2025. 7. 13. 오후 5:27:00
‘공룡 연구’ 허민 신임 국가유산청장…고생물학자로는 첫 청장
출처: 경향 신문 ( 한국 / 한국어 )
익룡은 중생대에 번성했던 하늘을 나는 파충류로, 뼈가 속이 비어 가볍고 공기로 채워져 비행에 적합한 구조를 가졌다. 초기 익룡은 긴 꼬리를 가졌지만, 후기 익룡은 꼬리가 짧아지고 다양한 크기와 형태를 보이며, 날개는 피부막으로 이루어져 있었다. 익룡은 다양한 식성을 가졌으며, 볏과 털과 같은 특징을 보이기도 했다. 최초의 익룡 화석은 1784년에 발견되었으며, 1960년대 이후 익룡 연구가 활발하게 이루어졌다. 익룡은 다양한 분류군으로 나뉘며, 멸종 원인에 대해서는 여러 가설이 존재한다. 익룡은 대중문화에서 공룡만큼 자주 등장하지는 않지만, 영화, 소설 등에서 묘사되며, 종종 실제와 다른 모습으로 묘사되기도 한다.
| 익룡 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Pterosauria |
| 명명자 | Kaup, 1834 |
| 동의어 | Ornithosauria Seeley, 1870 |
| 어원 | 그리스어 pteron(날개) + sauros(도마뱀) |
| 의미 | 날개 도마뱀 |
| 분포 | 화석 발견 위치 지도 참고 |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 파충강 |
| 아강 | 쌍궁아강 |
| 미분류 | 주룡형류 |
| 미분류 | 주룡류 |
| 미분류 | 조경류 |
| 미분류 | 익룡형류 |
| 목 | 익룡목 (Pterosauria) |
| 하위 분류군 | |
| 특징 | |
| 크기 | 다양함 |
| 가장 큰 익룡 | 케찰코아틀루스, 하체고프테릭스 |
| 깃털 유무 | 깃털 존재 |
| 생존 시기 | |
| 시기 | 트라이아스기 후기 - 백악기 후기 ( 백만 년 전) |
| 참고자료 | |
| 웹사이트 | Pterosaur.net :: 육상 이동 |
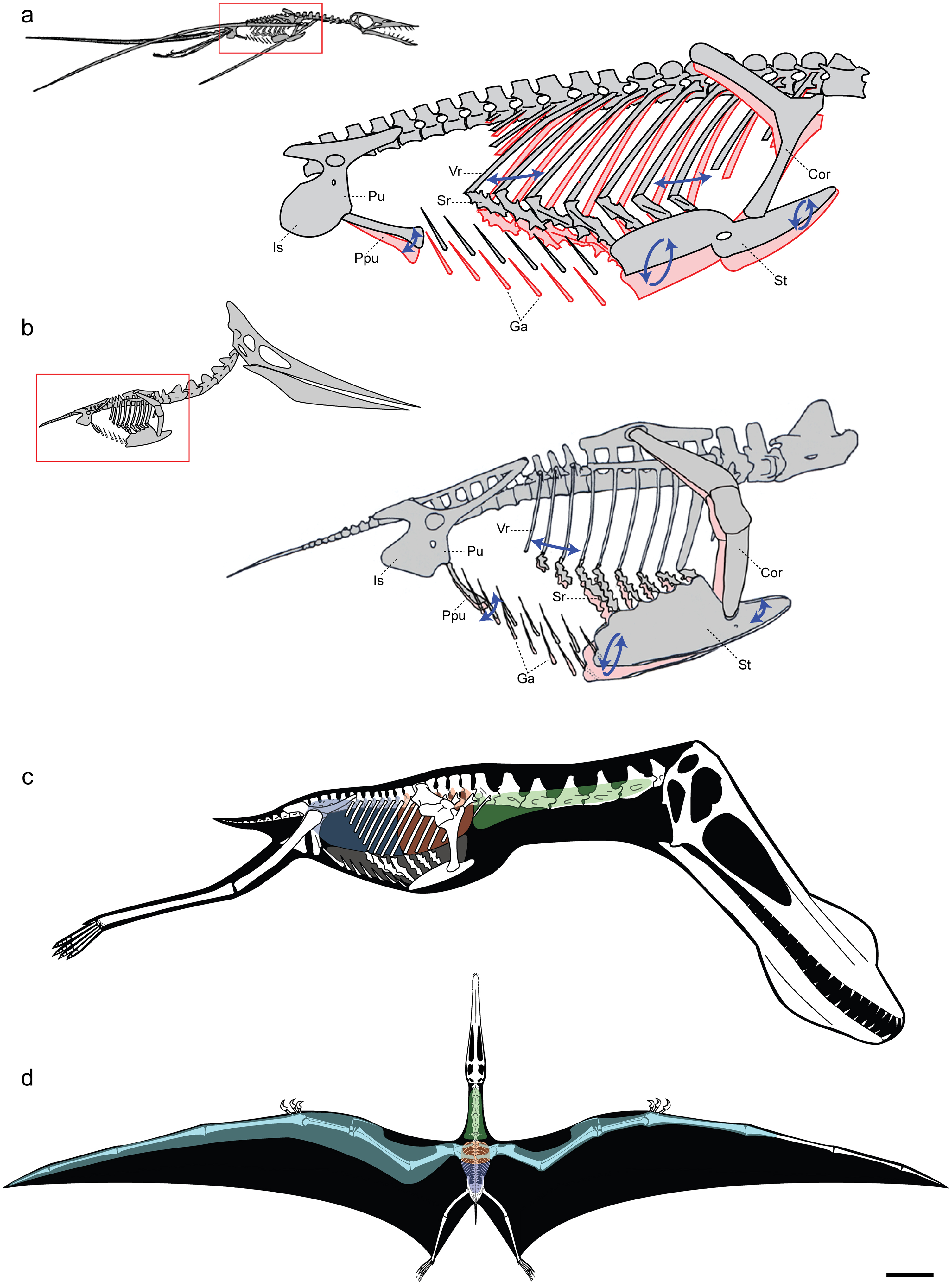
익룡은 해부학적으로 비행을 위해 조상인 파충류로부터 많은 변화를 겪었다. 익룡의 골격은 새의 뼈와 마찬가지로 속이 비어 공기로 차 있었다. 가슴뼈에는 비행 근육이 부착될 수 있도록 용골돌기가 발달했으며, 커다란 뇌는 비행에 특화된 기능을 가졌던 것으로 보인다.[171] 후기 익룡에서는 어깨 위 척추뼈가 합쳐져 '노타리움'(notarium)이라는 뼈가 되어 비행 중 몸통을 단단하게 해주고 견갑골을 안정적으로 지지해 주었다.
익룡의 뼈는 조류처럼 속이 비어 공기로 채워져 가벼우면서도 근육 부착 면적이 넓었다. 뼈 벽은 종이처럼 얇은 경우도 많았다. 익룡은 비행 근육을 위한 크고 융기된 흉골과 복잡한 비행 행동을 조정하는 확대된 뇌를 가졌다.[18] 익룡 골격은 융합된 경우가 많아, 두개골 구성 요소 사이 봉합이 사라지기도 했다. 일부 후기 익룡은 어깨 위 척추가 노타리움으로 융합되어 비행 중 몸통을 뻣뻣하게 하고 어깨뼈를 안정적으로 지지했다. 천추 뼈는 단일 합선골을, 골반뼈도 융합될 수 있었다.
기저 익룡에는 디모르포돈, 에우디모르포돈, 캄필로그나토이데스, 람포린쿠스, 스카포그나투스 등이 포함된다. 프테로닥틸로이드에는 이스티오닥틸루스, 오르니토케이루스, 프테라노돈, 크테노카스마, 프테로닥틸루스, 게르마노닥틸루스, 둥가리프테루스, 타페자라(익룡), 투푹수아라, 케찰코아틀루스 등이 포함된다. 두 그룹은 시간상 겹치지만, 화석 기록상 초기 익룡은 기저 익룡이고, 후기 익룡은 프테로닥틸로이드이다.[19]
안우로그나투스, 제홀로프테루스, 베스페로프테릴루스를 포함하는 안우로그나투스과는 논쟁의 여지가 있다.[20] 이들은 짧은 턱과 넓은 입을 가진 작은 비행체로, 일부는 야행성 또는 황혼형 습성을 시사하는 큰 눈, 입 수염, 매달리기 적응된 발을 가졌다. 이는 비행 중 곤충을 잡아먹는 조류 및 박쥐와 유사한 적응이다.
익룡은 일반적으로 컸지만, 다양한 크기를 보였다. 가장 작은 종은 날개 길이가 25cm 이상이었고,[21] 가장 큰 종은 날개 길이가 최대 10m에서 11m에 달해, 알려진 가장 큰 비행 동물에 속한다.[22] 거대 익룡은 서 있을 때 기린 키에 이를 수 있었다. 익룡은 크기에 비해 매우 가벼울 것으로 추정되었으나, 이후 연구에서는 최대 250kg까지 나갈 수 있다고 추정한다.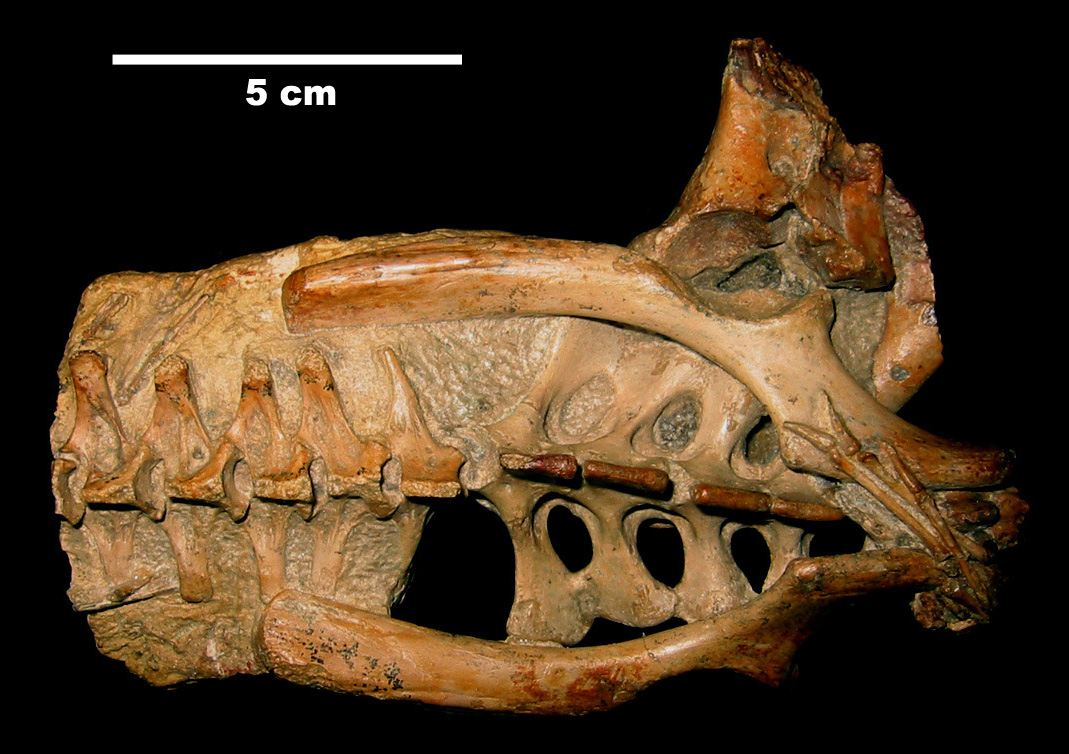
익룡의 골반은 중간 크기였고, 세 개의 골반 뼈가 융합된 경우가 많았다. 장골은 길고 낮았으며, 앞뒤 날은 아래쪽 골반 뼈 가장자리 너머로 수평으로 튀어나와 뒷다리 근육 힘을 제한했다. 좁은 두덩뼈는 넓은 궁둥뼈와 융합되어 궁둥두덩뼈 날을 형성했고, 때로는 양쪽 날이 융합되어 골반관을 형성했다. 고관절은 뚫려 있지 않아 다리 운동성을 허용했고, 비스듬히 위쪽으로 향해 다리의 수직 위치를 방지했다. 두덩뼈 앞쪽은 독특한 쌍을 이룬 전두덩뼈와 연결되어 골반과 배 갈비뼈 사이 뒷배를 덮는 뾰족한 부분을 형성했다. 이 요소의 수직 운동성은 가슴강 강성을 보완해 호흡에 기여했을 것으로 보인다.
익룡 뒷다리는 튼튼했지만, 날개 길이에 비해 새보다 작았다. 몸통 길이에 비해 다리는 길었다. 넙다리뼈는 비교적 곧았고, 머리 부분은 몸통과 작은 각도를 이루어 다리가 약간 벌어진 상태였다. 종아리뼈는 위 발목뼈와 융합되어 넙다리뼈보다 긴 경골을 이루었고, 걸을 때 수직 자세를 취할 수 있었다. 종아리뼈는 가늘어지는 경향이 있었고, 진화된 형태에서는 길이가 1/3로 줄어들기도 했다. 종아리뼈는 정강이뼈와 융합된 경우가 많았다. 발목은 단순한 "중족골" 경첩이었다. 길고 가느다란 중족골은 항상 어느 정도 벌어져 있었다. 발은 족저보행을 해, 걷는 동안 중족골 바닥이 땅에 닿았다.
초기 익룡과 진화된 종 사이에는 다섯 번째 발가락 형태에 차이가 있었다. 원래 다섯 번째 중족골은 튼튼했고 짧지 않았으며, 다른 중족골보다 높은 위치에서 발목에 연결되었다. 두 마디로 구성된 길고 구부러진, 발톱 없는 다섯 번째 발가락을 지지했다. 이 발가락은 한때 매달려 자는 데 사용되거나 팔꿈치막을 늘리는 데 사용되었을 것으로 추정되었으나, 이후 유로파타기움 또는 크루로파타기움을 확장시켜 비행 중 방향타 역할을 했을 것으로 추정된다. 일부 표본은 발가락 사이에 막을 보여줘,[116] 비행 제어 표면으로 기능했을 수 있다. 프테로닥틸로이드에서는 다섯 번째 중족골이 크게 감소했고, 다섯 번째 발가락은 짧은 뼈 조각에 불과했다. 이는 막이 갈라져 비행 기동성이 증가했음을 시사한다. 첫 번째에서 네 번째 발가락은 길었고, 각각 2, 3, 4, 5개 마디가 있었다. 세 번째 또는 네 번째 발가락이 가장 길었다. 평평한 관절은 제한된 이동성을 나타냈고, 발톱은 손톱보다 작았다.
익룡 유해 화석화를 가능하게 한 희귀 조건은 때때로 연조직도 보존했다. 싱크로트론이나 자외선 촬영법은 육안으로 보이지 않는 많은 흔적을 밝혀냈다. 이는 뿔 볏, 부리, 발톱 외피, 비막 등을 포함하며, 드물게 근육도 보존되었다. 피부 반점은 발바닥, 발목, 중족골 끝 부분에 작고 겹치지 않는 비늘을 보여주며, 걷는 동안 충격을 완화하는 패드를 덮고 있었다. 비늘은 신체 다른 부분에서는 알려져 있지 않다.
익룡은 최초로 하늘을 난 척추동물이다. 조류만큼 능숙하게 하늘을 날거나 걷지는 못했다. 공룡처럼 트라이아스기에 나타나 백악기 말에 멸종했다. 크기는 작은 새 정도부터 날개 길이 12m가 넘는 것까지 다양했다. 모두 큰 머리와 날개, 작은 몸통을 가졌고, 긴 꼬리가 있거나 없는 것도 있었다.
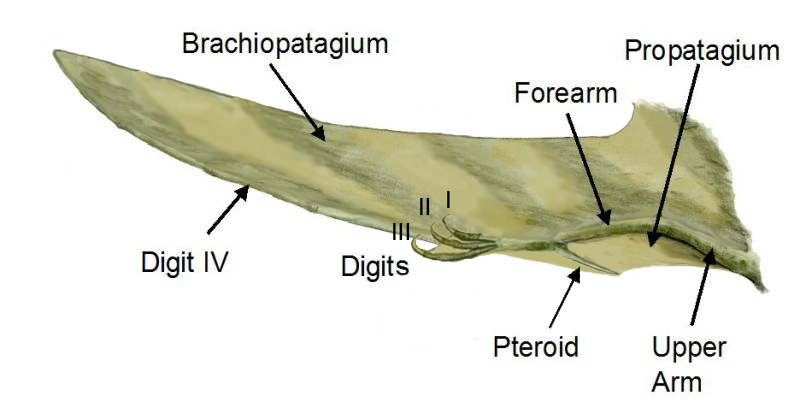
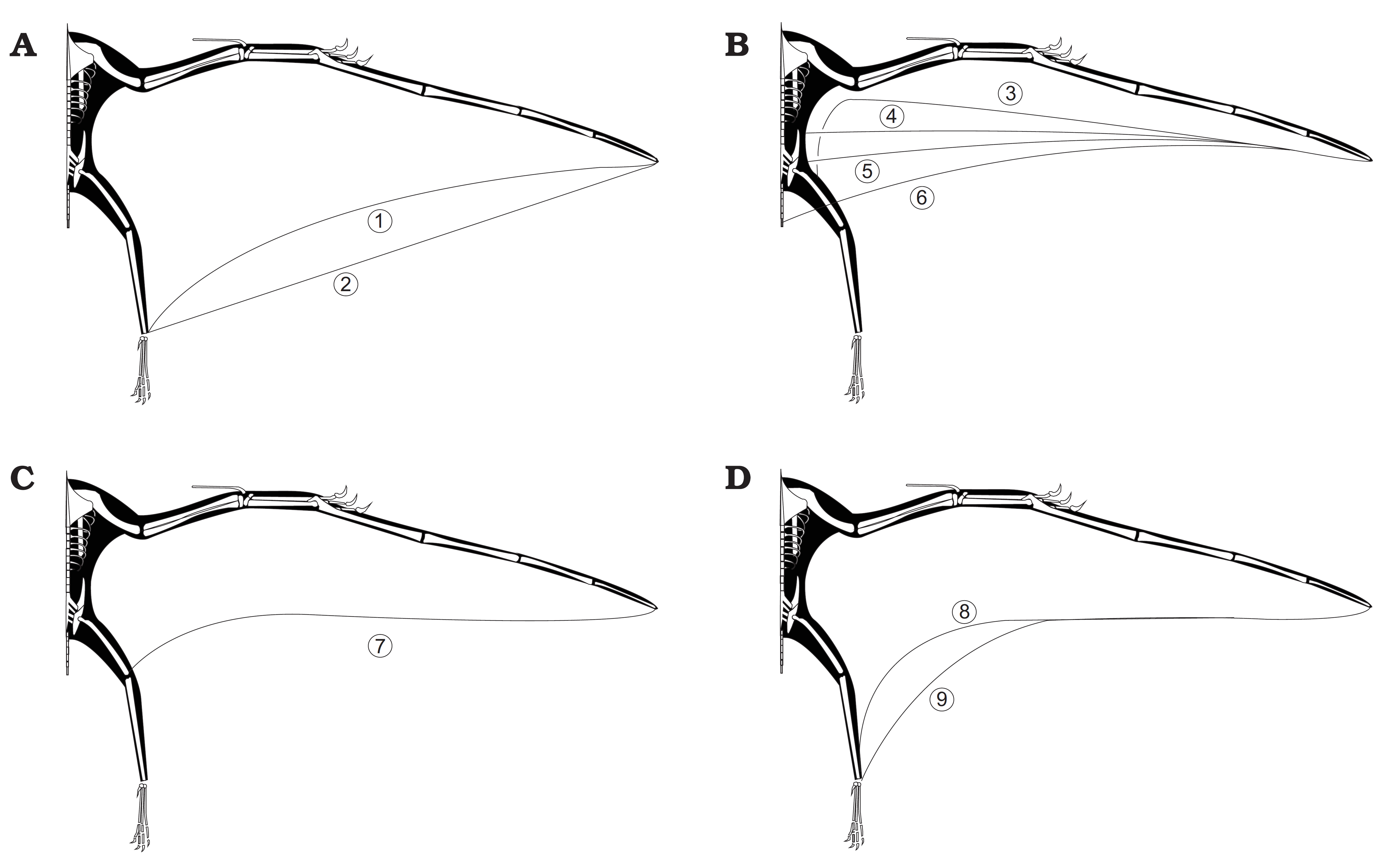
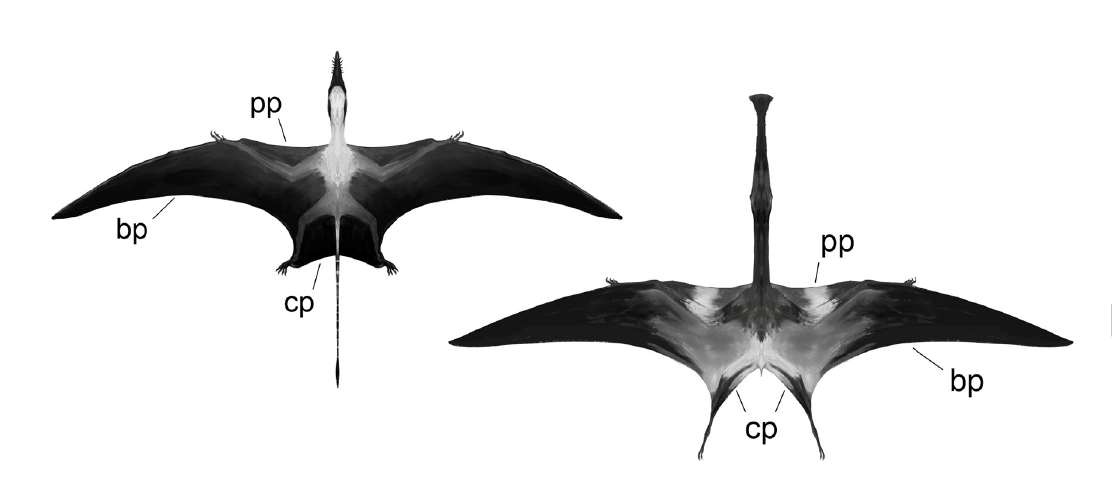
날개는 막 구조였을 것으로 생각된다. 박쥐 날개와 비슷하게 길게 뻗은 앞발 손가락에 얇은 막을 펼치는 구조다. 박쥐는 엄지손가락을 제외한 모든 손가락이 막을 지지하지만, 익룡 날개는 넷째 손가락(다섯째 손가락은 퇴화)과 다리 사이에만 막이 쳐져 있었다. 날개에서 독립된 손가락이 많아 박쥐보다 자유롭게 물건을 잡았겠지만, 손가락 하나만으로 막을 지지하는 날개는 비행 자유로움 등에서 박쥐에 미치지 못했을 것이다. 그러나 비막에 신경과 근육이 뻗어 있었을 것으로 보이는 흔적이 있어, 막 형태를 변화시켜 고도 비행 제어를 했을 가능성도 있다. 보행이나 지상 활동은 조류에 비해 뒤떨어졌지만, 발자국 연구에서 익룡이 척행성 사족 보행을 했으며, 지상에서 기어 다니는 박쥐보다 지상 적응성이 높았음이 시사되었다.
등심뼈를 가졌고, 골격이 속 빈 가볍고 공기를 함유한 뼈로 이루어진 점은 조류와 같다. 공기 함유 뼈를 가졌다는 점에서 조류처럼 기낭도 갖췄을 가능성이 높다. (공룡도 기낭을 가졌으며, 공룡과 익룡 공통 조상 단계에서 이미 기낭을 획득했을 가능성이 높다.) 체중은 매우 가벼워, 날개 길이 10m가 넘는 케찰코아틀루스도 70kg 정도였을 것이다. 익룡 화석에서도 공룡·조류와 유사한 깃털이 발견되었으며[164], 공룡류 공통 조상(조경류) 단계에서부터 깃털을 가졌을 것으로 생각된다. 하늘에서는 역광으로 잘 보이지 않아 색깔은 현재 바닷새와 비슷하며 화려한 색상은 적고, 등은 자외선을 막기 위해 검은색, 복부는 색소 절약으로 흰색이었다는 설도 있다. 볏이 있는 종도 많으며, 암수 간 의사소통이나 동종과 별종 구별 등에 사용되었다는 설이 있다. 쿤펭고프테루스 위에서 발견된 펠릿을 통해, 조류 등에서 보이는 소화되지 않은 것을 덩어리로 토해내는 습성을 가졌음이 시사되었다.[165][166]
## 두개골, 이빨, 볏
익룡 두개골은 새나 박쥐에 비해 일반적으로 컸다. 대부분 익룡 두개골은 길쭉한 턱을 가졌으며, 성체는 두개골 뼈가 융합되는 경향이 있었다. 초기 익룡은 종종 이형치를 가졌고 구조가 다양했으며, 일부는 구개에 이빨이 있었다. 후기 그룹에서는 이빨이 대부분 원뿔 모양이 되었다.[97] 앞니는 종종 더 길어 가로로 확장된 턱 끝에서 "먹이 획득"을 형성했지만, 크기와 위치는 종마다 다양했다. 파생된 익룡류에서는 두개골이 더욱 길어져 목과 몸통 길이를 합친 것보다 길어지기도 했다. 이는 앞쪽 주둥이 뼈인 전상악골이 위턱 뼈인 상악골과 뻗어 융합되면서 발생했다. 시조류와 달리, 익룡류 익룡의 비공과 안와전공은 ''비안와공''이라는 단일 큰 구멍으로 합쳐졌다. 이는 비행을 위해 두개골 무게를 줄이기 위해 진화했을 수 있다.[97] 반대로, 안와 뒤 뼈는 수축하고 회전하여 뒷머리를 기울이고 턱 관절을 앞으로 가져왔다. 뇌상자는 파충류치고는 비교적 컸다.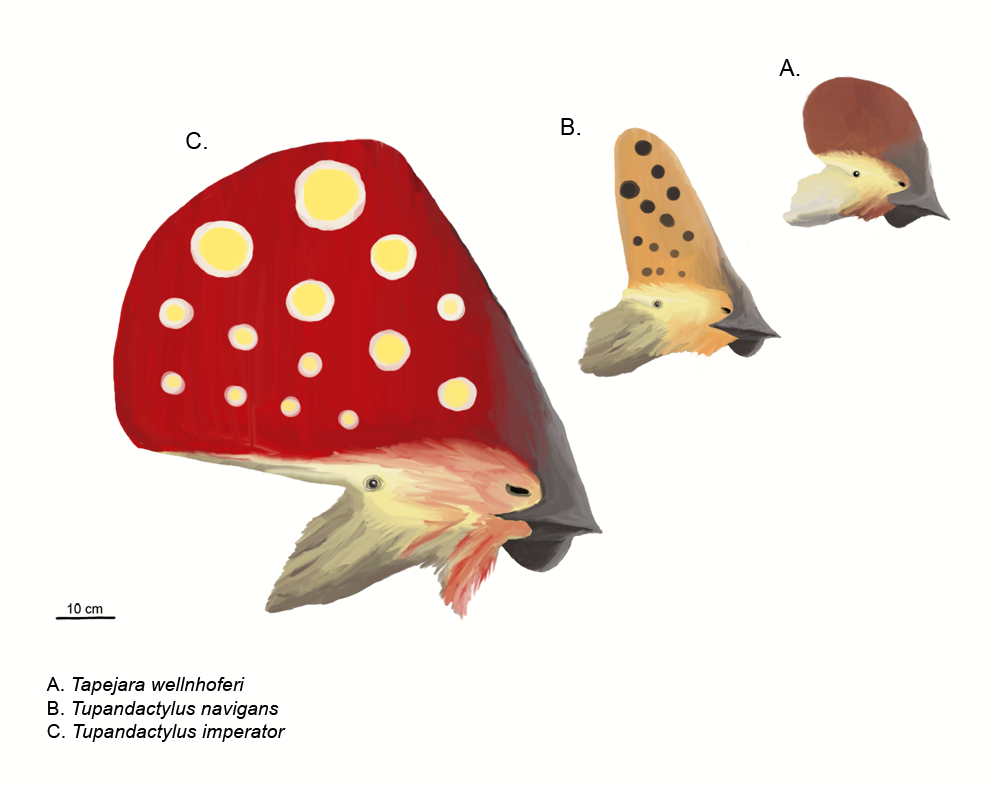
화석화된 케라틴질 부리 조직이 보존된 경우도 있지만, 이빨이 있는 형태에서는 부리가 작고 턱 끝에 국한되며 이빨과 관련이 없었다.[23] 일부 진화된 부리 형태는 프테라노돈과와 아즈다르코과처럼 이빨이 없었고 더 크고 광범위하며 새와 같은 부리를 가졌다.[97] 일부 그룹은 특화된 이빨 형태를 가졌다. 이스티오닥틸루스과는 고기를 먹기 위해 구부러진 이빨을 가졌다. 크테노카스마과는 여과 섭식을 위해 수많은 바늘 모양 이빨 빗을 사용했다. ''프테로다우스트로''는 천 개 이상 털 모양 이빨을 가졌을 수 있다. 둥가리프테루스과는 턱뼈 조직으로 이빨을 덮어 으깨는 기능을 했다. 이빨이 있는 경우, 이빨은 별도 치아 소켓에 위치했다. 교체 이빨은 오래된 이빨 아래가 아닌 뒤쪽에서 생성되었다.
익룡의 대중적 이미지는 정교한 머리 볏으로 정의된다. 이는 ''프테라노돈''의 뒤쪽을 향한 볏의 영향을 받았다. 볏 주요 위치는 주둥이 앞쪽(전상악골 성장) 또는 두개골 뒤쪽(두정골 확장)으로, 이 경우 "상후두정골 볏"이라 불린다. 앞쪽 볏과 뒤쪽 볏이 동시에 존재할 수 있으며 단일 더 큰 구조로 융합될 수 있고, 가장 확장된 형태는 테이페자리과에서 나타난다. ''닉토사우루스''는 뿔 모양 볏을 가졌다. 볏은 가로 방향으로 몇 밀리미터 두께였다. 뼈대 볏 기저부는 케라틴 등으로 확장될 것이다.
1990년대 이후, 새로운 발견과 오래된 표본 연구를 통해 볏이 이전에 생각했던 것보다 익룡에게 더 널리 퍼져 있음이 밝혀졌다. 케라틴으로 확장되었거나 구성되었기 때문에 이전 연구를 오도했다.[27] ''프테로린쿠스''와 ''프테로닥틸루스''의 경우, 볏 실제 범위는 자외선 사진을 통해서야 밝혀졌다.[23][24] 화석 볏은 더 진화된 익룡류에 국한되었지만, ''프테로린쿠스''와 ''오스트리아다크틸루스''는 초기 익룡도 이를 가졌다는 것을 보여준다.[27]
## 피크노파이버

최소한 몇몇 익룡들은 '피크노파이버'라고 알려진 가는 실 모양 털 같은 것을 머리와 몸에 가졌다. 포유류 털과는 유사하지만 상동(homologous)은 아니었을 것이다. 1831년 골드퍼스(Georg August Goldfuss)에 의해 처음 보고되었고,[185], 최근 발견된 표본들과 조직학 기술 발전 및 자외선 검사로 익룡이 피크노파이버 외피를 가졌다는 증거가 확실해졌다. 피크노파이버는 포유류 털과 다른, 겉보기에 유사하게 발달한 독특한 구조다. 소르데스(Sordes pilosus, 털이 많은 악마)나 제홀옵테루스(Jeholopterus ninchengensis) 화석에는 머리와 몸에 피크노파이버 자국이 남아있어 수렴진화의 예시로 박쥐와도 유사하다.[179]
일부 연구자들은 피크노파이버가 원시깃털 선행형태가 아닌가 생각하지만, 익룡 외피는 마니랍토라 공룡 깃털과 다르다.[185] 피크노파이버는 유연하고 짧은 섬유질로, "어떤 표본에서는 5-7mm 정도"이며, "중앙 관(canal)을 제외하면 그 어떤 내부구조도 없는" 단순한 구조다.[185] 익룡 "털가죽"은 "화석화된 포유류에서 발견된 것과 유사하게 섬유로 촘촘하게 덮인 상태로 보존되어" 있어, 중생대 포유류 털가죽과 비견될 만한 두께였을 것이다.
피크노파이버와 비행은 익룡이 내열성(온혈)이었을 가능성을 높인다. 익룡 날개에 피크노파이버가 없는 것은 털에 기체역학적 기능이 없었으며, 내열성 신진대사에 의해 발생된 열을 보존하는 단열이 체온조절에 중요했음을 뒷받침한다.[185]
익룡 "털"은 포유류 털가죽 등과 뚜렷하게 구분되어 "피크노파이버"(pycnofiber영어)라는 이름이 붙었다. "촘촘한 섬유"라는 의미로, 2009년 고생물학자 알렉산더 켈너(Alexander W. A. Kellner)와 동료들 논문에서 처음 만들어졌다.[187]


최초의 익룡 화석은 1784년 이탈리아인 박물학자인 코시모 콜리니에 의해 기재되었다. 콜리니는 익룡 표본을 바다에 살던 동물로, 긴 앞다리를 노처럼 사용했던 것으로 잘못 해석했다.[188] 몇몇 과학자들은 1830년에 독일 동물학자 요한 게오르그 바글러가 프테로닥틸루스의 날개가 물갈퀴처럼 이용되었을 것이라는 제안을 할 때까지 이 해석을 지지했다.[189] 1801년에 조르쥬 퀴비에가 처음으로 익룡이 하늘을 날던 동물이라고 제안했으며,[190] 1809년에는 독일에서 발견된 표본에 "프테로-닥틸"이라는 이름을 붙였다.[191] 학명이 표준화되면서 이 속의 이름은 프테로닥틸루스(''Pterodactylus'')가 되었지만, "프테로닥틸"이라는 이름은 대중적으로 계속 쓰였고 익룡 전체를 가리키는 명칭으로도 부정확하게 쓰이고 있다.[192] 고생물학자들은 "프테로닥틸"이라는 단어를 사용하는 대신 "익룡(pterosaur)"이라는 용어를 선호한다. "프테로닥틸"이라는 용어는 "프테로닥틸루스" 속에 속하는 익룡들, 혹은 좀 더 넓은 의미로 프테로닥틸루스아목의 익룡을 가리키는 것으로 사용된다.[193]
익룡 화석은 가벼운 뼈 구조로 인해 매우 희귀하다. 완전한 골격은 일반적으로 예외적인 보존 상태를 가진 지층, 즉 소위 ''라거슈테텐''에서만 발견될 수 있다. 1784년, 이탈리아의 박물학자 코시모 알레산드로 콜리니가 최초로 익룡 화석을 기술했다. 당시 진화와 멸종에 대한 개념은 불완전하게 발전되어 있었다. 익룡의 기이한 구조는 기존 동물 그룹에 명확하게 할당할 수 없었기 때문에 충격적이었다. 따라서 익룡의 발견은 현대 고생물학과 지질학의 발전에 중요한 역할을 하게 되었다. 당시의 과학적 의견은 그러한 생물이 아직 살아 있다면 바다만이 믿을 만한 서식지라는 것이었다. 콜리니는 긴 앞다리를 노처럼 사용하는 수영 동물일 것이라고 제안했다. 몇몇 과학자들은 1830년까지 수생 생물 해석을 계속 지지했는데, 독일의 동물학자 요한 게오르크 바글러는 ''프테로닥틸루스''가 날개를 지느러미로 사용했으며 어룡류 및 플레시오사우루스와 관련이 있다고 제안했다.
1800년, 요한 헤르만은 조르주 퀴비에에게 보낸 편지에서 이것이 나는 생물을 나타낸다고 처음 제안했다. 퀴비에는 1801년에 동의하며 이것이 멸종된 나는 파충류라는 것을 이해했다. 1809년, 그는 "날개-손가락"이라는 이름인 ''프테로-닥틸''을 만들었다. 이것은 1815년에 라틴어로 ''프테로닥틸루스''로 바뀌었다. 처음에는 대부분의 종이 이 속으로 지정되었고, 궁극적으로 "익룡"은 익룡류의 모든 구성원에게 대중적이고 부정확하게 적용되었다. 오늘날 고생물학자들은 이 용어를 프테로닥틸로이드의 속인 ''프테로닥틸루스'' 또는 구성원으로 제한한다.
1812년과 1817년, 사무엘 토마스 폰 죔머링은 원래 표본과 추가 표본을 재기술했다. 그는 그것들을 새와 박쥐와 관련이 있다고 보았다. 비록 그는 이 점에서 실수했지만, 그의 "박쥐 모델"은 19세기에 영향력이 있었다. 1843년, 에드워드 뉴먼 (곤충학자)은 익룡이 나는 유대류라고 생각했다. 아이러니하게도 "박쥐 모델"은 익룡을 온혈 동물이고 털이 있는 것으로 묘사했기 때문에, 장기적으로 퀴비에의 "파충류 모델"보다 특정 측면에서 더 정확한 것으로 밝혀졌다. 1834년, 요한 야코프 카우프는 익룡류라는 용어를 만들었다.
1828년, 메리 애니는 독일 외 지역에서 최초의 익룡 속을 발견했는데, 이는 리처드 오웬에 의해 ''디모르포돈''(Dimorphodon)으로 명명되었으며, 알려진 최초의 비-익룡류 익룡이기도 하다. 그 후 세기 동안, 초기 백악기 케임브리지 그린샌드에서 수천 개의 익룡 화석이 발견되었지만, 대부분 심하게 침식된 조각들로 구성되어 품질이 좋지 않았다. 그럼에도 불구하고, 이를 바탕으로 수많은 속과 종이 명명되었다. 당시 이 분야의 주요 영국 전문가였던 해리 고비어 실리는 많은 종을 묘사했으며, 최초의 익룡 책인 ''Ornithosauria'',[57] 1901년 최초의 대중서인 ''Dragons of the Air''를 저술하기도 했다. 실리는 익룡을 온혈적이고 역동적인 생물로, 새와 밀접한 관련이 있다고 생각했다.[58] 이전에는 진화론자 세인트 조지 잭슨 미버트가 익룡이 새의 직접적인 조상이라고 제안했었다.[59] 오웬은 두 사람의 견해에 반대하며, 익룡을 냉혈적인 "진정한" 파충류로 보았다.
미국에서는 1870년 오스니엘 찰스 마시가 니오브라라 백악에서 ''프테라노돈''(Pteranodon)을 발견했는데, 당시 알려진 가장 큰 익룡이었으며, 최초의 이빨 없는 익룡이자 미국에서 발견된 최초의 익룡이었다. 이 지층에서도 수천 개의 화석이 산출되었으며, 솔른호펜 표본처럼 심하게 압착되지 않고 3차원적으로 보존된 비교적 완전한 골격도 포함되어 있었다. 이는 뼈의 속이 빈 특징과 같은 많은 해부학적 세부 사항에 대한 훨씬 더 나은 이해로 이어졌다.
한편, 솔른호펜에서 발견된 표본들은 계속 이어졌으며, 고품질의 완전한 표본 대부분을 차지했다. 이들은 ''람포린쿠스''(Rhamphorhynchus), ''스카포그나투스''(Scaphognathus) 및 ''도리그나투스''(Dorygnathus)와 같은 새로운 기저 분류군을 식별할 수 있게 했다. 이 자료는 독일 익룡 연구 학파를 탄생시켰으며, 이 학파는 날아다니는 파충류를 현대 박쥐와 새의 온혈적이고 털이 많으며 활동적인 중생대 대응물로 보았다. 1882년, 마시와 칼 알프레트 치텔은 ''람포린쿠스'' 표본의 날개 막에 대한 연구를 발표했다.[60][61] 독일 연구는 1930년대까지 계속되었으며, ''아누로그나투스''(Anurognathus)와 같은 새로운 종을 묘사했다. 1927년, 페르디난트 브로일리는 익룡 피부에서 털집을 발견했으며,[62] 고신경학자 틸리 에딩거는 익룡의 뇌가 현대의 냉혈 파충류보다 새의 뇌와 더 유사하다는 것을 밝혀냈다.[63]
20세기 중반까지 영국과 미국의 고생물학자들은 익룡에 대한 관심을 크게 잃었다. 그들은 익룡을 실패한 진화 실험으로, 냉혈적이고 비늘이 있으며, 날기 어려우며, 더 큰 종은 활공만 가능하고, 이륙하기 위해 나무에 오르거나 절벽에서 몸을 던져야 한다고 보았다. 1914년, 어니스트 햄버리 행킨과 데이비드 메러디스 시어스 왓슨은 처음으로 익룡의 공기역학을 정량적으로 분석했지만, 그들은 ''프테라노돈''을 순수한 글라이더로 해석했다.[64] 1940년대와 1950년대에는 이 그룹에 대한 연구가 거의 이루어지지 않았다.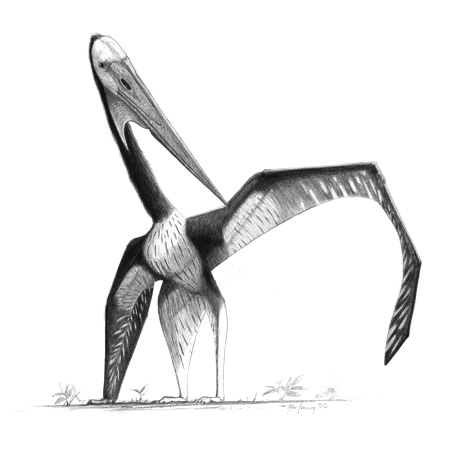
1960년대부터 공룡 르네상스가 일어났다. 데이노니쿠스의 추가적인 화석 발견으로 인해 연구 및 비판적 아이디어가 급증했는데, 이는 기존의 정설을 반박하는 놀라운 특징을 보여주었다. 1970년, 털이 있는 익룡인 ''소르데스''의 묘사 역시 로버트 배커가 익룡 르네상스라고 명명한 시발점이 되었다.[65] 특히 케빈 패디언은 새로운 관점을 널리 알렸으며, 익룡을 온혈 동물, 활동적이고, 달리는 동물로 묘사하는 일련의 연구를 발표했다.[66][67][113] 이는 1970년대 페터 벨른호퍼의 연구를 통해 독일 학파가 부활한 시기와 일치하며, 그는 현대 익룡 과학의 토대를 마련했다. 1978년, 그는 첫 번째 익룡 교과서인 ''Handbuch der Paläoherptologie, Teil 19: Pterosauria''를 출판했고, 1991년에는 두 번째 대중 과학 익룡 서적인 ''Encyclopedia of Pterosaurs''를 출판했다.
1990년대부터 연조직 잔해를 종종 보여주는 수백 개의 정교하게 보존된 2차원 화석을 가져온 랴오닝의 초기 백악기 중국 제홀 생물군은 더욱 생산적인 곳이었다. 뤼쥔창과 같은 중국 연구원들은 다시 많은 새로운 분류군을 명명했다. 세계 다른 지역에서도 발견이 증가하면서 명명된 속의 총 수가 갑자기 급증했다. 2009년까지 약 90개로 증가했을 때, 이러한 증가는 둔화될 기미를 보이지 않았다.[69] 2013년, M.P. 위튼은 발견된 익룡 종의 수가 130종으로 증가했음을 나타냈다.[70] "르네상스" 동안 알려진 분류군의 90% 이상이 명명되었다. 이 중 많은 수가 존재가 알려지지 않았던 그룹에서 나왔다. 컴퓨팅 능력의 발전은 연구자들이 계통분류학의 양적 방법을 통해 복잡한 관계를 결정할 수 있게 했다. 새롭고 오래된 화석은 현대 자외선 또는 뢴트겐 사진, 또는 CAT 스캔을 적용했을 때 훨씬 더 많은 정보를 얻었다. 생물학의 다른 분야에서 얻은 통찰력이 얻은 데이터에 적용되었다. 이 모든 것은 익룡 연구의 실질적인 발전을 가져왔으며, 대중 과학 서적의 오래된 설명은 완전히 구식이 되었다.
2017년, 1억 7천만 년 된 익룡 화석이 발견되었으며, 2022년 종으로 ''디어크 스기아나치''로 명명되었다. 이는 스코틀랜드의 스카이 섬에서 발견되었다. 스코틀랜드 국립 박물관은 이 화석이 쥐라기 시대에 발견된 동종의 가장 큰 화석이라고 주장하며, 세계에서 가장 잘 보존된 익룡 골격으로 묘사되었다.[71]
익룡의 고생물학은 1960년대부터 공룡 르네상스와 비슷한 변화를 겪었다. 1970년, 털이 있는 익룡인 ''소르데스''의 발견은 로버트 배커가 익룡 르네상스라고 명명한 시기의 시작이었다.[65] 케빈 패디언은 익룡을 온혈 동물이며 활동적이고, 달리는 동물로 묘사하는 연구를 발표하여 새로운 관점을 널리 알렸다.[66][67][113] 페터 벨른호퍼는 현대 익룡 과학의 토대를 마련하였고, 1978년에는 첫 번째 익룡 교과서인 ''Handbuch der Paläoherptologie, Teil 19: Pterosauria''를 출판했다.
]
1970년대 브라질의 초기 백악기 산타나 층에서 3차원 보존 상태가 뛰어난 익룡 골격 화석이 발견되고, 1990년대에는 중국 랴오닝의 제홀 생물군에서 연조직 잔해를 포함한 수백 개의 2차원 화석이 발견되면서 익룡 연구가 가속화되었다.[69] 이러한 발견으로 2009년까지 약 90개, 2013년에는 130종의 익룡이 명명되었다.[70] 컴퓨팅 능력의 발전으로 계통분류학의 양적 방법을 통해 익룡의 관계를 분석하고, 자외선, 뢴트겐 사진, CAT 스캔과 같은 현대 기술을 통해 화석에서 더 많은 정보를 얻게 되었다.
2017년, 스코틀랜드 스카이 섬에서 1억 7천만 년 된 익룡 화석이 발견되었고, 2022년 ''디어크 스기아나치''로 명명되었다. 이 화석은 세계에서 가장 잘 보존된 익룡 골격으로 묘사되었다.[71]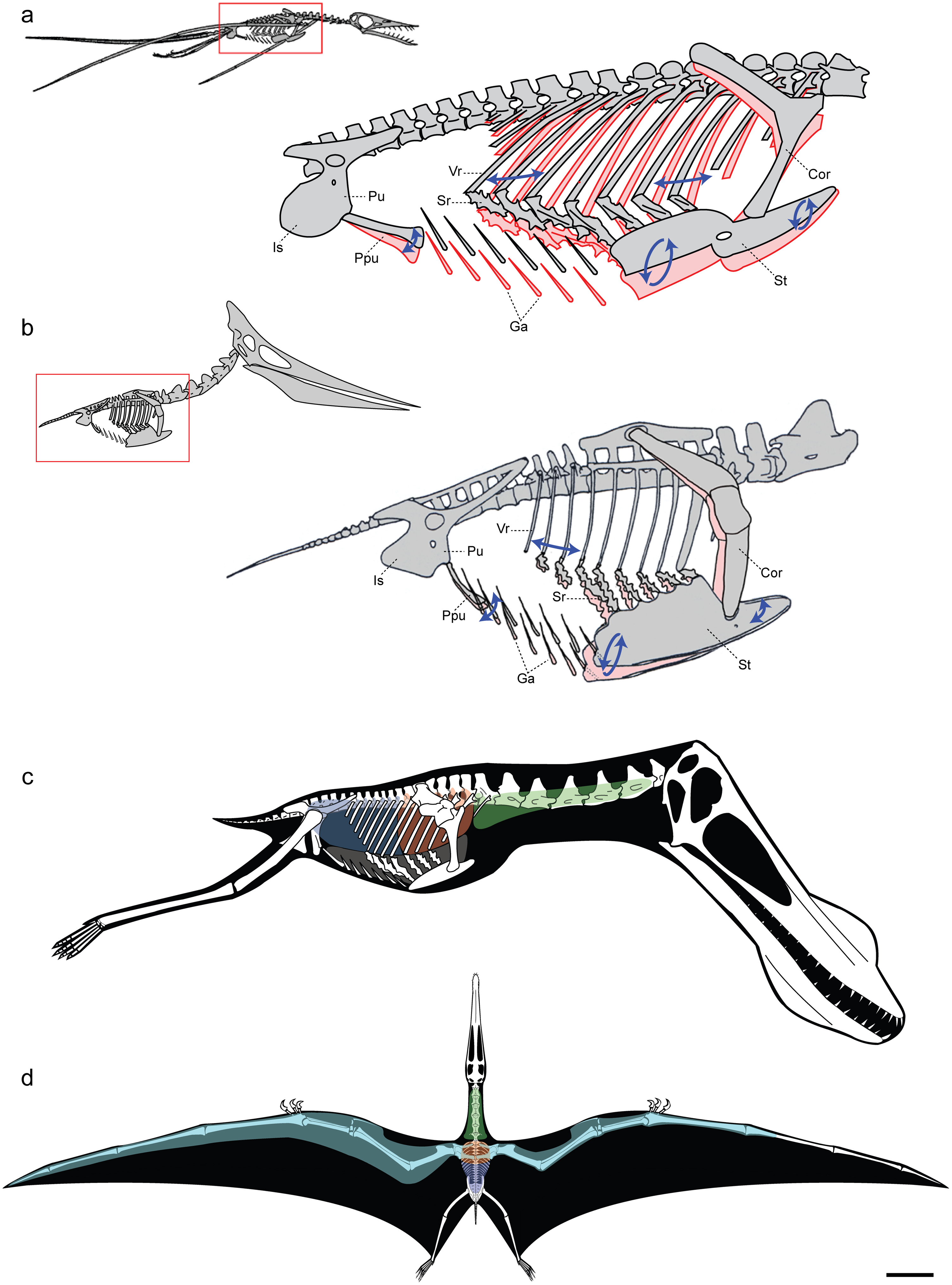
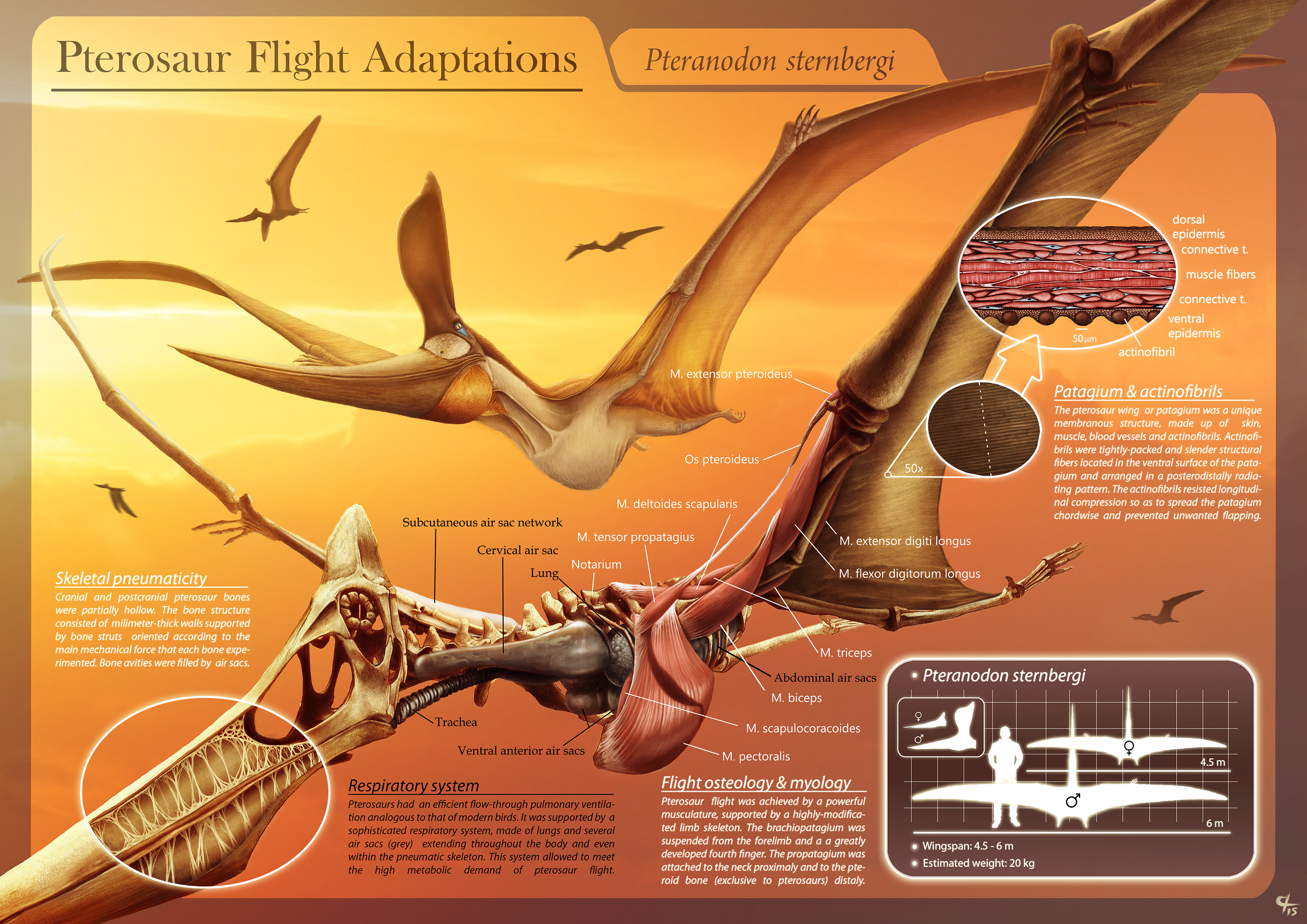
익룡의 비행 역학은 아직 완전히 이해되지 않았다.[100][101] 현대 조류를 이용한 계산에 따르면 익룡이 공중에 떠 있을 수 없다는 결론이 나왔지만,[100] 후기 백악기의 산소가 풍부하고 밀도가 높은 대기 덕분에 비행이 가능했다는 이론도 제기되었다.[102] 그러나 다렌 네이시는 대기 차이가 익룡의 거대한 크기에 필수적이지 않다고 결론 내렸다.[103] 익룡은 도약 메커니즘을 사용하여 비행했을 것으로 추정된다.[104][105] 일단 공중에 뜨면 익룡은 최대 120km/h의 속도로 수천 킬로미터를 이동할 수 있었다.[105]
익룡은 조류와 유사한 관류 폐 환기 모델을 가진 폐와 기낭 시스템, 골격 호흡 펌프를 가지고 있었다.[28] 피하 기낭 시스템은 익룡의 밀도를 더욱 감소시켰을 것이다.[28] 익룡은 악어류와 마찬가지로 간 피스톤을 가지고 있었던 것으로 보인다.[109]
익룡의 뇌 연구 결과, 거대한 소구가 발견되었는데, 이는 동물 전체 뇌 질량의 7.5%를 차지하며 다른 어떤 척추동물보다 높은 수치이다.[18] 소구는 감각 정보를 처리하는 데 중요한 역할을 하며, 익룡의 큰 날개 크기와 관련이 있을 수 있다.[18]
익룡의 엉덩이 관절과 넙다리뼈는 익룡이 직립 자세를 가졌음을 시사한다. 익룡은 사족 보행을 했는지, 이족 보행을 했는지에 대한 논쟁이 있었으나, 화석 발자국을 통해 네 발로 걷는다는 것이 밝혀졌다.[114][115] 익룡은 발 전체를 땅에 딛는 척행 보행을 했으며, 아즈다르크과는 직립 자세로 걸었다.[116][104]
익룡은 육상에서 유능한 보행자이자 주자였다.[118] 아즈다르크과는 건조한 땅에서 걷는 데 적응했으며, 크테노카스마토이드는 부드러운 진흙 토양에서 걷는 데 적응했다.[116] 익룡은 뒷다리를 사용하여 수영했을 가능성이 있으며, 닉토사우루스과의 ''알키온''은 날개를 이용한 잠수에 적응했을 수 있다.[99]
익룡은 표면에서 물고기를 먹는 어식성 동물로 여겨졌으나, 육상 육식 동물, 잡식성 동물, 곤충을 먹는 동물 등 다양한 식성을 가진 무리가 있었다. 익룡은 수각류 공룡이나 모사사우루스과 파충류에게 잡아먹혔다.[132]
익룡은 알을 낳아 번식했으며, 알 껍질은 가죽질이었다.[134] 익룡은 알을 묻었을 가능성이 높다.[140] 익룡 배아는 부화 후 곧바로 날 수 있었을 것으로 보이지만,[142] ''하미프테루스''의 경우 걷기에는 잘 발달했지만 비행에는 약한 가슴을 가지고 있었다.[143] 익룡은 부모의 보살핌 없이 조숙성을 보였을 가능성이 크다.[147] 익룡의 성장 속도는 그룹마다 달랐다. 람포린쿠스는 앨리게이터보다 약간 빠른 성장률을 보였고, 프테로닥틸로이드 익룡은 생애 첫 해 안에 성체 크기로 성장했다.[144] 익룡의 일일 활동 패턴은 주행성, 야행성, 주야혼성 등 다양했다.[151]
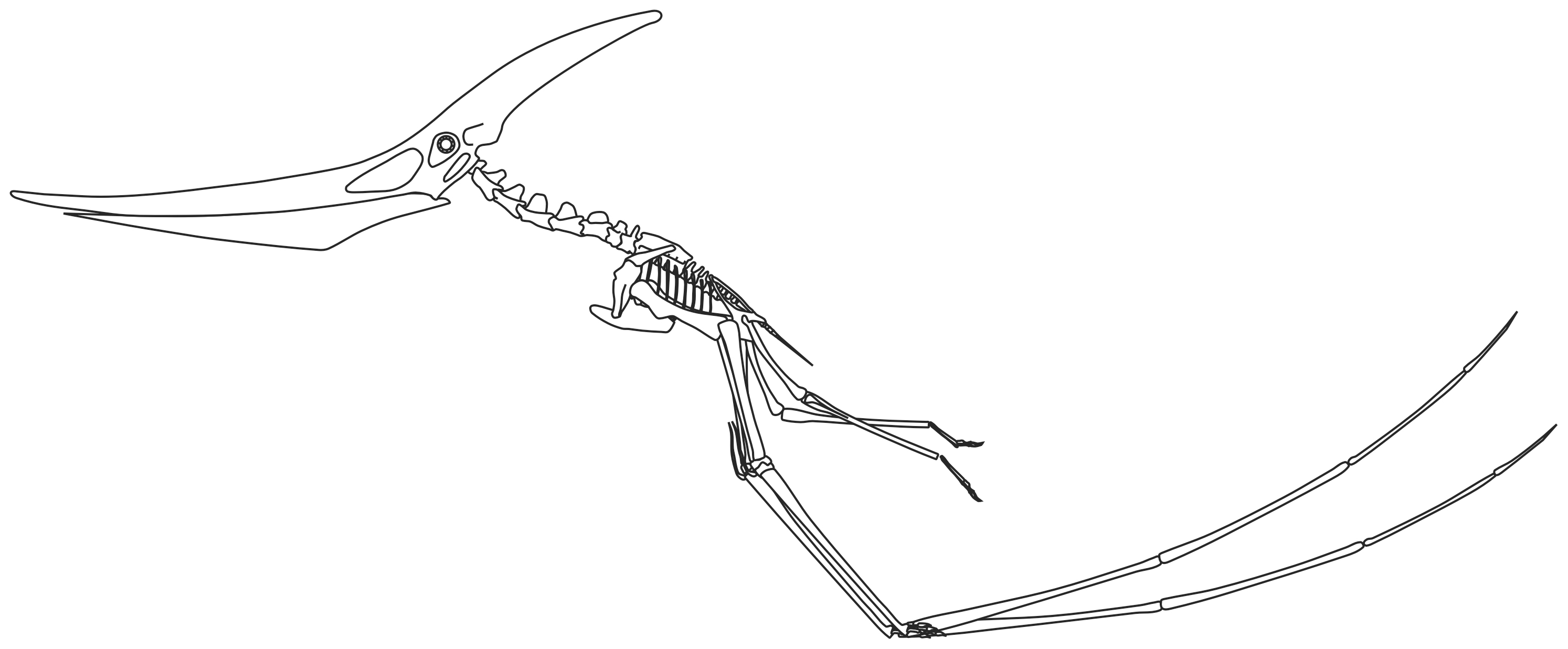

1960년대부터 공룡 르네상스가 일어나 익룡 연구에도 큰 변화가 생겼다. 1970년 털 달린 익룡 ''소르데스''가 발견되면서 로버트 배커는 이를 '익룡 르네상스'의 시작이라고 불렀다.[65] 케빈 패디언은 익룡을 온혈 동물이며 활동적이고, 달리는 동물로 묘사하는 연구를 발표했다.[66][67][113] 페터 벨른호퍼는 1978년 최초의 익룡 교과서인 ''Handbuch der Paläoherptologie, Teil 19: Pterosauria''를 출판하고,[70] 1991년에는 ''Encyclopedia of Pterosaurs''를 출판하여 현대 익룡 과학의 기초를 다졌다.
1970년대 브라질 산타나 층에서 3차원 익룡 골격 화석이 발견되고, 1990년대 중국 랴오닝의 제홀 생물군에서 연조직 잔해를 포함한 수백 개의 2차원 익룡 화석이 발견되면서 익룡 연구가 더욱 가속화되었다.[69] 알렉산더 켈너를 포함한 브라질 연구원들과 뤼쥔창과 같은 중국 연구원들은 많은 새로운 익룡 종을 발견하고 명명했다. 2013년에는 발견된 익룡 종의 수가 130종으로 증가했다.[70]
컴퓨터 기술의 발전으로 계통분류학의 양적 방법을 통해 익룡 간의 관계를 분석하고, 자외선, 뢴트겐, CAT 스캔과 같은 현대 기술을 통해 화석에서 더 많은 정보를 얻을 수 있게 되었다.
2017년 스코틀랜드 스카이 섬에서 1억 7천만 년 된 익룡 화석 ''디어크 스기아나치''가 발견되었는데, 스코틀랜드 국립 박물관은 이 화석을 쥐라기 시대에 발견된 가장 큰 익룡 화석이자 세계에서 가장 잘 보존된 익룡 골격이라고 평가했다.[71]
과거에는 초기 조류와의 경쟁이 익룡의 멸종을 야기했다고 생각되었지만,[84] 최근 연구에 따르면 익룡과 조류 사이의 생태적 중복은 거의 없었으며, 익룡의 쇠퇴는 조류의 다양성과 관련이 없는 것으로 보인다.[86] 백악기-고생대 멸종 사건으로 인해 비조류 공룡과 함께 익룡도 멸종한 것으로 보인다.
2010년대 이후 ''픽시''(Piksi)와 ''오르니토케이루스''("Ornithocheirus")를 포함한 새로운 익룡들이 캄파니아절/마스트리히트절에 존재했던 것으로 밝혀졌고,[87][88] 프테라노돈과, 닉토사우루스과, 여러 테페자리드와 ''나바호닥틸루스''(Navajodactylus) 등이 발견되면서 백악기 후기 익룡의 다양성이 이전에 생각했던 것보다 훨씬 컸음을 알 수 있다. 소형 아즈다르크과 익룡도 캄파니아절에 존재했다.[89]
최근 연구에 따르면, 백악기 후기 소형 익룡의 생태적 지위는 대형 익룡 종의 유년기가 차지했으며, 익룡은 조류와의 경쟁보다는 중생대 이전부터 진행된 전문화 경향을 따랐다는 것이 밝혀졌다.[92]


익룡은 사촌 격인 공룡만큼 영화, 문학이나 다른 예술 형식에서 두드러지지는 않았지만 공룡만큼이나 오랜 시간 동안 대중 문화의 한 자리를 차지해 왔다. 게다가 대중매체에서 묘사된 공룡들은 고생물학의 발전에 발맞추어 크게 바뀌어왔지만 20세기 중반부터 익룡의 모습은 계속해서 시대에 뒤떨어진 상태로 남아 있었다.[233]
일반적인 용어인 "프테로닥틸"이 익룡 전체를 가리키는 단어로 사용되어 왔지만, 익룡의 모습은 프테라노돈이나 람포린쿠스, 혹은 그 둘이 합쳐진 가상의 모습이 주로 보여졌다.[233] 아이들 장난감이나 만화에서 묘사된 "프테로닥틸"은 "프테라노돈"의 것처럼 생긴 볏과 람포린쿠스의 것처럼 생긴 긴 꼬리와 이빨로 이루어진, 실제로는 전혀 존재한 적이 없던 조합으로 이루어졌다. 최소한 한 종류의 익룡은 실제로 "프테라노돈"처럼 생긴 볏과 이빨을 가지고 있었다. 루도닥틸루스가 그 예인데, 이 이름은 "장난감 손가락"이라는 의미로, 과학적으로 정확하지 않던 아이들 장난감을 닮았기 때문에 붙여진 이름이다.[234] 또한, 어떤 익룡 그림은 조류라는 설명이 붙어 있기도 한데, 사실 익룡은 하늘을 날던 파충류이며 조류는 수각류 공룡의 후손이다.

익룡은 아서 코난 도일의 1912년 소설 잃어버린 세계와 그 후에 발표된 영화 잃어버린 세계에서도 등장한다. 이후, 1933년의 영화 킹콩, 1966년의 영화 공룡 100만년 등 여러 영화 및 TV 프로그램에서도 볼 수 있다. 영화 공룡 100만년에서 애니메이터였던 레이 해리하우젠은 비막을 지탱하기 위해 과학적으로는 정확하지 않지만 박쥐의 날개와 같은 형태의 손가락을 스톱모션에 사용된 익룡 모형에 추가해야 했다. 이 오류는 영화가 만들어지기 전의 그림 등에서도 흔한 것이었다. 1956년 영화 라돈에서 처음 등장한 가상의 거대 괴수(또는 ''괴수'')인 라돈은 거대한 방사선에 노출된 ''프테라노돈'' 종으로 묘사된다.[154] 라돈은 1960년대, 1970년대, 1990년대, 2000년대에 개봉된 여러 일본 고질라 영화에 출연했으며, 2019년 미국에서 제작된 영화 고질라: 킹 오브 몬스터에도 출연했다.[154][155][156]
익룡은 2001년의 영화 쥬라기 공원 3 이전까지는 영화에 그리 자주 등장하지 않았다. 하지만 고생물학자 데이브 혼은 40년이 지났음에도 영화에 묘사된 익룡은 현대적인 연구결과를 반영하지 못하고 있다고 지적했다. 1960년대부터 2000년대까지 꾸준히 이어진 오류들은 이빨이 없는 종류 (쥐라기 공원 3의 익룡은 프테라노돈인데 이 이름은 "이빨이 없는 날개" 란 의미이다.) 가 이빨을 가지고 있다거나, 2001년이면 익룡이 둥지에서 새끼를 돌보지 않는다는 것이 알려졌는데 그런 행동을 보인다거나, 익룡의 비행에 필요했고 실제로 익룡이 가지고 있었던 근육섬유로 된 팽팽한 비막 대신 펄럭이는 가죽 같은 날개를 가지고 있는 것 등이 있다.[233] ''랜드 비포 타임''(1988)의 페트리는 애니메이션 영화의 주목할 만한 예이다.[157]
대부분의 미디어 출연에서 익룡은 어식성으로 묘사되는데, 이는 그들의 모든 식단 변이를 반영하지 않는다. 또한 그들은 종종 맹금류와 유사한 공중 포식자로 묘사되어 발톱으로 인간 희생자를 움켜쥐고 있다. 그러나 작은 안우로그나투스과 ''베스페로프테릴루스''와 작은 우콩고프테루스과 ''쿤펭고프테루스''만이 각각 잡는 발과 손을 가지고 있는 것으로 알려져 있다.[158] 다른 알려진 모든 익룡은 대항할 수 없는 발가락이 없는 평평한 지상 보행형 발을 가지고 있으며, 발은 일반적으로 적어도 프테라노돈과에서 비례적으로 작다.[15]
[1]
논문
The Earliest Pterodactyloid and the Origin of the Group
[2]
논문
Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques.
[3]
서적
Pterosaurs: Natural History, Evolution, Anatomy
Princeton University Press
2013
[4]
간행물
"Darwinopterus'' and its implications for pterosaur phylogeny"
[5]
서적
English Pronouncing Dictionary
Cambridge University Press
[6]
웹사이트
Pterosaur
[7]
서적
The dinosaur book: the ruling reptiles and their relatives
https://archive.org/[...]
McGraw-Hill
1951
[8]
논문
Pterosaur distribution in time and space: an atlas
http://epub.ub.uni-m[...]
[9]
논문
The Extent of the Pterosaur Flight Membrane
[10]
웹사이트
Pterosaur.net :: Terrestrial Locomotion
https://pterosaur.ne[...]
2020-02-01
[11]
웹사이트
"It's Official: Those Flying Reptiles Called Pterosaurs Were Covered in Fluffy Feathers"
https://www.livescie[...]
2018-12-17
[12]
논문
Pterosaur from the Latest Cretaceous of West Texas: Discovery of the Largest Flying Creature
1975-03
[13]
논문
A new giant pterosaur with a robust skull from the latest cretaceous of Romania
http://doc.rero.ch/r[...]
2002-04
[14]
서적
The Dinosauria
https://archive.org/[...]
University of California Press
[15]
웹사이트
Pterosaurs: Myths and Misconceptions
http://www.pterosaur[...]
Pterosaur.net
2011-06-18
[16]
서적
Nature's Flyers: Birds, Insects, and the Biomechanics of Flight
https://books.google[...]
JHU Press
[17]
뉴스
Hundreds of Fossilized Pterosaur Eggs Uncovered in China
https://www.nytimes.[...]
2017-11-30
[18]
논문
Neuroanatomy of flying reptiles and implications for flight, posture and behaviour
http://doc.rero.ch/r[...]
[19]
웹사이트
Pterosaur.net :: Origins and Relationships
https://pterosaur.ne[...]
2020-02-01
[20]
논문
A new rhamphorhynchid pterosaur from the Upper Jurassic of Xinjiang, China, and the phylogenetic relationships of basal pterosaurs
http://doc.rero.ch/r[...]
2010-01-29
[21]
논문
Discovery of a rare arboreal forest-dwelling flying reptile (Pterosauria, Pterodactyloidea) from China
[22]
논문
Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity
[23]
논문
Soft tissue preservation in a specimen of ''Pterodactylus kochi'' (Wagner) from the Upper Jurassic of Germany
[24]
서적
Feathered Dinosaurs and the Origin of Flight
The Dinosaur Museum: Blanding, Utah
[25]
논문
Taxonomy and systematics of the Late Cretaceous pterosaur ''Pteranodon'' (Pterosauria, Pterodactyloidea)
https://www.biodiver[...]
[26]
논문
Pterosaur flight: the role of actinofibrils in wing function
[27]
논문
Pterosaurs – a successful invasion of prehistoric skies
[28]
논문
Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism
[29]
논문
''Sordes pilosus'' and the nature of the pterosaur flight apparatus
[30]
논문
A nearly completely articulated rhamphorhynchoid pterosaur with exceptionally well-preserved wing membranes and "hairs" from Inner Mongolia, northeast China
[31]
논문
New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion
[32]
논문
Limb disparity and wing shape in pterosaurs
2006-07
[33]
논문
High lift function of the pteroid bone and forewing of pterosaurs
[34]
논문
Articulation and Function of the Pteroid Bone of Pterosaurs
http://doc.rero.ch/r[...]
[35]
논문
New material of the non-pterodactyloid pterosaur Changchengopterus pani Lü, 2009 from the Late Jurassic Tiaojishan Formation of western Liaoning
[36]
논문
New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China
[37]
논문
High lift function of the pteroid bone and forewing of pterosaurs
[38]
논문
Pterosaurs evolved a muscular wing–body junction providing multifaceted flight performance benefits: Advanced aerodynamic smoothing, sophisticated wing root control, and wing force generation
[39]
논문
The soft tissue of ''Jeholopterus'' (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane
[40]
논문
Beiträge zur Erkentniss verschiedner Reptilien der Vorwelt
[41]
논문
Pterosaur integumentary structures with complex feather-like branching
https://research-inf[...]
2019-01
[42]
뉴스
Fur flies over new pterosaur fossils
https://www.bbc.com/[...]
2018-12-17
[43]
논문
No protofeathers on pterosaurs
2020-12
[44]
논문
The Soft Tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the Structure of the Pterosaur Wing Membrane
2009
[45]
논문
Sordes pilosus and the nature of the pterosaur flight apparatus
1994-09
[46]
논문
Reply to: No protofeathers on pterosaurs
2020-12
[47]
논문
Pterosaur plumage
2019-01
[48]
논문
Pterosaur melanosomes support signalling functions for early feathers
2022
[49]
간행물
"Sur quelques Zoolithes du Cabinet d'Histoire naturelle de S. A. S. E. Palatine & de Bavière, à Mannheim."
1784
[50]
서적
Natürliches System der Amphibien
Munich
1830
[51]
논문
"[Reptile volant]. In: Extrait d'un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossemens dans l'intérieur de la terre"
[52]
간행물
"Mémoire sur le squelette fossile d'un Reptil volant des environs d'Aichstedt, que quelques naturalistes ont pris pour un oiseau, et donc nous formons un genre de Sauriens, sous le nom de Ptero-Dactyle"
Paris
1809
[53]
서적
Analyse de la Nature ou tableau de l'univers et des corps organisés
Palermo
1815
[54]
간행물
"Über einen Ornithocephalus oder über das unbekannten Thier der Vorwelt, dessen Fossiles Gerippe Collini im 5. Bande der Actorum Academiae Theodoro-Palatinae nebst einer Abbildung in natürlicher Grösse im Jahre 1784 beschrieb, und welches Gerippe sich gegenwärtig in der Naturalien-Sammlung der königlichen Akademie der Wissenschaften zu München befindet"
München: mathematisch-physikalische Classe
1812
[55]
논문
Note on the Pterodactyle Tribe considered as Marsupial Bats
[56]
논문
Versuch einer Eintheilung der Säugethiere in 6 Stämme und der Amphibien in 6 Ordnungen
https://www.biodiver[...]
[57]
서적
Ornithosauria – an elementary study of the bones of Pterodactyles
Cambridge University Press
1870
[58]
서적
Dragons of the Air: An account of extinct flying reptiles
Londen: Methuen
1901
[59]
논문
A popular account of chamaeleons
[60]
논문
The wings of Pterodactyles
[61]
논문
Über Flugsaurier aus dem lithografischen Schiefer Bayerns
[62]
간행물
"Ein Ramphorhynchus mit Spuren von Haarbedeckung"
1927
[63]
논문
Das Gehirn der Pterosaurier
http://bigcat.fhsu.e[...]
2019-10-27
[64]
간행물
"On the Flight of Pterodactyls"
1914-10
[65]
서적
The Dinosaur Heresies
Londen: Penguin Books
1986
[66]
논문
The wings of pterosaurs: A new look
[67]
간행물
'Studies of the structure, evolution, and flight of pterosaurs (reptilia: Pterosauria)'
1980
[68]
서적
Handbuch der Paläoherpetologie XIX. Pterosauria
Urban & Fischer, München
1978
[69]
논문
The shape of pterosaur evolution: evidence from the fossil record
[70]
서적
2013
[71]
웹사이트
'Superbly preserved' pterosaur fossil unearthed in Scotland
https://phys.org/new[...]
2022-02-22
[72]
논문
"''Scleromochlus taylori'' and the origin of dinosaurs and pterosaurs"
[73]
논문
The phylogenetic position of the Pterosauria within the Archosauromorpha
[74]
논문
A Late Triassic Dinosauromorph Assemblage from New Mexico and the Rise of Dinosaurs
http://doc.rero.ch/r[...]
[75]
논문
An evaluation of the phylogenetic relationships of the pterosaurs to the archosauromorph reptiles
[76]
논문
The early evolution of archosaurs: relationships and the origin of major clades
[77]
논문
The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms
2016-04-28
[78]
논문
Reassessment of the Triassic archosauriform ''Scleromochlus taylori'': neither runner nor biped, but hopper
[79]
논문
Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria
http://doc.rero.ch/r[...]
2020-12-17
[80]
웹사이트
Paleontologists find pterosaur precursors that fill a gap in early evolutionary history
https://phys.org/new[...]
2020-12-14
[81]
웹사이트
Pterosaur Origins Flap into Focus
https://www.scientif[...]
2020-12-14
[82]
논문
The origin of Pterosaurs
2021-10
[83]
간행물
"Über die Ursprung der Flugsaurier"
Weltenberger Akademie, Erwin Rutte-Festschrift
[84]
기타
BBC Documentary: Walking with dinosaurs (episode 4 ) – Giant Of The Skies
[85]
논문
Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution
2006-06
[86]
논문
Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement
[87]
논문
Lone Star Pterosaurs
[88]
논문
Systematic reinterpretation of ''Piksi barbarulna'' Varricchio, 2002 from the Two Medicine Formation (Upper Cretaceous) of Western USA (Montana) as a pterosaur rather than a bird
http://www.mnhn.fr/m[...]
2012-12-29
[89]
논문
Does morphology reflect osteohistology-based ontogeny? A case study of Late Cretaceous pterosaur jaw symphyses from Hungary reveals hidden taxonomic diversity
http://real.mtak.hu/[...]
[90]
논문
A small azhdarchoid pterosaur from the latest Cretaceous, the age of flying giants
[91]
논문
Pterosaur remains (Archosauria, Ornithodira) from the early Late Cretaceous of "La Buitrera", Río Negro, Argentina
http://www.conicet.g[...]
[92]
논문
Small, immature pterosaurs from the Cretaceous of Africa: implications for taphonomic bias and palaeocommunity structure in flying reptiles
https://figshare.com[...]
2021-10-16
[93]
논문
Pterosaur phylogeny and comments on the evolutionary history of the group
[94]
서적
"Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin"
Geological Society of London
[95]
서적
Phylonyms: A Companion to the PhyloCode
https://books.google[...]
CRC Press Boca Raton, FL
[96]
서적
"Pterosauromorpha"
[97]
서적
The Pterosaurs: From Deep Time
Pi Press
[98]
논문
A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution
[99]
논문
Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary
[100]
뉴스
Pterodactyls were too heavy to fly, scientist claims
https://www.telegrap[...]
2008-10-01
[101]
뉴스
Were pterosaurs too big to fly?
https://www.newscien[...]
2008-10-02
[102]
서적
Posture, locomotion, and paleoecology of pterosaurs
https://books.google[...]
Geological Society of America
[103]
웹사이트
Pterosaurs breathed in bird-like fashion and had inflatable air sacs in their wings
http://scienceblogs.[...]
2009-02-18
[104]
뉴스
Why pterosaurs weren't so scary after all
https://www.theguard[...]
2013-08-11
[105]
뉴스
Did giant pterosaurs vault aloft like vampire bats?
https://www.newscien[...]
2010-11-16
[106]
논문
The Great Pterodactyl Project
http://calteches.lib[...]
[107]
뉴스
With Wings Flapping, Model Pterodactyl Takes to Air
https://www.nytimes.[...]
1986-01-28
[108]
웹사이트
The wingtips of the pterosaurs: Anatomy, aeronautical function and 3 ecological implications
https://qmro.qmul.ac[...]
2022-06-25
[109]
논문
Breathing in a box: Constraints on lung ventilation in giant pterosaurs
[110]
논문
Relative Brain Size and Behavior in Archosaurian Reptiles
[111]
뉴스
Coldblooded Does Not Mean Stupid
https://www.nytimes.[...]
2013-11-18
[112]
논문
A Jurassic pterosaur from Patagonia and the origin of the pterodactyloid neurocranium
[113]
논문
A Functional Analysis of Flying and Walking in Pterosaurs
[114]
논문
Pterosaur Stance and Gait and the Interpretation of Trackways
http://doc.rero.ch/r[...]
[115]
논문
New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea
http://edoc.hu-berli[...]
[116]
논문
A reappraisal of azhdarchid pterosaur functional morphology and paleoecology
[117]
논문
Were early pterosaurs inept terrestrial locomotors?
[118]
논문
Pterosaur tracks and the terrestrial ability of pterosaurs
http://doc.rero.ch/r[...]
[119]
서적
[120]
논문
A second specimen of the pterosaur ''Anurognathus ammoni''
[121]
문서
Dietary tendencies of the Early Jurassic pterosaurs Campylognathoides Strand, 1928, and Dorygnathus Wagner, 1860, with additional evidence for teuthophagy in Pterosauria
[122]
논문
A new rhamphorhynchid pterosaur from the Upper Jurassic of Xinjiang, China, and the phylogenetic relationships of basal pterosaurs
http://doc.rero.ch/r[...]
[123]
논문
A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications
[124]
논문
Evidence for tactile foraging in pterosaurs: a sensitive tip to the beak of Lonchodraco giganteus (Pterosauria, Lonchodectidae) from the Upper Cretaceous of southern England
https://www.scienced[...]
2021-01-01
[125]
논문
The toothless pterosaur ''Jidapterus edentus'' (Pterodactyloidea: Azhdarchoidea) from the Early Cretaceous Jehol Biota and its paleoecological implications
[126]
문서
Preliminary mandibular myological reconstruction of ''Thalassodromeus sethi'' (Pterodactyloidea: Tapejaridae)
[127]
논문
Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"?
2015
[128]
논문
Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators
2017
[129]
간행물
Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania
http://svpca.org/abs[...]
2013
[130]
논문
An unusual modification of the jaws in cf. Alanqa, a mid-Cretaceous azhdarchid pterosaur from the Kem Kem beds of Morocco
https://researchport[...]
2015-03
[131]
논문
Reconstruction of the adductor chamber and predicted bite force in pterodactyloids (Pterosauria)
2021-09-24
[132]
논문
Pterosaurs as part of a spinosaur diet
2004-07
[133]
웹사이트
Prehistoric sharks feasted on flying reptiles, fossil reveals
https://www.national[...]
2018-10-03
[134]
논문
Palaeontology: pterosaur egg with a leathery shell
http://doc.rero.ch/r[...]
2004-12
[135]
논문
Primer reporte de un embrión de pterosaurio (Cretácico inferior, San Luis, Argentina)
https://www.ameghini[...]
2014
[136]
논문
Argentinian unhatched pterosaur fossil
https://doi.org/10.1[...]
2004-12
[137]
논문
First occurrence of stomach stones in pterosaurs
https://doi.org/10.1[...]
2013-05
[138]
논문
An egg-adult association, gender, and reproduction in pterosaurs
[139]
논문
Sexually Dimorphic Tridimensionally Preserved Pterosaurs and Their Eggs from China
[140]
논문
A note on pterosaur nesting behavior
[141]
논문
Eggshell and Histology Provide Insight on the Life History of a Pterosaur with Two Functional Ovaries
[142]
논문
Palaeontology: pterosaur embryo from the Early Cretaceous
2004-06
[143]
웹사이트
Pterosaur hatchlings needed their parents, trove of eggs reveals (Update)
https://phys.org/new[...]
2020-03-21
[144]
논문
A statistical study of ''Rhamphorhynchus'' from the Solnhofen Limestone of Germany: Year-classes of a single large species
[145]
웹사이트
First 3D pterosaur eggs found with their parents
https://phys.org/new[...]
2020-03-21
[146]
논문
Prenatal development in pterosaurs and its implications for their postnatal locomotory ability
[147]
논문
Powered flight in hatchling pterosaurs: Evidence from wing form and bone strength
[148]
논문
Allometric wing growth links parental care to pterosaur giantism
2023-07-26
[149]
웹사이트
'July: Pterosaurs parents {{!}} News and features {{!}} University of Bristol'
https://www.bristol.[...]
2023-08-22
[150]
논문
Life history of ''Rhamphorhynchus'' inferred from bone histology and the diversity of pterosaurian growth strategies
[151]
논문
Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology
[152]
웹사이트
Pterosaurs In Popular Culture.
http://www.pterosaur[...]
Pterosaur.net
2010-08-27
[153]
서적
"A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur" in: Buffetaut, E., and Mazin, J.-M. (eds.). Evolution and Palaeobiology of Pterosaurs
Geological Society Special Publication
[154]
간행물
The One Born of Fire: a pterosaurological analysis of Rodan
https://jgeekstudies[...]
Journal of Geek Studies
[155]
웹사이트
A Monster-Sized Breakdown of Every Insane 'Godzilla' Movie
https://www.thrillis[...]
2019-07-11
[156]
웹사이트
'Godzilla: King of the Monsters' Trailer Turns Mothra, Rodan, and More Into Epic Spectacle
https://www.indiewir[...]
2019-07-11
[157]
서적
From Abba to Zoom A Pop Culture Encyclopedia of the Late 20th Century
Andrews MacMeel Publishing
2005
[158]
논문
A new darwinopteran pterosaur reveals arborealism and an opposed thumb
[159]
논문
The Earliest Pterodactyloid and the Origin of the Group
[160]
논문
Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques.
[161]
뉴스
翼竜につながる特徴 三畳紀の二足歩行爬虫類―国際チーム
JIJI.COM
2020
[162]
서적
Pterosauromorpha
Academic Press
[163]
논문
A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution
[164]
뉴스
Pterosaur melanosomes support signalling functions for early feathers
https://doi.org/10.1[...]
2022
[165]
웹사이트
翼竜の「ペリット」の化石を初めて発見!翼竜の食性知る手掛かりに--人民網日本語版--人民日報
http://j.people.com.[...]
2024-09-30
[166]
웹사이트
Like Owls, Some Prehistoric Flying Reptiles May Have Regurgitated Pellets
https://www.smithson[...]
2024-09-30
[167]
논문
Discovery of a rare arboreal forest-dwelling flying reptile (''Pterosauria, Pterodactyloidea'') from China
[168]
논문
Pterosaur from the Latest Cretaceous of West Texas: Discovery of the Largest Flying Creature
1975-03
[169]
논문
A new giant pterosaur with a robust skull from the latest cretaceous of Romania
2002-04
[170]
서적
The Dinosauria
https://archive.org/[...]
University of California Press
[171]
논문
Neuroanatomy of flying reptiles and implications for flight, posture and behaviour
[172]
논문
Pterosaur flight: the role of actinofibrils in wing function
[173]
논문
Pterosaurs — a successful invasion of prehistoric skies
[174]
논문
Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism
[175]
논문
High lift function of the pteroid bone and forewing of pterosaurs
[176]
저널
Articulation and Function of the Pteroid Bone of Pterosaurs
[177]
저널
On the nature of the pteroid in pterosaurs
[178]
저널
High lift function of the pteroid bone and forewing of pterosaurs
[179]
저널
"Sordes pilosus" and the nature of the pterosaur flight apparatus
[180]
저널
A nearly completely articulated rhamphorhynchoid pterosaur with exceptionally well-preserved wing membranes and "hairs" from Inner Mongolia, northeast China
[181]
저널
New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion
[182]
저널
Limb disparity and wing shape in pterosaurs
2006-07
[183]
저널
Soft tissue preservation in a specimen of ''Pterodactylus kochi'' (Wagner) from the Upper Jurassic of Germany
[184]
간행물
A new rhamphorhynchoid with a headcrest and complex integumentary structures
The Dinosaur Museum
[185]
서적
Pterosaurs: Natural History, Evolution, Anatomy
Princeton University Press
[186]
문서
Unwin 2005
[187]
문서
The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane.
2009-08-05
[188]
문서
Sur quelques Zoolithes du Cabinet d’Histoire naturelle de S. A. S. E. Palatine & de Bavière, à Mannheim.
1784
[189]
문서
Natürliches System der Amphibien
1830
[190]
저널
'[Reptile volant]. In: Extrait d'un ouvrage sur les espèces de quadrupèdes dont on a trouvé les ossemens dans l'intérieur de la terre'
[191]
서적
Pterosaurs: rulers of the skies in the dinosaur age
http://books.google.[...]
Houghton Mifflin Harcourt
[192]
웹인용
Pterosaurs: Myths and Misconceptions
http://www.pterosaur[...]
Pterosaur.net
2011-06-18
[193]
서적
Nature's Flyers: Birds, Insects, and the Biomechanics of Flight
https://books.google[...]
JHU Press
[194]
뉴스
Pterodactyls were too heavy to fly, scientist claims
http://www.telegraph[...]
2008-10-01
[195]
뉴스
Were pterosaurs too big to fly?
http://www.newscient[...]
2008-10-02
[196]
뉴스
Pterodactyls were too heavy to fly, scientist claims
http://www.telegraph[...]
2008-10-01
[197]
서적
Posture, locomotion, and paleoecology of pterosaurs
http://books.google.[...]
Geological Society of America
[198]
웹인용
Pterosaurs breathed in bird-like fashion and had inflatable air sacs in their wings
http://scienceblogs.[...]
2009-02-18
[199]
뉴스
Why pterosaurs weren't so scary after all
http://www.theguardi[...]
2013-08-11
[200]
뉴스
Did giant pterosaurs vault aloft like vampire bats?
http://www.newscient[...]
2010-11-16
[201]
저널
A Functional Analysis of Flying and Walking in Pterosaurs
[202]
저널
Pterosaur Stance and Gait and the Interpretation of Trackways
[203]
저널
New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea
[204]
저널
A reappraisal of azhdarchid pterosaur functional morphology and paleoecology
http://www.plosone.o[...]
[205]
저널
Pterosaur tracks and the terrestrial ability of pterosaurs
[206]
저널
Pterosaurs as part of a spinosaur diet
2004-07
[207]
저널
Palaeontology: pterosaur egg with a leathery shell
2004-12
[208]
저널
An egg-adult association, gender, and reproduction in pterosaurs
[209]
저널
A note on pterosaur nesting behavior
[210]
저널
Palaeontology: pterosaur embryo from the Early Cretaceous
2004-06
[211]
논문
A statistical study of ''Rhamphorhynchus'' from the Solnhofen Limestone of Germany: Year-classes of a single large species
[212]
논문
Life history of ''Rhamphorhynchus'' inferred from bone histology and the diversity of pterosaurian growth strategies
[213]
논문
Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology
[214]
논문
A Late Triassic Dinosauromorph Assemblage from New Mexico and the Rise of Dinosaurs
[215]
논문
An evaluation of the phylogenetic relationships of the pterosaurs to the archosauromorph reptiles
[216]
논문
The early evolution of archosaurs: relationships and the origin of major clades
http://digitallibrar[...]
2014-04-27
[217]
간행물
Pterosaur phylogeny and comments on the evolutionary history of the group.
Geological Society, London, Special Publications
[218]
간행물
Pterosauromorpha.
Currie, P.J. and Padian, K. The Encyclopedia of Dinosaurs. Academic Press.
[219]
서적
The Pterosaurs: From Deep Time
https://archive.org/[...]
Pi Press
[220]
논문
A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution
[221]
기타
Walking with dinosaurs (episode 4 ) – Giant Of The Skies
BBC Documentary
[222]
논문
Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution
http://mbe.oxfordjou[...]
2006-06
[223]
논문
Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement
[224]
doi
[225]
논문
Systematic reinterpretation of ''Piksi barbarulna'' Varricchio, 2002 from the Two Medicine Formation (Upper Cretaceous) of Western USA (Montana) as a pterosaur rather than a bird
http://www.mnhn.fr/m[...]
2014-04-28
[226]
서적
Pterosaurs: Natural History, Evolution, Anatomy
Princeton University Press
[227]
문서
Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania
[228]
서적
The Illustrated Encyclopedia of Pterosaurs
[229]
논문
Year-classes of pterosaurs from the Solnhofen Limestone of Germany: Taxonomic and Systematic Implications
[230]
서적
The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals
https://archive.org/[...]
Marshall Editions
[231]
논문
Pterosaurs
[232]
웹인용
Hatzegopteryx
http://www.bbc.co.uk[...]
BBC
2012-03-02
[233]
웹사이트
Pterosaurs In Popular Culture.
http://www.pterosaur[...]
2010-08-27
[234]
간행물
A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur.
Buffetaut, E., and Mazin, J.-M. (eds.). Evolution and Palaeobiology of Pterosaurs.
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com