
[93]
논문
A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids
https://www.research[...]
2015-10
[94]
논문
Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation
https://www.research[...]
2016-03
[95]
웹사이트
J-GLOBAL 科学技術総合リンクセンター jglobal.jst.go.jp
https://jglobal.jst.[...]
2020-08-17
[96]
논문
Opisthosomal fusion and phylogeny of Palaeozoic Xiphosura
https://www.research[...]
1997
[97]
논문
Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura
https://academic.oup[...]
2013-01-01
[98]
문서
J. A. Dunlop, D. Penney & D. Jekel 2020. A summary list of fossil spiders and their relatives
https://wsc.nmbe.ch/[...]
[99]
문서
杉田博昭・関口晃一・宍倉文夫・山道祥郎「カブトガニの種間雑種の発生と系統」『動物分類学会誌』第22号、日本動物分類学会、1982年、1-6頁。
https://doi.org/10.1[...]
[100]
문서
関口晃一「カブトガニの適応戦略と種分化」『化石』第56巻、日本古生物学会、1994年、47-50頁。
https://doi.org/10.1[...]
[101]
문서
Matthias Obst, Søren Faurby, Somchai Bussarawit, & Peter Funch (2012). “Molecular phylogeny of extant horseshoe crabs (Xiphosura, Limulidae) indicates Paleogene diversification of Asian species”
https://doi.org/10.1[...]
[102]
문서
Sname
[103]
논문
A Carboniferous Synziphosurine (xiphosura) from the Bear Gulch Limestone, Montana, Usa
https://onlinelibrar[...]
2007
[104]
문서
Novojilov, Nestor I. (1959). "Mérostomates du Dévonien inférieur et moyen de Sibérie"
https://iris.univ-li[...]
[105]
논문
Synziphosurines (Xiphosura: Chelicerata) from the Silurian of Iowa
https://www.cambridg[...]
2011-01
[106]
문서
原記載では[[多足類]]とされ、Lamsdell (2013) では {{Snamei||Willwerathia}} との類似性に基づいてカブトガニ類として分類される。
[107]
서적
大むかしの生きもの
https://www.worldcat[...]
講談社
2020
[108]
논문
A new synziphosurine (Chelicerata : Xiphosura) from the Late Llandovery (Silurian) Waukesha Lagerstatte, Wisconsin, USA
https://www.research[...]
2005-03
[109]
논문
On the Emsian (Lower Devonian) arthropods of the Rhenish Slate Mountains: 2. The synziphosurineWillwerathia
https://www.research[...]
1998-11
[110]
문서
"{{Sname||Planaterga}}:背甲両後側の頬棘は退化的。後体の背板は発達したこぶを欠けて第3/4節で最も幅広い。第7体節の背板はmicrotergiteを形成して付属肢は退化的。"
[111]
논문
Revision of the pseudoniscine merostome genus Cyamocephalus Currie. American Museum novitates ; no. 2557
https://hdl.handle.n[...]
1974
[112]
논문
A new specimen of the Silurian synziphosurine arthropod Cyamocephalus
http://www.sciencedi[...]
1999-01-01
[113]
문서
狭義のカブトガニ類 {{Sname||Xiphosura}} sensu stricto:第7体節は退化的で付属肢は[[唇様肢]]。後体は前端で最も幅広い。心域は背甲の後半を超えるほど前へ伸びる。眼部隆起線は前方で会合してMの字形となる。
[114]
논문
Revision of ''“Bellinurus” carteri'' (Chelicerata: Xiphosura) from the Late Devonian of Pennsylvania, USA
https://www.research[...]
2019-12
[115]
논문
Patesia n. gen., a new Late Devonian stem xiphosurid genus
http://www.sciencedi[...]
2020-09-06
[116]
문서
カブトガニ亜目 {{Sname||Xiphosurida}}:第8体節(後体第2節)以降の全ての背板は癒合してthoracetronをなす。
[117]
논문
First Fossil Xiphosuran (Chelicerata, Xiphosura) Egg Clutch from the Carboniferous of Khakassia
http://link.springer[...]
2018-07
[118]
문서
巌佐庸・倉谷滋・斎藤成也・塚谷裕一編「生物分類表」『岩波 生物学辞典 第5版』岩波書店、2013年、1531-1666頁。
[119]
논문
One name to rule them all: ''Belinurus trilobitoides'' (Buckland, 1837) is senior synonym to fourteen named species
https://www.cambridg[...]
2022-01
[120]
논문
Xiphosurid from the Upper Permian of Tasmania confirms Palaeozoic origin of Austrolimulidae
https://www.research[...]
2019
[121]
논문
AN EFFACED HORSESHOE CRAB (ARTHROPODA: CHELICERATA: XIPHOSURA) FROM THE UPPER CARBONIFEROUS OF THE CARNIC ALPS (FRIULI, NE ITALY)
https://riviste.unim[...]
2019
[122]
논문
A reappraisal of Paleozoic horseshoe crabs from Russia and Ukraine
https://www.research[...]
2020-10-03
[123]
문서
小野展嗣「鋏角亜門分類表」[[石川良輔]]編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉6、岩槻邦男・馬渡峻輔監修、裳華房、2008年、410-420頁。
[124]
문서
Rodney Steel & Authony P. Harvey (ed.)「カブトガニ類 Horseshoe crabs」『古生物百科事典』小畠郁生監訳、朝倉書店、1982年、37-38頁。
[125]
논문
An earliest Triassic age for ''Tasmaniolimulus'' and comments on synchrotron tomography of Gondwanan horseshoe crabs
https://peerj.com/ar[...]
2022-04-22
[126]
논문
Critical re‐evaluation of Limulidae uncovers limited ''Limulus'' diversity
https://onlinelibrar[...]
2021-08
[127]
논문
Revision of the suborder Synziphosurina (Chelicerata, Merostomata) : with remarks on merostome phylogeny. American Museum novitates ; no. 2543
https://hdl.handle.n[...]
1974
[128]
문서
Briggs, D.E.G., Bruton, D.L., Whittington, H.B., 1979. [https://www.palass.org/publications/palaeontology-journal/archive/22/1/article_pp167-180 Appendages of the arthropod Aglaspis spinifer (Upper Cambrian, Wisconsin) and their significance]. Palaeontology 22, 167-180.
[129]
서적
Arthropods of the Lower Cambrian Chengjiang fauna, southwest China
https://foreninger.u[...]
Scandinavian University Press
1997
[130]
논문
The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda
https://doi.org/10.1[...]
2012-08-15
[131]
논문
Trilobite and xiphosuran affinities for putative aglaspidid arthropods Caryon and Drabovaspis, Upper Ordovician, Czech Republic
https://onlinelibrar[...]
2010
[132]
논문
A chasmataspidid affinity for the putative xiphosuran Kiaeria Størmer, 1934
https://doi.org/10.1[...]
2019-11-04
[133]
문서
"{{ITIS|ID=82699 | taxon = Xiphosura}}"
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com