색소체
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
색소체는 식물 세포 내에서 발견되는 세포 소기관으로, 다양한 기능을 수행하며, 내부 공생을 통해 시아노박테리아에서 유래되었다. 주요 기능으로는 광합성, 색소 합성 및 저장, 녹말, 지방, 단백질 저장 등이 있다. 색소체는 엽록체, 유색체, 백색체 등으로 분화하며, 식물 종류와 환경에 따라 다양한 형태로 존재한다. 색소체는 자체 DNA를 가지고 있으며, 핵 유전자와 상호 작용하여 발달과 기능을 조절한다.
더 읽어볼만한 페이지
- 색소 - 색소포
색소포는 냉혈 척추동물과 두족류에서 발견되는 색소 함유 세포로, 흑색소포, 적색소포, 황색소포, 백색소포, 무지개 색소포 등으로 나뉘며 생체의 색을 나타내고 체색 변화를 통해 환경에 적응하거나 사회적 신호를 전달한다. - 세포소기관 - 소포 (세포)
소포는 막으로 둘러싸인 작은 주머니 모양의 구조물로, 세포 내 물질 수송, 소화, 분비, 세포간 신호 전달, 세포 외부 물질 소화 및 배출 등 다양한 기능을 수행하며, 특히 세포외소포체는 질병과의 연관성으로 주목받고 있고, 클라트린, COPI, COPII, SNARE 단백질 등이 소포 형성 및 수송에 관여한다. - 세포소기관 - 세포막
세포막은 세포질을 둘러싸고 세포 내외부 환경을 구분하는 선택적 투과성 막으로, 인지질 이중층과 단백질, 탄수화물로 구성되어 물질 수송, 세포 형태 유지, 세포 간 신호 전달, 세포 접합 등의 기능을 수행하며, 유동 모자이크 모델로 설명된다. - 식물생리학 - 광합성
광합성은 생물이 빛에너지를 이용하여 이산화탄소와 물로부터 유기물을 합성하고 산소를 방출하는 과정으로, 엽록체 내 틸라코이드 막에서 일어나는 명반응과 스트로마에서 일어나는 암반응으로 구성되며, 환경에 따라 탄소 농축 메커니즘을 통해 효율을 높이기도 하고, 지구 대기의 산소를 생성하는 주요 원천이다. - 식물생리학 - 저분자
저분자는 분자량이 900달톤 이하인 작은 분자로, 세포막 투과가 용이하여 의약품으로 많이 사용되지만, 표적 적용에 제한이 있고 내성 문제가 발생할 수 있으며, 의약품 외에도 이차 대사산물, 연구 도구, 항게놈 치료제 등으로 활용된다.
| 색소체 | |
|---|---|
| 개요 | |
 | |
| 종류 | 세포 소기관 |
| 기능 | 광합성, 녹말 저장 |
| 상세 정보 | |
| 정의 | 광합성을 수행하고 녹말을 저장하는 식물 세포 소기관 |
| 어원 | 그리스어 πλαστός (plastós, "형성된", "주조된") |
| 역사 | 1880년대에 에른스트 헤켈(으)로부터 유래 |
| 기원과 진화 | 시아노박테리아의 내부 공생에서 기원 |
| 유전체 | 독자적인 유전체를 가짐 시아노박테리아와 유사한 리보솜을 가짐 |
| 특징 | 세포당 하나의 색소체를 갖는 경우가 많음 막으로 둘러싸여 있음 |
| 종류 | |
| 주요 종류 | 엽록체 색소체 백색체 단백질체 |
| 기타 종류 | 아밀로플라스트 엘라이오플라스트 크로모플라스트 로도플라스트 탄노솜 |
| 관련 용어 | |
| 관련 용어 | 세포 소기관 내부 공생 식물 세포 광합성 녹말 |
| 학술적 정보 | |
| 학명 | Plastids |
2. 역사

엽록체에 관한 최초의 기록은 1837년 폰 몰(von Mohl)에 의한 것이다. 그러나 1800년대 후반까지 광합성의 기본적인 사실이 거의 발견되지 않았기 때문에 엽록체는 이보다 훨씬 앞서 관찰되었음이 틀림없다. 1818년 페르티에(Pelletier)는 녹색 색소에 '클로로필(엽록소)'이라는 이름을 붙였다. 윌스테터(Willstätter) 등은 클로로필의 화학적 성질을 규명하였고, 우드워드(Woodward) 등의 연구를 통해 클로로필의 인공 합성이 가능해졌다.
1950년대 후반부터 전자 현미경이 엽록체 연구에 도입되면서 엽록체의 내부 구조가 자세히 밝혀졌다. 성능이 좋은 광학 현미경으로 엽록체를 관찰하면 암록색을 띤 지름 0.2 ~ 0.3μm의 입자인 그라나를 많이 볼 수 있다.
엽록체는 핵분열과는 무관하게 분열하여 증식한다. 이는 핵 DNA와는 다른 성질의 DNA를 함유하며, 매우 높은 자율성을 가진 세포 기관으로 생각되었기 때문이다. 또한 세포질 내의 단백 합성계와는 다른 특유한 합성계를 갖고 있다는 것도 밝혀졌다. 그러나 연구가 진보됨에 따라 엽록체의 자율성은 완전한 것이 아니라 핵 유전자의 지배를 받는 부분이 많다는 것도 밝혀졌다.
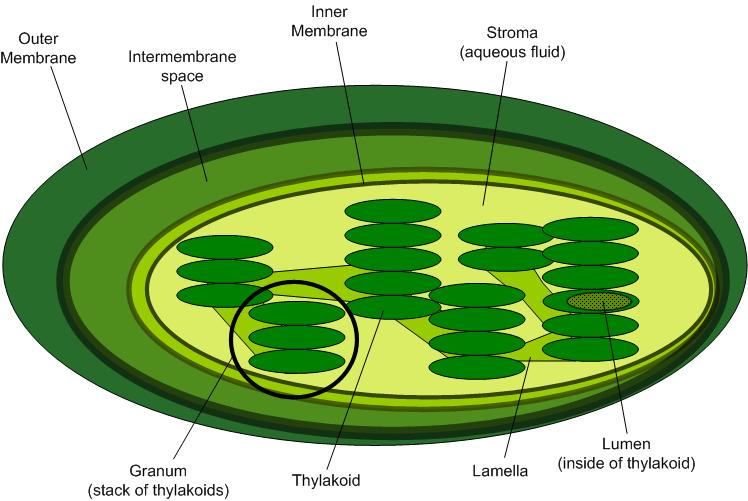
3. 구조
엽록체는 외막, 내막, 막 사이 공간으로 둘러싸인 이중막 구조를 가진다. 엽록체 내부에는 스트로마(stroma)라고 불리는 기질과 틸라코이드(thylakoid)라고 불리는 납작한 주머니 모양의 막 구조가 존재한다. 틸라코이드는 여러 개가 쌓여 그라나(grana)를 형성하며, 그라나 사이는 스트로마 틸라코이드(stroma thylakoid) 또는 라멜라(lamella)로 연결된다. 틸라코이드 막에는 엽록소, 카로티노이드 등 광합성 색소와 전자 전달계 단백질이 존재하여 광합성의 명반응이 일어난다. 스트로마에는 캘빈 회로(탄소 고정 반응)에 관여하는 효소들이 존재하여 광합성의 암반응이 일어난다.
엽록체는 세포핵과는 별도로 분열하여 증식하며, 자체 DNA와 리보솜을 가지고 있어 일부 단백질을 자체적으로 합성한다. 그러나 엽록체의 자율성은 완전하지 않으며, 엽록소 합성에 관여하는 효소나 탄산 고정 회로에 관여하는 효소 등은 세포핵 유전자에 의해 조절된다.
식물 세포에서 스트로뮬이라고 하는 가늘고 긴 융기가 형성되어 색소체에서 세포질 기질로 뻗어 나가거나, 색소체 사이를 연결하기도 한다. 단백질과 더 작은 분자도 스트로뮬 안을 이동할 수 있다.
3. 1. 엽록체 DNA와 핵양체
각 색소체는 고유한 게놈인 플라스타 게놈('색소체 게놈'에서 유래)의 여러 복사본을 생성하며, 엽록소 색소체(또는 엽록체)의 경우 '엽록체 게놈' 또는 '엽록체 DNA'와 동일하다.[11][12] 색소체당 생성되는 게놈 복사본 수는 다양하며, 몇 개의 색소체만 포함하는 빠르게 분열하는 새로운 세포에서는 1000개 이상, 수많은 색소체를 포함하는 성숙한 세포에서는 100개 이하로 다양하다.
플라스타 게놈은 일반적으로 ''전달'' 리보핵산 (tRNA)과 ''리보솜'' 리보핵산 (rRNA)을 암호화하는 게놈을 포함한다. 또한 광합성과 색소체 유전자 전사 및 번역에 관여하는 단백질도 포함한다. 그러나 이러한 단백질은 특정 유형의 색소체를 만들고 유지하는 데 필요한 전체 단백질 세트의 작은 부분만을 나타낸다. 핵 유전자(식물의 세포 핵 내)는 대부분의 색소체 단백질을 암호화하며, 핵 및 색소체 유전자의 발현은 색소체의 발달 및 분화를 조정하기 위해 공동으로 조절된다.
많은 색소체, 특히 광합성을 담당하는 색소체는 수많은 내부 막 층을 가지고 있다. 색소체 DNA는 색소체의 내부 외피 막 내의 국소적인 ''영역''으로 관련된 단백질-DNA 복합체로 존재하며, 이러한 복합체를 '색소체 뉴클레오이드'라고 한다. 진핵 세포의 핵과는 달리, 색소체 뉴클레오이드는 핵막으로 ''둘러싸여 있지 않다''. 각 뉴클레오이드의 영역은 10개 이상의 색소체 DNA 사본을 포함할 수 있다.
원색소체(''분화되지 않은 색소체'')는 원색소체의 중앙 근처에 위치한 단일 뉴클레오이드 영역을 포함하는 반면, ''발달하는(또는 분화하는) 색소체''는 색소체의 주변부에 위치하고 내부 외피 막에 결합된 많은 뉴클레오이드를 가지고 있다. 원색소체가 엽록체로 발달/분화하는 동안, 그리고 색소체가 한 유형에서 다른 유형으로 분화될 때 뉴클레오이드는 형태, 크기 및 세포 소기관 내 위치에서 변화한다. 색소체 뉴클레오이드의 재구성은 뉴클레오이드 단백질의 풍부함과 조성의 수정에 의해 일어나는 것으로 여겨진다.
4. 기능
엽록체의 가장 중요한 기능은 광합성이다. 광합성은 빛 에너지를 이용하여 이산화탄소와 물로부터 포도당과 같은 유기물을 합성하고 산소를 방출하는 과정이다. 광합성은 명반응과 암반응(캘빈 회로)으로 구분된다. 명반응은 틸라코이드 막에서 일어나며, 빛 에너지를 흡수하여 ATP와 NADPH를 생성하고 물을 분해하여 산소를 방출한다. 암반응은 스트로마에서 일어나며, ATP와 NADPH를 이용하여 이산화탄소를 고정하여 포도당을 합성한다. 엽록체는 광합성 외에도 지방산, 테르펜 등 다양한 화합물의 합성과 저장에도 관여한다.
엽록체에는 여러 가지 색소가 들어 있는데, 가장 중요한 것은 클로로필이다. 클로로필 외에도 카로티노이드 색소와 피코시아닌(남조소)·피코에리트린(홍조소)·푸코키산틴(녹조소) 등이 있다. 보조 색소가 받아들인 빛 에너지는 최종적으로는 클로로필 a에 전달된다. 홍조나 녹조에는 다량의 피코에리트린이나 푸코키산틴이 함유되어 있기 때문에 붉은색이나 녹색으로 보이는데, 반드시 클로로필 a를 함유하며, 광합성을 한다.[1]
성능이 좋은 광학 현미경으로 엽록체를 관찰하면 암록색을 띤 지름 0.2 ~ 0.3μ의 입자를 많이 볼 수 있는데, 이 입자를 그라나라고 부른다. 그라나 이외의 기질 부분을 스트로마라고 한다. 엽록체를 얇게 절단하여 전자 현미경으로 보면 그라나 부분은 동전이 여러 개 쌓여 있는 막 구조로 되어 있다. 이 막 구조는 매우 평평한 자루로 되어 있기 때문에 틸라코이드 또는 그냥 라메라라고 한다.[1]
5. 엽록체의 분화
식물 세포 내에서 엽록체는 전구체인 전색소체(proplastid)로부터 분화한다. 전색소체는 빛의 유무, 조직의 종류, 세포의 기능 등에 따라 다양한 형태의 색소체로 분화할 수 있다.
모든 색소체는 식물의 분열조직 부위에 존재하는 전색소체에서 유래한다. 전색소체와 어린 엽록체는 일반적으로 이분법으로 분열하지만, 더 성숙한 엽록체도 이 능력을 가지고 있다.
식물 ''전색소체''(미분화 색소체)는 세포 내에서 수행하는 기능에 따라 여러 형태로 세포 분화될 수 있다.[10]
- 엽록체: 광합성을 수행하는 녹색 색소체.
- * 에티오플라스트: 엽록체의 전구체.
- 유색체: 색소를 합성하고 저장하는 유색 색소체.
- 노화엽록체: 식물 노화 동안 광합성 장치의 해체를 제어하는 색소체.
- 백색체: 테르펜의 일종인 모노테르펜을 합성하는 무색 색소체.
- 알레우로플라스트;
- * 전분체: 녹말을 저장하고 중력을 감지하여 지구굴성을 유지한다.
- * 유지체: 지방을 저장한다.
- * 단백질체: 단백질을 저장하고 수정한다.
- 탄닌체: 탄닌과 폴리페놀을 합성하고 생성한다.
색소체는 형태와 목표 기능에 따라 이러한 형태와 다른 형태 사이에서 분화하거나 재분화할 수 있다.
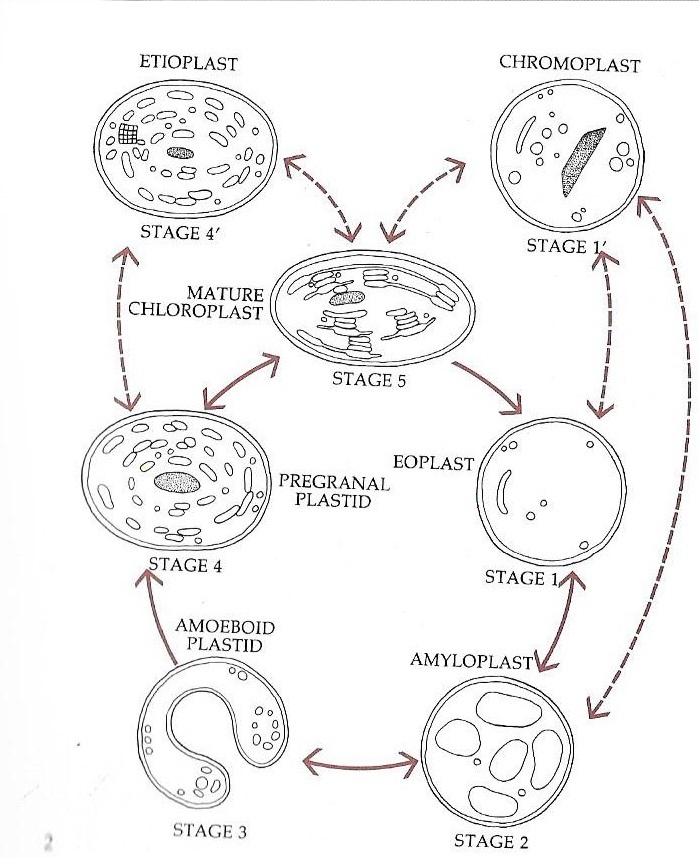
1977년 J.M. 왓리(J.M. Whatley)는 색소체 발달 주기를 제안했는데, 이는 색소체의 발달이 항상 단방향이 아니라 복잡한 순환 과정임을 나타낸다.[28]
색이나 기능에 따라 엽록체, 백색체, 유색체 등으로 나뉘지만, 이것들은 고정된 것이 아니라 서로 분화하거나 재분화할 수 있다.
식물에서는 색소체는 각 세포에 필요한 기능에 따라 여러 종류의 형태로 변화한다. 식물체의 모든 색소체는 분열 조직에 있는 미분화된 색소체인 '''원색소체'''(proplastid, 이전에는 '''에오플라스트'''(eoplast)라고 불렀다. eo-는 새벽, 빠름의 의미)에서 유래하며, 다음의 어떤 색소체로도 발달할 수 있다.
- 아밀로플라스트(amyloplast): 녹말 저장
- 에티오플라스트(전엽록체, etioplast): 아직 빛을 받지 않은 엽록체
- 엘라이오플라스트(elaioplast): 지질 저장
- 유색체(chromoplast): 색소의 합성 및 저장
원색소체와 어린 엽록체는 잘 분열하지만, 보다 성숙한 엽록체도 분열 능력이 있다.
한편 조류에서는 엽록체의 구조와 색소 조성이 계통에 따라 다르며, 각각 다른 이름으로 불리는 경우가 있다. 예를 들어 홍조류에서는 '''홍색체'''(rhodoplast), 회색조류나 ''Paulinella chromatophora''(케르코조아의 유각 아메바)에서는 '''시아넬'''(cyanelle)이라고 불린다. 또한 백색체라는 용어는 색소가 없는 모든 색소체에 사용되며, 이미 퇴화하여 엽록체로의 분화 능력을 잃은 것도 포함된다. 에티오플라스트, 아밀로플라스트, 유색체는 식물 특이적이며 조류에는 없다. 조류의 색소체는 피레노이드를 포함한다는 점에서도 식물의 색소체와 다르다.
5. 1. 엽록체 (Chloroplast)
광합성을 수행하는 녹색 색소체이다. 1837년 폰 몰이 처음으로 엽록체에 대한 기록을 남겼다. 고등 식물의 엽록체는 지름 5μm 정도의 볼록 렌즈 모양이며, 보통 하나의 세포 내에 수십 개가 존재한다. 엽록체에는 엽록소(클로로필)를 비롯하여 카로티노이드 색소, 피코시아닌, 피코에리트린, 푸코키산틴 등 다양한 색소가 함유되어 있다. 이 중 엽록소 a는 빛에너지를 받아 광합성에 이용된다.[9]엽록체의 내부 구조는 전자 현미경을 통해 자세히 밝혀졌다. 엽록체 내부에는 그라나와 스트로마가 존재한다. 그라나는 동전이 여러 개 쌓여 있는 듯한 막 구조로, 틸라코이드 또는 라멜라라고 불리는 평평한 주머니 모양의 구조로 이루어져 있다. 라멜라 막에는 엽록소, 카로티노이드, 지질, 단백질 등이 포함된 쿠온타좀이라는 입자가 박혀 있다. 엽록체는 세포핵과는 독립적으로 분열하여 증식하며, 자체적인 DNA를 가지고 있어 높은 자율성을 가진다. 그러나 엽록체의 기능은 핵 유전자의 지배를 받는 부분도 많다.[10]
육상 식물에서 엽록소를 함유한 색소체는 광합성을 통해 빛 에너지를 화학 에너지로 전환하고, 대기 중 이산화탄소를 고정하며 산소를 방출한다.
5. 2. 에티오플라스트 (Etioplast)
에티오플라스트는 엽록체의 전구체이다. 에티오플라스트는 빛이 없는 환경에서 엽록체가 되기 전 단계의 색소체이다.5. 3. 유색체 (Chromoplast)
유색체는 색소를 합성하고 저장하는 색소체이다. 카로티노이드 등 다양한 색소를 함유하여 꽃잎, 열매 등에 색을 부여한다.[10] 유색체는 미세 구조에 따라 다음과 같이 세 가지로 분류할 수 있다.- 당근 뿌리처럼 결정체를 갖고 있는 것.
- 황엽·홍엽·귤류의 과피 등에 들어 있는 것처럼 프라스토 과립(오스뮴산으로 검게 물드는 공모양의 입자)을 갖고 있는 것.
- 옥수수·피망 등의 과피처럼 섬유상 구조를 한 것.
발생적으로 보면, 당근의 유색체처럼 백색체 내에 카로틴 결정이 축적되어 유색체로 이행하는 경우가 있다. 또한 잎이나 과피의 엽록체처럼 라메라막과 엽록소가 분해 소실하지만 카로티노이드는 잔존하여 유색체가 되는 경우도 있다.
5. 4. 백색체 (Leucoplast)
백색체는 색소가 없는 색소체로, 모노테르펜을 합성한다. 녹말, 지방, 단백질을 포함한 다양한 구성 요소를 저장하는 기능을 한다.[9] 백색체는 더 분화하여 특화된 기능을 가질 수 있는데, 녹말을 저장하고 중력을 감지하는 전분체(Amyloplast), 지방을 저장하는 유지체(Elaioplast), 단백질을 저장하고 수정하는 단백질체(알레우로플라스트)가 그 예시이다.[10]Elaioplast는 오일 합성과 저장에 관여하며 꽃밥 세포에서 pollen coat를 형성하는 기능을 한다. oilseed나 endosperm cell과 같이 기름과 관련된 색소체라고 볼 수 있다. Amyloplast는 감자에서 발견되는 저장 색소체로, 감자에서는 starch를 granule형태로 저장한다. 주로 뿌리나 저장 조직에서 발견되며 식물의 굴지성에 관여한다. 굴지성이란 식물이 중력을 감지하여 중력 방향으로 성장하는 것을 의미한다. 식물이 위아래로 뒤집히면 amyloplast가 밑쪽으로 이동하고 그에 따라 auxin도 밑쪽에서 나오게 되면서 식물이 중력 방향으로 성장하게 된다. (Auxin은 식물의 길이 생장에 관여하는 호르몬이다.)
5. 5. 노화엽록체 (Gerontoplast)
식물 노화 과정에서 광합성 장치가 분해되는 것을 제어하는 색소체이다.[10]5. 6. 탄닌체 (Tannosome)
탄닌과 폴리페놀을 합성하고 생성한다.[10]6. 조류의 색소체
조류는 식물과 마찬가지로 엽록체를 가지지만, 종류에 따라 구조와 색소 조성이 다를 수 있다. 조류의 엽록체는 피레노이드를 포함한다는 점에서 식물의 엽록체와 다르다.
조류의 색소체는 계통에 따라 다른 이름으로 불리기도 한다. 예를 들어, 홍조류에서는 로도플라스트(홍색체), 회색조류나 ''Paulinella chromatophora'' (케르코조아의 유각 아메바)에서는 시아넬이라고 불린다. 또한, 백색체라는 용어는 색소가 없는 모든 색소체를 통칭하며, 이미 퇴화하여 엽록체로의 분화 능력을 잃은 것도 포함한다.
6. 1. 홍조류의 로도플라스트 (Rhodoplast)
홍조류에서 발견되는 붉은색 색소체인 로도플라스트는 최대 268m 해양 깊이까지 광합성을 가능하게 한다.[10] 로도플라스트는 식물의 엽록체와 달리 전분을 색소체 내가 아닌 세포질에 플로리데아 전분 형태로 합성하여 저장한다.[16]6. 2. 회색조류의 무로플라스트 (Muroplast)
무로플라스트는 글라우코조류 조류의 색소체로, 시아노플라스트 또는 시아넬이라고도 불린다. 식물의 엽록체와 유사하지만 세균과 유사한 펩티도글리칸 세포벽을 가지고 있다는 점이 다르다.[10]6. 3. 녹조류의 엽록체 (Chloroplast)
녹조류의 엽록체는 식물의 엽록체와 유사한 구조와 기능을 가지고 있다. 엽록체는 광합성을 담당하는 세포 소기관으로, 엽록소를 포함하고 있어 빛 에너지를 이용하여 이산화탄소와 물로부터 포도당과 같은 유기물을 합성한다.6. 4. 기타
- 백색체: 조류에서 이 용어는 모든 무색소체에 사용된다. 그 기능은 식물의 백색체와는 다르다.
- 아피코플라스트: 2차 공생에서 유래한 아피콤플렉사의 비광합성 색소체이다.
7. 색소체의 유전
대부분의 식물은 번식 과정에서 한쪽 부모로부터만 색소체를 물려받는다. 일반적으로 속씨식물은 암컷 배우자로부터 색소체를 물려받으며, 많은 겉씨식물은 수컷 꽃가루로부터 색소체를 물려받는다. 조류 또한 한쪽 부모로부터만 색소체를 물려받는다. 따라서 다른 부모의 색소체 DNA는 완전히 소실된다.[21]
정상적인 종내 교배(한 종의 정상적인 잡종 생산)에서 색소체 DNA의 상속은 엄격하게 단일 부모, 즉 암컷으로부터 이루어지는 것으로 보인다. 그러나 종간 잡종화에서는 상속이 더 불규칙하게 나타난다. 종간 잡종화에서 색소체는 주로 암컷으로부터 상속되지만, 수컷으로부터 색소체를 생성하는 속씨식물의 잡종에 대한 많은 보고가 있다.[21]
알팔파(''Medicago sativa'')를 포함한 약 20%의 속씨식물은 일반적으로 양쪽 부모로부터 색소체를 물려받는 것으로 나타난다.[21]
8. 색소체 DNA 손상 및 복구
색소체 DNA는 묘목이 성장함에 따라 손상 정도가 증가한다.[22] DNA 손상은 광산화 반응과 광합성/호흡 전자 전달에 의해 생성된 산화 환경으로 인해 발생한다. 일부 DNA 분자는 DNA 복구되지만, 복구되지 않은 손상된 DNA는 기능이 없는 조각으로 분해되는 것으로 보인다.
DNA 복구 단백질은 세포의 핵 게놈에 의해 암호화된 후 색소체로 이동하여 색소체의 DNA를 복구함으로써 게놈의 안정성/온전성을 유지한다.[23] 예를 들어, 이끼 ''Physcomitrella patens''의 엽록체에서 DNA 불일치 복구에 사용되는 단백질(Msh1)은 재조합 복구에 사용되는 단백질(RecA 및 RecG)과 상호 작용하여 색소체 게놈 안정성을 유지한다.[24]
9. 기원
색소체는 내부 공생을 한 시아노박테리아에서 유래한 것으로 생각된다. 고세균식물(Archaeplastida)의 주요 내부 공생 사건은 약 15억 년 전에 발생한 것으로 추정되며,[25] 진핵생물이 산소 발생 광합성을 수행할 수 있게 했다.[26] 이후 고세균식물에서 세 개의 진화 계통이 출현했으며, 그 중 색소체는 녹조류 및/또는 식물에서는 엽록체, 홍조류에서는 홍조체, 회색조식물에서는 무로플라스트로 다르게 명명된다. 이들은 착색과 미세 구조 모두에서 차이가 있다. 예를 들어, 식물과 녹조류의 엽록체는 시아노박테리아, 홍조류, 회색조식물에서 발견되는 광 수확 복합체인 모든 피코빌리소좀을 잃었지만, 대신 스트로마와 그라나 틸라코이드를 포함한다. 회색조식물 색소체는 엽록체와 홍조체와 달리 시아노박테리아 세포벽의 잔해에 여전히 둘러싸여 있다. 이러한 1차 색소체는 모두 두 개의 막으로 둘러싸여 있다.
복잡한 색소체는 2차 내부 공생에 기인하며, 이 과정에서 진핵생물이 홍조류 또는 녹조류를 삼켜 그 색소체를 유지한다.[27] 홍조류를 2차 공생시킨 복잡한 색소체를 가진 조류에는 이형편모조류, 갈조류, 은편모조류 및 대부분의 와편모조류가 있다. 녹조류를 2차 공생시킨 것(엽록체)으로는 유글레나류와 클로라라크니온 조류가 있다. 아피콤플렉사(말라리아(`Plasmodium`속), 톡소플라스마증(`Toxoplasma gondi`)을 비롯한 많은 인간과 동물의 병의 병원체를 포함하는 절대 기생 원생생물 문)도 복잡한 색소체를 가지고 있다. 아피코플라스트라고도 불리는 이 색소체는 광합성 능력을 잃었지만 필수적인 세포 소기관이며, 항기생충 약을 개발하기 위한 표적으로 유망하다.
어떤 종류의 와편모조류나 일부 섬모충은 다른 조류를 섭취하여 광합성의 혜택을 받기 위해 소화한 조류의 색소체를 보존하고 있다. 잠시 후 색소체도 소화된다. 이러한 붙잡힌 색소체는 클렙토클로로플라스트라고 불린다.
참조
[1]
서적
The Structure and Function of Plastids
Springer Netherlands
2007
[2]
학술지
An Expanded Ribosomal Phylogeny of Cyanobacteria Supports a Deep Placement of Plastids
2019
[3]
학술지
The monoplastidic bottleneck in algae and plant evolution
https://jcs.biologis[...]
2018-01-15
[4]
학술지
The ancestor of the Paulinella chromatophore obtained a carboxysomal operon by horizontal gene transfer from a Nitrococcus-like γ-proteobacterium
2007-06-05
[5]
학술지
The plastid ancestor originated among one of the major cyanobacterial lineages
2014-01-29
[6]
간행물
Ueber die Gestalten der Stärkebildner und Farbkörper
https://www.biodiver[...]
1882
[7]
간행물
Morphologische Individuen erster Ordnung: Plastiden oder Plasmastücke
https://www.biodiver[...]
1866
[8]
웹사이트
Picozoans Are Algae After All: Study
https://www.the-scie[...]
[9]
서적
Biosynthetic pathways of cutin and waxes, and their sensitivity to environmental stresses
BIOS Scientific publishers Ltd.
1996
[10]
서적
The Structure and Function of Plastids
Springer
2006
[11]
학술지
The evolution of the plastid chromosome in land plants: gene content, gene order, gene function
2011
[12]
학술지
Molecular evolution of plastid genomes in parasitic flowering plants
https://www.scienced[...]
2018
[13]
웹사이트
Plants Without Plastid Genomes
http://www.the-scien[...]
2015-09-26
[14]
학술지
Why are plastid genomes retained in non-photosynthetic organisms?
2006-02
[15]
웹사이트
DNA of Giant 'Corpse Flower' Parasite Surprises Biologists
https://www.quantama[...]
2021-04
[16]
학술지
The unique features of starch metabolism in red algae
2001-07
[17]
학술지
Evolutionary dynamics of the chromatophore genome in three photosynthetic Paulinella species
2019-02-22
[18]
학술지
Paulinella , a model for understanding plastid primary endosymbiosis
Wiley
2020-05-05
[19]
학술지
Early photosynthetic eukaryotes inhabited low-salinity habitats
2017-09-12
[20]
Q
[21]
학술지
Why does biparental plastid inheritance revive in angiosperms?
2010-03
[22]
학술지
Changes in DNA damage, molecular integrity, and copy number for plastid DNA and mitochondrial DNA during maize development
2014-12
[23]
학술지
DNA maintenance in plastids and mitochondria of plants
2015
[24]
학술지
MSH1 maintains organelle genome stability and genetically interacts with RECA and RECG in the moss Physcomitrella patens
2017-08
[25]
학술지
The plastid ancestor originated among one of the major cyanobacterial lineages
2014-09
[26]
학술지
A molecular timescale of eukaryote evolution and the rise of complex multicellular life
2004-01
[27]
학술지
The Origin of Plastids
http://www.nature.co[...]
[28]
학술지
A Suggested Cycle of Plastid Developmental Interrelationships
1978
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com