토레토크네무스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
토레토크네무스는 트라이아스기 후기에 북아메리카 태평양 연안에 서식했던 어룡의 한 속이다. 안와가 크고 관골이 좁으며, 턱뼈와 이빨의 특징을 보인다. 꼬리 척추, 사지, 골반 등의 해부학적 특징은 종에 따라 차이를 보이며, 두 종은 유사한 사지를 가지고 있다. 1903년 메리엄에 의해 명명되었으며, 화석은 캘리포니아주 등지에서 발견되었다. 수영 방식은 꼬리를 흔드는 방식이었으며, 새끼를 낳는 난태생이었다. 분류학적으로는 롱기피나테에 속하며, 토레토크네무스과에 속한다.
더 읽어볼만한 페이지
- 1903년 기재된 화석 분류군 - 브라키오사우루스
브라키오사우루스는 최대 25m에 달하는 거대한 용각류 공룡으로, 긴 목과 앞다리가 긴 체형을 가졌으며 북아메리카에서 발견되었고, 대중문화에서 상징적인 공룡으로 묘사된다. - 1903년 기재된 화석 분류군 - 오르니톨레스테스
오르니톨레스테스는 쥐라기 후기 북아메리카에 살았던 "새 도둑"이라는 뜻의 소형 수각류 공룡으로, 날렵한 몸집과 날카로운 이빨로 작은 동물을 사냥했을 것으로 추정되며 헨리 페어필드 오스본에 의해 명명된 이후 깃털 존재 여부와 식성 등에 대한 다양한 가설이 제시되었다. - 미국의 화석 - 트리케라톱스
트리케라톱스는 북아메리카에서 발견된 각룡류 공룡의 한 속으로, 거대한 두개골과 뿔이 특징이며 백악기 말기에 번성했고, 현재는 두 종으로 분류된다. - 미국의 화석 - 티라노사우루스
티라노사우루스는 백악기 후기 북아메리카에 서식했던 몸길이 15m에 달하는 거대한 육식 공룡으로, 강력한 턱힘과 짧은 앞다리, 긴 꼬리를 가진 최상위 포식자였으며, '수'와 '스카티' 등의 표본과 대중문화 속 인기 덕분에 그 생태와 행동에 대한 연구가 지속되고 있다. - 어룡 - 운도로사우루스
운도로사우루스는 후기 쥐라기 시대에 살았던 해양 파충류로, 불완전하게 융합된 좌골치골과 강한 치열을 가지고 있으며, 삼각형 이빨로 해양 생물을 잡아먹는 육식성 포식자로 추정되지만, 계통 발생적 위치는 아직 논쟁 중이다. - 어룡 - 겐가사우루스
쥐라기에 이탈리아 겐가 지역에서 서식한 겐가사우루스는 약 2.4m의 몸길이를 가진 오프탈모사우루스과의 어룡으로, 비교적 완전한 화석 골격 특징으로 오프탈모사우루스와 구별된다.
| 토레토크네무스 | |
|---|---|
| 토레토크네무스 정보 | |
 | |
| 학명 | Toretocnemus |
| 명명자 | Merriam, 1903 |
| 화석 범위 | 트라이아스기, }} |
| 동의어 | Leptocheirus Merriam, 1903 Merriamia Boulenger, 1904 |
| 하위 분류 | †T. californicus Merriam, 1903 (모식종) †T. zitelli (Merriam, 1903) |
2. 특징

토레토크네무스는 멸종된 어룡의 한 속으로, 미국 캘리포니아주의 트라이아스기 지층에서 화석이 발견되었다. 1903년 존 캠벨 메리암에 의해 명명되었으며, '토레토스'와 '크네메'라는 그리스어에서 유래한 속명은 '구멍이 있는 다리'를 의미한다.
토레토크네무스는 2m 이상 자라지 않았을 것으로 보이며, 일부 추정으로는 두 종 모두 약 1m 정도의 작은 크기였을 것으로 보인다.[13][14] 수중 생활에 잘 적응한 해양 파충류로, 큰 눈과 긴 주둥이를 가지고 있었으며, 매끄러운 피부에는 비늘이 없거나 매우 작았다. 어룡의 사지는 단단한 지느러미로 변형되었으며, 등지느러미와 꼬리지느러미를 가지고 있었다.
1908년에 메리암은 토레토크네무스와 같은 트라이아스기 어룡이 수영할 때 꼬리 외에 지느러미를 추진력으로 사용했을 것이라고 추정했다.[15] 그러나 이후 연구들은 어룡의 지느러미가 주로 몸의 균형을 잡고 방향을 전환하는 데 사용되었을 것이라고 보고 있다.[10][9] 어룡은 꼬리를 좌우로 흔들어 앞으로 나아갔을 것으로 보이며, 이는 지속적인 수영에 더 효율적인 방법이었을 것이다.
어룡은 육지에서 움직일 수 없었지만 공기를 호흡해야 했다.[9] 높은 신진대사를 가진 항온 동물이었으며,[23] 알을 낳는 대신 물속에서 새끼를 낳았다.[9] 트라이아스기 어룡은 후대의 어룡들만큼 수영 실력이 뛰어나지 않았기 때문에, 먹이를 숨어서 기다리는 매복 포식자였을 가능성이 있다.[24]
2. 1. 해부학적 특징
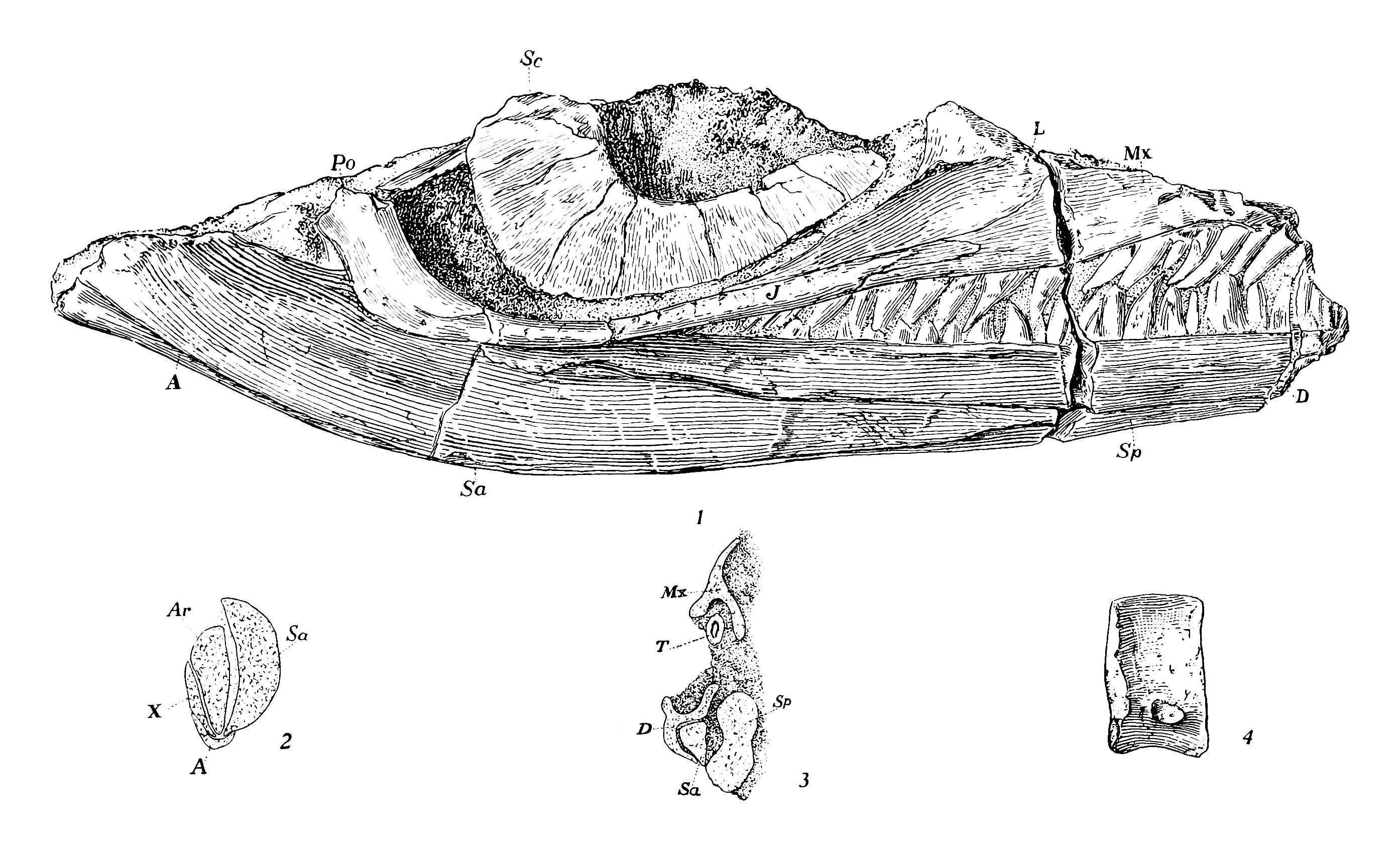
''토레토크네무스''(Toretocnemus)는 바다 생활에 잘 적응한 해양 파충류인 어룡의 한 속이다.[5] 어룡은 큰 눈과 일반적으로 긴 주둥이를 가진 유선형 몸매를 가지고 있었고, 매끄러운 피부에는 비늘이 없거나 매우 작았다.[9][10][11] 어룡의 사지는 단단한 지느러미로 변형되었으며, 등지느러미와 꼬리지느러미를 가지고 있었다.[9][10][12][20] ''토레토크네무스''는 몸길이가 2m를 넘지 않았을 것으로 보이며, 일부는 두 종 모두 약 1m 정도로 작았을 것으로 추정한다.[13][14] 2019년 앤더슨과 동료들은 알래스카 표본 UAMES 3599의 전체 길이를 1.5m 미만으로 추정했다.[8]
토레토크네무스는 두개골, 척추, 갈비뼈, 부속 골격에서 다른 어룡들과 구분되는 특징들을 보인다.
- 두개골: 안와(눈구멍)가 매우 크고, 관골이 좁다. 공막환을 이루는 골판은 매우 크다. 각골 노출은 크기가 매우 제한적이며, 뒤쪽 부분의 외부 표면 대부분은 상각골이 차지한다.[13][5] 분절골과 관절골은 모두 확대되어 있다.[5] 이빨은 모두 같은 모양이며, 홈에서 솟아난다.[5]
- 척추와 갈비뼈: ''T. californicus''의 가장 뒷부분 등뼈는 뒤로 굽은 신경 가시와 관절돌기 대신 노치를 가지고 있다. 얇은 신경궁은 측면에 얇은 능선을 가지고 있다.[15][5] 등쪽 중심은 ''T. zitteli''에서 길이와 높이가 유사한 반면, ''T. californicus''는 중심이 훨씬 덜 길쭉하다.[13] 등쪽 중심은 폭보다 높고 앞뒤 면이 모두 오목하다.[7] 갈비뼈와 연결되는 돌기인 횡돌기와 방횡돌기는 ''T. californicus''에서 작고 분리되어 있으며, 그 사이에 상당한 공간이 있다.[5] 이것은 속의 특징이다.[7] 등쪽 갈비뼈의 앞뒤 면은 하단을 제외하고 길이의 대부분을 따라 홈이 있다. ''T. zitteli''의 유일하게 잘 보존된 등쪽 갈비뼈 머리는 이분되지 않았지만, ''T. californicus''의 등쪽 갈비뼈 머리는 몸통의 중간에서 뒤쪽 부분까지 강하게 이분된다. 척주에서 더 앞쪽에 위치한 갈비뼈 머리는 더 두드러지게 이분된다. ''T. zitteli''는 5세트의 복늑골을 가지고 있다.[5] 꼬리 척추의 신경 가시는 매우 길쭉하다.[13] ''T. californicus''에서 꼬리 척추는 등쪽 척추보다 더 곧은 신경 가시와 더 긴 관절돌기를 가지며, 매우 높은 횡돌기를 가진다. 이 종의 모든 꼬리 중심은 양쪽 오목형이다. 꼬리 갈비뼈는 등쪽 갈비뼈와 달리 이분된 머리를 가지고 있지 않다. ''T. zitteli''는 긴 꼬리 척추를 가진다. 유일하게 양호한 상태로 알려진 꼬리 중심은 꼬리의 앞부분에서 왔으며, 높이보다 넓고 양쪽 오목형이다. 중심의 측면은 바깥쪽으로 구부러지지 않으며, 척추 몸체는 육각형 단면을 가진다.[5] 꼬리 아래쪽 굽힘은 약하며, 상단이 하단보다 긴 꼬리 중심에 의해 형성되어 쐐기 모양을 하고 있다. 2019년 앤더슨과 동료들은 굽힘 각도가 최소 6.45도였을 것이라고 발견했다.[8] 횡돌기는 측면으로 멀리 뻗어 있고 세브론을 위한 면은 잘 구분된다. ''T. californicus''의 길쭉한 세브론은 Y자 모양이다.[13][5]
- 부속 골격: 둥근 오구돌기는 폭보다 길이가 두 배 길며 가장자리에 오목한 부분이 없다. 내측은 외측보다 더 튼튼하다.[5] 짧은 어깨뼈의 앞 가장자리에는 갈고리 모양의 돌출부가 있다.[15] 오구돌기와 상완골이 연결되는 어깨뼈의 표면은 분리되어 있다. 어깨뼈의 하단은 넓다. 서로 접촉하는 빗장뼈도 상당히 넓다. 메리엄은 삼각형 뼈를 ''T. zitteli''의 간쇄골로 잠정적으로 확인했다.[5] 추정되는 간쇄골은 작고 뒤쪽으로의 돌출부가 매우 짧다.[15] ''T. californicus''의 긴 장골은 꽤 좁다. 치골과 좌골(하부 엉덩이뼈)은 편평하고 각 쌍은 정중선을 따라 광범위하게 접촉한다.[15][16] 넓은 치골(앞쪽 하부 엉덩이뼈)은 각각 폐쇄공이라고 하는 작은 구멍으로 뚫려 있으며, 이는 ''킴보스폰딜루스''와 유사하다.[5][15] 좌골은 ''샤스타사우루스''보다 넓지 않다.[5] 두 종은 유사한 사지를 가지고 있다.[15] 1903년 메리엄은 ''T. zitteli''의 앞다리가 뒷다리보다 길다고 언급한 반면, ''T. californicus''의 뒷다리는 앞다리만큼 길거나 더 길다고 말했다.[5] 그러나 1999년, 모타니는 두 표본 모두 분리되고 불완전하므로 이러한 차이는 의심스럽다고 지적했다.[17] 두 종의 상부 및 하부 사지 뼈는 매우 길다.[5] 넓은 하단을 가진 좁은 상완골(위팔뼈)을 가지고 있으며, 이 뼈는 요골보다 두 배 길다. 상완골의 상단은 뼈의 앞면과 거의 일치하는 반면, 상완골의 뒷 가장자리는 대각선이다. 상완골 앞 가장자리의 중간 지점에 잘 정의된 노치가 있으며, 뼈의 뒷 가장자리에도 노치가 있지만 강하게 구분되지는 않는다. 아래팔뼈의 면은 안쪽으로 휘어져 있다. ''T. zitteli''의 상완골 상단은 상단과 하단 모두에 상완골 길이의 절반에 해당하는 능선이 있으며, 하단의 능선이 더 크다. 또 다른 상완골의 융기된 영역은 척골의 유사한 구조로 보완된다.[5] 두 종의 하부 사지 뼈(말단골) 사이에는 구멍이 있다.[5] 요골의 내측과 외측 가장자리는 모두 오목하지만 척골의 내측 가장자리에만 노치가 있다.[15] 잘 알려져 있지는 않지만, ''T. californicus''의 앞다리는 ''T. zitteli''의 앞다리와 유사해 보이지만, ''T. californicus''의 요골은 ''T. zitteli''의 요골보다 약간 더 넓고 전자의 앞다리 뼈의 노치는 더 뚜렷하다. ''T. zitteli''의 손목뼈는 세 개의 행으로 배열된다. 하나의 손목뼈만 중간 상완골의 하단 가장자리에 연결된다. 중간 뼈는 요골보다 척골과 더 광범위하게 접촉하며, 요골골이 척골골(각각 앞과 뒤의 상완골)보다 넓기 때문이다. 지느러미의 바깥쪽 가장자리를 형성하는 손목뼈(손가락 뼈)는 바깥쪽을 향한 가장자리에 노치가 있다.[13][5] 첫 번째 손가락의 중수골의 앞면과 뒷면 가장자리 모두에 노치가 있다.[5] 각 사지는 세 개의 주요 손가락과 매우 작은 네 번째 손가락을 가지고 있다. ''T. zitteli''의 앞다리의 바깥쪽 두 개의 주요 손가락은 각각 일곱 개의 손가락뼈로 구성되어 있으며 가운데 손가락은 여덟 개를 가지고 있다. 손가락 끝 부분의 손가락뼈가 매우 섬세하기 때문에 메리엄은 두 개 이상이 누락되었을 가능성은 낮다고 생각했다. ''T. californicus''의 앞다리에서 첫 번째 손가락의 모든 손가락뼈의 양쪽은 노치가 있고, 그 바로 뒤의 손가락의 모든 손가락뼈(가장 위쪽 손가락뼈 제외)에도 노치가 있다. ''T. zitteli''의 손가락뼈 중에서 가운데 주요 손가락의 첫 번째와 일부 말단 손가락뼈에만 노치가 없다. 앞지느러미에서 중간 손가락의 모든 노치된 손가락뼈는 배쪽을 향한 노치만 가지고 있다.[5] 바깥쪽 주요 손가락의 일부 손가락뼈는 이중 노치가 있고, 모두 바깥쪽을 향한 가장자리에 노치가 있다.[13][5] 앞다리에서 부속 손가락에는 단 두 개의 매우 작은 손가락뼈만 알려져 있지만, 더 많았을 수 있다. 이러한 손가락뼈는 다른 손가락의 위쪽 영역 근처에 위치한다.[5] 넙다리뼈의 길쭉한 부분(몸통)은 좁지만, 뼈의 하단은 매우 넓다.[13] 넙다리뼈의 중간 부분은 비틀어져 있고, 뼈의 길이는 경골과 비골의 길이의 약 두 배이다. 넙다리뼈의 상단은 윗면에 큰 돌출된 선반을 가지고 있고, 넙다리뼈의 하단은 정강이뼈에 부착되는 부분에서 안쪽으로 굽어 있다. 경골(앞 정강이뼈)은 비골(뒤 정강이뼈)보다 넓고, 비골은 넙다리뼈를 넘어 뒤쪽으로 기울어져 있지 않다.[5][17][13] 비골의 뒷 가장자리는 경골의 오목한 가장자리와 달리 직선이며, 비골의 앞쪽도 직선이다. ''T. zitteli''의 경골과 비골은 앞면과 뒷면에 모두 노치가 있으며, 전자의 뼈가 더 넓다. 손목뼈와 마찬가지로 ''T. zitteli''의 발목뼈는 세 개의 행으로 배열된다.[5] ''T. californicus''의 발목뼈 중 많은 부분이 사각형 윤곽을 가지고 있지만, 중간 상부 발목뼈는 오각형이다. 첫 번째 손가락 위의 발목뼈는 앞쪽 가장자리에 노치가 있다. 그러나 III번 손가락 위의 발목뼈에는 노치가 없고, 그 중족골에도 노치가 없다. 비골 아래의 발목뼈(비골골)의 뒷 가장자리는 매우 약하게 노치되어 있다. ''T. zitteli''의 중간 하부 발목뼈는 볼록한 가장자리를 가지고 있는 반면, 그 바로 뒤에 있는 발목뼈는 뒷 가장자리에 큰 들여쓰기가 있다. 그 아래의 중족골은 모양이 비슷하지만, 가장 뒤쪽의 중족골은 뒷면 노치 외에도 앞면 노치가 있다. ''T. californicus''의 비골골은 그 아래의 발목뼈와 함께 부속 손가락의 부착을 위한 표면을 가지고 있다. ''T. zitteli''의 뒷다리의 손가락 배열은 앞다리의 손가락 배열과 유사하다. 뒷다리에 있는 부속 손가락은 앞다리의 부속 손가락보다 더 발달되어 있으며, 각각 네 개의 뼈로 구성되어 있고 발목뼈 뒤에서 시작된다.[5]
2. 1. 1. 두개골
토레토크네무스의 안와(눈구멍)는 매우 크고, 관골(각 안와의 하단을 형성하는 뼈)은 좁다. 안와 내에 위치한 구조인 공막환을 이루는 골판은 매우 크다. 토레토크네무스의 각골(아래턱 뒤쪽 외부 하악골) 노출은 크기가 매우 제한적이며, 뒤쪽 부분의 외부 표면 대부분은 상각골에 의해 차지된다.[13][5] 분절골(한 쌍의 내부 하악골)과 관절골(턱 관절에 관여하는 하악골) 모두 ''토레토크네무스''에서 확대되었으며, 특히 후자의 경우 더욱 그렇다. ''토레토크네무스''의 이빨은 모두 같은 모양이며, 홈에서 솟아난다.[5]2. 1. 2. 척추와 갈비뼈
''T. californicus''의 가장 뒷부분 등뼈는 뒤로 굽은 신경 가시와 관절돌기 (앞 또는 뒤로 향하는 두 쌍의 돌기) 대신 노치를 가지고 있다. ''T. californicus''의 얇은 신경궁은 측면에 얇은 능선을 가지고 있다.[15][5] 등쪽 중심은 ''T. zitteli''에서 유사한 길이와 높이를 가지는 반면, ''T. californicus''는 중심이 훨씬 덜 길쭉하다.[13] 등쪽 중심은 폭보다 높고 앞면과 뒷면 모두 오목하다(양쪽 오목형).[7] 갈비뼈와 연결되는 돌기인 등뼈의 횡돌기와 방횡돌기는 ''T. californicus''에서 작고 분리되어 있으며 그 사이에 상당한 공간이 있는데,[5] 이것은 속의 특징이다.[7] ''Toretocnemus''의 등쪽 갈비뼈의 앞면과 뒷면은 하단을 제외하고 길이의 대부분을 따라 홈이 있다. ''T. zitteli''의 유일하게 잘 보존된 등쪽 갈비뼈 머리는 이분되지 않았지만, 반대로 ''T. californicus''의 등쪽 갈비뼈 머리는 몸통의 중간에서 뒤쪽 부분까지 강하게 이분된다. 척주에서 더 앞쪽에 위치한 갈비뼈 머리는 더 두드러지게 이분된다. ''T. zitteli''는 5세트의 복늑골을 가지고 있다.[5]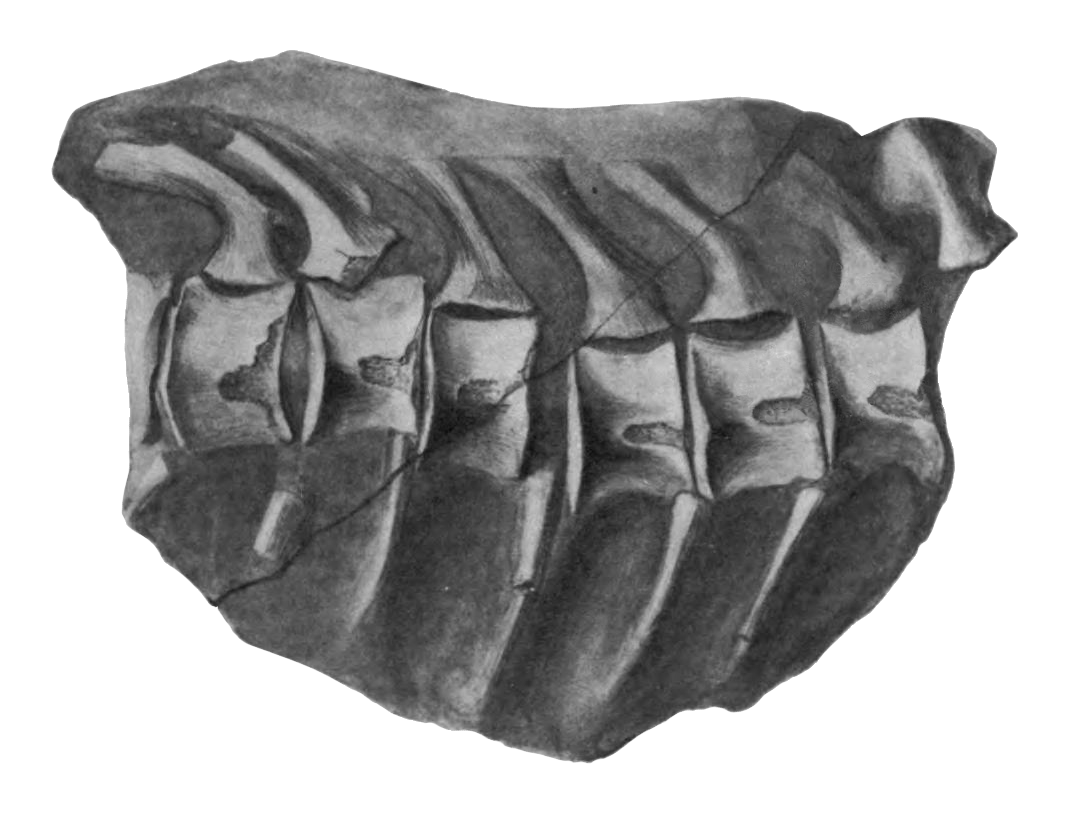
꼬리 척추의 신경 가시(척추의 위쪽 돌기)는 ''Toretocnemus''에서 매우 길쭉하다.[13] ''T. californicus''에서 꼬리 척추는 등쪽 척추보다 더 곧은 신경 가시와 더 긴 관절돌기를 가지고 있으며, 이 꼬리 척추는 또한 매우 높은 횡돌기를 가지고 있다. 이 종의 모든 꼬리 중심은 양쪽 오목형이다. ''T. californicus''의 꼬리 갈비뼈는 등쪽 갈비뼈와 달리 이분된 머리를 가지고 있지 않다. ''T. zitteli''는 긴 꼬리 척추를 가지고 있다. ''T. zitteli''의 유일하게 양호한 상태로 알려진 꼬리 중심은 꼬리의 앞부분에서 왔으며, 높이보다 넓고 양쪽 오목형이다. 중심의 측면은 바깥쪽으로 구부러지지 않으며, 척추 몸체는 육각형 단면을 가지고 있다.[5] ''Toretocnemus''의 꼬리 아래쪽 굽힘은 약하며, 상단이 하단보다 긴 꼬리 중심에 의해 형성되어 쐐기 모양을 하고 있다. 2019년 앤더슨과 동료들은 굽힘 각도가 최소 6.45도였을 것이라고 발견했다.[8] 횡돌기는 측면으로 멀리 뻗어 있고 세브론을 위한 면은 잘 구분된다. ''T. californicus''의 길쭉한 세브론은 Y자 모양이다.[13][5]
2. 1. 3. 부속 골격
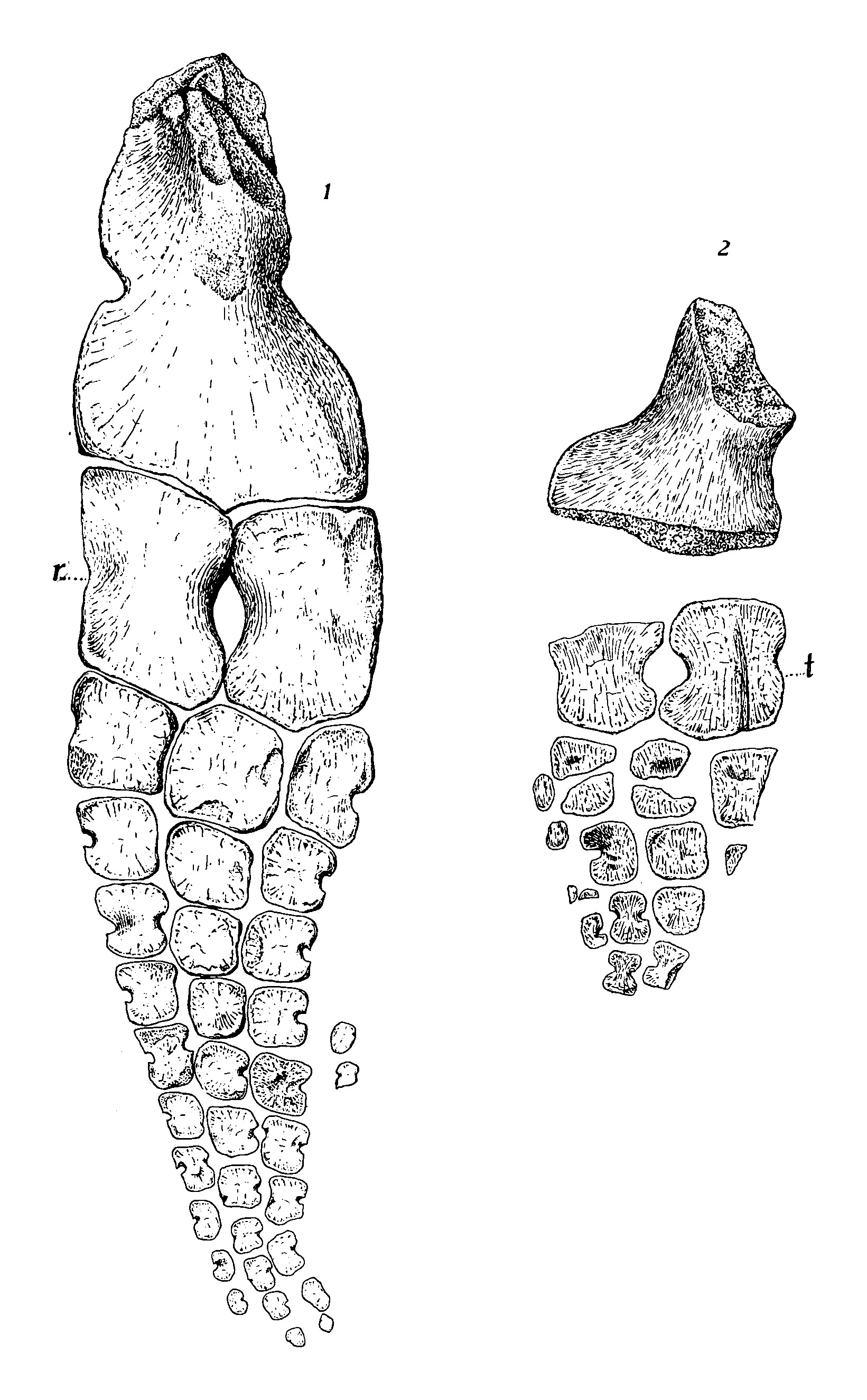
둥근 오구돌기는 폭보다 길이가 두 배 길며 가장자리에 오목한 부분이 없다. 내측은 외측보다 더 튼튼하다.[5] 짧은[5] 어깨뼈의 앞 가장자리에는 갈고리 모양의 돌출부가 있다.[15] 오구돌기와 상완골이 연결되는 어깨뼈의 표면은 분리되어 있다. 어깨뼈의 하단은 넓다. 서로 접촉하는 빗장뼈도 상당히 넓다. 메리엄은 삼각형 뼈를 ''T. zitteli''의 간쇄골로 잠정적으로 확인했다.[5] 추정되는 간쇄골은 작고 뒤쪽으로의 돌출부가 매우 짧다.[15] ''T. californicus''의 긴 상부 엉덩이뼈인 장골은 꽤 좁다. ''T. californicus''의 치골과 좌골(하부 엉덩이뼈)은 편평하고[15] 각 쌍은 정중선을 따라 광범위하게 접촉한다.[16] ''T. californicus''의 넓은 치골(앞쪽 하부 엉덩이뼈)은 각각 폐쇄공이라고 하는 작은 구멍으로 뚫려 있으며,[5] 이는 ''킴보스폰딜루스''와 유사하다.[15] 좌골은 ''샤스타사우루스''보다 넓지 않다.[5]
두 종의 ''Toretocnemus''는 유사한 사지를 가지고 있다.[15] 1903년 메리엄은 ''T. zitteli''의 앞다리가 뒷다리보다 길다고 언급한 반면, ''T. californicus''의 뒷다리는 앞다리만큼 길거나 더 길다고 말했다.[5] 그러나 1999년, 모타니는 두 표본 모두 분리되고 불완전하므로 각 표본에 할당된 사지가 모두 동일한 개체에서 나온 것인지 확인할 수 없으므로 이러한 차이는 의심스러운 효용성이 있다고 지적했다.[17] 두 종의 상부 및 하부 사지 뼈는 매우 길다.[5] ''Toretocnemus''는 넓은 하단을 가진 좁은 상완골(위팔뼈)을 가지고 있으며[13] 이 뼈는 요골보다 두 배 길다. 상완골의 상단은 뼈의 앞면과 거의 일치하는 반면, 상완골의 뒷 가장자리는 대각선이다. 상완골 앞 가장자리의 중간 지점에 잘 정의된 노치가 있으며, 뼈의 뒷 가장자리에도 노치가 있지만 강하게 구분되지는 않는다. 아래팔뼈의 면은 안쪽으로 휘어져 있다. ''T. zitteli''의 상완골 상단은 상단과 하단 모두에 상완골 길이의 절반에 해당하는 능선이 있으며, 하단의 능선이 더 크다. 또 다른 상완골의 융기된 영역은 척골의 유사한 구조로 보완된다.[5]
두 종의 하부 사지 뼈(말단골) 사이에는 구멍이 있다.[5] 요골의 내측과 외측 가장자리는 모두 오목하지만 척골의 내측 가장자리에만 노치가 있다.[15] 잘 알려져 있지는 않지만, ''T. californicus''의 앞다리는 ''T. zitteli''의 앞다리와 유사해 보이지만, ''T. californicus''의 요골은 ''T. zitteli''의 요골보다 약간 더 넓고 전자의 앞다리 뼈의 노치는 더 뚜렷하다. ''T. zitteli''의 손목뼈는 세 개의 행으로 배열된다. 하나의 손목뼈만 중간 상완골의 하단 가장자리에 연결된다. 중간 뼈는 요골보다 척골과 더 광범위하게 접촉하며, 요골골이 척골골(각각 앞과 뒤의 상완골)보다 넓기 때문이다. 지느러미의 바깥쪽 가장자리를 형성하는 ''Toretocnemus''의 손목뼈(손가락 뼈)는 바깥쪽을 향한 가장자리에 노치가 있다.[13][5] 첫 번째 손가락의 중수골의 앞면과 뒷면 가장자리 모두에 노치가 있다.[5]
''Toretocnemus''의 각 사지는 세 개의 주요 손가락과 매우 작은 네 번째 손가락을 가지고 있다. ''T. zitteli''의 앞다리의 바깥쪽 두 개의 주요 손가락은 각각 일곱 개의 손가락뼈로 구성되어 있으며 가운데 손가락은 여덟 개를 가지고 있다. 손가락 끝 부분의 손가락뼈가 매우 섬세하기 때문에 메리엄은 두 개 이상이 누락되었을 가능성은 낮다고 생각했다. ''T. californicus''의 앞다리에서 첫 번째 손가락의 모든 손가락뼈의 양쪽은 노치가 있고, 그 바로 뒤의 손가락의 모든 손가락뼈(가장 위쪽 손가락뼈 제외)에도 노치가 있다. ''T. zitteli''의 손가락뼈 중에서 가운데 주요 손가락의 첫 번째와 일부 말단 손가락뼈에만 노치가 없다. ''T. zitteli''의 앞지느러미에서 중간 손가락의 모든 노치된 손가락뼈는 배쪽을 향한 노치만 가지고 있다.[5] 바깥쪽 주요 손가락의 일부 손가락뼈는 이중 노치가 있고, 모두 바깥쪽을 향한 가장자리에 노치가 있다.[13][5] ''T. zitteli''의 앞다리에서 부속 손가락에는 단 두 개의 매우 작은 손가락뼈만 알려져 있지만, 더 많았을 수 있다. 이러한 손가락뼈는 다른 손가락의 위쪽 영역 근처에 위치한다.[5]
''Toretocnemus''의 넙다리뼈의 길쭉한 부분(몸통)은 좁지만, 뼈의 하단은 매우 넓다.[13] 넙다리뼈의 중간 부분은 비틀어져 있고, 뼈의 길이는 경골과 비골의 길이의 약 두 배이다. 넙다리뼈의 상단은 윗면에 큰 돌출된 선반을 가지고 있고, 넙다리뼈의 하단은 정강이뼈에 부착되는 부분에서 안쪽으로 굽어 있다. 경골(앞 정강이뼈)은 비골(뒤 정강이뼈)보다 넓고,[5] 비골은 넙다리뼈를 넘어 뒤쪽으로 기울어져 있지 않다.[17][13] 비골의 뒷 가장자리는 경골의 오목한 가장자리와 달리 직선이며, 비골의 앞쪽도 직선이다. ''T. zitteli''의 경골과 비골은 앞면과 뒷면에 모두 노치가 있으며, 전자의 뼈가 더 넓다. 손목뼈와 마찬가지로 ''T. zitteli''의 발목뼈는 세 개의 행으로 배열된다.[5] ''T. californicus''의 발목뼈 중 많은 부분이 사각형 윤곽을 가지고 있지만, 중간 상부 발목뼈는 오각형이다. 첫 번째 손가락 위의 발목뼈는 앞쪽 가장자리에 노치가 있다. 그러나 III번 손가락 위의 발목뼈에는 노치가 없고, 그 중족골에도 노치가 없다. 비골 아래의 발목뼈(비골골)의 뒷 가장자리는 매우 약하게 노치되어 있다. ''T. zitteli''의 중간 하부 발목뼈는 볼록한 가장자리를 가지고 있는 반면, 그 바로 뒤에 있는 발목뼈는 뒷 가장자리에 큰 들여쓰기가 있다. 그 아래의 중족골은 모양이 비슷하지만, 가장 뒤쪽의 중족골은 뒷면 노치 외에도 앞면 노치가 있다. ''T. californicus''의 비골골은 그 아래의 발목뼈와 함께 부속 손가락의 부착을 위한 표면을 가지고 있다. ''T. zitteli''의 뒷다리의 손가락 배열은 앞다리의 손가락 배열과 유사하다. ''T. zitteli''의 뒷다리에 있는 부속 손가락은 앞다리의 부속 손가락보다 더 발달되어 있으며, 각각 네 개의 뼈로 구성되어 있고 발목뼈 뒤에서 시작된다.[5]
2. 2. 생태
''토레토크네무스''(Toretocnemus)는 바다 생활에 잘 적응한 해양 파충류 그룹인 어룡에 속한다.[5] 어룡은 큰 눈과 일반적으로 길쭉한 주둥이를 가진 유선형 몸매를 가지고 있었으며, 매끄러운 피부에는 매우 작은 비늘이 있거나 비늘이 전혀 없었다.[9][10][11] 어룡의 사지는 단단한 지느러미로 크게 변형되었고, 등지느러미와 꼬리지느러미를 가지고 있었는데, 꼬리지느러미는 꼬리의 아래쪽 굽힘으로 지지되었다.[10][12][20] ''토레토크네무스''는 2m보다 크게 자라지 않았을 것이며,[13] 일부 추정치는 두 종 모두 약 1m에 불과하여 이 그룹의 작은 구성원이었다.[14] 2019년, 앤더슨과 동료들은 알래스카 표본 UAMES 3599의 전체 길이를 1.5m 미만으로 추정했다.[8]
2. 2. 1. 수영 방식
1908년 메리암은 토레토크네무스와 같은 트라이아스기 어룡이 수영할 때 꼬리 외에도 사지를 추진력으로 사용했을 것이라고 생각했다.[15] 그러나 물고기와 비교한 더 많은 연구는 어룡의 사지를 노처럼 사용하는 것에 대해 의문을 제기했다. 대신, 그들은 아마도 느린 수영에 국한된 추진 기능인 동물을 똑바로 유지하고 회전시키는 역할을 했을 것이다.[10][9] 어룡은 꼬리를 앞뒤로 흔들면서 앞으로 나아갔을 것이고, 이는 지속적인 수영에 더 효율적인 방법이었다.[10][9] T. zitteli의 사지 앞쪽 가장자리는 위팔뼈의 상단부가 하단부에 상대적으로 위치해 있기 때문에 뒤쪽 가장자리보다 근력이 약간 더 강했을 것이다.[5]어룡은 골격 지지 구조의 상당 부분을 잃었기 때문에 육지에서 이동할 수 없었으며, 대신 평생을 물속에서 보냈지만 여전히 공기를 호흡했다.[9] 현대의 변온 동물 ("냉혈") 파충류와 달리, 어룡은 높은 신진대사를 가지고 있었고, 일정한 체온을 유지할 수 있는 항온 동물이었다.[23] 어룡은 알을 낳기 위해 육지로 나갈 수 없었기 때문에 (파충류 알은 물속에서 익사할 것이다), 새끼를 낳았다.[9] 트라이아스기 어룡은 후기 친척들만큼 효율적인 수영을 하지 못했기 때문에 매복 포식자였을 수 있다.[24] 어룡의 큰 눈은 시각이 중요한 감각이었음을 나타내며,[9] 콧구멍이 폐가 아닌 입으로 이어져 있었기 때문에 물속에서 먹이와 다른 동물의 냄새를 맡을 수 있었다.[25]
3. 분류
1903년 메리엄은 토레토크네무스의 분류에 대해 확신하지 못했다. 그는 ''T. zitteli''(당시 ''Leptocheirus'')가 믹소사우루스와 상당히 유사하지만, 치아와 부속지 해부학에서 뚜렷한 차이를 보인다고 여겼다. 또한 ''T. californicus''가 ''T. zitteli'', 샤스타사우루스, 어룡과 유사한 점이 있다고 언급했다.[5] 1904년 불랑제는 어룡의 세 가지 주요 계통, 즉 넓은 지느러미의 ''Ichthyosaurus'' 종, 좁은 지느러미의 ''Ichthyosaurus''(''프로테오사우루스''로 지칭), ''안룡''으로 이어지는 계통이 있다고 보았다. ''Toretocnemus''와 ''Merriamia''(''T. zitteli'')는 두 번째 계통에 속하는 것으로 여겨졌다.[6]
1908년 메리엄은 어룡에 대한 잠정적인 분류 체계를 제안했다. 그는 트라이아스기 종이 모두 믹소사우루스과에 속하며, 이는 믹소사우루스아과와 샤스타사우루스아과로 나뉘고, ''Merriamia''는 후자에 속한다고 보았다. ''T. californicus'' 역시 팔다리가 ''Merriamia''와 유사하여 샤스타사우루스아과로 분류되었지만, 메리엄은 이 종의 두 개의 머리를 가진 갈비뼈가 그러한 위치를 반박하는 것처럼 보여 더 신중하게 접근했다.[15]
1923년 프리드리히 폰 후에네는 어룡을 다른 방식으로 분류하여, 그들이 가진 손가락의 수에 따라 롱기피나테와 라티피나테로 나누었다. 폰 후에네는 ''Toretocnemus''가 ''스테노프테리지우스'', ''플라티프테리지우스'', ''난노프테리지우스''로 이어지는 계열에서 롱기피나테에 속한다고 여겼다.[18][19]
1999년 모타니는 ''Toretocnemus''를 전통적으로 분류되었던 샤스타사우루스과 대신 ''캘리포르노사우루스'' 및 파르비펠비아와 함께 진어룡류 내에서 회복시켰다.[17] 2000년 마이쉬와 마츠케는 ''Toretocnemus''와 ''치아니치오사우루스''를 결합하기 위해 새로운 과인 토레토크네무스과를 명명했다. 그들은 이 그룹이 심보스폰딜루스 및 파르비펠비아를 포함한 더 큰 그룹인 롱기피나티 내에서 처음으로 분화된 그룹임을 발견했다.[13] 2002년 엘리자베스 니콜스와 동료들은 거의 완전한 ''치아니치오사우루스'' 골격을 묘사하고 ''Toretocnemus''와 많은 유사점을 발견했다. 그들은 둘이 토레토크네무스과 내에서 자매 분류군이라는 데 동의했다.[2]
2016년 청 지와 동료들의 분석 결과 ''Toretocnemus''와 ''치아니치오사우루스''가 토레토크네무스과 내에서 결합되었지만, 이 그룹이 진어룡류 내에 있다는 것을 발견했다.[16] 2017년 벤자민 문은 어룡 관계에 대한 여러 포괄적인 분석을 수행했다. ''Toretocnemus''는 여러 번 잘 뒷받침되는 클레이드(조상의 모든 후손을 포함하는 자연 그룹)로 발견되었지만, 진어룡류 내에서 자주 발견되었지만, ''치아니치오사우루스''의 자매 분류군으로 회복되지 않았다. 그러나 그의 결과 중 하나는 ''Toretocnemus''를 ''치아니치오사우루스'', ''캘리포르노사우루스'', ''위마니우스'', ''팔라로돈 메이저''와 함께 클레이드에 포함시켰지만, 이 그룹화는 지지도가 높지 않았다.[20]
4. 고생물학
''토레토크네무스''(Toretocnemus)는 바다 생활에 잘 적응한 해양 파충류인 어룡의 한 속이다.[5] 어룡은 큰 눈과 길쭉한 주둥이를 가진 유선형 몸체를 가지고 있었고, 매끄러운 피부에는 비늘이 없거나 매우 작았다.[9][10][11] 어룡의 사지는 단단한 지느러미로 변형되었으며, 등지느러미와 꼬리지느러미를 가지고 있었다. 꼬리지느러미는 꼬리 아래쪽이 굽은 형태로 지지되었다.[10][12][20] ''토레토크네무스''는 이상 자라지 않았을 것으로 추정되며,[13] 일부에서는 두 종 모두 약 에 불과하여 어룡 중에서도 작은 크기였다고 본다.[14] 2019년 앤더슨과 동료들은 알래스카 표본 UAMES 3599의 전체 길이를 미만으로 추정했다.[8]
어룡은 골격 구조상 육지에서 이동할 수 없었지만, 공기 호흡을 했다.[9] 현대의 변온 동물 파충류와 달리, 어룡은 높은 신진대사를 가진 항온 동물이었다.[23] 어룡은 알을 낳기 위해 육지로 올라올 수 없었기 때문에 (파충류 알은 물속에서 익사한다), 새끼를 낳았다.[9]
1908년, 메리암은 ''토레토크네무스''와 같은 트라이아스기 어룡이 수영할 때 꼬리 외에 사지를 추진력으로 사용했다고 생각했다.[15] 그러나 이후 연구와 어류와의 비교를 통해 어룡이 사지를 노처럼 사용했을 것이라는 주장에 의문이 제기되었다. 대신 사지는 몸을 똑바로 유지하고 회전하는 데 사용되었으며, 추진 기능은 느린 수영에 국한되었을 것으로 보인다. 어룡은 꼬리를 좌우로 흔들어 앞으로 나아갔으며, 이는 지속적인 수영에 더 효율적인 방법이었다.[10][9] ''T. zitteli''는 위팔뼈의 상단이 하단에 상대적으로 위치하기 때문에 사지 앞쪽 가장자리가 뒤쪽 가장자리보다 근력이 약간 더 강했을 것이다.[5]
어룡은 해양 환경에 살면서 염분이 많은 먹이를 먹었기 때문에, 몸에서 여분의 소금을 제거하기 위한 염분샘을 가지고 있었을 것이다. 트라이아스기 어룡은 후대 어룡들만큼 수영을 잘 하지 못했기 때문에 매복 포식자였을 수 있다.[24] 어룡의 큰 눈은 시각이 중요한 감각이었음을 나타내며,[9] 콧구멍이 폐가 아닌 입으로 이어졌기 때문에 물속에서 먹이나 다른 동물의 냄새를 맡을 수 있었다.[25]
4. 1. 서식 환경
1903년에 미국 캘리포니아주의 트라이아스기 지층에서 미국의 고생물학자들에 의해 처음으로 화석이 발견되어 새롭게 명명된 ''토레토크네무스''는 중생대 트라이아스기 후기(2억 년 전~1억 8000만 년 전)에 북아메리카를 중심으로 하는 북동부 태평양에서 주로 서식했던 어룡이다.과거에는 네바다주와 밀접한 관련이 있는 것으로 추정되었지만, 멕시코 안티모니오 지층에서 ''토레토크네무스''가 발견되면서 루카스는 2002년에 안티모니오 테란이 동부 클래머스 테란과 연결되어 있었으며, 이곳이 호셀쿠스 석회암이 퇴적된 판의 일부라고 주장했다. 루카스는 안티모니오 지층의 대부분의 동물과는 달리, ''토레토크네무스''는 매우 제한적인 분포를 보이며 네바다주에서는 발견되지 않았다는 점에 주목했다. ''토레토크네무스''가 이동성이 있는 생물임에도 불구하고 그렇게 좁은 지역에서만 발견되는 이유에 대해, 루카스는 작은 크기 때문에 개방된 해역에서 수영을 잘 하지 못했을 것이라고 추측했다. 그러나 루카스는 만약 ''키아니크티오사우루스''가 ''토레토크네무스''와 동일한 종이라면, 이 고생물학적 연관성은 훨씬 약해질 것이라고 언급했다.
4. 2. 제한된 분포
멕시코 안티모니오 지층에서 토레토크네무스가 발견되면서, 2002년 루카스는 안티모니오 테란이 한때 호셀쿠스 석회암이 퇴적된 구조판인 동부 클래머스 테란과 연결되어 있었다고 주장했다. 루카스는 안티모니오 지층의 대부분의 동물들과 달리 토레토크네무스는 네바다주에서는 발견되지 않아 매우 제한된 분포를 보였다고 언급했다. 토레토크네무스가 이동성이 있는 생물임에도 불구하고 그렇게 좁은 지역에서만 발견되는 이유에 대해, 루카스는 작은 크기 때문에 개방된 해역에서 수영을 잘 하지 못했을 것이라고 추측했다. 그러나 루카스는 만약 키아니크티오사우루스가 토레토크네무스와 동의어라면, 이렇게 추정된 고생물학적 연관성은 훨씬 약해질 것이라고 언급했다.[1]5. 화석 발견
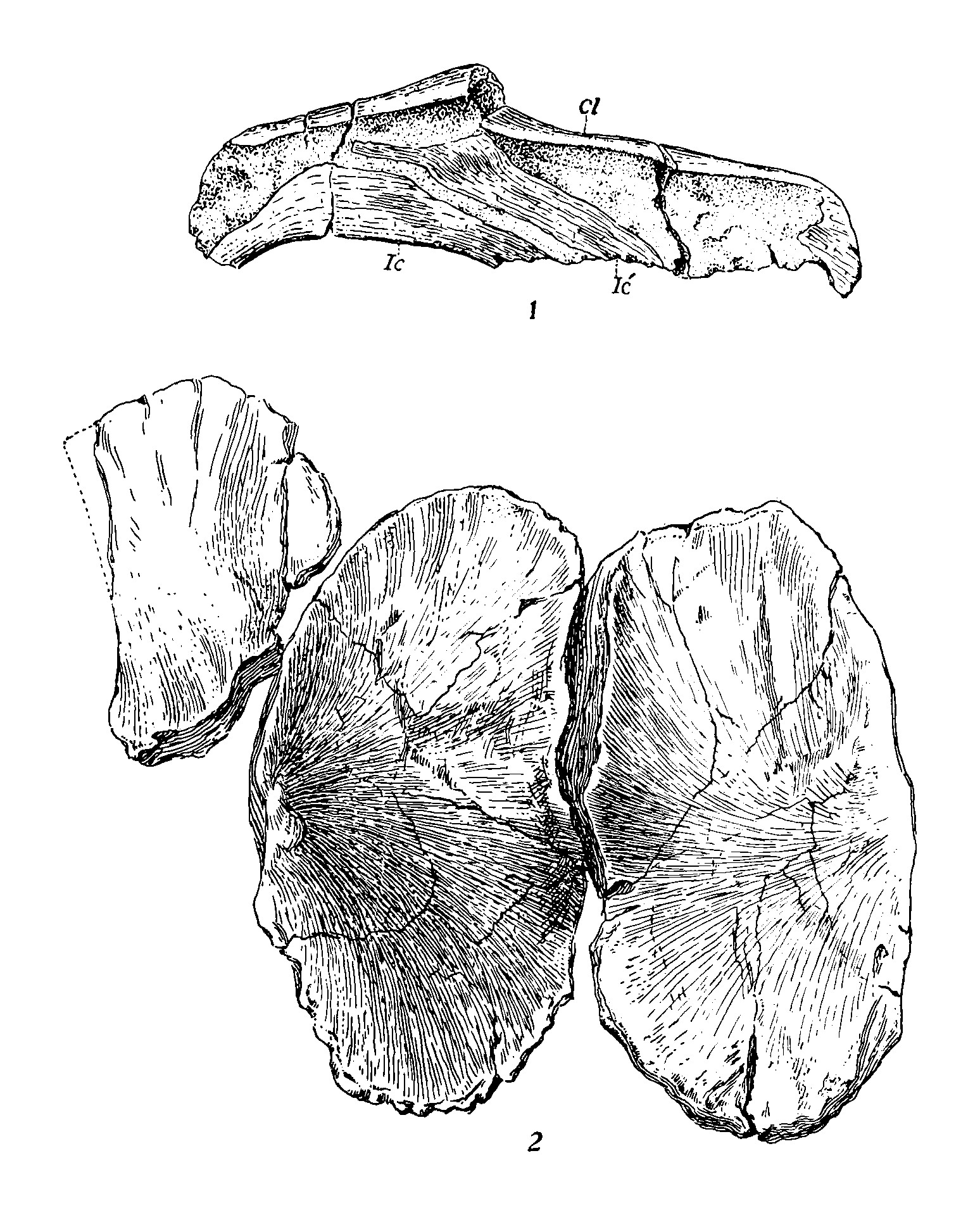
UCMP 8100으로 최종 분류될 표본은 미국 캘리포니아주 샤스타 군의 핏 강을 따라 애니 몬태규 알렉산더가 발견했다. 표본의 많은 부분이 침식으로 손실되었지만, 일부 두개골 조각, 등뼈를 포함한 약 30개의 척추뼈 (대부분은 앞쪽 꼬리뼈) , 많은 수의 갈비뼈, 골반, 오른쪽 앞다리, 왼쪽과 오른쪽 뒷다리 등 많은 요소들이 보존되었다.[15] 또 다른 표본인 UCMP 8099도 샤스타 군에서 알렉산더가 발견했다. 목장에서 발굴된 이 표본은 부분적인 두개골, 여러 불완전한 척추뼈, 잘 보존되지 않은 갈비뼈, 복부 갈비뼈, 어깨뼈, 앞다리, 부분적인 뒷다리로 구성되어 있다.[15] 이 두 표본은 모두 Hosselkus Limestone의 ''Trachyceras''대에서 발견되었다. 알렉산더는 1903년경 다른 여러 화석 파충류와 함께 이들을 캘리포니아 대학교에 기증했다.[5]
1903년 존 캠벨 메리엄이 ''토레토크네무스''를 명명했다.[5][3] 속명은 그리스어 ("뚫린")와 ("정강이뼈")에서 유래했다.[4] 모식종은 UCMP 8100을 완모식표본으로 하는 ''Toretocnemus californicus''이다. 같은 출판물에서 메리엄은 UCMP 8100에 대해 ''Leptocheirus zitteli''라는 다른 속과 종을 명명했다.[5] 그러나 ''Leptocheirus''라는 이름은 이미 다른 곳에 사용되고 있었고, 조지 앨버트 불렌저는 1904년에 이를 ''Merriamia''로 개명했다.[6] 1908년에 메리엄은 완모식표본과 같은 지역에서 나온 추가 표본들이 ''T. californicus''와 관련이 있을 수 있지만, 아직 처리되지 않았다고 언급했다.[15]
모타니 료스케는 ''Toretocnemus''와 ''Merriamia'' 사이의 여러 해부학적 유사점과 발견 장소의 근접성에 주목하여 두 속을 동의어로 만들고, 더 오래된 이름인 ''Toretocnemus''를 우선시했다. 그는 표본을 재연구하면서 메리엄이 제안한 특징들을 보존 상태 때문에 제대로 관찰할 수 없다는 것을 발견했다. 그러나 그는 표본 간의 겹치는 자료가 거의 없어 비교가 복잡했기 때문에 두 종(''T. californicus''와 ''T. zitteli'')을 잠정적으로 유효한 것으로 유지했다.[17] 2000년에 마이클 마이쉬와 안드레아스 마츠케는 이 동의어에 동의하면서도 ''T. californicus''와 ''T. zitteli''를 별개의 종으로 유지했다.[13]
2001년에는 ''Toretocnemus californicus''의 척추뼈가 멕시코 소노라의 엘 안티모니 광산 마을 근처 안티모니 형성에서 발견되었다. 이곳에서는 어룡 척추뼈가 흔히 발견되지만, 대부분은 더 큰 샤스타사우루스과 척추뼈였다. 안티모니 형성에서 발견된 척추뼈는 직경이 1.3cm에서 2.8cm로 훨씬 작았다. 신경 가시는 없었지만, 관절을 위한 명확한 홈이 있었다.[7]
2019년 논문에서 캐서린 앤더슨은 패트릭 드루켄밀러 및 짐 바이트찰과 함께 알래스카에서 ''Toretocnemus''로 분류한 두 표본을 기술했다. 이 중 하나인 UAMES 3599는 그라비나 섬 해안의 네헨타 형성에서 나왔다. 이 표본은 1969년에 발견되어 2004년에 발굴되었다. 다른 표본인 UAMES 34994는 하운드 섬의 하운드 아일랜드 화산암 형성에서 발견되었다.[8]
참조
[1]
웹사이트
†Toretocnemus Merriam 1903 (ichthyosaur)
https://paleobiodb.o[...]
Fossilworks
2021-12-17
[2]
논문
New material of ''Qianichtyosaurus'' Li, 1999 (Reptilia, Ichthyosauria) from the Late Triassic of southern China, and implications for the distribution of Triassic ichthyosaurs
2003
[3]
논문
Triassic Ichthyopterygia from California and Nevada
https://www.biodiver[...]
[4]
웹사이트
Genus: ''Toretocnemus'' MERRIAM, 1902
http://www.paleofile[...]
2022-05-17
[5]
논문
New ichthyosaurs from the Upper Triassic of California
https://www.biodiver[...]
[6]
논문
A remarkable ichthyosaurian right anterior paddle
https://www.biodiver[...]
[7]
논문
''Toretocnemus'', a Late Triassic ichthyosaur from California, U.S.A. and Sonora, Mexico
https://www.research[...]
2002-01-01
[8]
간행물
Paleobiology of ichthyosaurs: Using osteohistology to test hypotheses of growth rates and metabolism in a clade of secondarily aquatic marine tetrapods
https://scholarworks[...]
University of Alaska
[9]
논문
Ichthyosauria: Their diversity, distribution, and phylogeny
https://www.research[...]
[10]
논문
Swimming Capabilities of Mesozoic Marine Reptiles: Implications for Method of Predation
[11]
논문
A review of ichthyosaur (Reptilia, Ichthyopterygia) soft tissues with implications for life reconstructions
[12]
논문
New findings reveal that the Middle Triassic ichthyosaur ''Mixosaurus cornalianus'' is the oldest amniote with a dorsal fin
[13]
논문
The Ichthyosauria
https://www.biodiver[...]
[14]
논문
Evolutionary trends in longipinnate ichthyosaurs, with particular reference to the skull and fore fin
https://www.biodiver[...]
[15]
논문
Triassic Ichthyosauria: With special reference to the American forms
https://www.biodiver[...]
[16]
논문
Phylogeny of the Ichthyopterygia incorporating recent discoveries from South China
[17]
논문
Phylogeny of the Ichthyopterygia
http://mygeologypage[...]
[18]
논문
Lines of phyletic and biological development of the Ichthyopterygia
https://www.biodiver[...]
[19]
논문
The distinction between longipinnate and latipinnate ichthyosaurs
https://archive.org/[...]
[20]
논문
A new phylogeny of ichthyosaurs (Reptilia: Diapsida)
https://research-inf[...]
[21]
논문
Cranial anatomy of ''Besanosaurus leptorhynchus'' Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications
[22]
논문
Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans
https://faculty.umb.[...]
[23]
논문
Osteohistology of the Early Triassic ichthyopterygian reptile ''Utatsusaurus hataii'': Implications for early ichthyosaur biology
[24]
논문
The affinities and ecology of Triassic ichthyosaurs
[25]
논문
The skull and endocranium of a Lower Jurassic ichthyosaur based on digital reconstructions
https://www.research[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com