태형동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
태형동물은 몸길이가 약 1mm 안팎의 작은 동물들이 모여 군체를 이루는 무척추동물로, 전 세계에 약 5,000종이 알려져 있다. 키틴질 외골격과 촉수관을 가지며, 해양 및 담수 환경에 서식한다. 여과 섭식을 하며, 무성 생식으로 번식한다. 과거에는 산호 등과 함께 식충류로 불리기도 하였으나, 1831년 Ehrenberg에 의해 이끼벌레류(Bryozoa)로 분류되었다. 태형동물은 형태, 서식지, 외골격 물질 등에 따라 협후강, 나후강, 피후강으로 분류된다.
더 읽어볼만한 페이지
- 태형동물 - 나후강
나후강은 순구강류와 빗후강류로 나뉘는 외항동물문에 속하는 강이다. - 태형동물 - 협후강
협후강은 현존하는 원구목과 더불어 Cryptostomata, Cryptostomida, Cystoporata, Cystoporida, Fenestrida, Melicerititida, Trepostomatida 등 멸종된 다양한 목들을 포함하는 태형동물문의 한 분류군이다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 태형동물 - [생물]에 관한 문서 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
 | |
| 학명 | Bryozoa |
| 명명자 | 크리스티안 고트프리트 에렌베르크, 1831년 |
| 이명 | 외항동물 (Ectoprocta) (니체, 1869년) (이전에는 외항동물문의 하위문) 폴리조아(Polyzoa) (이전 명칭) |
| 로마자 표기 | Bryozoa |
| 분류 | |
| 계 | 진핵생물 |
| 역 | 동물계 |
| 문 | 외항동물문 |
| 강 | 협후강 Stenolaemata 나후강 Gymnolaemata 은후강 Phylactolaemata |
| 생물학적 특징 | |
| 크기 | 0.5mm |
| 서식지 | 수생, 군체 생활 |
| 특징 | 촉수관을 가진 로포포아 좌우대칭 군체 생활 석회질 또는 키틴질 껍질 각 개체는 조이드라고 불림 |
| 화석 기록 | |
| 출현 시기 | 초기 캄브리아기 (이의 있음) 오르도비스기 ~ 현재 |
| 역사 | |
| 최초 발견 | 1830년, 존 본햄 톰슨 |
| 최초 명명 | 1831년, 크리스티안 고트프리트 에렌베르크 |
| 명칭 혼란 | 초기에는 폴리조아(Polyzoa)와 브리오조아(Bryozoa) 혼용 1911년, T.R.R. 스테빙에 의해 용어 정리 |
| 생태 및 중요성 | |
| 생태적 역할 | 해양 생태계의 중요한 구성원 서식지 제공 및 생물 다양성 유지에 기여 |
| 경제적 영향 | 일부 종은 침입종으로 작용하여 해양 환경에 부정적인 영향 양식업 및 선박에 피해를 입힘 |
| 참고 문헌 | |
| 참고 자료 | 캘리포니아 대학교 고생물학 박물관의 브리오조아 소개 |journal=Zootaxa |volume=3703 |issue=1 |pages=67–74 |doi=10.11646/zootaxa.3703.1.14 |url=https://www.researchgate.net/publication/279928935}} |
2. 특징
태형동물은 몸길이가 1mm 안팎인 매우 작은 동물로서, 이들이 많이 모여 나뭇가지, 부채, 덩어리 등 다양한 모양의 군체를 이룬다. 각각의 개체는 개충(zooid)이라고 불리며, 주로 키틴질이나 탄산칼슘으로 된 외골격(蟲室, zooecium)과 그 속에 들어있는 부드러운 몸체(충체, polypide)로 구성된다.[24]
가장 두드러진 특징은 입 주위를 둘러싼 촉수관(lophophore)이다. 이는 속이 빈 촉수들이 원형 또는 말굽 모양으로 배열된 기관으로, 표면의 섬모를 이용하여 물의 흐름을 만들고 식물성 플랑크톤과 같은 작은 먹이 입자를 걸러 먹는다.[24] 이러한 촉수관은 포로니다(Phoronida)나 완족류(Brachiopoda)와 같은 다른 촉수동물에서도 발견되지만, 태형동물은 대부분 무성생식으로 증식한 클론 개충들이 모여 군체를 형성한다는 점에서 차이가 있다.[16][24]
소화관은 입에서 시작해 항문으로 이어지는 U자 형태이며, 항문은 촉수관 바깥쪽에 위치한다. 몸 크기가 매우 작기 때문에 별도의 순환계(혈관계)나 호흡계, 배설계(신관)는 가지고 있지 않으며, 가스 교환 등은 몸 표면 전체, 특히 촉수관을 통해 이루어진다.[24][34] 간단한 신경계와 몸의 형태를 지지하는 정수압 골격계는 존재한다.[34]
군체는 원칙적으로 자웅동체이며, 대부분 바위, 해조류, 조개껍데기 등 다른 물체 표면에 붙어서 생활한다. 전 세계적으로 약 5,000종 이상이 알려져 있으며, 한국 연안에서도 120여 종이 보고되었다.
일부 종은 생물광물화된 단단한 외골격을 만들어 작은 산호와 비슷하게 보이기도 한다. 그러나 태형동물 군체는 일반 개충과 다른 둥근 모양의 조상충(ancestrula)이라는 첫 개충에서 시작하며, 각 개충이 체강과 촉수관을 가진다는 점에서 폴립에 체강이나 촉수관이 없는 산호와 근본적으로 다르다.[34] 또한, 내새류(Entoprocta)도 촉수관과 유사한 먹이 섭취 기관을 가지지만, 내새류의 촉수는 속이 비어있지 않고 단단하며, 항문이 촉수관 안쪽에 열리고 체강이 없다는 점에서 태형동물과 구별된다.[19]
일부 태형동물 종은 인간에게 유용한 물질을 생산하기도 한다. 예를 들어, ''Bugula neritina''라는 종은 세포독성 화학물질인 브리오스타틴(bryostatin)을 생산하는데, 이는 항암제로서의 가능성 때문에 임상 연구가 진행 중이다.
2. 1. 개충의 구조
태형동물 군체를 이루는 기본 단위는 개충(個蟲, zooid)이라고 불리는 매우 작은 개체이다. 개충 하나의 크기는 보통 0.5mm에서 1mm 정도이며,[24] 완전히 독립적인 생활을 하지 않기 때문에 '개체' 대신 '개충'이라는 용어를 사용한다.[24] 이 개충들이 무성생식을 통해 클론으로 증식하여 모여 나뭇가지, 부채, 덩어리 등 다양한 형태의 군체를 형성한다.[24][34] 군체 내의 개충들은 유전적으로 동일하며, 마치 하나의 큰 동물을 이루는 기관처럼 서로 협력한다.[24]모든 군체에는 영양 섭취를 담당하는 자충체(autozooid)가 있으며, 일부 종에서는 방어, 생식, 청소 등 특수한 기능을 수행하는 이충체(heterozooid)가 분화되기도 한다.[23] 어떤 종류의 개충이 군체 내 특정 위치에서 발달할지는 군체 전체의 화학적 신호나 외부 환경 요인(포식자, 경쟁 군체 등)에 따라 결정된다.[23]

모든 개충은 크게 두 부분으로 나눌 수 있다.
1. 낭체(囊體, cystid): 개충의 몸 바깥 부분을 이루는 구조물이다.
- 체벽: 가장 바깥쪽의 표피, 그 아래의 기저막, 결합조직, 근육층으로 구성된다.[24]
- 외골격(蟲室, zooecium): 체벽의 표피에서 분비되어 만들어지는 딱딱한 집이다. 성분은 키틴이나 단백질 같은 유기물이거나, 탄산칼슘과 같은 광물질일 수 있다.[26] 광물질 외골격은 주로 해양 태형동물에서 발견되며, 담수 종에서는 보통 유기물로만 이루어진다.[26] 외골격은 군체 전체의 형태(둔덕 모양, 부채꼴, 가지 모양 등)를 만들며, 표면의 작은 구멍 하나하나가 개충이 사는 공간이 된다.[34] 결정학적 분석에 따르면, 광물질 골격은 방해석이나 아라고나이트 결정이 유기 기질 내에 복잡하게 배열된 구조를 보인다.[20]
- 체강: 체벽 안쪽의 공간으로, 중피라는 조직으로 둘러싸여 있다.[24] 일부 분류군에서는 중피가 두 층으로 나뉘어, 안쪽 층이 체강을 포함한 막 주머니를 형성하고 바깥쪽 층이 체벽에 붙어 가체강과 같은 구조를 만들기도 한다.[27]
2. 폴립체(polypide): 낭체 안에 들어있는 부드러운 몸체 부분이다. 필요에 따라 낭체 밖으로 나왔다가 안으로 완전히 수축될 수 있다.[24]
- 촉수관(觸手冠, lophophore): 입 주위를 둘러싼 촉수들의 모임으로, 태형동물의 가장 큰 특징 중 하나이다.[24] 촉수 표면에는 수많은 섬모가 나 있어, 물의 흐름을 만들어 식물성 플랑크톤과 같은 작은 먹이 입자를 걸러 입으로 보낸다.[24] 촉수관의 형태는 분류군에 따라 다른데, 해산 종이 많은 나후강(Gymnolaemata)과 협후강(Stenolaemata)에서는 주로 촉수가 원형으로 배열되고, 담수 종인 은후강(Phylactolaemata)에서는 말굽 모양(U자형)으로 배열되는 경우가 많다 (단, ''Fredericella'' 속은 예외적으로 원형).[19] 원형 촉수관은 촉수가 모인 중심에 입이 열리지만, U자형 촉수관은 U자의 굽은 부분 바닥에 입이 열린다.[19]
- 소화계: 입에서 시작하여 인두, 식도, 위 (분문부, 맹낭, 유문부로 구성), 장, 직장을 거쳐 항문으로 이어지는 U자형 관이다.[24] 항문은 촉수관의 바로 바깥쪽에 열린다 (은후강에서는 U자형 촉수관 바닥의 입 근처).[19] 일부 종에서는 위 앞에 모래주머니와 같은 구조가 있어 먹이를 잘게 부수는 것을 돕는다.[24]
- 신경계 및 근육: 폴립체를 움직이고 외부 자극에 반응하는 신경 조직과 근육들이 포함되어 있다.[24]
- 퇴화와 재생: 폴립체는 주기적으로 퇴화하여 갈색체(brown body)라는 덩어리로 변했다가 다시 재생성되는 과정을 반복하기도 한다. 특히 군체에서 오래된 부분의 개충들은 폴립체가 퇴화하고 외골격 입구가 격막으로 막힌 휴면 상태로 남아있는 경우가 많다.[24]

개충은 크기가 매우 작기 때문에 별도의 순환계(혈관계)나 호흡계(아가미 등)가 없다.[24] 필요한 산소 공급과 이산화탄소 배출 같은 가스 교환은 몸 표면 전체, 특히 촉수관의 넓은 표면적을 통해 직접 이루어진다.[24] 노폐물 배설을 위한 신관과 같은 전문적인 배설 기관도 없다.[24] 하지만 간단한 신경계와 몸의 형태를 유지하는 골격계(외골격 및 정수압 골격)는 가지고 있다.[34]
2. 2. 촉수관
태형동물의 가장 일반적인 개체 유형인 자가영양충(autozooid)은 먹이를 섭취하는 역할을 담당하며, 속이 빈 촉수가 모여 만들어진 '관(crown)' 형태의 기관인 촉수관(lophophore)을 가지고 물속의 먹이 입자를 걸러낸다.[23] 촉수관은 태형동물, 포로니다, 완족류가 공유하는 특징적인 기관이기도 하다.[16] 모든 군체에서 상당수의 개충이 자가영양충이며, 일부 군체는 전적으로 자가영양충으로만 구성되기도 한다. 이들 중 일부는 생식에도 관여한다.[28]촉수관의 촉수 표면에는 섬모(cilium)라고 하는 미세한 털들이 나 있다. 이 섬모들의 규칙적인 운동은 물의 흐름을 만들어내는데, 물은 촉수의 끝부분에서 안쪽으로 흘러들어와 촉수관의 기저부(밑부분)를 통해 밖으로 빠져나간다.[16][24] 이 물의 흐름에 섞여 들어온 식물성 플랑크톤, 규조류 같은 작은 미생물이나 유기물 입자가 촉수와 부딪히면, 촉수 표면의 점액에 붙잡히고, 촉수 안쪽 면의 다른 섬모 작용으로 입으로 운반된다.[16][24] "상류 수집(upstream collecting)"이라고 불리는 이 섭식 방식은 먹이 입자가 물 흐름을 만드는 섬모 영역을 통과하기 전에 포획하는 것이 특징이며, 포로니다, 완족류, 익새류에서도 유사하게 나타난다.[29]
촉수관의 기본적인 형태는 촉수가 완전한 원 모양으로 배열된 것이다. 이는 주로 바다에 서식하는 나후강(Gymnolaemata)과 협후강(Stenolaemata)에서 관찰된다.[24] 반면, 담수에 서식하는 피락토라에마타(Phylactolaemata)에서는 촉수관이 말굽(U자) 모양으로 깊게 패인 형태를 보인다. 이는 촉수 배열 자체의 틈이 아니라 관 가장자리가 깊게 패여 U자처럼 보이는 것이다.[24] 예외적으로 피락토라에마타에 속하는 ''Fredericella'' 속은 원형의 촉수관을 가진다. 원형 촉수관의 경우, 촉수가 모이는 중심부에 바로 입이 열려 있다. U자형 촉수관에서는 U자의 굽어진 바닥 부분에 입이 위치한다.[24]
"내측(invert)"이라 불리는 유연한 관 구조 위에 촉수관과 입이 있으며, 이는 신축성이 있어 몸 안으로 완전히 수축될 수 있다.[24] 촉수관이 수축되면 내측 안에 접혀서 보호받게 된다. 이 수축은 한 쌍의 수축근에 의해 일어나며, 때로는 60밀리초 이내의 매우 빠른 속도로 이루어진다.[27] 촉수 끝에는 감각 수용체가 있어 위험을 감지하면 촉수관이 완전히 펴지기 전에 빠르게 수축할 수 있다. 반대로 촉수관을 펼 때는 내부 체액 압력을 높이는 방식을 사용한다. 유연한 외골격을 가진 종은 체벽 안쪽 원형 근육을 수축시키고, 막성 주머니를 가진 종은 이 주머니를 압축하여 압력을 높인다.[27] 단단한 외골격을 가진 일부 종은 외골격 일부를 대체하는 유연한 막을 가지며, 외골격 안쪽 가로 근육이 이 막을 당겨 체액 압력을 높인다.[24] 또 다른 종들은 단단한 골격에 틈이 없는 대신, 작은 구멍으로 외부 물과 연결된 유연한 주머니를 이용한다. 가로 근육이 이 주머니를 당기면 내부 체액 압력이 증가하여 내측과 촉수관을 밖으로 밀어낸다.[24] 일부 종은 수축된 내측과 촉수관을 보호하기 위해 근육으로 닫고 체액 압력으로 여는 덮개인 조개판(operculum)을 가진다. 특정 강(class)에서는 입 위를 "입술판(epistome)"이라는 속이 빈 돌기가 덮기도 한다.[24]
U자 모양의 소화관은 촉수관 중앙의 입에서 시작하여 몸 안쪽으로 내려갔다가 다시 위로 올라와 항문으로 이어진다. 항문은 보통 촉수관 바깥쪽 아래, 내측(invert) 위에 위치하며,[24] 피락토라에마타에서는 U자형 촉수관 바닥의 입 근처에 열린다. 인두 주변에는 신경 고리가 있고, 그 한쪽에는 뇌 역할을 하는 신경절이 있다. 신경은 이 고리와 신경절에서 촉수를 포함한 몸의 다른 부위로 뻗어 나간다.[24] 특별한 감각 기관은 없지만, 촉수의 섬모가 감각 수용체 역할을 하는 것으로 보인다. 예를 들어, ''부굴라''(Bugula) 속 군체는 빛을 향해 자라는 경향이 있어 빛 감지 능력이 있음을 시사한다.[24] 일부 종의 군체에서는 체벽 구멍을 통해 신경 신호가 개체 간에 전달되어 먹이 섭취나 촉수관 수축 같은 활동을 조절하기도 한다.[24]
크기가 매우 작아 별도의 혈액계나 호흡계는 없다. 기체 교환은 몸 전체 표면, 특히 촉수관의 촉수를 통해 이루어진다.
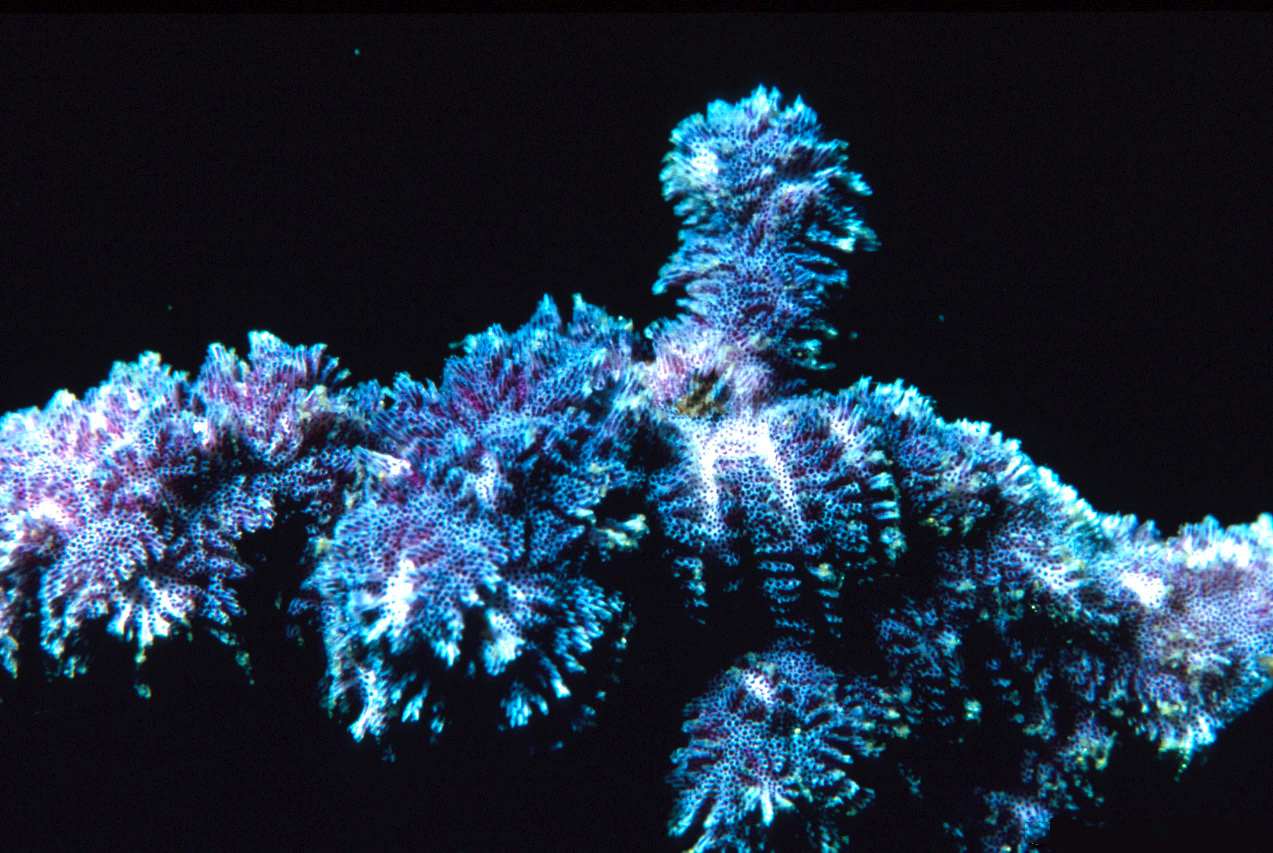
2. 3. 군체 형태
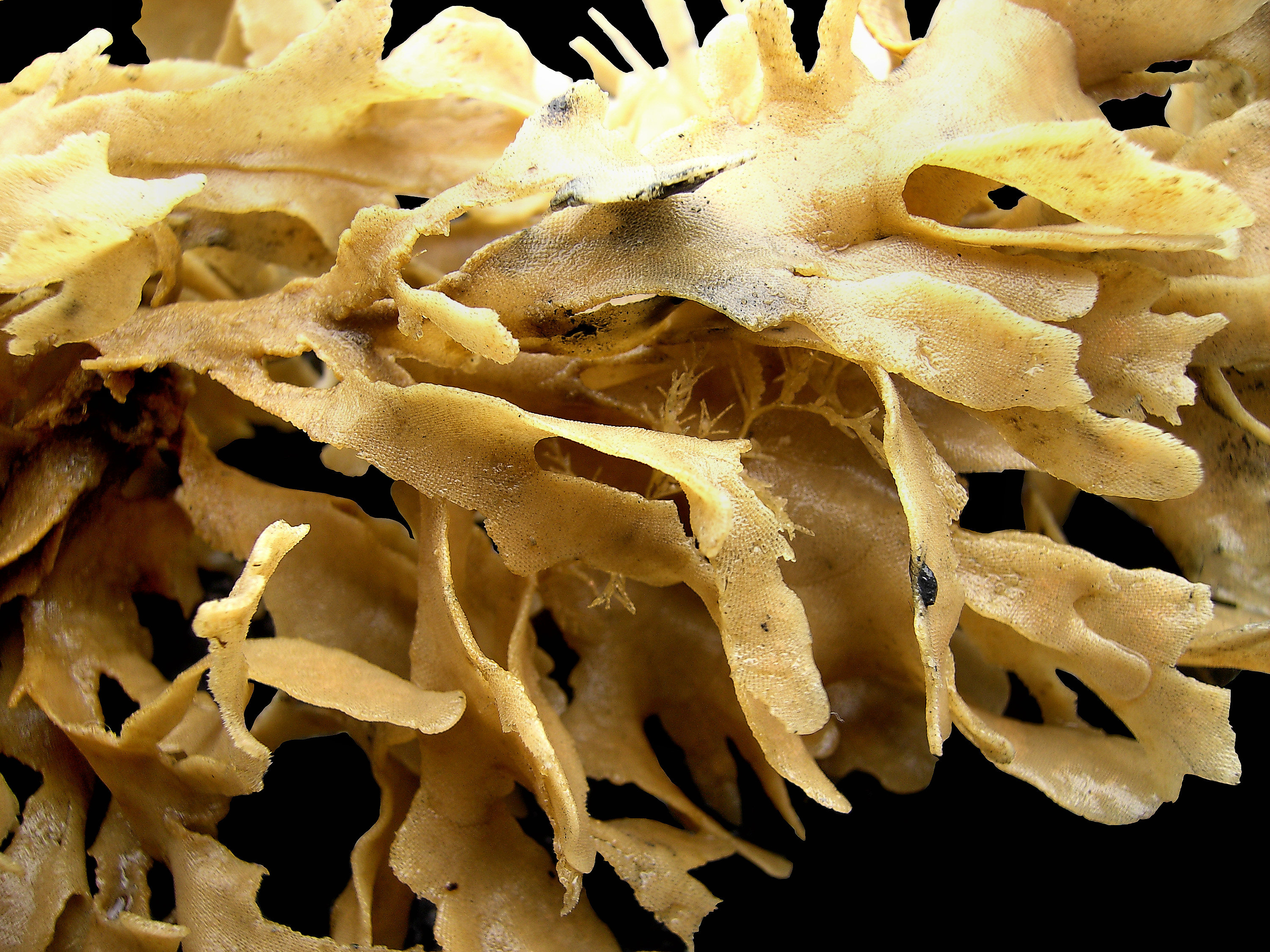

개개의 개충(zooid)은 몸길이가 1mm 안팎으로 현미경적 크기이지만, 이들이 모여 이루는 군체의 크기는 1cm에서 1m가 넘는 범위까지 다양하다.[24] 그러나 대부분은 10cm 미만이다.[34] 군체의 형태는 매우 다양하며, 성장 방식인 출아 패턴, 존재하는 개충의 다양성, 그리고 분비하는 골격 물질의 종류와 양에 따라 달라진다.[24] 각각의 개충은 주로 키틴질로 된 외골격(蟲室)과 그 속의 충체(蟲體)로 이루어져 있다.
일부 해양 종은 뿌리와 가지를 형성하는 헛개충에 의해 지지되는 관목형 또는 부채꼴 형태를 띤다. 이러한 형태의 군체는 일반적으로 생광물화되지 않지만, 키틴으로 이루어진 외골격을 가질 수 있다.[24] 다른 종들은 작은 산호처럼 보이며, 무거운 석회질 골격을 생성한다.[35] 많은 종들이 여러 개의 개충으로 구성된 군체를 형성하는데, 이는 잎, 뭉치 또는 속 ''Thalamoporella''의 경우 상추의 열린 머리와 유사한 구조를 형성할 수 있다.[24]
그러나 가장 흔한 해양 형태는 피복형으로, 한 층의 개충이 단단한 표면이나 해조류 위에 퍼져 자란다. 일부 피복형 군체는 50cm가 넘게 자라며, 약 2,000,000개의 개충을 포함할 수 있다.[24] 이러한 종들은 일반적으로 탄산칼슘으로 강화된 외골격을 가지고 있으며, 촉수관이 돌출되는 구멍은 위쪽 또는 바깥쪽 표면에 있다.[24] 피복형 군체의 이끼와 같은 외형은 태형동물이라는 이름(고대 그리스어 단어 βρύονgrc는 '이끼'를, ζῷονgrc는 '동물'을 의미함)의 유래가 된다.[36] 대형 피복종 군체는 종종 "굴뚝"이라고 하는 촉수관 캐노피의 틈이 있는데, 이를 통해 걸러진 물을 빠르게 배출하여 이미 여과된 물을 다시 여과하는 것을 피한다.[37] 이것은 먹이를 먹지 않는 잡개충의 무리에 의해 형성된다.[38] 새로운 굴뚝은 확장하는 군체의 가장자리 근처, 유출 속도가 이미 빠른 지점에 나타나며, 물 흐름이 변해도 위치가 바뀌지 않는다.[39]
일부 담수종은 최대 1m 지름의 젤라틴 물질 덩어리를 분비하고, 개충이 이에 부착된다. 다른 담수종은 "줄기"와 "가지"를 가진 식물과 같은 형태를 가지며, 곧게 서 있거나 표면에 퍼져 있을 수 있다. 일부 종은 하루에 약 2cm 정도 이동할 수 있다.[24]
각 군체는 조상충(ancestrula)이라고 알려진 단일 개충에서 무성 출아에 의해 자란다.[24] 조상충은 일반적인 개충과 달리 둥글다.[34] 군체 성장은 이러한 구조를 가진 형태의 "줄기" 또는 "가지" 끝에서 일어나며, 피복형 군체는 가장자리를 따라 자란다. 석회질 외골격을 가진 종의 경우, 개충이 완전히 자랄 때까지 무기질화되지 않는다. 군체의 수명은 1년에서 약 12년까지 다양하며, 수명이 짧은 종은 한 계절에 여러 세대를 거친다.[24]
방어 개충을 생성하는 종은 위협이 이미 나타난 경우에만 48시간 이내에 방어 개충을 만든다.[23] "유도 방어" 이론은 방어 생성이 비용이 많이 들기 때문에, 너무 일찍 또는 과도하게 방어하는 군체는 성장률과 수명이 감소할 것이라고 설명한다. 이러한 "막바지" 방어 접근 방식은 단일 공격으로 인한 개충 손실이 크지 않기 때문에 가능하다.[23] 일부 피복종의 군체는 다른 피복 유기체, 특히 다른 태형동물의 확장을 제한하기 위해 특수한 잡개충을 생성하기도 한다. 어떤 경우에는 상대 군체가 작을수록 이 반응이 더 공격적인데, 이는 군체 가장자리의 개충이 상대의 크기를 감지할 수 있음을 시사한다. 일부 종은 특정 다른 종에 대해 지속적으로 우세하지만, 대부분의 영역 다툼은 결정적이지 않으며, 경쟁자들은 곧 경쟁이 없는 지역에서 자라기 시작한다.[23] 영역을 놓고 경쟁하는 태형동물은 해면이나 산호가 사용하는 정교한 기술을 사용하지 않는데, 아마도 태형동물의 수명이 짧아 영역 다툼에 많은 투자를 하는 것이 비효율적이기 때문일 것이다.[23]
태형동물은 오르도비스기 이후로 해양 생물의 탄산염 퇴적에 기여해 왔다. 태형동물은 다양한 분류군에서 진화했으며, 퇴적물 생성 능력이 다양한 여러 군체 형태를 보인다. 9가지 기본적인 태형동물 군체 형태는 다음과 같다.[40]
| 형태 | 설명 |
|---|---|
| 피복형 | 단단한 표면 위에 한 층으로 퍼져 자람 |
| 돔형 | 반구형 또는 덩어리 형태 |
| 손바닥형 | 넓고 평평하며 손바닥 모양 |
| 엽상형 | 잎사귀 모양 |
| 창살형 | 격자 또는 그물 모양 |
| 굵은 가지형 | 굵은 가지 모양으로 분지 |
| 가는 가지형 | 가는 가지 모양으로 분지 |
| 연결형 | 여러 부분이 연결된 형태 |
| 자유생활형 | 고착하지 않고 자유롭게 움직이는 형태 |
이러한 퇴적물의 대부분은 돔형, 가는 가지형, 굵은 가지형, 손바닥형 군체 그룹과 창살형 군체 그룹에서 유래한다. 창살형 군체는 퇴적물과 스트로마토포로이드 산호초의 구성 요소 모두로 거친 입자를 생성한다. 반면, 가는 가지형 군체는 거친 퇴적물을 생성하고 심해 광선이 미치지 않는 생물 기원 언덕의 핵을 형성한다. 고생대 이후의 태형동물 퇴적물은 자유생활형 군체 외에도 다양한 성장 형태를 포함하며, 입자 크기가 진흙에서 모래, 자갈로 다양하게 변하는 퇴적물을 생성했다.[40]
외항동물의 군체는 산형, 부채형, 작은 가지형 등 다양한 형태를 취한다. 외벽에는 작은 구멍이 무수히 뚫려 있으며, 각각은 개충이라고 불리는 개체이다. 이들은 입에서 항문까지 이어지는 소화관으로 이루어진 진체강 구조를 가지고 있다. 입 주변의 촉수에는 섬모가 나 있는 총담(lophophore)이라고 불리는 구조가 있으며, 규조나 조류를 포함한 미생물을 잡아 먹이로 한다. 이 섬모가 난 촉수들이 입을 둘러싸 배열되어 전체적으로 촉수관을 이룬다. 촉수관은 완전히 몸 안으로 끌어들일 수 있다.
2. 4. 다형성
모든 태형동물 군체는 단 하나의 속인 ''Monobryozoon''을 제외하고는 군체 생활을 한다.[22][23] 군체를 이루는 개별 구성원은 개충(zooideng)이라고 불리며, 크기는 보통 약 0.5mm 정도이다.[24] 개충은 무성생식으로 만들어진 클론이므로 유전적으로 동일하며,[121] 완전히 독립적인 동물이 아니라 마치 더 큰 동물의 기관처럼 서로 협력한다.[24]모든 군체에는 영양 섭취를 담당하는 자충체(autozooideng)가 있다. 일부 그룹에서는 자충체 외에 영양 섭취 외의 특수한 기능을 수행하는 다양한 형태의 이충체(heterozooideng)가 나타나는데, 이를 다형성(polymorphism)이라고 한다.[23] 어떤 종류의 개충이 군체의 특정 위치에서 자랄지는 군체 전체의 화학적 신호나 포식자, 경쟁 군체의 존재 여부에 따라 결정될 수 있다.[23]
모든 개충은 기본적으로 두 부분으로 구성된다. 몸통 벽과 외골격으로 이루어진 낭체(cystideng)와, 그 안에 들어있는 신경계, 소화계, 근육, 촉수관 등을 포함하는 폴립체(polypideeng)이다.[24] 외골격은 키틴, 다당류, 단백질 같은 유기물이거나 탄산칼슘 같은 광물질일 수 있다.[26] 이충체는 이러한 기본 구조가 변형된 형태이다.
주요 이충체의 종류와 기능은 다음과 같다.
| 이충체 종류 | 어원/원어 표기 | 주요 기능 | 비고 (형태 등) |
|---|---|---|---|
| Aviculariaeng (작은 새) | 방어 (포식자 퇴치, 침입자 제거), 청소 | 새 부리 모양 또는 상자 모양. 덮개(아래턱)가 움직여 물체를 잡음. 일부는 자루(줄기)에 달림.[24][28] 찰스 다윈은 "목에 앉아 움직일 수 있는 작은 독수리의 머리와 부리" 같다고 묘사했다.[24][28] | |||
| Vibraculaeng | 방어, 청소, 이동 (일부 종) | 길고 움직이는 털 또는 채찍 모양의 돌기.[24][28] | |||
| κενός|케노스grc 빈 | 군체 지지, 줄기 형성, 간격 유지, 영양 저장 | 폴립체가 퇴화하고 낭체만 남은 형태. 군체의 구조를 형성하고 강화함.[24][23][28] 일부는 영양 저장 기능 추정. | |||
| Mesozooideng | 자충체 간 간격 유지 (주로 멸종한 해면벌레류) | 자충체 사이의 작은 방 형태. 격막으로 나뉨. | |||
| Gonozooideng | 생식 (알 육아) | 수정된 알을 품는 육아낭 역할. 주로 관산호류에서 발견. 유생이 나가는 난공(ooecioporeeng)이 있음. 단일배 발생(monozygotic polyembryonyeng)을 통해 여러 유생을 만듦. 아르마딜로도 사용하는 생식 전략. | |||
| Ovicelleng | 생식 (배아 육아) | 주로 틈새산호류에서 발견. 자충체에 부착된 주머니 형태로 배아를 보호. 난실을 가진 자충은 먹이 활동을 계속하므로 엄밀히 이형충으로 보지 않기도 함. | |||
| Androzooideng | 생식 (정자 생성 및 전달) | "수컷" 다형체. 특정 종(예: 게 껍질에 공생하는 Odontoporella bishopi)에서 발견. 자충체보다 작고 촉수 수가 다름. 이동성을 가지기도 함. 다른 군체와 만나 정자 교환에 사용될 가능성 있음. | |||
| Spinozooideng | 방어 추정 | 속이 빈 움직이는 가시 모양. 군체 표면에 존재. | |||
| Nanozooideng | 불명확 | 작은 단일 촉수 폴립을 가진 소형 개충. 다른 개충 위나 퇴화된 개충 내에서 자람.[28] |
태형동물(Bryozoa)은 포로니다(Phoronida), 완족류(Brachiopoda)와 함께 촉수관이라는 속이 빈 촉수 다발을 이용해 물에서 먹이를 걸러 먹는 촉수동물이다.[16] 태형동물은 대부분 군체를 이루어 생활하며, 군체를 구성하는 각 개체는 개충(zooid)이라고 불린다. 개충은 무성생식으로 증식된 클론이며, 크기는 보통 약 0.5mm 정도이다.[24] 이는 비슷한 구조를 가졌지만 군체를 형성하지 않는 포로니다나[17] 이매패류와 유사한 껍데기를 가진 완족류와 구별되는 특징이다.[18] 작은 산호 군체와 비슷하게 보일 수 있지만, 태형동물 군체는 일반적으로 둥근 형태의 조상충(ancestrula)이라는 첫 개체에서 시작하며, 이후 성장하는 개충들과는 다른 모양을 가진다. 반면 산호의 첫 개체는 이후 개체들과 같은 모양이며, 체강이나 촉수관이 없다.[34] 또한 내새류도 여과 섭식을 하지만, 촉수가 단단하고 항문이 촉수관 안쪽에 위치하며 체강이 없다는 점에서 태형동물과 다르다.[19]
군체 내의 개충들은 모두 무성생식으로 만들어져 유전적으로 동일하다. 따라서 생식 능력이 없는 이충체라도 다른 개충(특히 생식 개충)의 생존과 번식을 도움으로써 자신의 유전자를 다음 세대에 전달하는 데 기여할 수 있다. 이러한 개충의 기능 분화는 사회성 곤충의 카스트 분화처럼 혈연 선택 이론으로 설명할 수 있다.[121]
3. 생태
단 하나의 속인 ''Monobryozoon''을 제외한 모든 태형동물은 군체 생활을 한다.[22][23] 군체를 이루는 개충들은 완전히 독립적인 동물이 아니며,[24] 유전적으로 동일하여 마치 더 큰 동물의 기관처럼 서로 협력한다.[24] 모든 군체에는 먹이 섭취를 담당하는 자충체(autozooid)가 있으며, 일부 종에서는 먹이 섭취 외에 방어, 생식, 지지 등 다양한 특수 기능을 수행하는 이충체(heterozooid)가 존재한다.[23] 어떤 종류의 개충이 군체의 어느 위치에서 자랄지는 군체 전체의 화학적 신호나 포식자, 경쟁 군체의 존재 여부에 따라 결정되기도 한다.[23] 예를 들어, 방어 기능을 하는 개충(avicularia, vibracula, spinozooids 등)은 위협이 감지될 때만 생성되는데, 이는 방어 구조를 만드는 데 드는 비용을 절약하기 위한 "유도 방어" 전략으로 해석된다. 이러한 빠른 대응(48시간 이내 생성 가능)은 단일 공격으로 인한 손실이 군체 전체에 치명적이지 않기 때문에 가능하다.[23]
개충은 현미경 수준의 크기지만, 군체 전체의 크기는 수 밀리미터에서 1m 이상까지 매우 다양하다.[24] 군체의 형태 또한 매우 다양한데, 이는 개충의 증식(출아) 방식, 존재하는 개충의 종류, 분비하는 골격 물질의 종류와 양에 따라 달라진다.[24] 해양에서는 단단한 표면을 덮는 피복형이 가장 흔하며,[24] 이끼와 같은 모습 때문에 태형동물이라는 이름(그리스어로 '이끼 동물'이라는 뜻)이 유래했다.[36] 그 외에도 뿌리와 가지를 형성하는 관목형, 부채꼴형, 작은 산호와 유사한 형태 등 다양하다.[24][35] 담수 종 중에는 젤라틴 덩어리를 분비하거나 식물처럼 가지를 뻗는 형태도 있다.[24]
각 군체는 조상충(ancestrula)이라는 단일 개충이 무성생식의 일종인 출아를 통해 성장하여 형성된다.[24] 군체의 수명은 종에 따라 1년에서 약 12년까지 다양하다.[24] 일부 피복형 군체는 다른 피복성 생물, 특히 다른 태형동물의 성장을 막기 위해 특수한 개충을 생성하며 영역 다툼을 벌이기도 한다. 경쟁의 결과는 종종 결정적이지 않으며, 경쟁자들은 곧 비경쟁 지역으로 성장을 이어간다.[23] 태형동물은 해면이나 산호처럼 정교한 경쟁 기술을 사용하지 않는데, 이는 상대적으로 짧은 수명 때문에 영역 다툼에 많은 에너지를 투자하는 것이 비효율적이기 때문일 수 있다.[23]
태형동물은 오르도비스기 이후 해양 생태계에서 탄산칼슘 퇴적물 형성에 중요한 역할을 해왔다. 다양한 군체 형태(피복형, 돔형, 가지형, 창살형 등)는 각기 다른 종류와 크기의 퇴적물을 생성하여 해저 지형 형성에 기여해왔다.[40]
3. 1. 서식지
대부분의 태형동물은 바다에 서식하지만, 담수에 사는 종도 약 70종 알려져 있다.[61]
해양 서식지해양 태형동물은 주로 수심 100m 미만의 열대 해역에 서식한다.[94] 그러나 일부 종은 냉수순환류 주변의 깊은 해구[94]나 극지방 근처에서도 발견된다.[95][96] 대부분 부착성으로, 암석, 모래, 조개껍질을 포함한 단단한 기질에 붙어 산다.[97] 얕은 바다에서는 표면을 덮는 피막형 군체가 흔하지만, 수심이 깊어질수록 곧게 서는 직립형 군체가 더 많아진다.[95] 일본 북부의 플라이스토세 지층에서는 자갈이나 조약돌 하나에 20종 이상의 태형동물이 붙어 있는 화석이 발견되기도 했다. 모래나 실트처럼 입자가 작은 퇴적물은 일반적으로 태형동물이 살기 어렵지만, 거친 모래 알갱이에 부착된 작은 군체가 발견된 사례도 있다. 일부 종은 해조류, 잘피, 심지어 맹그로브 뿌리에 서식하는 데 특화되어 있다. 예를 들어, ''Amphibiobeania'' 속은 맹그로브 잎에 살면서 썰물 때 공기에 노출되어도 생존할 수 있어 "양서류"라는 이름이 붙었다.
기질에 부착하지 않고 자유롭게 생활하는 "자유생활성" 태형동물도 다양하게 존재한다. ''Cristatella'' 같은 일부 담수 종은 이동이 가능하다. 해양에서는 초승달 모양의 측판류(Lunulite)가 기질에 부착되지 않은 채 이동하는 군체를 형성한다. ''Selenaria'' 속의 군체는 강모(setae)라는 털 같은 구조를 이용해 움직이는 것이 관찰되었으며, 수족관에서는 시간당 1m 속도로 이동하고, 빛을 향해 움직이며, 뒤집혔을 때 스스로 제자리로 돌아오는 모습이 기록되었다. Cupuladriidae 과의 태형동물도 비슷한 형태와 이동성을 독립적으로 진화시켰다. 이러한 이동 능력은 원래 퇴적물 속에서 몸을 파내기 위해 진화시킨 긴 강모의 부수적인 결과일 수 있다는 가설이 있다.
다른 자유생활성 태형동물은 파도나 해류에 의해 수동적으로 이동한다. 남극해에 서식하는 ''Alcyonidium pelagosphaera''는 속이 빈 공 모양의 부유성 군체를 이루며 떠다닌다. 이 군체의 지름은 5mm에서 23mm 사이이며, 일생 동안 계속 떠다니는 것인지 아니면 특정 시기의 모습인지는 아직 불확실하다.[95][98] 북극의 진흙 바닥에 사는 원반 모양의 ''Alcyonidium disciforme''는 흡수한 모래 알갱이를 이용해 몸의 균형을 잡고 뒤집혔을 때 제자리로 돌아온다. 어떤 종들은 해저를 따라 구르면서 모든 방향으로 성장하는 구형 군체, 즉 브리올리스(bryoliths)를 형성하기도 한다.
최근 지구 온난화의 영향으로 남극 일부 지역에서는 빙산에 의한 해저 삭마가 증가했는데, 태형동물의 일종인 ''Fenestrulina rugula''가 이렇게 새로 생긴 공간을 빠르게 차지하며 우점종이 되었다는 보고가 있다.[99] 이는 환경 변화에 대한 태형동물의 적응 능력을 보여주는 사례이다.
담수 서식지내부강태형류는 호수, 연못, 강, 개울, 하구 등 모든 유형의 담수 환경에 서식하며,[61] 가장 풍부하게 발견되는 부착성 담수 동물 중 하나이다.[100] 일부 관태형류는 오직 담수에서만 살지만, 다른 종들은 염분이 약간 섞인 기수를 선호하면서도 담수 환경에서 생존할 수 있다.[61] 많은 지역의 담수 태형동물 분포에 대한 정보는 아직 부족하며, 과거에는 특정 담수 종이 전 세계적으로 분포한다고 여겨졌으나, 2002년 이후 연구를 통해 실제로는 더 좁은 지역에 제한적으로 분포하는 여러 종으로 나뉘어야 한다는 사실이 밝혀졌다.[61]
전반적으로 태형동물은 매우 다양한 수중 환경에 적응하여 서식하고 있으며, 대부분은 단단한 표면에 고착하여 살아가지만, 일부는 퇴적물 위에서 살거나 자유롭게 이동 또는 부유하며 생활하는 등 독특한 생존 방식을 보여준다. 서식 가능한 수심 범위도 매우 넓어, 얕은 연안에서부터 최대 8200m의 심해까지 분포한다.
3. 2. 섭식
태형동물은 촉수관이라는 속이 빈 촉수 다발을 이용하여 물속의 먹이를 걸러 먹는 여과섭식 동물이다.[24][16] 대부분의 종은 주로 식물플랑크톤(미세한 부유 식물)과 같은 작은 입자를 먹이로 삼는다.[24] 예를 들어, 담수종인 ''Plumatella emarginata''는 규조류, 녹조류, 남세균, 비광합성 세균, 와편모조류, 윤충류, 원생동물, 작은 선형동물, 미세 갑각류 등을 먹는다.[107]
촉수관의 기본적인 형태는 원형이지만, 담수 태형동물(피락토라에마타)에서는 U자형으로 보이기도 한다.[24] 촉수 측면에는 섬모라는 미세한 구조가 있어, 이 섬모의 움직임으로 촉수 끝에서 밑부분을 향하는 물의 흐름을 만든다. 먹이 입자가 촉수에 닿으면 점액에 잡히고, 촉수 안쪽 면의 다른 섬모들이 입자를 중앙의 입으로 운반한다.[16] 이러한 방식은 "상류 수집" 또는 "상류 포획"이라고 불리며, 먹이 입자는 물 흐름을 만드는 섬모 영역을 통과하기 전에 포획된다. 이는 포로니다, 완족류, 익새류에서도 사용되는 방식이다.[29] 태형동물은 촉수를 이용해 더 큰 입자를 입 쪽으로 튕기거나, 일부 종은 촉수를 우리처럼 사용해 동물플랑크톤을 잡기도 한다. 또한, 촉수 표면의 미세융모를 통해 물에 녹아 있는 유기 화합물을 직접 흡수할 수도 있다.[24] 원치 않는 입자는 촉수로 튕겨내거나 입을 닫아 걸러낸다.[24] 연구에 따르면, 부착성 군체와 직립성 군체 모두 강한 물 흐름보다는 약한 흐름에서 더 빨리 먹이를 먹고 성장하는 경향이 있다.[83]
촉수관과 입은 "내번관(invert)"이라는 유연한 관 구조 끝에 달려 있는데, 이 내번관은 마치 고무장갑 손가락처럼 안팎으로 뒤집히면서 촉수관을 몸 안(개충벽(cystid))으로 빠르게 수축시킬 수 있다.[24] 수축은 한 쌍의 수축근에 의해 약 60밀리초 만에 이루어지기도 한다.[27] 촉수 끝의 감각 수용기는 위험을 감지하여 완전히 펴지기 전에 수축을 유발할 수 있다. 반대로 촉수관을 밖으로 내미는 팽창은 몸 내부 체액의 압력을 높여서 이루어진다. 외골격이 유연한 종은 체벽 안쪽의 근육을 수축시켜 압력을 만들고, 단단한 외골격을 가진 종은 외골격의 일부인 유연한 막을 근육으로 당기거나, 외부와 연결된 작은 주머니를 당겨 내부 압력을 높이는 등 다양한 방식으로 팽창시킨다.[24][27] 일부 종에서는 수축된 촉수관을 보호하기 위해 근육으로 여닫는 덮개인 조개판(operculum)을 가지고 있기도 하다.[24]
소화관은 U자 모양으로, 촉수관 중앙의 입에서 시작하여 몸 안쪽으로 내려갔다가 다시 위로 올라와 항문으로 이어진다. 항문은 보통 촉수관 바깥쪽 아래에 위치한다.[24] 일부 종의 위 첫 부분에는 키틴질 이빨이 있는 근육질의 모래주머니가 있어 규조류처럼 단단한 껍질을 가진 먹이를 부순다.[24] 음식물은 연동 운동을 통해 위에서 소화되고, 위의 마지막 부분에서는 섬모가 소화되지 않은 찌꺼기를 뭉쳐 장으로 보낸 뒤 항문을 통해 배출시킨다.[24] 태형동물은 신장과 같은 특별한 배설 기관이 없다.[23] 질소 노폐물인 암모니아는 체벽과 촉수관을 통해 물속으로 바로 확산시키는 것으로 추정된다.[24] 더 복잡한 노폐물은 배출되지 않고 폴립체(polypide, 개충의 내부 기관 부분) 내에 쌓인다. 폴립체는 주기적으로(몇 주마다) 퇴화하는데, 이때 노폐물이 축적된 죽은 세포 덩어리인 "갈색체(brown body)"가 형성된다. 이 갈색체는 새 폴립체가 만들어질 때 함께 배출되거나, 일부는 재활용되기도 한다.[24]
군체를 이루는 각 개충(zooid)들은 서로 연결되어 있어, 먹이를 섭취하는 자가개충(autozooid)이 다른 자가개충이나 먹이 섭취 기능이 없는 이형개충(heterozooid)에게 영양분을 나눠줄 수 있다.[24] 이 연결 방식은 태형동물의 강(class)에 따라 다른데, 체벽에 큰 구멍이 있어 체강액이 직접 통하거나, 소삭(funiculus)이라는 내부의 끈 구조가 작은 구멍을 통해 연결되어 영양분을 전달하기도 한다.[24][27]
3. 3. 생식
태형동물은 유성생식과 무성생식을 모두 한다.[23]
=== 유성생식 ===
대부분의 태형동물 개충(zooid)은 자웅동체이다. 모든 담수 종은 동시 자웅동체이며, 많은 해양 종은 순차적 자웅동체(먼저 수컷, 이후 암컷으로 기능)이지만 군체 전체적으로는 수컷과 암컷 단계의 개충이 섞여 있다.[23] 난소는 체벽 안쪽에, 정소는 위를 체벽에 연결하는 끈(funiculus)에 발달한다.[23]
난자와 정자는 체강으로 방출된다. 정자는 일부 촉수 끝의 구멍을 통해 물로 나가고, 이후 난자를 생산하는 개충의 섭식 수류에 의해 포획된다.[24] 수정 방식은 종에 따라 다르다. 어떤 종은 난자를 두 촉수 사이의 구멍(때로는 "촉수간 기관"이라는 작은 돌기 끝)을 통해 방출하여 외부에서 수정하고, 다른 종은 촉수간 기관이나 체강 내부에서 내부 수정을 한다.[24]
일부 종에서는 정자 생성에 특화된 수충(androzooids)이 발견된다. 예를 들어 게와 공생하는 ''Odontoporella bishopi'' 종의 수충은 다른 충체보다 작고 촉수 개수도 다르며(4개의 짧은 촉수와 4개의 긴 촉수), 이동성이 있어 군체 간 정자 교환에 관여할 수 있다.
=== 육아 및 유생 발생 ===
많은 태형동물 종은 배아를 군체 내에서 보호하고 키운다. 모든 담수 태형동물, 협구목, 대부분의 관구목은 태반과 유사한 구조를 통해 영양을 공급받는 비섭식성 유생(난황영양성 유생)을 갖는다.[85][86] 발생 중인 배아는 난황이나 모체로부터의 영양 공급(배외 영양), 또는 둘 다에 의존한다.[87]
육아 방식은 다양하다. 담수 태형동물은 내부 육아낭에서 배아를 품는다. 관구목은 외부 막낭, 골격으로 된 방(난실, ovicell), 내부 육아낭 등 다양한 구조를 이용한다.[85][86] 섬모구목에서는 어미 개체의 폴립체가 붕괴되어 배아에게 영양분을 제공한다. 협구목은 육아실 역할을 하는 특수한 개충(곤충충, gonozooid)을 생성하며, 이곳에서 난자가 분열하여 최대 100개의 동일한 배아(복제체)를 만드는 단일배 발생을 한다.[23] 틈새산호류(Cheilostome bryozoans)는 흔히 자충(autozooid)에 부착된 난낭(ovicells)에서 배아를 품는다.
섭식성 유생(플랑크톤영양성 유생)은 일부 관구목(Membraniporidae, Electridae 등)과 섬모구목(Alcyonidiidae, Farrellidae, Hislopiidae 등)에서만 발견된다.[88][89][90] 대표적인 예로 키포나우테스 유생이 있는데, 키틴질의 삼각형 껍질을 가지며 상당 기간 플랑크톤으로 살다가 정착한다.[23] 이 유생의 섬모는 성체와 유사하게 먹이를 포획한다.[91] 배아를 품는 종의 유생은 큰 난황으로 영양을 공급받고 내장이 없어 먹이를 먹지 않으며, 빠르게 표면에 정착한다.[24]
태형동물 알의 세포분열은 이방사상이며 초기 단계는 양측 대칭이다. 유생에서 성체로의 변태 시 내부 조직이 파괴되므로 체강 형성 과정은 불명확하다. 초기 배아의 원구는 닫히고 새로운 개구가 입이 된다.[24]
모든 태형동물 유생은 헤엄치기 위한 섬모띠, 꼭대기의 섬모 다발, 정착 시 사용되는 접착낭을 가진다.[24] 해양 종 유생은 정착 후 완전히 변태하는 고치를 생성하며, 유생의 표피는 체강 안감이 되고 내부 조직은 새로운 개체의 영양 저장고가 된다.[24] 담수 태형동물 유생은 여러 개의 폴립체를 생성하여 군체는 여러 개체로 시작한다.[24] 모든 종에서 처음 정착한 개충인 초충(ancestrula)은 출아를 통해 복제체를 만들어 군체를 성장시킨다.[24]
=== 무성생식 ===
태형동물은 출아를 통해 새로운 개충을 형성하여 군체를 성장시킨다. 군체 일부가 파괴되면 각 파편이 새로운 군체로 성장할 수도 있으며, 이렇게 만들어진 군체는 유전적으로 동일한 클론이다.
특히 담수 태형동물은 환경 변화에 적응하기 위해 정아(statoblast)라는 특수한 무성생식체를 만든다.[23] 정아는 해면의 아구(gemmule)와 유사한 원반 모양의 세포 덩어리로, 일종의 "생존 캡슐"이다.[24] 부모 개체로부터 영양을 공급받아[23] 키틴질의 보호 껍질을 발달시킨다. 성숙한 정아는 부모 군체에 붙어있거나(sessoblasts), 바닥에 가라앉거나, 공기 공간을 이용해 물에 뜨거나(floatoblasts),[24] 부모 군체가 죽은 후 재건을 위해 시스티드(cystid) 내에 남기도 한다.[23]
정아는 장기간 휴면하며 동결, 건조 등 극한 환경을 견딜 수 있다. 동물, 식물, 물살,[24] 바람[23] 등을 통해 장거리 이동도 가능하다.[92] 환경 조건이 좋아지면 껍질이 열리고 내부 세포가 새로운 개충으로 발달하여 군체를 형성한다. 예를 들어, ''Plumatella emarginata''는 서식지 유지를 위한 sessoblasts와 확산을 위한 floatoblasts를 모두 생산하며, ''Plumatella repens''는 성장 단계에 따라 다른 종류의 정아를 주로 생산한다.[107] 1m2 면적의 군체 무리가 800,000개의 정아를 생산할 수도 있다.[24]
피각목(Phylactolaemata)에서는 휴지아(hibernacula)라는 내구성이 강한 무성생식체를 만들어 겨울을 나거나 분포를 넓힌다.
컵룰라드리아과(Cupuladriidae) 태형동물은 유성생식으로 만들어진 군체가 무성생식(조각화)을 하거나, 무성생식으로 만들어진 군체가 유성생식을 하는 등 복잡한 생활사를 보인다.[93]
3. 4. 포식자와의 관계
해양 환경에서 태형동물은 다양한 포식자들의 먹이가 된다. 예를 들어, 레이스 모양의 해양 태형동물인 ''Membranipora membranacea''는 여러 종의 민달팽이 포식에 대응하여 방어용 가시를 만들기도 한다.[102] 그 외에도 어류, 성게, 바다거미, 갑각류, 진드기,[103] 불가사리 등이 태형동물을 포식한다.[104] 포식 방식은 포식자 그룹에 따라 다른데, 극피동물이나 연체동물은 군체의 일부를 통째로 뜯어 먹으며 광물화된 골격을 부수지만, 대부분의 절지동물 포식자는 개별 개체인 개충(zooid)을 먹는다.[105]
담수 환경에서도 태형동물은 달팽이, 곤충, 어류 등 다양한 포식자들의 먹이가 된다.[107] 특히 태국에서는 외래 침입종인 황금사과달팽이( ''Pomacea canaliculata'' )가 유입된 지역에서 담수 태형동물이 거의 사라지는 현상이 나타났다. 이 달팽이는 일반적인 담수 태형동물뿐만 아니라 섬모관태형동물도 먹지만, 후자에 대한 피해는 상대적으로 덜한 편이다. 반면, 태국의 토착 달팽이들은 태형동물을 먹이로 삼지 않는다.[108]
태형동물은 다른 생물과 다양한 상호작용을 한다. 히드라류나 Zancleidae과의 일부 종들은 태형동물과 함께 살아가는데, 이 관계는 서로에게 이익이 되는 공생이기도 하고 한쪽만 이득을 보는 기생 관계이기도 하다.[109] 남아프리카 연안의 태형동물 ''Alcyonidium nodosum''는 포식자인 바위가재 ''Jasus lalandii''로부터 고둥의 일종인 ''Burnupena papyracea''를 보호해준다. 태형동물이 고둥 껍데기를 덮으면 물리적으로 더 단단해지기도 하지만, 태형동물이 만드는 화학 물질이 포식자를 막는 데 더 중요한 역할을 하는 것으로 보인다.[110] 마우리타니아 연안에서는 태형동물 ''Acanthodesia commensale''가 특정 집게(''Pseudopagurus cf. granulimanus'')와 독특한 공생 관계를 맺는다. 이 태형동물은 집게가 사는 빈 고둥 껍데기 위에 자라면서 여러 겹의 골격을 만들어 알 크기만 한 둥근 공 모양의 구조물, 즉 브리올리스(bryolith)를 형성하고 집게의 생활 공간을 넓혀준다.[111]
한편, 일부 담수 태형동물은 믹소조아라는 기생충의 중간 숙주 역할을 한다. 이 기생충은 연어과 어류에게 치명적인 증식성 신장 질환(PKD)을 일으키는 원인으로 밝혀졌다.[112][61] 또한, 담수에 사는 일부 태형동물(굴족강)은 미포자충류(Microsporidia)에 속하는 부덴브로키아(''Buddenbrockia'') 같은 특수한 기생충에 감염되기도 한다.
일부 태형동물 종은 침입종으로 분류되어 기존 생태계에 영향을 미치기도 한다. 대표적인 예로 해양 태형동물인 ''Membranipora membranacea''는 1987년 북미 메인만에 처음 나타난 이후 빠르게 번식하여 켈프 숲을 뒤덮는 가장 흔한 생물이 되었다.[83] 이 태형동물이 켈프 잎을 약하게 만들어 부서지게 함으로써 켈프 군집이 크게 줄어들었다.[24] 그 결과, 켈프가 사라진 자리에는 또 다른 침입종인 해조류 ''Codium fragile'' 아종 ''tomentosoides''가 대신 번성하게 되었고, 이는 결국 지역 어류와 무척추동물의 서식지를 감소시키는 결과를 낳았다.[83] ''M. membranacea''는 미국의 북서부 연안에도 침입하여 비슷한 문제를 일으키고 있다.[24] 담수 태형동물 중 일부 종들도 원래 서식지에서 멀리 떨어진 곳에서 발견되는데, 휴면 상태의 스태토블라스트(statoblast)가 자연적으로 이동했을 수도 있지만, 수입된 수생 식물에 붙어오거나 선박 평형수 등을 통해 인간 활동에 의해 확산되었을 가능성이 더 큰 것으로 여겨진다.[92]
3. 5. 인간과의 관계
태형동물은 인간 활동에 다양한 영향을 미친다.
일부 해양 태형동물은 선박 선체, 부두, 선착장, 해양 구조물 표면에 달라붙어 생물 부착의 주요 원인이 된다. 이들은 새롭거나 깨끗하게 청소된 구조물에 가장 먼저 정착하는 생물 중 하나이다.[114] 정치망(生簀)과 같은 어업용 그물에 번식하여 그물눈을 막거나, 선저(船底)에 부착하여 선박의 속도를 늦추는 등 경제적 손실을 유발하기도 한다. 담수 종 역시 수도관, 정수 설비, 하수 처리 시설, 발전소의 냉각관 등에 부착하여 문제를 일으킬 수 있다.[61][115] 북해의 일부 어부들은 그물이나 통발에 붙은 태형동물과의 접촉으로 인해 "도거뱅크 가려움증"이라는 습진(피부 질환)을 앓기도 한다.[95][113] 일부 종은 독성을 지녀 어부에게 피부병을 유발하기도 한다.
특정 태형동물 종은 침입종으로 분류되어 생태계에 영향을 미치기도 한다. 예를 들어, 해양 태형동물인 ''Membranipora membranacea''는 1987년 메인만에서 처음 발견된 이후 빠르게 번식하여 켈프 군락을 뒤덮고 파괴했으며, 이는 지역 생태계의 변화를 초래했다.[83][24] 이 종은 미국 북서부 해안에도 침입했다.[24] 담수 태형동물 중 일부 종들은 원래 서식지에서 수천 킬로미터 떨어진 곳에서 발견되기도 하는데, 이는 수입된 수생 식물이나 선박 이동과 같은 인간 활동에 의해 확산되었을 가능성이 높다.[92] 담수산 큰공이끼벌레는 거대한 젤라틴질 군체를 형성하여 수중에 떠다니며 미관을 해치거나 수질 악화를 유발하는 경우도 있다.
일부 담수 태형동물 종은 연어과 어류에 치명적인 증식성 신장 질환을 일으키는 믹소조아 기생충의 중간 숙주 역할을 한다.[112] 이 질병은 유럽과 북미의 야생 어류 개체 수를 심각하게 감소시켰으며,[61] 양식장에서도 어류 폐사를 유발한다.[112]
한편, 태형동물은 의학 분야에서 주목받기도 한다. 해양 태형동물인 깃털이끼벌레(''Bugula neritina'')에서 추출되는 브리오스타틴이라는 화학 물질은 항암 효과[116] 및 알츠하이머병 치료[117][118] 가능성으로 연구되었다. 2001년 제약회사 GPC 바이오텍이 애리조나 주립대학교로부터 브리오스타틴 1의 라이선스를 취득하여 상업 개발을 시도했으나, 2003년 효과 부족과 독성 부작용을 이유로 개발을 중단했다.[116] 이후 알츠하이머병 치료 효과에 대한 임상 시험이 시도되었으나[117] 초기에는 어려움을 겪었다. 최근 연구에서는 알츠하이머병 환자의 인지 기능 개선 효과와 적은 부작용이 보고되었다.[118] 그러나 브리오스타틴은 1000kg의 태형동물에서 겨우 1g 정도만 추출될 정도로 양이 매우 적기 때문에,[119] 생산이 용이하고 효과가 유사한 합성 대체물 개발이 이루어졌다.[119]
4. 분류
태형동물문은 크게 다음 세 개의 강으로 나뉜다.[122]
태형동물은 다른 동물 문들과 다음과 같은 특징에서 구별된다.
| 태형동물[24](외항동물) | 다른 촉수동물[16] | 다른 후구동물 | 비슷하게 생긴 문 | |||
|---|---|---|---|---|---|---|
| 포로니다[17] | 완족류[18] | 환형동물, 연체동물 | 내항동물[19] | 산호류(자포동물 문의 강)[34] | ||
| 체강 | 세 부분, 만일 입술의 공간이 포함된다면 | 세 부분 | 기본 형태에서는 분절당 하나; 일부 분류군에서는 융합 | 없음 | ||
| 체강 형성 | 애벌레가 성체로 변태하기 때문에 추적이 불가능하여 불확실 | 장체강형성 | 열체강형성 | 해당 없음 | ||
| 촉수관 | 속이 빈 촉수가 있음 | 없음 | 비슷하게 생긴 먹이 섭취 구조, 하지만 단단한 촉수가 있음 | 없음 | ||
| 섭식 흐름 | 촉수 끝에서 밑으로 | 해당 없음 | 밑에서 끝으로 | 해당 없음 | ||
| 상피의 섬모 세포 | 있음[78] | 없음[78] | 있음[78] | 해당 없음 | ||
| 항문 위치 | 촉수관 기저부 바깥쪽 | 다양함, 일부 종에서는 없음 | 뒷끝, 하지만 시보글리니과에서는 없음 | 촉수관 비슷한 기관 기저부 안쪽 | 없음 | |
| 군체성 | 대부분 복제된 개체의 군체; 하나의 고독성 속 | 고착성 종은 종종 덩어리를 형성하지만, 능동적인 협력은 없음 | 일부 종에서는 복제된 개체의 군체; 일부 고독성 종 | 복제된 개체의 군체 | ||
| 창립자 개체의 모양 | 둥글며, 일반적인 개체와 다름[34] | 해당 없음 | 다른 개체와 같음 | |||
| 생물광물화된 외골격 | 일부 분류군 | 없음 | 이매패류와 같은 껍데기 | 일부 고착성 환형동물은 광물화된 관을 만듦;[20] 대부분의 연체동물은 껍데기를 가지지만, 대부분의 현대 두족류는 내부 껍데기 또는 없음.[21] | 없음 | 일부 분류군 |
태형동물(Bryozoa), 포로니다(Phoronida), 완족류(Brachiopoda)는 모두 촉수관(lophophore)이라는 속이 빈 촉수 다발을 이용해 물속의 먹이를 걸러 먹는다. 태형동물은 보통 약 0.5mm 길이의 개충(zooid)들이 무성생식으로 복제되어 군체를 이루는 반면,[24] 포로니다는 개충과 비슷하지만 크기가 2cm 에서 20cm 정도로 크고 군체를 이루지 않는다.[17] 완족류는 이매패류처럼 두 장의 껍데기를 가진다는 점에서 구별된다.[18] 이 세 문은 모두 체강을 가지고 있다.[24][17][18]
생물광물화된 외골격을 가진 일부 태형동물 군체는 작은 산호와 비슷하게 보일 수 있다. 그러나 태형동물 군체는 보통 둥근 모양의 특별한 창립 개충(ancestrula)에서 시작하는 반면, 산호 군체의 시작점인 폴립은 이후 만들어지는 폴립들과 모양이 같다. 또한 산호는 체강이나 촉수관이 없다.[34] 내항동물(Entoprocta) 역시 물을 걸러 먹는 동물이지만, 촉수관과 비슷한 기관의 촉수가 속이 차 있고, 항문이 촉수관 기저부 바깥쪽이 아닌 안쪽에 있으며, 체강이 없다는 점에서 태형동물과 다르다.[19]


태형동물 문은 원래 "Polyzoa"라고 불렸으나, 이후 에렌베르크가 제안한 "Bryozoa"라는 이름으로 대체되었다.[10][46][44] 처음 "Bryozoa"는 항문이 촉수관 바깥쪽에 있는 현재의 태형동물(Ectoprocta, ἐκτός|ektós|바깥grc + πρωκτός|prōktós|항문grc)만을 지칭했다.[41] 이후 항문이 촉수관 안쪽에 있는 내항동물(Entoprocta, ἐντός|entós|안grc + πρωκτός|prōktós|항문grc)[42]이 발견되면서 "Bryozoa"는 이 두 그룹을 포함하는 문(門)의 이름으로 사용되기도 했다.[43] 그러나 1869년 니체(Hinrich Nitsche)는 두 그룹 간의 뚜렷한 해부학적 차이(항문 위치, 촉수 구조, 체강 유무 등)를 근거로 기존의 "Bryozoa"에 해당하는 그룹을 "Ectoprocta"로 명명할 것을 제안했다.[74][72] 내항동물은 체강이 없고 나선분할을 하는 반면, 외항동물(현재의 태형동물)은 체강이 있고 방사분할을 한다는 중요한 차이가 있다. 분자 연구 결과도 이 둘의 가까운 관계를 지지하지 않아, 현재 내항동물은 약 150종을 포함하는 독립된 문으로 간주된다.[120][122]
이로 인해 현재는 두 그룹을 별개의 문으로 보는 것이 일반적이며, '태형동물(Bryozoa)'은 '외항동물(Ectoprocta)'과 사실상 동의어가 되었다.[43] 하지만 명칭 사용에 혼란이 있어왔기 때문에[45], 일부 학자들은 여전히 "Ectoprocta"라는 용어를 선호하기도 한다.[46][47] 일반적인 이름인 "이끼벌레"는 그리스어 βρυόν|bryón|이끼grc와 ζῷα|zōa|동물grc에서 유래했으며, 일부 종의 외형이 이끼와 비슷한 데서 비롯되었다.[50]
형태적으로 명명된 태형동물 종의 수는 4,000~4,500종으로 추산된다.[58] 특히 나후강의 입술입목은 다양한 종류의 특수화된 개충을 가져 가장 많은 종을 포함하는 것으로 여겨진다.[23]
과학자들은 태형동물(Ectoprocta)이 단계통군(단일 조상과 그 모든 후손으로 이루어진 그룹)인지, 동물 계통수에서 가장 가까운 친척이 누구인지, 심지어 원구동물인지 후구동물인지에 대해서도 의견이 분분하다. 분자 계통학 연구는 아직 "소규모 문"에 대한 데이터 부족으로 명확한 결론을 내리지 못하고 있다.[43][72] 현재 태형동물과 가장 가까운 관계는 완족동물로 보이며, 이 그룹의 자매군은 촉수동물일 가능성이 제기된다.
4. 1. 나후강 (Gymnolaemata)
태형동물문의 한 강으로, 주로 해양 환경에서 서식한다.[24][23] 입 위쪽에 입술 모양의 돌기(피후강의 특징)가 없으며, 촉수관은 원형이다.[24][23] 나후강은 크게 빗입목(Ctenostomata)과 입술입목(Cheilostomata)으로 나뉜다.[24]- '''빗입목''' (Ctenostomata)
- 군체는 직립형 또는 피복형이다.[62]
- 외골격은 키틴, 젤라틴질 또는 막질로 이루어져 있으며, 무기질화되지 않는다.[24][23]
- 대부분의 종에서 입을 덮는 뚜껑(Operculum)이 없다.[24][23]
- 촉수관을 내밀 때는 몸 전체의 벽을 압축한다.[24][23]
- 개충(zooid)은 기본적인 자가충 외에 줄기나 가시 형태를 가지기도 한다.[64]
- 예시: 갯솜벌레류, 끈벌레류, 미역벌레류, 차미드로벌레류.
- '''입술입목''' (Cheilostomata)
- 군체는 직립형, 피복형 또는 드물게 자유생활형이다.[24][23]
- 외골격은 탄산 칼슘 등으로 단단하게 무기질화되어 있다.[24][23]
- ''Bugula'' 속 등 일부를 제외하고는 입을 덮는 뚜껑(Operculum)이 있다.[24][23]
- 촉수관을 내밀 때는 몸 벽의 유연한 부분을 안쪽으로 당기거나 내부 주머니를 확장한다.[24][23]
- 자가충 외에도 방어, 생식 등 특수화된 다양한 이형 개충(heterozooids)을 가진다.[64] 이러한 다양한 개충 덕분에 태형동물 중 가장 많은 종을 포함하는 것으로 여겨진다.[23]
- 예시: 잔벌레류, 그물벌레류, 이타벌레류, 깃털벌레류, 작은벌레류, 모래알벌레류, 그물벌레류.
4. 2. 협후강 (Stenolaemata)
협후강(Stenolaematala)은 태형동물 문의 한 강이다.[122] 이 강에 속하는 대부분의 목은 중생대 말에 멸종했으며, 현재는 원구목만이 남아있다.[122]| 목(目) | 라틴어명 | 상태 | 비고 |
|---|---|---|---|
| †은구목 | Cryptostomidala | 멸종 | 중생대 말 멸종[122] |
| †변구목 | Trepostomatidala | 멸종 | 중생대 말 멸종[122] |
| †포공목 | Cystoporidala | 멸종 | 중생대 말 멸종[122] |
| †창격목 | Fenestridala | 멸종 | 중생대 말 멸종[122] |
| 원구목 | Cyclostomatala | 현존 | 국화벌레류, 실벌레류, 얇은벌레류 포함[122] |
4. 3. 피후강 (Phylactolaemata)
피후강(Phylactolaemata)은 태형동물 문에 속하는 강 중 하나이다. 주로 담수 환경에서 서식하며, 다른 두 강인 나후강(Gymnolaemata)과 협후강(Stenolaemata)과는 여러 차이점이 있어 다른 계통에 속할 가능성도 제기된다.[122]피후강의 주요 특징은 다음과 같다.[24][23]
- 입 위쪽에 입술 모양의 돌기(상순, epistome)가 있다.
- 군체는 젤라틴질 덩어리나 관 모양의 가지 구조를 이룬다.[61]
- 외골격은 젤라틴이나 막질이며, 단단하게 생물광물화되지 않는다.
- 개충의 입구를 덮는 뚜껑(개구덮개, operculum)이 없다.
- 촉수관(lophophore)은 일반적으로 U자 모양이지만, ''Fredericella'' 속은 원형이다.
- 촉수관을 확장할 때는 몸 전체의 벽을 압축하는 방식을 사용한다.
- 군체를 구성하는 개충은 먹이를 섭취하는 자가충(autozooid)만으로 이루어져 있다.[64]
피후강은 하나의 목으로 분류된다.
5. 진화
약 15,000종의 태형동물 화석이 발견되었으며, 고생대 화석의 3대 주요 그룹 중 하나이다.[65] 방해석 골격을 가진 태형동물은 석회암을 구성하는 탄산염 광물의 주요 원천이었고, 그 화석은 오르도비스기 이후 전 세계 해양 퇴적물에서 매우 흔하게 발견된다. 그러나 산호 및 다른 군체 동물과 달리 태형동물 군체는 큰 크기에 도달하지 못했다.[66] 태형동물 군체 화석은 일반적으로 심하게 파편화되고 산발적으로 발견되며, 완전한 군체(zoaria)가 보존되는 경우는 드물다.[67] 가장 큰 것으로 알려진 화석 군체는 미국 오르도비스기 암석에서 발견된 가지가 있는 트레포스토메 태형동물로, 높이가 66cm에 달한다.[66]
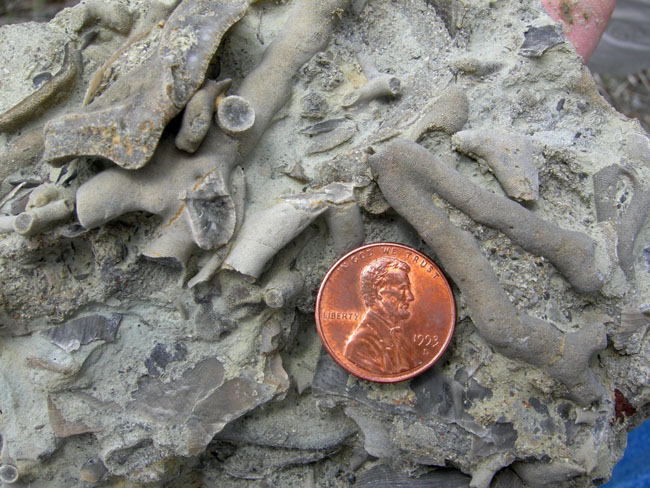






가장 오래된 광물화된 골격을 가진 종은 하부 오르도비스기(약 4억 8천만 년 전)에 나타난다.[13][34][68] 이는 오르도비스기 방산의 일부로 여겨진다. 최초의 태형동물은 훨씬 이전에 출현하여 완전히 연체였을 가능성이 높으며, 오르도비스기 화석은 이 문에서 광물화된 골격의 출현을 기록한다.[74] 오르도비스기 초기 무렵에는 모든 현생 목의 스테놀라에마타가 존재했고,[76] 크테노스토마티다 목의 짐놀라에마타는 중기 오르도비스기(약 4억 6천 5백만 년 전) 무렵에 출현했다. 초기 오르도비스기 화석은 이 문의 원래 구성원과 이미 상당히 달라진 형태를 나타낼 수도 있다.[76] 인산염으로 된 연조직을 가진 크테노스토마티다는 데본기에서 알려져 있다.[69] 다른 유형의 여과 섭식자가 거의 같은 시기에 출현했는데, 이는 어떤 변화가 이러한 생활 방식에 더 유리한 환경을 만들었음을 시사한다.[34] 키일로스토마타(광물화된 골격을 가진 짐놀라에마타 목)의 화석은 중기 쥐라기(약 1억 7천 2백만 년 전)에 처음 나타나며, 백악기부터 현재까지 가장 풍부하고 다양한 태형동물이었다.[34] 지난 1억 년 동안 수집된 증거에 따르면 키일로스토마티드는 영역 다툼에서 꾸준히 사이클로스토마티드보다 성장했는데, 이는 키일로스토마티드가 해양 태형동물의 우점종으로 사이클로스토마티드를 대체한 방법을 설명하는 데 도움이 될 수 있다.[70]
고생대(약 2억 5천 1백만 년 전 종료)의 해양 화석은 주로 직립 형태이며, 중생대의 화석은 직립 형태와 피복 형태로 거의 동등하게 나뉘며, 최근의 화석은 대부분 피복 형태이다.[71] 부드러운 담수 필락톨라에마타의 화석은 매우 드물며,[34] 후기 페름기(약 2억 6천만 년 전 시작) 이후에 나타나며, 내구성이 있는 휴면아(statoblast)로만 구성된다.[61] 다른 강의 담수 구성원의 화석은 알려진 것이 없다.[61]
태형동물 진화 동안 가장 중요한 사건 중 하나는 석회질 골격의 획득과 촉수 돌출 메커니즘의 관련 변화였다. 외벽의 강성은 더 큰 정도의 동물실 인접성과 거대하고 다열의 군체 형태의 진화를 허용했다.
과학자들은 태형동물(Ectoprocta)이 단계통군(단일 조상 종과 그 모든 자손을 포함하는 그룹)인지, 동물 계통수에서 가장 가까운 친척이 무엇인지, 심지어 원구동물(protostome)인지 후구동물(deuterostome)인지(모든 중간 정도로 복잡한 동물을 설명하는 두 가지 주요 그룹)에 대해 의견이 분분하다.
분자 계통 발생학은 잘 알려진 무척추동물 문들 간의 관계를 명확히 하는 데 많은 기여를 했지만,[43] 태형동물과 내형동물(Entoprocta)과 같은 "소규모 문"에 대한 유전자 데이터 부족으로 인해 다른 그룹과의 관계가 불분명하다.[72]
전통적인 견해는 태형동물이 단계통군이며, 그중 키일로스토마타 강이 화석 기록에서 가장 먼저 나타나는 스테놀라에마타와 크테노스토마티다와 가장 가까운 관계에 있다는 것이다.[100] 그러나 2005년, 키일로스토마타에 초점을 맞춘 분자계통발생 연구는 이들이 다른 태형동물 강보다 촉수동물(Phoronida), 특히 군체를 이루는 유일한 촉수동물 종과 더 가까운 관계에 있다는 결론을 내렸다. 이는 태형동물이 단계통군이 아니라는 것을 의미한다.[100]
2009년 또 다른 분자계통발생 연구는 미토콘드리아와 세포핵의 유전자 조합을 사용하여 태형동물이 단계통군이라는 결론을 내렸다. 이 분석은 또한 강(class)인 키일로스토마타, 스테놀라에마타, 그리고 짐놀라에마타도 단계통군이라는 결론을 내렸지만, 스테놀라에마타가 키일로스토마타와 짐놀라에마타 중 어느 쪽과 더 가까운 관계인지는 판단할 수 없었다. 짐놀라에마타는 전통적으로 연체성 크테노스토마티다와 광물화된 키일로스토마타로 나뉘지만, 2009년 분석에서는 이러한 목(order) 중 어느 것도 단계통군이 아니며 광물화된 골격이 초기 짐놀라에마타 내에서 두 번 이상 진화했을 가능성이 더 높다고 여겨졌다.[74]
태형동물과 다른 문과의 관계는 불확실하고 논란의 여지가 있다. 해부학과 배아 발달을 기반으로 하는 전통적인 계통 발생학은 태형동물의 위치에 대한 지속적인 합의를 이끌어내지 못했다.[78] 동물의 계통수를 재구성하려는 시도는 태형동물과 다른 "소규모 문"을 대부분 무시해 왔는데, 이는 이들이 일반적으로 크기가 작고, 비교적 단순한 신체 계획을 가지고 있으며, 인간 경제에 미치는 영향이 적기 때문이다.[75]
Ruth Dewel, Judith Winston, Frank McKinney의 견해에 따르면, 태형동물 형태학과 배아 발생에 대한 표준 해석은 100년 이상 모든 무척추동물에 대한 단일 프레임워크를 종합하려는 시도의 결과로 만들어진 구성물이며, 태형동물의 특이한 특징을 거의 고려하지 않는다.[76] 태형동물에서 유충의 모든 내부 기관은 성체로 변태하는 동안 파괴되고 성체의 기관은 유충의 표피와 중배엽으로부터 만들어지는 반면, 다른 좌우대칭동물에서는 창자를 포함한 일부 기관이 내배엽으로부터 만들어진다. 대부분의 좌우대칭동물 배아에서 원구(blastopore)는 유충의 창자로 깊어지지만, 태형동물에서는 원구가 사라지고 새로운 움푹 들어간 곳이 창자가 자라는 지점이 된다. 태형동물의 체강은 다른 좌우대칭동물이 사용하는 과정인 장체강 형성(enterocoely)이나 분할체강 형성(schizocoely)으로 형성되지 않는다.[76]
19세기에 내형동물(Entoprocta)이 발견되었을 당시, 내형동물과 외형동물(Ectoprocta, 즉 현재의 태형동물)은 모두 고착성 동물이며 섬모가 있는 촉수관을 이용하여 여과 섭식을 한다는 공통점 때문에 태형동물(Bryozoa) 문 내의 강으로 분류되었다. 그러나 1869년부터 내형동물의 항문이 섭식 구조물 안에 위치한다는 점, 초기 배의 세포 분열 패턴 차이 등 여러 차이점이 인식되면서 과학자들은 두 그룹을 별개의 문으로 간주하게 되었다.[72] "태형동물(Bryozoa)"이라는 이름은 항문이 섭식 기관 바깥에 있는 외형동물(Ectoprocta)의 동의어로 남게 되었다.[43] 1996년부터 2006년까지 일련의 분자 계통 발생 연구에서도 태형동물(외형동물)과 내형동물이 자매 분류군이 아니라는 결론을 내렸다.[72] 그러나 클라우스 니엘센(Claus Nielsen)과 토마스 카발리어-스미스(Thomas Cavalier-Smith)와 같은 일부 저명한 동물학자들은 해부학적 및 발생학적 근거를 들어 태형동물과 내형동물이 여전히 태형동물(Bryozoa) 문에 속하는 구성원이라고 주장하며, 2007년의 한 분자 계통 발생 연구 또한 이러한 견해를 뒷받침했다.[72]
태형동물(외형동물)은 전통적으로 촉수동물(Phoronida) 및 완족동물(Brachiopoda)과 함께 촉수관동물(Lophophorata)이라는 그룹으로 묶여왔다. 이들은 모두 여과 섭식을 위해 촉수관(lophophore)이라는 구조를 사용한다는 공통점이 있다.[43][77] 1970년대에는 촉수동물 유충과 일부 태형동물 유충(사이포나우테스 유충)의 비교 분석을 통해 태형동물이 촉수동물로부터 진화했다는 가설이 제기되기도 했다.[77] 그러나 클라우스 니엘센 등 일부 학자들은 이러한 유사성이 피상적이라고 주장한다. 예를 들어, 태형동물의 촉수에는 다수의 섬모(cilia)를 가진 세포가 있지만, 촉수동물, 완족동물, 그리고 유사한 구조를 가진 반삭동물(Hemichordata)의 익새류(Pterobranchia) 촉수관 세포는 세포당 하나의 섬모만을 가지며, 태형동물 촉수에는 다른 세 문의 촉수에 있는 혈관(hemal canal)이 없다.[78]
태형동물을 촉수관동물로 분류하는 것이 정확하다면, 다음 문제는 촉수관동물이 원구동물(protostome)인지 후구동물(deuterostome)인지이다. 전통적인 견해는 이들이 두 그룹의 특징을 혼합하여 가지고 있다는 것이었다. 1970년대 이후 연구는 후구동물의 특징으로 여겨지는 3부분 체강, 방사형 난할, 장체강 형성 등을 근거로 촉수관동물을 후구동물로 보았다.[43][78] 그러나 태형동물 유충의 체강은 3부분으로 나뉜 흔적이 없으며,[77] 성체 태형동물의 체강은 변태 과정에서 유충 체강이 파괴된 후 표피와 중배엽에서 새로 생성되는 등 다른 체강동물과 다른 방식으로 형성된다.[76]
1995년 이후의 분자 계통 발생 분석 결과는 태형동물이 후구동물보다는 환형동물 및 연체동물과 밀접하게 관련된 원구동물이라는 것을 강력하게 시사한다.[43][80] 이들은 나선동물(Lophotrochozoa)이라는 상위 분류군에 속하는 것으로 여겨진다. 형태적 특징과 유전자를 함께 사용한 분석은 다양한 결론을 내렸지만, 대부분 태형동물과 후구동물 간의 관계보다는 원구동물과의 관계를 지지한다.[80] 2008년의 한 연구는 더 많은 유전자를 사용하여 태형동물이 후구동물보다 원구동물에 더 가깝다는 결론을 내렸지만, 동시에 태형동물이 단계통군이 아니며 오히려 내형동물과 가장 가까운 관계라는, "태형동물"의 원래 정의를 뒷받침하는 결론을 내리기도 했다.[80]
현재 태형동물의 계통 발생적 위치는 여전히 논쟁의 여지가 있지만, 원구동물, 특히 나선동물에 속한다는 점은 비교적 확실해 보인다. 이는 태형동물 유생이 트로코포어 유생(trochophore)과 유사하며, 섬모관(corona)이 원섬모환(prototroch)의 상동기관일 수 있다는 점 등으로 뒷받침된다.[81] 미토콘드리아 DNA 염기서열 연구에 따르면, 태형동물은 모악동물(Chaetognatha)과 관련이 있을 수도 있다.[82]
6. 역사
태형동물은 과거 산호 등과 함께 '식충류'(Zoophyta)로 불리기도 했으나, 내부 구조 연구를 통해 독립된 동물 문으로 인식되었다. 1831년 에렌베르크(Ehrenberg)는 이들을 'Bryozoa'로 명명했다.[10][46][44] 처음에는 현재 내항동물(Entoprocta)로 분류되는 동물들도 포함했으나, 이후 두 그룹의 뚜렷한 차이가 밝혀지면서 분리되었다.
'Bryozoa'라는 이름은 원래 항문이 촉수관 바깥쪽에 열리는 동물들, 즉 현재 외항동물(Ectoprocta)로 불리는 그룹에만 적용되었다.[41] 항문이 촉수관 안쪽에 열리는 내항동물이 발견되자,[42] 'Bryozoa'는 이 두 강을 모두 포함하는 문 수준의 분류군으로 격상되기도 했다.[43] 그러나 1869년 힌리히 니체(Hinrich Nitsche)는 항문 위치, 촉수 형태(외항동물은 속이 빈 촉수, 내항동물은 속이 찬 촉수), 체강 유무(외항동물은 체강이 있고, 내항동물은 없음) 등 해부학적 차이를 근거로 외항동물에 'Ectoprocta'(외부 항문)라는 이름을 부여하며 두 그룹을 명확히 구분했다.[74][72]
이후 외항동물과 내항동물은 별개의 문으로 간주되는 것이 일반적이며, 'Bryozoa'는 외항동물(Ectoprocta)과 동의어로 사용되는 경우가 많아졌다.[43] 하지만 명칭 사용에 혼란이 지속되어 1960년대와 1970년대에는 모호하지 않은 'Ectoprocta'를 사용하자는 제안도 있었으나,[45] 기존 문헌과의 연속성 문제 등으로 인해 'Bryozoa'가 여전히 선호되는 경향이 있다.[46] 2000년대 이후 연구에서도 "Bryozoa",[24][34] "Ectoprocta",[78][23] "Bryozoa (Ectoprocta)",[27] "Ectoprocta (Bryozoa)"[48] 등 다양한 표기가 혼용되고 있다.[49] 일반적인 이름인 '이끼벌레'는 그리스어 βρυόν|브리온grc('이끼')과 ζῷα|조아grc('동물')에서 유래했으며, 일부 종의 이끼 같은 외형 때문에 붙여졌다.[50]
태형동물 화석은 약 15,000종이 발견되었으며, 고생대 화석의 주요 그룹 중 하나이다.[65] 생물광물화된 골격을 가진 최초의 태형동물 화석은 오르도비스기 전기(아레니그절, 약 4억 8천만 년 전) 지층에서 발견된다.[13][34][68] 이보다 앞선 캄브리아기에도 태형동물이 존재했을 수 있으나, 연체성이었거나 다른 이유로 화석으로 보존되지 않았을 가능성이 제기된다.[74] 오르도비스기에는 이미 현생 스테놀라에마타(Stenolaemata) 강의 모든 목이 출현했으며,[76] 짐놀라에마타(Gymnolaemata) 강의 크테노스토마티다(Ctenostomatida) 목은 중기 오르도비스기(약 4억 6천 5백만 년 전)에 나타났다.[76] 오르도비스기 해저 생태계에서 태형동물은 퇴적물 안정화와 다른 저서 생물의 먹이원으로서 중요한 역할을 했다.[65] 특히 미시시피기(약 3억 5400만 년 ~ 3억 2300만 년 전)에는 태형동물이 매우 번성하여 그 골격 파편이 두꺼운 석회암 층을 형성하기도 했다.
광물화된 골격을 가진 짐놀라에마타 목인 키일로스토마타(Cheilostomata)는 중기 쥐라기(약 1억 7천 2백만 년 전)에 처음 나타나 백악기 이후 가장 풍부하고 다양한 태형동물 그룹이 되었다.[34] 지난 1억 년 동안 키일로스토마타는 사이클로스토마타(Cyclostomata)와의 영역 경쟁에서 우위를 점하며 해양 태형동물의 우점종으로 자리 잡은 것으로 보인다.[70] 화석 기록을 보면 고생대에는 직립형 군체가 우세했으나, 중생대에는 직립형과 피복형이 비슷하게 나타나고, 이후로는 피복형이 주를 이루는 경향을 보인다.[71] 담수에 서식하는 필락톨라에마타(Phylactolaemata) 강의 화석은 매우 드물며, 후기 페름기(약 2억 6천만 년 전 시작) 이후의 지층에서 내구성이 있는 휴면아(statoblast) 형태로만 발견된다.[34][61] 태형동물 진화에서 가장 중요한 사건 중 하나는 석회질 골격의 획득과 그에 따른 촉수 돌출 메커니즘의 변화로, 이는 군체 형태의 다양화와 대형화를 가능하게 했다.[65]
태형동물의 계통 발생학적 위치에 대해서는 오랫동안 논쟁이 있었다. 과거에는 발생 과정의 유사성 등을 근거로 후구동물로 분류되기도 했으나, 1995년 이후 분자 계통 발생 분석 결과는 이들이 연체동물, 환형동물과 함께 원구동물에 속한다는 것을 강력히 시사한다.[43][80] 하지만 형태학적 특징과 유전자 정보를 함께 분석한 일부 연구에서는 여전히 후구동물과의 관계를 지지하는 등 다양한 결과가 제시되기도 했다.[80] 2008년의 한 연구는 더 많은 유전자를 분석하여 태형동물(외항동물)이 원구동물에 더 가깝다는 결론을 내렸으나, 동시에 외항동물이 단계통군이 아니며 오히려 내항동물과 가장 가까운 관계일 수 있다는 결과를 제시하기도 했다. 이 연구는 완족류와 포로니다가 자매군을 형성한다고 보았다.[80]
최근에는 저진공 주사 전자 현미경과 같은 현대 기술을 이용하여 기존에 분류가 불분명했던 Oncousoeciidae 과의 모식종들을 재검토하고 분류 체계를 수정하는 연구가 이루어졌다.[56] 또한 Stomachetosellidae 과에서 새로운 속인 ''Junerossia''와 함께 10종의 새로운 태형동물 종이 발견되는 등[57] 활발한 연구가 계속되고 있다.
참조
[1]
논문
The oldest known bryozoan: ''Prophyllodictya'' (Cryptostomata) from the lower Tremadocian (Lower Ordovician) of Liujiachang, south-western Hubei, central China
2015-09-01
[2]
논문
A cystoporate bryozoan species from the Zechstein (Late Permian)
2007-01-01
[3]
서적
Common intertidal invertebrates of the Gulf of California
University of Arizona Press
1980-01-01
[4]
논문
Phylum Bryozoa {{small|Ehrenberg, 1831}}
https://www.research[...]
2013-08-01
[5]
논문
Key novelties in the evolution of the aquatic colonial phylum Bryozoa: evidence from soft body morphology
2020-06-01
[6]
논문
Aethozooides uraniae, a new deep-sea genus and species of solitary bryozoan from the Mediterranean Sea, with a revision of the Aethozoidae
https://doi.org/10.1[...]
2019-08-01
[7]
서적
Zoological researches and illustrations; or Natural history of nondescript or imperfectly known animals
https://archive.org/[...]
King and Ridings
1830-01-01
[8]
서적
Symbolae physicae, seu lcones et descriptiones corporum naturalium novorum aut minus cognitorum.
Berolini Ex officina Academica
1831-01-01
[9]
논문
The terms Polyzoa and Bryozoa
https://archive.org/[...]
1911-01-01
[10]
서적
A history of the classification of the phylum Brachiopoda
https://archive.org/[...]
British Museum (Natural History)
1955-01-01
[11]
서적
The Mechanistic Benefits of Microbial Symbionts
Springer International Publishing
2016-01-01
[12]
웹사이트
Introduction to the Bryozoa
https://ucmp.berkele[...]
University of California Museum of Paleontology
2019-12-08
[13]
논문
Reinterpretation of the Cambrian 'bryozoan' ''Pywackia'' as an octocoral
https://zenodo.org/r[...]
2013-11-01
[14]
논문
Fossil evidence unveils an early Cambrian origin for Bryozoa
2021-01-01
[15]
논문
Protomelission is an early dasyclad alga and not a Cambrian bryozoan
https://durham-repos[...]
2023-03-08
[16]
서적
Invertebrate Zoology
Brooks / Cole
[17]
서적
Invertebrate Zoology
Brooks / Cole
[18]
서적
Invertebrate Zoology
Brooks / Cole
[19]
서적
Invertebrate Zoology
Brooks / Cole
[20]
서적
Invertebrate Zoology
Brooks / Cole
[21]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[22]
서적
Meiobenthology
Springer Verlag
2009-07-07
[23]
서적
Invertebrate Zoology
Oxford University Press
2001-01-01
[24]
서적
Invertebrate Zoology
Brooks / Cole
[25]
서적
Shorter Oxford English Dictionary
Oxford University Press
[26]
간행물
Bryozoans and Palaeoenvironmental Interpretation
http://palaeontologi[...]
[27]
서적
Encyclopedia of Life Sciences
John Wiley & Sons, Ltd.
2001-01-01
[28]
서적
Bryozoan evolution
University of Chicago Press
2009-07-29
[29]
논문
Downstream collecting in ciliary suspension feeders: the catch-up principle
https://www.int-res.[...]
2009-09-12
[30]
서적
Random House Dictionary
Random House
2009-08-02
[31]
서적
Ctenostome Bryozoans
Linnean Society of London
2009-08-02
[32]
서적
Meiobenthology
Springer-Verlag
2009-08-02
[33]
서적
A Greek-English Lexicon
Clarendon Press
2009-08-01
[34]
서적
The fossil book
https://archive.org/[...]
Dover Publications
2009-08-07
[35]
서적
Two Oceans – A Guide to the Marine Life of Southern Africa
Struik
2009-08-02
[36]
서적
Shorter Oxford English Dictionary
Oxford University
[37]
학술지
A Model of Particle Capture by Bryozoans in Turbulent Flow: Significance of Colony Form
1998-12-01
[38]
서적
Life in moving fluids
Princeton University Press
2009-08-05
[39]
학술지
Function-Dependent Development in a Colonial Animal
http://www.biolbull.[...]
2006-08-01
[40]
학술지
Secular changes in colony-forms and bryozoan carbonate sediments through geological history
2013-08-01
[41]
서적
Shorter Oxford English Dictionary
Oxford University Press
[42]
서적
Shorter Oxford English Dictionary
Oxford University Press
[43]
학술지
The new view of animal phylogeny
http://www-fourier.u[...]
2016-08-26
[44]
학술지
Bryozoa versus Ectoprocta
1968-06-01
[45]
학술지
Bryozoa versus Ectoprocta – The Necessity for Precision
[46]
학술지
Names of Invertebrate Phyla
[47]
학술지
On Changing the Names of Higher Taxa
[48]
학술지
Complete nucleotide sequences of mitochondrial genomes of two solitary entoprocts, ''Loxocorone allax'' and ''Loxosomella aloxiata'': Implications for lophotrochozoan phylogeny
2008-05-01
[49]
학술지
Taxonomically Important Features on the Surface of Floatoblasts in ''Plumatella'' (Bryozoa)
[50]
서적
Shorter Oxford English Dictionary
Oxford University Press
[51]
웹사이트
ITIS Standard Report Page: Ectoprocta
https://www.itis.gov[...]
Integrated Taxonomic Information System
2009-08-23
[52]
서적
Shorter Oxford English Dictionary
Oxford University Press
1964
[53]
서적
Shorter Oxford English Dictionary
Oxford University Press
1964
[54]
학술지
Bryozoa versus Ectoprocta – The Necessity for Precision
1969-01-01
[55]
웹사이트
ITIS Standard Report Page: Bryozoa
https://www.itis.gov[...]
Integrated Taxonomic Information System
2009-08-23
[56]
문서
Taylor, Zaton 2008
[57]
학술지
Taxonomy of the bryozoan genera Oncousoecia, Microeciella and Eurystrotos
2008-10-01
[58]
서적
Numbers of Living Species in Australia and the World
http://www.environme[...]
Department of the Environment and Heritage, Australian Government
2009-08-07
[59]
서적
Invertebrate Zoology
Brooks / Cole
[60]
웹사이트
ITIS Standard Report Page: Phylactolaemata
https://www.itis.gov[...]
Integrated Taxonomic Information System
2009-08-12
[61]
학술지
Global diversity of bryozoans (Bryozoa or Ectoprocta) in freshwater
[62]
서적
A student's guide to the seashore
Cambridge University Press
[63]
학술지
Comparative Ecology of Bryozoan Radiations: Origin of novelties in cyclostomes and Cheilostomes
[64]
서적
Cyclostome bryozoans
Linnean Society of London
2009-08-09
[65]
뉴스
Bryozoan Evolution
Boston: Unwin & Hyman, 1989
[66]
서적
Phylum Bryozoa
2020-01-01
[67]
학술지
Epizoan and endoskeletozoan distribution across reassembled ramose stenolaemate bryozoan zoaria from the Upper Ordovician (Katian) of the Cincinnati Arch region, USA
2019-01-01
[68]
학술지
Cambrian-Ordovician paleogeography of Baltica
1991-01-01
[69]
학술지
Exceptional soft-tissue preservation in boring ctenostome bryozoans and associated "fungal" borings from the Early Devonian of Podolia, Ukraine
[70]
학술지
One hundred million years of competitive interactions between bryozoan clades: asymmetrical but not escalating
[71]
서적
Reef evolution
https://books.google[...]
Oxford University Press
2009-08-11
[72]
논문
Spiralian Phylogenomics Supports the Resurrection of Bryozoa Comprising Ectoprocta and Entoprocta
[73]
논문
The boring ctenostomate bryozoa: taxonomy and paleobiology based on cavities in calcareous substrata
[74]
논문
The first comprehensive molecular phylogeny of Bryozoa (Ectoprocta) based on combined analyses of nuclear and mitochondrial genes
2009-07-00
[75]
논문
The Essential Role of "Minor" Phyla in Molecular Studies of Animal Evolution
[76]
서적
Bryozoan studies 2001: proceedings of the Twelfth International Bryozoology Conference
Swets and Zeitlinger
2009-08-13
[77]
서적
Animal evolution: interrelationships of the living phyla
Oxford University Press
2009-08-14
[78]
논문
The Phylogenetic Position of Entoprocta, Ectoprocta, Phoronida, and Brachiopoda
2002-07-00
[79]
웹사이트
Introduction to the Hemichordata
http://www.ucmp.berk[...]
University of California Museum of Paleontology
2008-09-22
[80]
논문
Phylogenomic analyses of lophophorates (brachiopods, phoronids and bryozoans) confirm the Lophotrochozoa concept
[81]
논문
Structure and occurrence of cyphonautes larvae (Bryozoa, Ectoprocta)
2010-09-00
[82]
논문
Complete mitochondrial genome of ''Membranipora grandicella'' (Bryozoa: Cheilostomatida) determined with next-generation sequencing: The first representative of the suborder Malacostegina
2012-09-00
[83]
논문
Living where the flow is right: How flow affects feeding in bryozoans
https://scholarworks[...]
[84]
논문
Respiration in polyzoa (ectoprocta)
[85]
논문
Three in one: Evolution of viviparity, coenocytic placenta and polyembryony in cyclostome bryozoans
2021-00-00
[86]
논문
The placental analogue and the pattern of sexual reproduction in the cheilostome bryozoan Bicellariella ciliata (Gymnolaemata)
2012-00-00
[87]
논문
Unravelling the Evolution of Bryozoan Larvae
https://ui.adsabs.ha[...]
2024-00-00
[88]
논문
Morphology and life cycle of an epiphytic pherusellid ctenostome bryozoan from the Mediterranean Sea
2020-00-00
[89]
서적
Phylum Bryozoa
https://books.google[...]
Walter de Gruyter GmbH & Co KG
2020-11-23
[90]
논문
The first deep-sea ctenostome bryozoan from the Indian Ocean: ''Aethozoon flavum'' sp. nov.
2024-00-00
[91]
논문
Versatile ciliary behaviour in capture of particles by the bryozoan cyphonautes larva
2006-03-00
[92]
논문
''Asajirella gelatinosa'' in Panama: a bryozoan range extension in the Western Hemisphere
1998-12-00
[93]
논문
Modes of reproduction in recent and fossil cupuladriid bryozoans
[94]
서적
Planet Earth: Cosmology, Geology, & the Evolution of Life & the Environment
https://archive.org/[...]
Cambridge University Press
2009-08-11
[95]
서적
Applied palaeontology
Cambridge University Press
2009-08-11
[96]
논문
Comparison of bryozoan assemblages from two contrasting Arctic shelf regions
[97]
서적
Invertebrates (2nd Edition)
Sunderland, MA: Sinauer Associates
[98]
논문
A pelagic bryozoan from Antarctica
https://link.springe[...]
2017-08-28
[99]
뉴스
'Weedy thing' thrives as Antarctic shores warm
https://www.bbc.co.u[...]
2014-06-16
[100]
서적
Bryozoan Studies 2004: Proceedings of the 13th International Bryozoology Association
Taylor & Francis Group
2009-08-24
[101]
웹사이트
Bryozoans: The Fascinating Colonies Of Phylum Ectoprocta
https://www.earthlif[...]
2020-03-05
[102]
논문
Specificity of cues inducing defensive spines in the bryozoan Membranipora membranacea
http://cat.inist.fr/[...]
2009-08-18
[103]
서적
Cyclostome bryozoans: keys and notes for the identification of the species
Brill Archive
2009-08-18
[104]
논문
Predation by ''Patiria miniata'' (Asteroidea) on bryozoans
1981-01-00
[105]
서적
Predator-prey interactions in the fossil record
Springer
2009-08-18
[106]
논문
Freshwater Bryozoans of Thailand (Ectoprocta and Entoprocta)
http://www.wright.ed[...]
2009-08-24
[107]
논문
Summer dormancy as a refuge from mortality in the freshwater bryozoan ''Plumatella emarginata''
2002-06
[108]
논문
Heavy Predation on Freshwater Bryozoans by the Golden Apple Snail, ''Pomacea canaliculata''
https://web.archive.[...]
2006-05
[109]
논문
Symbiotic relationships between hydroids and bryozoans
http://cat.inist.fr/[...]
[110]
논문
A symbiotic shell-encrusting bryozoan provides subtidal whelks with chemical defence against rock lobsters
2005-12
[111]
논문
Bryoliths constructed by bryozoans in symbiotic associations with hermit crabs in a tropical heterozoan carbonate system, Golfe d'Arguin, Mauritania
https://doi.pangaea.[...]
2013-07-30
[112]
논문
Molecular data implicate bryozoans as hosts for PKX (Phylum Myxozoa) and identify a clade of bryozoan parasites within the Myxozoa
http://cat.inist.fr/[...]
[113]
논문
Professional photosensitive eczema of fishermen by contact with bryozoans: disabling occupational dermatosis
http://www.imh.mug.e[...]
[114]
서적
Five kingdoms: an illustrated guide to the phyla of life on earth
https://archive.org/[...]
Elsevier
[115]
논문
Biofouling of wastewater treatment plants by the freshwater bryozoan, ''Plumatella vaihiriae''
1999-02
[116]
웹사이트
Bryostatin 1
https://web.archive.[...]
2006-06-19
[117]
웹사이트
Safety, Efficacy, Pharmacokinetics, and Pharmacodynamics Study of bryostatin 1 in Patients With Alzheimer's Disease
http://clinicaltrial[...]
National Institutes of Health
2009-08-19
[118]
논문
Bryostatin Effects on Cognitive Function and PKCɛ in Alzheimer's Disease Phase IIA and Expanded Access Trials
[119]
논문
The Practical Synthesis of a Novel and Highly Potent Analogue of Bryostatin
2002-11-20
[120]
서적
On the origins of phyla
University of Chicago Press
[121]
서적
無脊椎動物の多様性と系統(節足動物を除く)
裳華房
[122]
서적
On the origins of phyla
University of Chicago Press
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com