내항동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
내항동물(Entoprocta)은 1870년에 명명된 동물로, 입과 항문이 촉수 왕관 안쪽에 위치하는 특징을 갖는다. 대부분 군체 생활을 하며, 개체는 1mm 내외의 크기를 가진다. 외형적으로 태형동물과 유사하나, 먹이 섭취 방식, 내부 해부학, 유충 변태 과정 등에서 차이를 보인다. 분류는 4개 과, 약 150여 종으로 이루어져 있으며, 대부분 해양에 서식하지만 일부는 담수에서도 발견된다. 내항동물은 자웅동체이며, 출아에 의한 무성 생식도 활발하게 이루어진다. 화석 기록은 드물며, 캄브리아기 대폭발 시기에 존재했던 것으로 추정된다. 계통적으로는 트로코조아에 속하며, 환형동물과 가까운 관계에 있다.
더 읽어볼만한 페이지
| 내항동물 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
| 학명 | Entoprocta |
| 명명자 및 연도 | Nitsche, 1870 |
| 어원 | 그리스어로 '안쪽'을 의미하는 'entos'와 '항문'을 의미하는 'proktos'의 합성어임. |
| 특징 | 촉수관 내부로 항문이 열리는 특징을 가짐. 몸 크기는 정도임. |
| 분류 | |
| 하위 분류군 | Barentsiidae (= Urnatellidae) Loxokalypodidae Loxosomatidae Pedicellinidae incertae sedis †Cotyledion |
| 생물학적 분류 | |
| 역 | 진핵생물 |
| 계 | 동물계 |
| 문 | 내항동물문 |
| 계통분류학적 위치 | |
| 상문 | 촉수담륜동물상문 |
| 화석 기록 | |
| 화석 범위 | 초기 캄브리아기 - 현재 |
2. 명칭
내항동물(Entoprocta)은 1870년에 "안쪽의 항문"을 의미하는 이름으로 명명되었다.[4] 1929년에는 "굽은" 또는 "휘어진" 동물을 의미하는 캄토조아(Kamptozoa)라는 대체 명칭이 붙여졌다.[3] 일부 저자는 "내항동물"(Entoprocta)을 사용하고,[6][10] 다른 저자는 "캄토조아"(Kamptozoa)를 선호한다.[4][22]
내항동물의 몸은 크게 몸통부와 줄기부로 나뉜다. 몸통부인 '''꽃받침'''에는 촉수관과 대부분의 내장이 있다. 줄기부는 꽃받침을 지지하는 부분으로, 막대 모양이거나 더 복잡한 형태를 띠기도 한다. 단독 생활을 하는 종은 '''족반'''을 이용해 기질에 부착하고, 군체 생활을 하는 종은 식물의 포복경(匍匐莖, 기는줄기)과 같은 '''주근'''을 뻗어 새로운 개체를 만든다.
내항동물이라는 명칭은 촉수관 안쪽에 항문이 있다는 데서 유래하며, 태형동물은 촉수관 바깥쪽에 항문이 있어 외항동물이라고 부르는 것과 대조를 이룬다.
3. 형태
꽃받침부는 윗면에 촉수관이 있으며, 대개 약간 비스듬히 기울어져 있다. 촉수는 다소 말굽 모양으로 배열되어 있으며, 개개의 촉수는 꽃받침부의 길이를 넘을 정도로 길지 않고, 그다지 유연하지도 않다. 촉수 표면은 온통 섬모로 덮여 있다. 촉수가 배열된 안쪽 영역의 한쪽 끝에는 입이, 다른 한쪽 끝에는 항문이 있다.
조건이 나빠지면 꽃받침부는 떨어져 나가기도 하지만, 조건이 좋아지면 바로 재생된다. 군체성 종에서는 주근에서 일종의 '''휴아'''(쉬는 눈)를 형성하는 것으로 알려져 있다.
3. 1. 개요
대부분의 내항동물 종은 군체 생활을 하며, 그 구성원들은 완전히 독립적인 동물이 아니기 때문에 "개체"라고 불린다.[7] 개체는 일반적으로 1mm 길이지만 0.1mm에서 7mm 길이 범위에 있다.[4]
내항동물은 겉으로는 태형동물(외항동물)과 유사하며, 두 그룹 모두 입으로 음식 입자를 끌어들이는 물의 흐름을 생성하는 섬모가 있는 촉수 "왕관"을 가지고 있다. 그러나 먹이 공급 방식과 내부 해부학이 다르며, 태형동물은 유충에서 성체로의 변태를 거치면서 유충 조직의 대부분을 파괴한다. 또한 그들의 군체는 "딸"과 다른 창시자 개체(zooid)를 가지고 있다.[4]
| 내항동물[4] | 태형동물 (외항동물)[4] | |
|---|---|---|
| 촉수 | 단단함 | 속이 빔 |
| 먹이 섭취 흐름 | 촉수 기부에서 끝 부분으로 | 촉수 끝 부분에서 기부로 |
| 항문 위치 | 촉수 "왕관" 안쪽 | 촉수 "왕관" 바깥쪽 |
| 체강 | 없음 | 세 부분으로 나뉨 |
| 군체 내 창시자 개체의 형태 | 다른 개체와 같음 | 일반적인 개체와 달리 둥긂[9] |
| 성체로의 변태 | 대부분의 유충 구조를 유지 | 대부분의 유충 구조를 파괴 |
| 배설 기관 | 전신관 | 없음 |
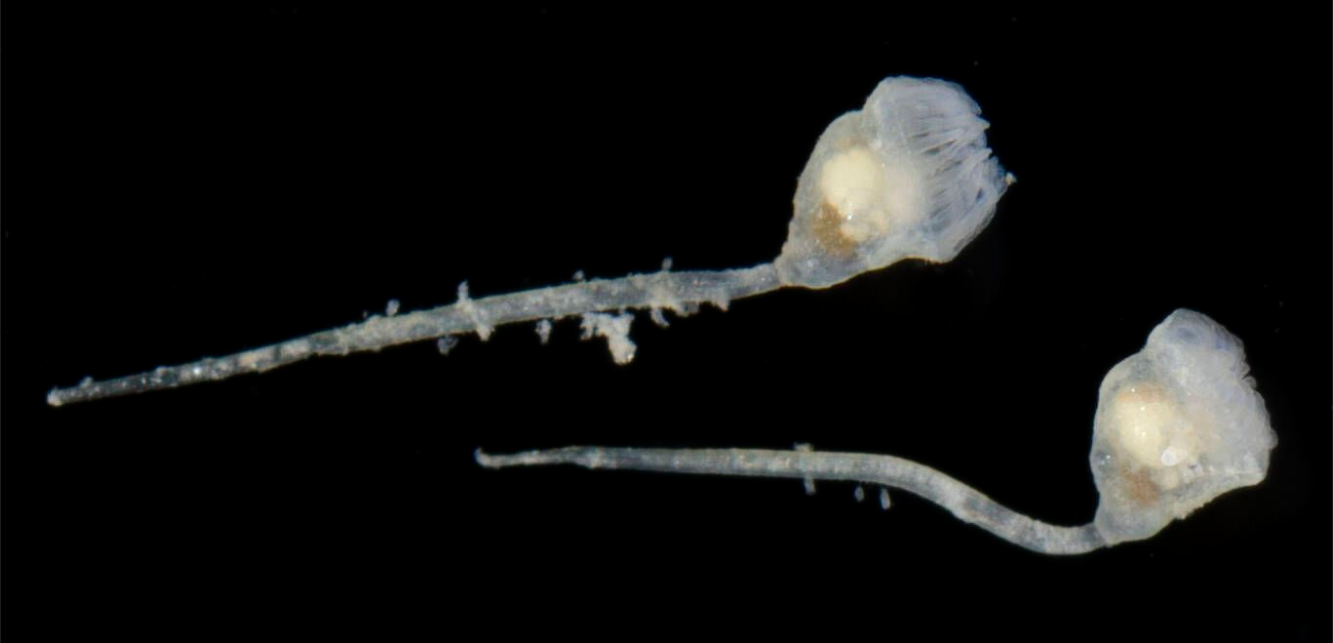
성숙한 내항동물 개충의 몸은 표면에 부착되는 비교적 긴 자루에 부착된 꽃받침이 있는 술잔 모양의 구조를 가지고 있다. 꽃받침의 가장자리에는 몸 벽의 연장인 8개에서 30개의 단단한 촉수 "왕관"이 있다. 촉수 "왕관"의 바닥은 촉수가 후퇴할 때 촉수를 부분적으로 덮는 막으로 둘러싸여 있다. 입과 항문은 심방(촉수 "왕관"으로 둘러싸인 공간)의 반대쪽에 위치하며, 둘 다 괄약근으로 닫을 수 있다. 창자는 U자형이며, 꽃받침의 바닥을 향해 굽어 위를 형성하기 위해 넓어진다. 이것은 각각 여러 개의 섬모를 가진 단일 세포층으로 구성된 막으로 덮여 있다.[4]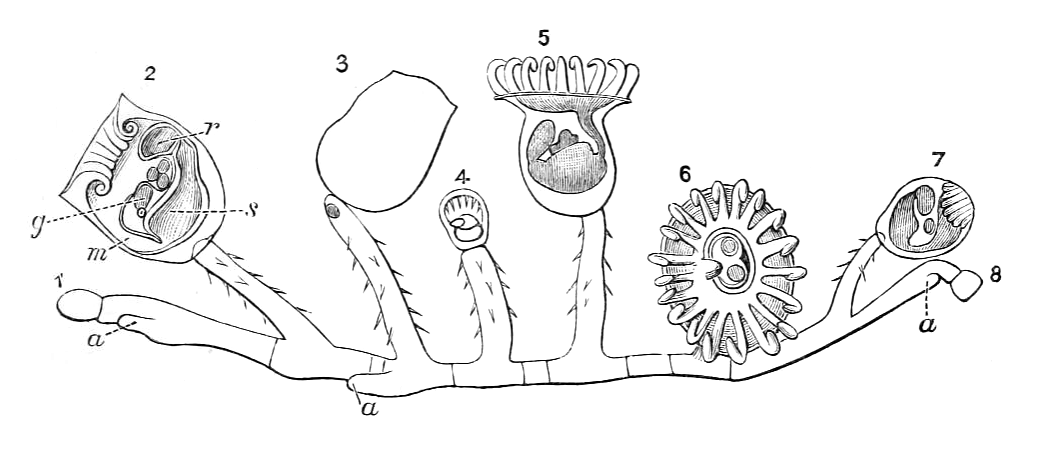
군체 종의 자루는 공유된 부착판 또는 표면을 가로질러 뻗어 있는 튜브인 스톨론 네트워크에서 나온다.[4] 단독 종에서 자루는 근육질 빨판 또는 유연한 발로 끝나거나 표면에 시멘트된다.[10] 자루는 근육질이며 특징적인 끄덕임 운동을 생성한다. 일부 종에서는 분절된다. 일부 단독 종은 근육질 발로 기어가거나 재주넘기를 함으로써 움직일 수 있다.[4]
몸 벽은 표피와 외부 큐티클로 구성되어 있으며,[4] 이는 주로 교차된 콜라겐 섬유로 구성된다. 표피는 단일 세포층만 포함하며, 각 세포는 여러 개의 섬모("털")와 큐티클을 관통하는 미세융모 (작은 "주름")를 가지고 있다.[4] 군체 종의 스톨론과 자루는 키틴으로 강화된 더 두꺼운 큐티클을 가지고 있다.[10]
체강(내부 유체로 채워진 복막으로 덮인 공간)은 없으며 다른 내부 기관은 위와 촉수 "왕관"의 바닥 사이에 있는 결합 조직에 내장되어 있다. 신경계는 결합 조직을 통과하여 표피 바로 아래에 있으며 한 쌍의 신경절에 의해 제어된다. 신경은 이들로부터 꽃받침, 촉수, 자루로 뻗어 있으며 이 모든 영역의 감각 기관으로 뻗어 있다.[4]
'''내항동물문'''은 군체 또는 단체성 동물로, 기껏해야 5mm 이하의 작은 동물이다. 오래전에는 태형동물과 혼동되기도 했지만, 현재는 독립된 분류군으로 여겨진다. 참고로 이 명칭은 항문이 촉수관 안쪽에 있다는 데서 유래하며, 태형동물은 그 바깥쪽에 있어 외항동물이라고 부르는 것과 대조를 이룬다.
3. 2. 꽃받침
개체는 일반적으로 1mm 길이지만 0.1mm 에서 7mm 길이 범위에 있다.[4] 성숙한 내항동물 개충의 몸은 표면에 부착되는 비교적 긴 자루에 부착된 꽃받침이 있는 술잔 모양의 구조를 가지고 있다. 꽃받침의 가장자리에는 몸 벽의 연장인 8개에서 30개의 단단한 촉수 "왕관"이 있다.[4]촉수 "왕관"의 바닥은 촉수가 후퇴할 때 촉수를 부분적으로 덮는 막으로 둘러싸여 있다. 입과 항문은 심방(촉수 "왕관"으로 둘러싸인 공간)의 반대쪽에 위치하며, 둘 다 괄약근으로 닫을 수 있다. 창자는 U자형이며, 꽃받침의 바닥을 향해 굽어 위를 형성하기 위해 넓어진다. 이것은 각각 여러 개의 섬모를 가진 단일 세포층으로 구성된 막으로 덮여 있다.[4] 신경은 신경절로부터 꽃받침, 촉수, 자루로 뻗어 있으며 이 모든 영역의 감각 기관으로 뻗어 있다.[4]
몸통부, 즉 '''꽃받침'''부는 촉수관과 대부분의 내장이 여기에 있다. 꽃받침부는 윗면에 촉수관이 있으며, 대개 약간 비스듬히 기울어져 있다. 촉수의 배치는 다소 말굽 모양을 하고 있으며, 개개의 촉수는 꽃받침부의 길이를 넘을 정도로 길지 않고, 그다지 유연하지도 않다. 그 표면은 온통 섬모로 덮여 있다. 촉수가 배열된 안쪽 영역의 한쪽 끝에 입이, 다른 한쪽 끝에 항문이 있다.
또한, 조건이 나빠지면 꽃받침부는 버려지는 경우가 있다. 조건이 좋아지면 바로 재생된다.
3. 3. 내부 구조
대부분의 내항동물 종은 군체 생활을 하며, 그 구성원들은 완전히 독립적인 동물이 아니기 때문에 "개체"라고 불린다.[7] 개체는 일반적으로 1mm 길이지만 0.1mm에서 7mm 길이 범위에 있다.[4] 내항동물은 겉으로는 태형동물(외항동물)과 유사하지만, 먹이 공급 방식과 내부 해부학이 다르다.[4]성숙한 내항동물 개충의 몸은 술잔 모양으로, 표면에 부착되는 비교적 긴 자루에 꽃받침이 부착되어 있다. 꽃받침 가장자리에는 8개에서 30개의 단단한 촉수가 있는데, 이는 몸 벽의 연장이다. 촉수 "왕관" 바닥은 막으로 둘러싸여 있어 촉수가 후퇴할 때 부분적으로 덮는다. 입과 항문은 심방(촉수 "왕관"으로 둘러싸인 공간)의 반대쪽에 있으며, 둘 다 괄약근으로 닫을 수 있다. 창자는 U자형이며, 꽃받침 바닥을 향해 굽어 위를 형성하기 위해 넓어진다. 막은 여러 개의 섬모를 가진 단일 세포층으로 구성된다.[4]
군체 종의 자루는 공유된 부착판 또는 표면을 가로질러 뻗어 있는 스톨론 네트워크에서 나온다.[4] 단독 종에서 자루는 근육질 빨판 또는 유연한 발로 끝나거나 표면에 시멘트된다.[10] 자루는 근육질이며 끄덕임 운동을 한다. 일부 종에서는 분절된다. 일부 단독 종은 근육질 발로 기어가거나 재주넘기를 하여 움직일 수 있다.[4]
몸 벽은 표피와 외부 큐티클로 구성되며,[4] 주로 교차된 콜라겐 섬유로 구성된다. 표피는 단일 세포층만 포함하며, 각 세포는 여러 개의 섬모와 큐티클을 관통하는 미세융모를 가진다.[4] 군체 종의 스톨론과 자루는 키틴으로 강화된 더 두꺼운 큐티클을 가진다.[10]
체강은 없으며 다른 내부 기관은 위와 촉수 "왕관"의 바닥 사이에 있는 결합 조직에 내장되어 있다. 신경계는 결합 조직을 통과하여 표피 바로 아래에 있으며 한 쌍의 신경절에 의해 제어된다. 신경은 이들로부터 꽃받침, 촉수, 자루로 뻗어 있으며 이 모든 영역의 감각 기관으로 뻗어 있다.[4]
내항동물의 몸은 대략 몸통부와 줄기부로 이루어진다. 몸통부인 '''꽃받침'''부는 촉수관과 대부분의 내장이 있다. 줄기부는 막대 모양이거나 더 복잡한 형태이다. 단체성인 것에서는 '''족반'''이 되어 기판상에 부착한다. 군체성인 것에서는 식물의 포복경과 같은 '''주근'''을 뻗어 새로운 개체를 생성한다.
꽃받침부는 윗면에 촉수관이 있으며, 약간 비스듬히 기울어져 있다. 촉수는 말굽 모양을 하고 있으며, 섬모로 덮여 있다. 촉수가 배열된 안쪽 영역의 한쪽 끝에 입이, 다른 한쪽 끝에 항문이 있다.
소화계는 U자 모양이며, 식도, 위, 창자, 직장으로 이어진다. 체강은 간충직세포로 채워진다. 배출계로는 원신관 한 쌍이 있다. 생식기도 꽃받침 부위에 있다.
| 내항동물 | 태형동물 (외항동물) | |
|---|---|---|
| 촉수 | 단단함 | 속이 빔 |
| 먹이 섭취 흐름 | 촉수 기부에서 끝 부분으로 | 촉수 끝 부분에서 기부로 |
| 항문 위치 | 촉수 "왕관" 안쪽 | 촉수 "왕관" 바깥쪽 |
| 체강 | 없음 | 세 부분으로 나뉨 |
| 군체 내 창시자 개체의 형태 | 다른 개체와 같음 | 일반적인 개체와 달리 둥긂[9] |
| 성체로의 변태 | 대부분의 유충 구조를 유지 | 대부분의 유충 구조를 파괴 |
| 배설 기관 | 원신관 | 없음 |
4. 생태
내항동물은 크기가 작고 동물학자들에 의해 거의 연구되지 않아, 표본이 이미 동일한 지역에서 발생하는 종에 속하는지, 아니면 인간의 활동으로 나타난 침입종인지 판단하기 어렵다.[34] 주근이나 족반으로 기질에 부착하는 고착성 동물이지만, 일부는 천천히 이동할 수 있다.
4. 1. 서식지
모든 내항동물은 부착 생활을 한다.[4] 대부분 해양 종이지만, ''Loxosomatoides sirindhornae''와 ''Urnatella gracilis'' 두 종은 담수에서 산다. ''Loxosomatoides sirindhornae''는 2004년 태국 중부에서 보고되었고, ''Urnatella gracilis''는 남극을 제외한 모든 대륙에서 발견된다.[3] 군체 생활을 하는 종은 모든 대양에서 발견되며, 바위, 조개 껍질, 조류 및 수중 건물에 서식한다.[4] 단독 생활을 하는 종은 해양 종이며,[3] 해면동물, 외항동물 및 부착 생활을 하는 환형동물과 같이 물의 흐름을 생성하여 먹이를 얻는 다른 동물에 서식한다.[10] 대부분의 종은 수심 50m보다 얕은 곳에 살지만, 몇몇 종은 심해에서 발견된다.[30]대부분 해산이며, 소수가 기수산이고, 시마미즈우동게 등 2종만이 담수산 종이다.
4. 2. 섭식
촉수 양쪽을 따라 여러 개의 섬모가 있는 세포 띠가 뻗어 있어 각 촉수를 이웃과 연결하지만, 항문과 가장 가까운 띠에는 틈이 있다. 별도의 섬모 띠는 "왕관" 기부 안쪽 면을 따라 홈을 따라 자라며, 각 촉수의 안쪽 표면 위로 좁게 연장되어 있다.[10] 촉수 측면의 섬모는 촉수의 기저부에서 "왕관"으로 흘러 들어가 "왕관" 중앙 위로 빠져나가는 흐름을 생성한다.[4] 이러한 섬모는 음식 입자를 촉수 안쪽 표면의 섬모로 전달하고, 안쪽 섬모는 입자를 홈 안과 주변으로 밀어넣어 입으로 이동시키는 하향 흐름을 생성한다.[10]내항동물은 일반적으로 섬모 체질 중 하나 또는 둘 다를 사용한다. 섬모 체질에서는 한 개의 섬모 띠가 먹이 섭취 흐름을 생성하고 다른 띠가 음식 입자를 가두는(체(sieve)) 방식이다. 그리고 다운스트림 수집은 음식 입자가 통과하려 할 때 가두는 방식이다. 내항동물에서는 다운스트림 수집이 흐름을 생성하는 동일한 섬모 띠에 의해 수행된다. 환형동물의 트로코포라(trochozoan) 유충도 다운스트림 수집을 사용하지만, 음식 입자를 가두기 위해 별도의 섬모 세트를 사용한다.[11]
또한, 촉수의 샘은 큰 입자를 포획하는 끈적한 실을 분비한다.[4] 1993년 남극 반도 주변에서 보고된 비군체 종은 표면적으로 자포동물의 자포(cnidocyte)와 유사하며 끈적한 실을 발사하는 세포를 가지고 있다. 이 특이한 세포는 입 주변에 위치하며, 먹이를 포획하는 추가적인 수단을 제공할 수 있다.[12]
위와 창자는 영양분을 흡수하는 것으로 생각되는 미세융모로 덮여 있다. "왕관" 안에 열려 있는 항문은 촉수가 물에서 음식을 걸러낸 후 고체 폐기물을 외부 흐름으로 배출한다. 일부 과에서는 입으로 음식을 전달하는 홈의 높이보다 원뿔 위에 솟아 있다.[4][13] 대부분의 종은 내부 체액에서 용해성 폐기물을 추출하여 입 근처의 구멍을 통해 제거하는 한 쌍의 원신관을 가지고 있다. 그러나 담수 종인 ''Urnatella gracilis''는 꽃받침과 자루에 여러 개의 신관을 가지고 있다.[4]
개체는 확산을 통해 산소를 흡수하고 이산화 탄소를 방출하며,[4] 이는 작은 동물에게 효과적이다.[14] 대부분이 해산이며, 소수가 기수산이고, 시마미즈우동게 등 2종만이 담수산 종이다. 촉수관을 통해 여과 섭식을 한다.
4. 3. 공생 및 천적
누디브런치(바다 달팽이)의 일부 종, 특히 ''Trapania'' 속의 종들과 터블러리아 편형동물은 내항동물을 포식한다.[31]수생 곤충 날도래 ''Corydalus cornutus''의 수생 유충에 담수 내항동물 ''Urnatella gracilis''의 작은 군체가 서식하는 것이 발견되었다. 외항동물은 포식자로부터의 보호와 분산 수단을 얻고, 유충의 아가미 옆에 군체가 서식하는 경우가 많으므로 산소와 영양소가 풍부한 물을 얻을 수 있다.[32] 백해에서는 군집을 이루지 않는 내항동물 ''Loxosomella nordgaardi''가 이끼벌레(외항동물) 군집, 주로 군집의 가장자리 또는 큰 이끼벌레 군집이 음식을 걸러낸 물을 배출하는 틈새인 "굴뚝"에 부착하여 사는 것을 선호한다. 관찰 결과 내항동물과 이끼벌레 모두 이 연관성에서 이점을 얻는 것으로 보인다. 서로가 먹이를 얻는 데 필요한 물의 흐름을 개선하고, 내항동물의 더 긴 섬모는 이끼벌레가 잡는 것과 다른 음식을 포획하는 데 도움이 될 수 있으므로 동물들이 동일한 음식을 놓고 경쟁하지 않는다.[33]
5. 생식과 발생
내항동물은 유성 생식과 무성 생식(출아)을 모두 할 수 있다. 대부분 자웅동체이지만, 일부는 성체가 되면서 수컷에서 암컷으로 성전환을 하기도 하고, 어떤 종은 평생 같은 성별을 유지한다. 수정된 알은 나선형동물 패턴에 따라 나선형 난할로 분열하며 유생으로 발달한다. 중배엽은 초기 배아에서 "4d"로 표시된 특정 세포에서 발달하며, 어떤 단계에서도 체강은 없다.[4]
일부 종의 유생은 트로코포라 유생과 유사하며, 플랑크톤 생활을 한다. Loxosomella 속 및 Loxosoma 속의 일부 종에서는 유생이 분리되어 새로운 개체를 형성하기도 한다. 그러나 대부분의 유생은 감각 털, 눈점, 원신관, 섬모를 가진 발을 가지며, 정착 후 복잡한 변태를 겪는다.[4][10]
모든 종은 복제에 의해 출아를 할 수 있다. 군체성 종은 스톨론 또는 자루에서 새로운 개충을 생산하여 큰 군체를 형성하고, 단독 종에서는 복제체가 심방 바닥에서 형성되어 방출된다.[4][10] 무성 생식에 의한 번식이 활발하며, 단독 생활 및 군체 생활 종 모두 출아를 통해 번식한다. 군체 생활 종은 주근을 뻗어 새로운 개체를 만들고, 단독 생활 종은 꽃받침부에 새로운 개체의 싹을 생성하여 독립시킨다.
5. 1. 생식
대부분의 내항동물 종은 자웅동체이지만, 일부는 성체가 되면서 수컷에서 암컷으로 성전환을 하기도 하고, 어떤 종은 평생 같은 성별을 유지한다. 개체는 심방과 위 사이에 위치하고 심방의 단일 생식공으로 열리는 한두 쌍의 생식선을 가지고 있다.[10] 알은 난소에서 수정되는 것으로 추정된다. 대부분의 종은 플랑크톤 유생으로 부화하는 알을 방출하지만, 일부는 생식공에서 알을 품는다. 작은 알을 품는 종은 태반과 유사한 기관으로 영양을 공급하고, 더 큰 알을 가진 종의 유생은 저장된 난황으로 살아간다.[4] 수정된 알이 유생으로 발달하는 것은 전형적인 나선형동물 패턴을 따른다. 세포는 나선형 난할로 분열하고, 중배엽은 초기 배아에서 "4d"로 표시된 특정 세포에서 발달한다.[16] 어떤 단계에서도 체강은 없다.[4]일부 종의 유생은 트로코포라 유생인데, 이는 플랑크톤이며, "적도" 주위의 두 개의 섬모 띠를 사용하여 부유하는 음식 입자를 입으로 쓸어 넣고, 입은 더 많은 섬모를 사용하여 위로 밀어 넣으며, 위는 더 많은 섬모를 사용하여 소화되지 않은 잔해를 항문을 통해 배출함으로써 먹이를 섭취한다.[17] Loxosomella 속 및 Loxosoma 속의 일부 종에서는 유생이 분리되어 새로운 개체를 형성하는 하나 또는 두 개의 싹을 생성하는 반면, 트로코포라 유생은 붕괴된다. 그러나 대부분은 상단과 전면에 감각 털, 한 쌍의 색소 컵 눈점, 한 쌍의 원신관, 그리고 하단에 큰 섬모를 가진 발을 가진 유생을 생산한다.[10] 정착 후, 발과 전면 털이 표면에 부착된다. 대부분의 종의 유생은 복잡한 변태를 겪으며, 내부 기관이 최대 180° 회전하여 입과 항문이 모두 위를 향할 수 있다.[4]
모든 종은 복제에 의해 출아를 할 수 있다. 군체성 종은 스톨론 또는 자루에서 새로운 개충을 생산하며, 이런 방식으로 큰 군체를 형성할 수 있다.[4] 단독 종에서는 복제체가 심방의 바닥에서 형성되며, 기관이 발달하면 방출된다.[10] 실제로는 무성 생식에 의한 번식이 활발하며, 단독 생활을 하는 종과 군체 생활을 하는 종 모두 출아에 의해 증가한다. 군체 생활을 하는 종은 주근이 뻗어 그 끝에 새로운 개체를 만들고, 단독 생활을 하는 종은 꽃받침부에 새로운 개체의 싹을 생성하여 그것이 성장하여 독립한다.
5. 2. 발생
대부분의 내항동물 종은 자웅동체이지만, 일부는 성체가 되면서 수컷에서 암컷으로 성전환을 하기도 하고, 어떤 종은 평생 같은 성별을 유지한다. 개체는 심방과 위 사이에 위치하고 심방의 단일 생식공으로 열리는 한두 쌍의 생식선을 가지고 있다.[10] 알은 난소에서 수정되는 것으로 추정된다. 대부분의 종은 플랑크톤 유생으로 부화하는 알을 방출하지만, 일부는 생식공에서 알을 품는다. 작은 알을 품는 종은 태반과 유사한 기관으로 영양을 공급하고, 더 큰 알을 가진 종의 유생은 저장된 난황으로 살아간다.[4] 수정된 알이 유생으로 발달하는 것은 전형적인 나선형동물 패턴을 따른다. 세포는 나선형 난할로 분열하고, 중배엽은 초기 배아에서 "4d"로 표시된 특정 세포에서 발달한다.[16] 어떤 단계에서도 체강은 없다.[4]일부 종의 유생은 트로코포라 유생인데, 이는 플랑크톤이며, "적도" 주위의 두 개의 섬모 띠를 사용하여 부유하는 음식 입자를 입으로 쓸어 넣고, 입은 더 많은 섬모를 사용하여 위로 밀어 넣으며, 위는 더 많은 섬모를 사용하여 소화되지 않은 잔해를 항문을 통해 배출함으로써 먹이를 섭취한다.[17] Loxosomella 속 및 Loxosoma 속의 일부 종에서는 유생이 분리되어 새로운 개체를 형성하는 하나 또는 두 개의 싹을 생성하는 반면, 트로코포라 유생은 붕괴된다. 그러나 대부분은 상단과 전면에 감각 털, 한 쌍의 색소 컵 눈점, 한 쌍의 원신관, 그리고 하단에 큰 섬모를 가진 발을 가진 유생을 생산한다.[10] 정착 후, 발과 전면 털이 표면에 부착된다. 대부분의 종의 유생은 복잡한 변태를 겪으며, 내부 기관이 최대 180° 회전하여 입과 항문이 모두 위를 향하게 된다.[4]
모든 종은 복제에 의해 출아를 할 수 있다. 군체성 종은 스톨론 또는 자루에서 새로운 개충을 생산하며, 이런 방식으로 큰 군체를 형성할 수 있다.[4] 단독 종에서는 복제체가 심방의 바닥에서 형성되며, 기관이 발달하면 방출된다.[10] 암수 이체이며, 발생은 암컷의 체내에서 진행된다. 초기 유생은 트로코포라에 가깝다.
그러나 실제로는 무성 생식에 의한 번식이 활발하며, 단독 생활을 하는 종과 군체 생활을 하는 종 모두 출아에 의해 증가한다. 군체 생활을 하는 종은 주근이 뻗어 그 끝에 새로운 개체를 만들고, 단독 생활을 하는 종은 꽃받침부에 새로운 개체의 싹을 생성하여 그것이 성장하여 독립한다.
6. 분류
내항동물은 4개 과에 약 150여 종으로 분류된다.[4][6] 대부분의 종이 해양성이지만, 일부 종(''우르나텔라 그라실리스'')은 담수에 널리 분포한다. 일본에는 약 30종이 확인되었다.
내항동물 문은 록소소마목, 페디켈리나목, 우르나텔라목으로 구성되어 있으며, 4개의 과로 분류된다:[4][6]
6. 1. 록소소마목 (Loxosomatida)
록소소마목(Loxosomatida)은 록소칼리푸스과와 록소소마과로 나뉜다.[4][6] 록소칼리푸스과는 록소칼리푸스속(''Loxokalypus'')을, 록소소마과에는 록소소마속(''Loxosoma''), ''Loxosomella'', ''Loxomitra'', ''Loxosomespilon'', ''Loxocore''를 포함한다.[21]
6. 1. 1. 록소칼리푸스과 (Loxokalypodidae)
록소칼리푸스과 (Loxokalypodidae)는 록소소마목에 속하는 내항동물 과이다.[20] 유일속인 록소칼리푸스속(''Loxokalypus'')을 포함하고 있다.[20] 록소칼리푸스과는 아래 표에서 다른 내항동물 과들과 비교할 수 있다.
6. 1. 2. 록소소마과 (Loxosomatidae)
록소소마과에는 록소소마속(''Loxosoma'')[21], ''Loxosomella'', ''Loxomitra'', ''Loxosomespilon'', ''Loxocore''[21]가 있다. 록소소마과는 단독성[22]이며, 컵과 자루 사이에 격막[22]과 별세포 기관[22]이 없다. 항문은 원뿔 모양이다.[4]6. 2. 페디켈리나목 (Pedicellinida)
페디켈리나과는 페디켈리나목의 한 과이다.| 과 | 페디켈리나과 |
|---|---|
| 속 | 키타스피스, 록소소마토이데스, 묘소마, 페디켈리나[19] |
| 군체성[22] | 군체성 |
| 컵과 자루 사이의 격막[22] | 예 |
| 별세포 기관[22] | 예 |
| 항문이 원뿔 모양[4] | 아니오 |
| 스톨론 존재[22] | 예 |
| 분절된 줄기[4][22] | 아니오 |
6. 2. 1. 페디켈리나과 (Pedicellinidae)
페디켈리나과에 속하는 속은 다음과 같다.[19]- ''Pedicellina''
- ''Myosoma''
- ''Chitaspis''
- ''Loxosomatoides''
- ''Coriella''
6. 3. 우르나텔라목 (Urnatellida)
''Barentsiidae''는 바렌치아, ''코리엘라'', ''페디켈리놉시스'', ''의페디켈리나'', ''요관충''[18] 등의 속을 포함한다.6. 3. 1. 바렌치아과 (Barentsiidae)
7. 진화
내항동물은 19세기에 발견되었을 때 이끼벌레(외항동물)와 함께 이끼벌레동물문에 속하는 강으로 여겨졌다. 두 그룹 모두 착생 생활을 하고 여과 섭식을 하며, 섬모가 있는 촉수 "왕관"으로 먹이를 걸러 먹었기 때문이다. 그러나 1869년부터 내항동물의 항문이 섭식 구조 안에 위치하고, 배아의 세포 분열 패턴의 차이 등 여러 차이점이 밝혀지면서, 과학자들은 이 두 그룹을 별개의 문으로 간주하게 되었다.[26] 이후 "이끼벌레동물"은 항문이 섭식 기관 밖에 있는 외항동물의 또 다른 이름이 되었다.[27] 그러나 2007년과 2008년 한 연구팀의 연구에 따르면 내항동물을 강으로 이끼벌레동물에 통합하고, 현재 확인된 이끼벌레를 지칭하는 이름으로 ''외항동물''을 부활시키는 것을 주장했다.[26][28]
1996년 이후의 연구 결과는 내항동물이 트로코조아에 속한다는 것이 일반적인 견해이다. 트로코조아는 원구동물 "상문"으로, 가장 기본적인 유생 형태인 트로코포라를 공통적으로 갖는 무리이다. 트로코조아에는 연체동물, 환형동물, 납작벌레, 끈벌레 등이 포함된다. 그러나 과학자들은 트로코조아 내에서 어떤 문이 내항동물과 가장 밀접한 관련이 있는지에 대해 의견이 일치하지 않는다.[29] 2008년의 분석에서는 내항동물과 외항동물이 서로 가장 가까운 친척 관계에 있는 그룹을 지칭하는 용어로 1869년 이전의 "이끼벌레동물"의 의미를 재도입했다.[26]
처음에는 에렌베르크가 1831년에 태형동물(Bryozoa)을 세웠을 때 태형동물과 동일시되어 이에 포함되었다. 그러나 니체(1870)가 항문의 위치에 따라 외항동물(Ectoprocta)과 내항동물(Endoprocta)로 나누었다. 더 나아가 코리(1929)는 근연 관계가 아닌데 이름이 비슷한 것은 좋지 않다며 곡형동물(Kamptozoa)이라는 이름을 제창했고, 이것이 사용되기도 한다.
7. 1. 화석 기록

내항동물은 작고 부드러운 몸을 가지고 있기 때문에 화석이 극히 드물다.[24] 1977년, 사이먼 콘웨이 모리스는 캐나다의 버제스 셰일에서 발견된, 컵, 자루 및 부착부를 가진 정착 생활을 하는 동물 ''디노미스쿠스''에 대한 최초의 설명을 제공했는데, 이는 약 5억 5백만 년 전에 형성되었다. 콘웨이 모리스는 이 동물을 가장 초기의 내항동물로 간주했는데, 그 이유는 입과 항문이 컵 위의 구조물 고리 안에 위치했지만, 이러한 구조물은 평평하고 다소 뻣뻣한 반면, 현대 내항동물의 촉수는 유연하고 둥근 단면을 가지고 있다는 점을 언급했다.[23]
1992년 J.A. 토드와 P.D. 테일러는 ''디노미스쿠스''가 전형적인 둥글고 유연한 촉수가 없고, 화석이 내항동물과 명확하게 유사한 다른 특징을 보이지 않기 때문에 내항동물이 아니라고 결론 내렸다. 그들의 의견으로는, 가장 초기의 화석 내항동물은 영국에서 후기 쥐라기 암석에서 발견한 표본들이었다. 이들은 많은 면에서 현대의 군체 속 ''바렌티시아''와 유사한데, 여기에는 군체가 부착된 표면을 덮는 스톨론 네트워크로 연결된 수직의 개체, 주름의 가로 띠가 있는 부피가 큰 소켓으로 스톨론에 연결된 곧은 자루, 현대의 ''바렌티시아'' 종과 유사한 전반적인 크기와 비율이 포함된다.[24]
또 다른 종인 ''코틸레디온 틸로데스''는 1999년에 처음 기술되었는데, 현존하는 내항동물보다 크고, 높이가 8mm~56mm에 달하며, 현대 종과 달리, 작은 판 모양의 구조물인 경화판으로 "갑옷"을 갖추고 있었다. ''C. 틸로데스''는 현대 내항동물과 유사한 정착 생활 방식을 가지고 있었다. 확인된 ''C. 틸로데스''의 화석은 중국 남부에서 5억 2천만 년 된 암석에서 발견되었다. 이것은 초기 내항동물을 캄브리아기 대폭발 시기에 위치시킨다.[25]
7. 2. 계통
내항동물은 19세기에 발견되었을 때 이끼벌레(외항동물)와 함께 이끼벌레동물문에 속하는 강으로 여겨졌다. 두 그룹 모두 착생 생활을 하고 여과 섭식을 하며, 섬모가 있는 촉수 "왕관"으로 먹이를 걸러 먹었기 때문이다. 그러나 1869년부터 내항동물의 항문이 섭식 구조 안에 위치하고, 배아의 세포 분열 패턴의 차이 등 여러 차이점이 밝혀지면서, 과학자들은 이 두 그룹을 별개의 문으로 간주하게 되었다.[26] 이후 "이끼벌레동물"은 항문이 섭식 기관 밖에 있는 외항동물의 또 다른 이름이 되었다.[27]1996년 이후의 연구 결과에 따르면 내항동물은 트로코조아에 속한다는 것이 일반적인 견해이다. 트로코조아는 원구동물 "상문"으로, 가장 기본적인 유생 형태인 트로코포라를 공통적으로 갖는 무리이다. 트로코조아에는 연체동물, 환형동물, 납작벌레, 끈벌레 등이 포함된다. 그러나 과학자들은 트로코조아 내에서 어떤 문이 내항동물과 가장 밀접한 관련이 있는지에 대해 의견이 일치하지 않는다.[29]
분자 유전학적 정보에 따르면, 내항동물과 외항동물은 모두 환형동물에 속하지만 자매군은 아닌 것으로 여겨진다.
참조
[1]
논문
A sclerite-bearing stem group entoproct from the early Cambrian and its implications
2013-01
[2]
논문
The first fossil entoproct
[3]
논문
"Loxosomatoides sirindhornae", new species, a freshwater kamptozoan from Thailand (Entoprocta)
[4]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks/Cole
[5]
서적
The names of plants
https://books.google[...]
Cambridge University Press
2009-09-10
[5]
논문
Camptomelic dysplasia
[6]
웹사이트
ITIS Standard Report Page: Entoprocta
https://www.itis.gov[...]
Integrated Taxonomic Information System
2009-08-26
[7]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[8]
서적
Shorter Oxford English Dictionary
Oxford University Press
[9]
서적
The fossil book
Dover Publications
2009-08-07
[10]
서적
Encyclopedia of Life Sciences
John Wiley & Sons, Ltd.
[11]
논문
Downstream collecting in ciliary suspension feeders: the catch-up principle
https://www.int-res.[...]
2009-09-12
[12]
논문
On Antarctic Entoprocta
http://www.biolbull.[...]
2009-09-12
[13]
서적
The invertebrates: a synthesis
Wiley-Blackwell
[14]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[15]
서적
Invertebrate Zoology
Brooks / Cole
2004
[16]
논문
Mesoderm in spiralians: the organizer and the 4d cell
Wiley InterScience
[17]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[18]
웹사이트
ITIS Standard Report Page: Barentsiidae
https://www.itis.gov[...]
2009-09-14
[19]
웹사이트
ITIS Standard Report Page: Pedicellinidae
https://www.itis.gov[...]
2009-09-14
[20]
웹사이트
ITIS Standard Report Page: Loxokalypodidae
https://www.itis.gov[...]
2009-09-14
[21]
웹사이트
ITIS Standard Report Page: Loxosomatidae
https://www.itis.gov[...]
2009-09-14
[22]
논문
Systematic revision of colonial kamptozoans (entoprocts) of the Pacific coast of North America
[23]
논문
A new entoproct-like organism from the Burgess Shale of British Columbia
http://palaeontology[...]
2009-09-13
[24]
논문
The first fossil entoproct
1992-07
[25]
뉴스
ScienceShot: Fossils of Enigmatic Sea Creature Emerge
ScienceNOW
2013-01-17
[26]
논문
Spiralian phylogenomics supports the resurrection of Bryozoa comprising Ectoprocta and Entoprocta
2007-12
[27]
논문
The new view of animal phylogeny
http://www-fourier.u[...]
[28]
논문
Phylogenomic analyses of lophophorates (brachiopods, phoronids and bryozoans) confirm the Lophotrochozoa concept
2008-08-22
[29]
서적
Phylogeny and evolution of the Mollusca
University of California Press
2009-09-13
[30]
웹사이트
A New Inside Anus Found Very Deep
http://www.deepseane[...]
[31]
논문
Predation on kamptozoans (Entoprocta)
2005-05-11
[32]
논문
The phoretic association of ''Urnatella gracilis'' (Entoprocta: Urnatellidae) and ''Nanocladius downesi'' (Diptera: Chironomidae) on ''Corydalus cornutus'' (Megaloptera: Corydalidae)
The North American Benthological Society
1983-11
[33]
논문
Substrate preferences of a non-colonial kamptozoan, and its interactions with bryozoan hosts
2002-12
[34]
논문
Detecting invasions of marine organisms: kamptozoan case histories
2000-03
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com