브라키오사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
브라키오사우루스는 최대 길이 25m, 키 15m, 무게 45톤에 달하는 대형 용각류 공룡으로, 긴 목과 키가 큰 외형으로 널리 알려져 있다. 어린 개체는 육식 공룡에게 잡아먹히기도 했지만, 성체는 천적이 없었을 것으로 추정된다. 브라키오사우루스는 작은 머리, 긴 목, 큰 몸통, 긴 꼬리, 기둥 모양의 사지를 가진 네 발 동물이었으며, 앞다리가 뒷다리보다 길어 기린과 유사한 체형을 가졌다. 북아메리카 서부의 모리슨 층에서 발견되며, 다양한 초식 공룡 및 육식 공룡과 공존했다. 브라키오사우루스의 목 자세, 식성, 콧구멍 위치 등 다양한 고생물학적 특징에 대한 논쟁이 있으며, 대중문화에서도 상징적인 공룡으로 묘사된다.
더 읽어볼만한 페이지
- 브라키오사우루스과 - 아비도사우루스
아비도사우루스는 신용각류 공룡의 한 속이며, 브라키오사우루스와 티타노사우루스상과에 속한다. - 1903년 기재된 화석 분류군 - 오르니톨레스테스
오르니톨레스테스는 쥐라기 후기 북아메리카에 살았던 "새 도둑"이라는 뜻의 소형 수각류 공룡으로, 날렵한 몸집과 날카로운 이빨로 작은 동물을 사냥했을 것으로 추정되며 헨리 페어필드 오스본에 의해 명명된 이후 깃털 존재 여부와 식성 등에 대한 다양한 가설이 제시되었다. - 1903년 기재된 화석 분류군 - 토레토크네무스
토레토크네무스는 트라이아스기 후기에 북아메리카 북동부 태평양에 살았던 어룡의 한 속으로, 캘리포니아에서 처음 발견되었으며 최대 2m까지 자라는 큰 눈과 유선형 몸체를 가진 진어룡류이다. - 모리슨층의 공룡 - 마르쇼사우루스
마르쇼사우루스는 1976년 미국 콜로라도주에서 발견된 몸길이 약 2.5미터의 작은 수각류 공룡으로, 가볍고 민첩한 골격과 모리슨 층에서의 발견, 그리고 장골 변형 등의 특징을 통해 행동 및 생태 연구가 진행되었으며, 초기에는 메갈로사우루스류, 이후에는 피아트니츠키사우루스과와의 관계 연구가 이루어졌다. - 모리슨층의 공룡 - 드리오사우루스
쥐라기 후기 북아메리카에 서식한 드리오사우루스는 긴 목과 다리, 각질 부리를 가진 최대 3m 크기의 조각류 공룡으로, 모리슨 층 등 다양한 지역에서 화석이 발견되었으며, 1894년에 'Dryosaurus altus'로 재분류되었다.
| 브라키오사우루스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Brachiosaurus altithorax |
| 명명자 | 엘머 S. 릭스, 1903년 |
| 어원 | '팔 도마뱀' |
| 시대 | 쥐라기 후기 (킴메리지절 ~ 티톤절) |
| 생존 시기 | 약 1억 5,400만 년 전 ~ 1억 5,000만 년 전 |
| 발견 장소 | 미국 콜로라도주 |
| 몸길이 | 18 ~ 22m |
| 무게 | 28.3 ~ 46.9 톤 |
| 키 | 9m |
| 생물학적 분류 | |
| 상목 | 공룡상목 |
| 목 | 용반목 |
| 소목 (계통군) | 마크로나리아 |
| 하목 (계통군) | 티타노사우루스형류 |
| 과 | 브라키오사우루스과 |
| 속 | 브라키오사우루스속 |
| 종 | |
| 타입 종 | Brachiosaurus altithorax |
2. 특징
Brachiosaurus영어는 최대 25m, 무게 45ton,[106] 키 15m로, 용각류 중에서도 큰 편에 속한다. 몸무게는 과거 80ton 정도로 추정되었으나, 기낭이 있었다는 연구 이후 추정치가 줄었다. 키가 크고 목이 긴 모습 때문에 세계적으로 가장 널리 알려진 공룡 중 하나이다. 케라토사우루스와 알로사우루스와 공존했고, 어린 개체는 이들의 먹이가 되기도 했지만, 다 자란 성체는 천적이 없었을 것으로 추정된다. 한때 물속에서 살았다는 수생설이 주장되기도 하였으나, 신체 구조가 수생 생활에 적합하지 않음이 밝혀지면서 기각되었다.
2. 1. 신체 구조

최대 길이 25m, 무게 45톤,[106] 키 15m로, 용각류 중에서도 큰 편에 속한다. 몸무게는 과거 80톤 정도로 추정되었으나, 기낭이 있었다는 연구 이후 추정치가 줄었다. 키가 크고 목이 긴 모습 때문에 세계적으로 가장 널리 알려진 공룡 중 하나이다. 육식공룡 케라토사우루스와 알로사우루스와 공존했고, 어린 개체는 이들의 먹이가 되기도 했지만, 다 자란 성체는 천적이 없었을 것으로 추정된다. 한때 물속에서 살았다는 수생설이 주장되기도 하였으나, 신체 구조가 수생생활에 적합하지 않음이 밝혀지면서 기각되었다.[104]
모든 용각류 공룡과 마찬가지로, ''브라키오사우루스''는 작은 두개골, 긴 목, 높은 타원체 단면을 가진 큰 몸통, 길고 근육질의 꼬리, 가늘고 기둥 모양의 사지를 가진 네 발로 걷는 동물이었다.[50] 허파꽈리와 연결된 큰 기낭이 목과 몸통에 존재하여, 뼈 흡수를 통해 척추와 갈비뼈를 침범하여 몸 전체의 밀도를 크게 감소시켰다.[51][52] 목은 모식 표본에서 보존되지 않았지만, 밀접하게 관련된 ''지라파티탄''에서 용각류 기준에서도 매우 길었으며, 13개의 길쭉한 경추 (목) 척추로 구성되어 있었다.[53] 목은 약간의 S자형 곡선을 이루며, 아래쪽과 위쪽 부분은 구부러지고 가운데 부분은 곧게 뻗어 있었다.[54] ''브라키오사우루스''는 아마도 ''지라파티탄''과 매우 길쭉한 경늑골 (목 갈비뼈)을 공유했을 것이며, 이는 목의 아랫부분을 따라 뻗어 나가 여러 개의 이전 척추를 겹치게 했다. 이 뼈 막대들은 목 근육의 끝에 부착되어 이 근육이 몸통에 더 가깝게 위치하면서 목의 원위 부분을 작동시켜 원위 목 부분을 가볍게 만들었다.[54][55]
''브라키오사우루스''와 ''지라파티탄''은 아마도 세 번째와 다섯 번째 흉추 (등) 척추 사이에 작은 어깨 혹을 가지고 있었을 것이며, 여기에서 옆으로 향하고 위로 향하는 척추 돌기가 더 길어져 목 근육 부착을 위한 추가적인 표면을 제공했다.[56] 갈비뼈는 다른 용각류에 비해 깊었다.[57] 비록 상완골 (위팔뼈)과 넙적다리뼈 (넓적다리뼈)가 길이가 거의 같았지만, 길쭉한 앞팔과 다른 브라키오사우루스과의 중수골을 통해 추론할 수 있듯이 전체 앞다리가 뒷다리보다 길었을 것이다. 이로 인해 어깨가 엉덩이보다 훨씬 높은 기울어진 몸통이 생겼고, 목은 몸통에서 가파른 각도로 나왔다. ''브라키오사우루스''의 전반적인 모습은 다른 어떤 살아있는 동물보다 기린과 더 유사하다.[58] 반면에, 대부분의 다른 용각류는 뒷다리보다 짧은 앞다리를 가지고 있었고, 앞다리는 동시대의 디플로도쿠스류에서 특히 짧다.[59]
''브라키오사우루스''는 몸의 비율에서 밀접하게 관련된 ''지라파티탄''과 달랐다. 몸통은 약 25~30% 더 길어서 상완골보다 긴 등 척추를 갖게 되었다. 단 하나의 완전한 미추 (꼬리) 척추만 발견되었지만, 그것의 큰 높이는 꼬리가 ''지라파티탄''보다 더 컸음을 시사한다. 이 척추는 넓어진 신경 가시로 인해 인대 부착을 위한 면적이 훨씬 넓었으며, 이는 꼬리가 ''지라파티탄''보다 20~25% 더 길었을 가능성을 나타낸다.[40] 1988년, 고생물학자 그레고리 S. 폴은 ''브라키오사우루스''의 목이 ''지라파티탄''의 목보다 짧다고 제안했지만, 2009년, 고생물학자 마이크 P. 테일러는 ''브라키오사우루스''에 속할 가능성이 있는 두 개의 경추가 동일한 비율을 가지고 있다고 지적했다.[40][58] 수직으로 지향된 앞다리를 가진 ''지라파티탄'' 및 다른 용각류와 달리, ''브라키오사우루스''의 팔은 견갑골 관절 표면의 옆으로 향하는 방향으로 나타나 어깨 관절에서 약간 벌어진 것으로 보인다. 상완골은 ''지라파티탄''보다 덜 가늘었고, 넙적다리뼈는 비슷한 비율을 가졌다. 이것은 ''브라키오사우루스''의 앞다리가 ''지라파티탄''의 경우보다 더 많은 신체 무게를 지탱했음을 나타낼 수 있다.[40]
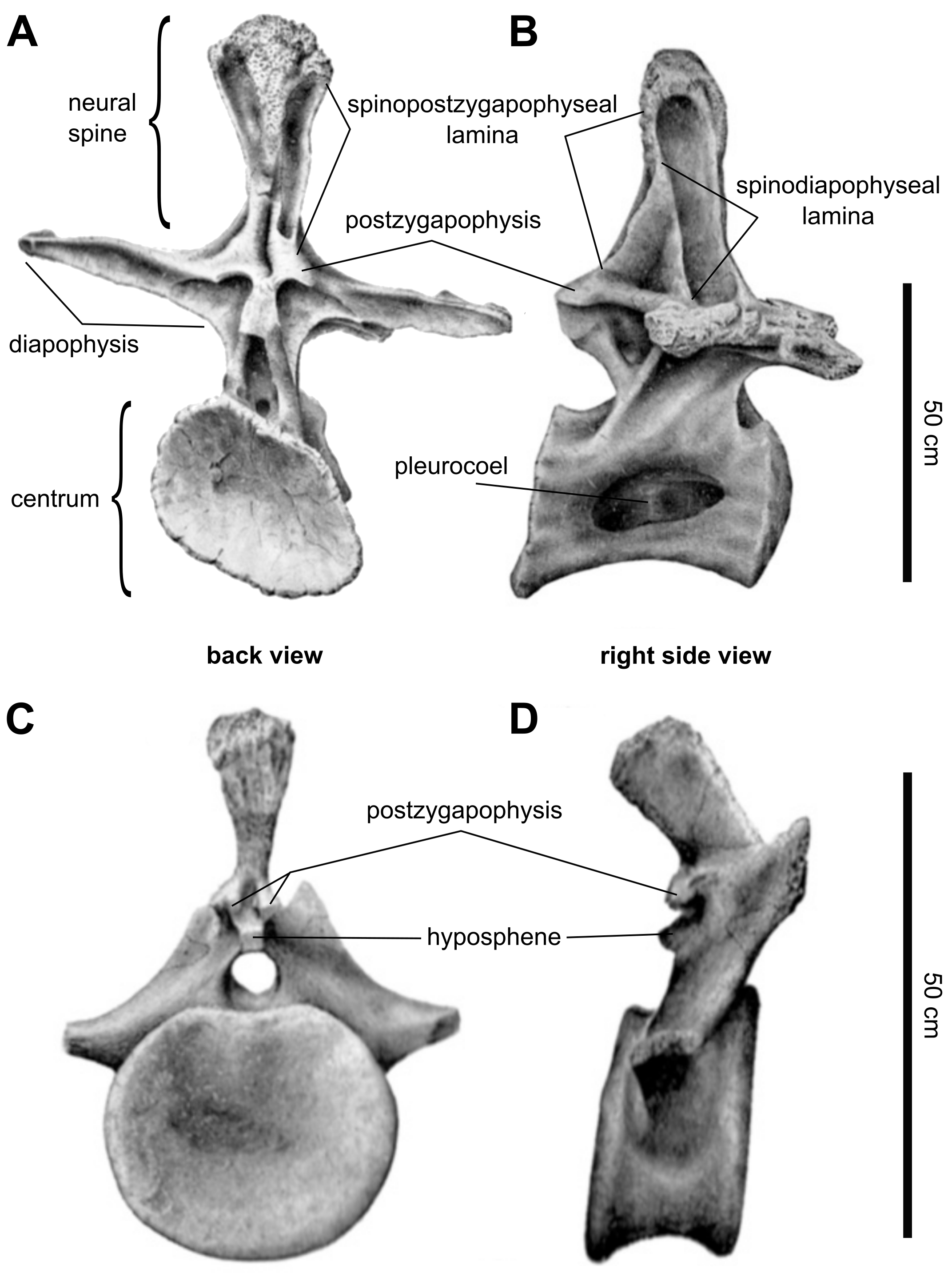
몸통 또는 흉부의 척추 기둥은 불완전하게 알려져 있지만, 브라키오사우루스의 등은 12개의 등뼈로 구성되었을 가능성이 가장 높다. 이는 명명되지 않은 브라키오사우루스과 표본, BMNH R5937에 보존된 완전한 등뼈 기둥에서 추론할 수 있다.[60] 등뼈 앞쪽 부분의 척추는 뒤쪽 부분의 척추보다 약간 더 컸지만 훨씬 더 길었다. 이는 앞부분의 척추가 훨씬 더 컸지만 약간 더 긴 ''지라파티탄''과는 대조적이다. 척추의 하부 부분인 중심(척추체)은 단면에서 더 길고 대략 원형이었지만, ''지라파티탄''의 중심은 높이보다 넓었다. 공기 주머니의 침입을 허용하는 중심 측면의 구멍은 ''지라파티탄''보다 더 컸다. 척추의 신경궁에서 옆으로 뻗어 나가는 큰 돌기인 횡돌기는 수평이었지만, ''지라파티탄''의 횡돌기는 위쪽으로 기울어져 있었다. 끝부분에서 이 돌기는 갈비뼈와 연결되었는데, 관절면은 ''지라파티탄''처럼 뚜렷한 삼각형이 아니었다. 옆면에서 위쪽으로 튀어나온 신경 가시는 수직으로 서 있었고, 밑부분이 위쪽보다 두 배 더 넓었다. ''지라파티탄''의 신경 가시는 뒤쪽으로 기울어져 있었고 밑부분이 넓어지지 않았다. 앞 또는 뒤에서 보면, 신경 가시는 위쪽으로 갈수록 넓어졌다.[40]
''브라키오사우루스''에서 이러한 넓어짐은 점진적으로 발생하여 노 모양을 만들었지만, ''지라파티탄''에서는 갑자기 발생하여 가장 윗부분에서만 나타났다. 앞면과 뒷면 모두에서, 신경 가시는 크고 삼각형이며 거친 표면을 특징으로 했는데, ''지라파티탄''에서는 반원형이고 훨씬 작았다. 다양한 척추 돌기는 얇은 뼈 시트 또는 융기(ridge)로 연결되어 있었으며, 이를 ''판(laminae)''이라고 한다. ''브라키오사우루스''에는 신경 가시의 뒷면을 따라 내려오는 ''지라파티탄''에 있는 척추 후방 판이 없었다. 신경 가시에서 횡돌기까지 뻗어 있는 척추 횡돌기 판은 신경 가시와 척추 뒤쪽의 관절 돌기 사이를 뻗어 있는 척추 후관절 돌기 판과 융합되어 신경 가시의 중간 높이에서 끝났다. ''지라파티탄''에서는 두 판이 융합되지 않았고, 척추 횡돌기 판은 신경 가시의 꼭대기까지 닿았습니다. ''브라키오사우루스''는 또한 후자 속에만 고유한 등뼈의 판에 있는 세 가지 세부 사항이 없다는 점에서 ''지라파티탄''과 구별된다.[40]
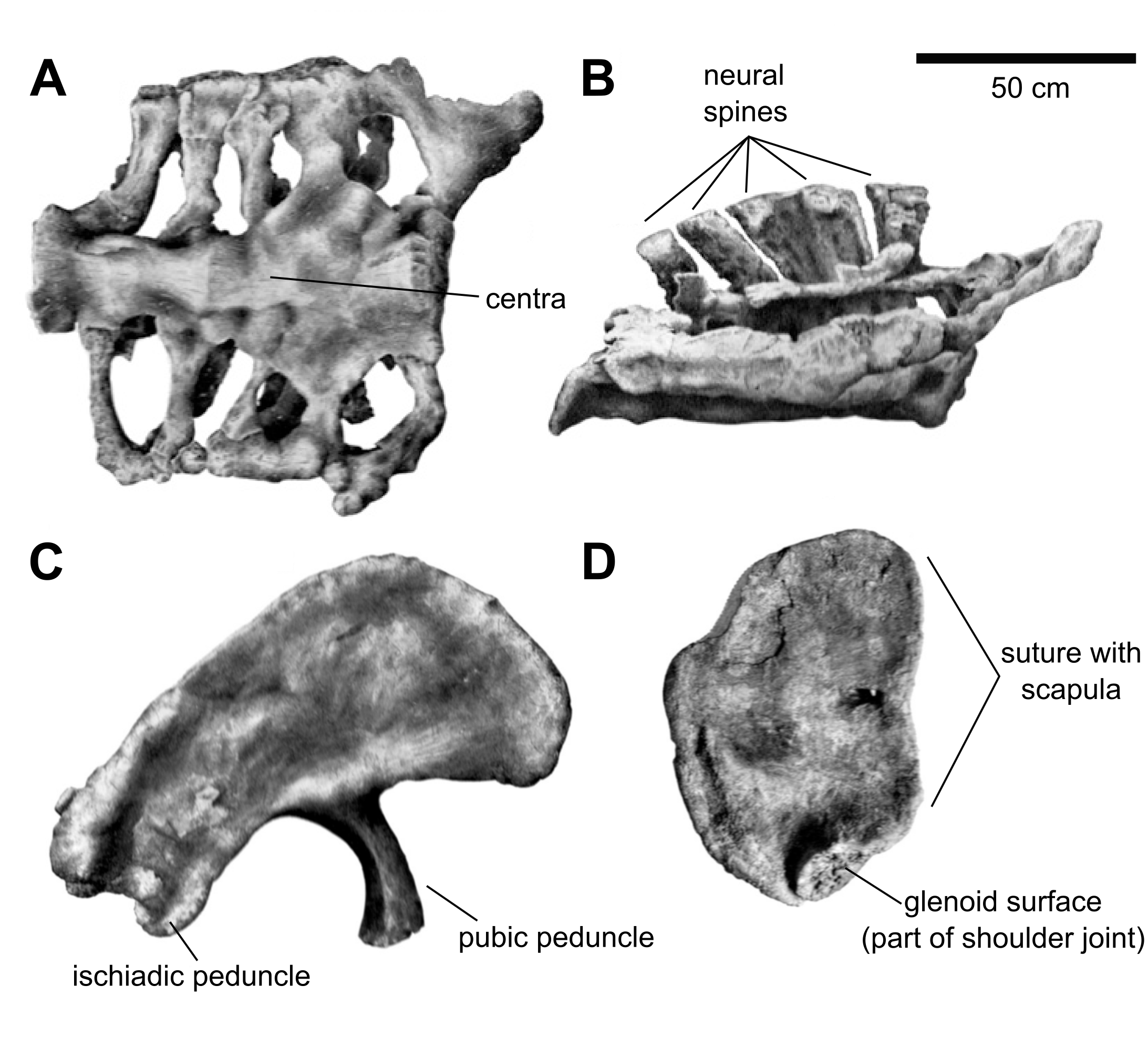
공기 주머니는 척추뿐만 아니라 갈비뼈에도 침투했다. ''브라키오사우루스''에서 공기 주머니는 갈비뼈 축의 앞쪽에 있는 작은 구멍을 통해 침투했지만, ''지라파티탄''에서는 척추의 횡돌기와 연결되는 뼈 돌기인 결절의 앞면과 뒷면에 모두 구멍이 있었다. 1988년 폴은 ''브라키오사우루스''의 갈비뼈가 ''지라파티탄''보다 더 길다고 주장했지만, 2009년 테일러는 이에 이의를 제기했다.[40] 등뼈 기둥 뒤에는, 엉치뼈가 5개의 공동 골화된 엉치뼈로 구성되어 있었다.[61] ''지라파티탄''과 마찬가지로 엉치뼈는 비례적으로 넓었고 매우 짧은 신경 가시를 특징으로 했다. ''지라파티탄''의 엉치뼈 재료가 제대로 보존되지 않아 두 속 간의 자세한 비교는 불가능하다. 꼬리에서는 두 번째 꼬리뼈만 잘 보존되어 있다.[40]
''지라파티탄''과 마찬가지로 이 척추는 약간 양쪽으로 오목했고(양쪽 끝이 오목함), 측면에 구멍이 없었으며 사각형이고 뒤쪽으로 기울어진 짧은 신경 가시를 가지고 있었다. ''지라파티탄''의 두 번째 꼬리뼈와는 대조적으로, ''브라키오사우루스''의 두 번째 꼬리뼈는 비례적으로 더 높은 신경궁을 가지고 있어 척추가 약 30% 더 높았다. 중심에는 ''지라파티탄''과는 달리 측면에 함몰부가 없었다. 앞 또는 뒤에서 보면, 신경 가시는 끝으로 갈수록 최소 너비의 약 세 배까지 넓어졌지만, ''지라파티탄''에서는 넓어짐이 나타나지 않는다. 신경 가시는 또한 ''지라파티탄''(20도)보다 더 많은 약 30도 기울어져 있었다. 꼬리 갈비뼈는 옆으로 튀어나왔고, ''지라파티탄''처럼 뒤로 기울어지지 않았습니다. 척추 뒤쪽의 관절 돌기의 관절면은 아래쪽으로 향했지만, ''지라파티탄''의 관절면은 측면을 더 향했습니다. 관절 돌기 외에도, 상하관절 결합은 척추 사이에 추가적인 관절을 형성하여 척추 기둥을 더 뻣뻣하게 만들었습니다. ''브라키오사우루스''에서 상관절은 ''지라파티탄''보다 훨씬 더 두드러졌습니다.[40]
오훼골은 반원형이고 폭보다 높았다. ''지라파티탄''과의 차이점은 옆면에서 형태와 관련이 있는데, 여기에는 어깨뼈와의 더 곧은 봉합선이 포함된다. 또한, 어깨 관절의 일부를 형성하는 관절면은 ''지라파티탄''과 다른 용각류보다 더 두껍고 측면으로 더 향해 있어 앞다리가 더 넓게 뻗어 있었을 가능성이 있다. 보존된 위팔뼈는 길이가 204cm이지만, 하단 일부는 침식으로 인해 손실되었다. 원래 길이는 216cm로 추정된다. 이 뼈는 ''브라키오사우루스''에서 다른 대부분의 용각류보다 더 가늘었으며, 가장 좁은 부분의 너비는 28.5cm에 불과했다. 그러나 ''지라파티탄''보다 더 튼튼하여 상단과 하단에서 약 10% 더 넓었다. 상단에는 옆면에서 보이는 낮은 융기가 있었는데, 이는 ''지라파티탄''에는 없었다.[40]
차별화되는 특징은 또한 골반의 엉덩이뼈에서도 찾을 수 있다. ''브라키오사우루스''에서, 좌골에 연결되는 아래쪽으로 뻗어 있는 연장부인 좌골 다리는 ''지라파티탄''보다 더 멀리 아래로 뻗어 있다. 후자의 속은 좌골 다리와 엉덩이뼈의 뒷부분 사이에 날카로운 노치가 있었지만, 이 노치는 ''브라키오사우루스''에서 더 둥글게 되어 있다. 엉덩이뼈 뒷부분의 윗면에는 ''브라키오사우루스''에 다른 용각류에는 없는 두드러진 결절이 있었다. 뒷다리의 경우, 넙다리뼈는 ''지라파티탄''의 넙다리뼈와 매우 유사했지만 약간 더 튼튼했으며, 길이는 203cm였다.[57] ''지라파티탄''과 마찬가지로, 단면에서 강하게 타원형이었으며, 앞 또는 뒤에서 보면 옆면보다 두 배 이상 넓었다.[40] 넙다리뼈의 축 뒤쪽에 있는 두드러진 돌기인 네 번째 돌기는 더 두드러졌고 더 아래쪽에 위치해 있었다. 이 돌기는 가장 중요한 운동 근육인 꼬리넙다리 근육의 고정점 역할을 했으며, 이는 꼬리에 위치하여 수축 시 위쪽 허벅지를 뒤로 당겼다. 넙다리뼈의 아래쪽 끝에서, 한 쌍의 관절 융기는 ''지라파티탄''만큼 뒤쪽으로 강하게 뻗어 있지 않았습니다. 두 개의 관절 융기는 ''브라키오사우루스''에서는 너비가 비슷했지만 ''지라파티탄''에서는 달랐다.[40]

카펜터와 티드웰이 재구성한 바에 따르면, 펠치 채석장으로 지정된 두개골은 머리 뒤쪽의 후두골 과에서 전상악골(위턱의 앞쪽 뼈) 앞쪽까지 약 81cm 길이로, 모리슨 층에서 발견된 가장 큰 용각류 두개골이었다.[20] 반면, D'Emic과 Carrano는 두개골의 길이를 70cm로 추정했고, ''지라파티탄''과 비례적으로 비슷하다면 높이는 약 55cm, 너비는 35cm로 추정했다.[22] 전반적으로 두개골은 ''지라파티탄''과 같이 높았으며, 콧구멍 사이의 비골 바 앞쪽의 주둥이는 길었다(카펜터와 티드웰에 따르면 두개골 길이의 약 36%). 위에서 보면 주둥이는 다소 무뎠으며(''지라파티탄''과 같음), 두개골의 나머지 부분과 각도를 이루고 있어서 아래쪽을 향하는 인상을 주었다.[20][22]
등쪽 및 측두개창 (두개골 위쪽 뒤쪽과 측면에 있는 구멍)은 컸는데, 이는 거대한 턱 부착 근육의 힘 때문일 수 있다. 두개골 상단의 전두골은 짧고 넓었으며(''지라파티탄''과 유사), 봉합선으로 융합되어 두정골과 연결되었으며, 두정골 또한 서로 융합되었다. 등쪽 개창 사이의 두정골 표면은 ''지라파티탄''보다 넓었지만 ''카마라사우루스''보다는 좁았다. 두개골은 전두골과 비골 사이의 U자형(W자형 대신) 봉합선에서 ''지라파티탄''과 달랐는데, 이 모양은 전두골이 안와 (눈구멍) 위로 앞으로 뻗어나오면서 더 뚜렷하게 나타난다. ''지라파티탄''과 마찬가지로 후두골 과의 목은 매우 길었다.[20][22]

전상악골은 ''카마라사우루스''보다 길어 보이며, 비골 바 쪽으로 더 완만하게 기울어져 매우 긴 주둥이를 만들었다. ''브라키오사우루스''는 길고 깊은 상악골 (위턱의 주요 뼈)을 가지고 있었는데, 치조 (이빨 소켓)이 있는 가장자리를 따라 두꺼웠고 위쪽으로 갈수록 얇아졌다. 상악골의 치간판은 얇고, 융합되어 있으며, 다공성이었고 삼각형이었다. 판 사이에는 삼각형의 영양 동맥이 있었으며, 각각 치아 맹출 중인 이빨의 끝부분이 포함되어 있었다. 뼈로 된 콧구멍 앞쪽의 비강 와 (우울증)는 길고 비교적 얕았으며, ''지라파티탄''보다 덜 발달했다. 이 와에는 하비강 개창이 있었는데, 이는 ''지라파티탄''과 ''카마라사우루스''보다 훨씬 컸다. 치아를 포함하는 아래턱의 뼈인 치골은 튼튼했지만 ''카마라사우루스''보다는 덜했다. 치골의 위쪽 가장자리는 옆에서 보면 아치형이었지만, ''카마라사우루스''만큼 심하지는 않았다. 치골의 치간판은 다소 타원형이었으며, 그 사이에 다이아몬드 모양의 구멍이 있었다. 치골에는 아홉 번째 치조 아래까지 열려 있는 메켈 홈이 있었으며, 그 이후에는 얕은 홈으로 이어졌다.[20][22]
각 상악골에는 14개 또는 15개의 이빨이 들어갈 공간이 있었지만, ''지라파티탄''은 11개, ''카마라사우루스''는 8~10개의 이빨을 가지고 있었다. 상악골에는 ''카마라사우루스''와 유사하게 거친 치아 에나멜을 가진 치아 대체가 있었지만, 가장자리에는 작은 치아 세선 (톱니)이 없었다. 상악골은 ''카마라사우루스''보다 넓었기 때문에 ''브라키오사우루스''는 더 큰 이빨을 가졌을 것이다. 전상악골의 대체 이빨은 주름진 에나멜을 가지고 있었으며, 이 이빨 중 가장 완전한 것은 치아 세선이 없었다. 그것은 다소 주걱 모양이었고, 세로 능선이 있었다. 각 치골에는 약 14개의 이빨이 들어갈 공간이 있었다. ''브라키오사우루스''와 ''지라파티탄''의 상악 치열은 안와전개창 (안와 앞쪽의 구멍) 앞에서 끝났지만, ''카마라사우루스''와 ''슈노사우루스''에서는 개창 바로 앞과 아래에서 끝났다.[20][22]

호흡계와 같이 용각류는 조류처럼 기낭을 사용했다. 포유류처럼 양방향 기류는 없었는데, 포유류는 허파가 풀무처럼 기능하여 먼저 공기를 들이마시고 내쉬는 방식이다. 대신 공기는 기관에서 배에 있는 복부 기낭으로 빨려 들어간 다음, 뻣뻣한 허파의 곁가지인 공기 고리를 통해 앞으로 펌핑되었다. 밸브는 복부 기낭이 다시 채워질 때 공기가 뒤로 흐르는 것을 막았다. 동시에 목 기저부의 경추 기낭은 허파에서 사용된 공기를 빨아냈다. 두 기낭은 동시에 수축하여 사용된 공기를 기관에서 밖으로 펌핑했다. 이 절차는 단방향 기류를 보장했으며, 공기는 허파 자체에서 항상 단일 전방 방향으로 이동했다. 이것은 산소 섭취와 이산화 탄소 배출을 크게 향상시켰다. 죽은 공기가 빠르게 제거되었을 뿐만 아니라 허파의 혈류도 기류와 반대 방향으로 흘러 훨씬 더 효과적인 가스 교환을 이끌었다.
용각류에서 기낭은 단순히 호흡을 돕는 역할만 한 것이 아니었다. 공기 채널을 통해 골격의 많은 부분과 연결되었다. 이러한 가지, 즉 ''돌기''는 공압 개구를 통해 많은 뼈에 침투하여 강력하게 속을 비게 했다. 이 현상의 진화적 이점이 무엇인지는 완전히 밝혀지지 않았지만, 어쨌든 골격을 상당히 가볍게 했다. 또한 체온 조절을 돕기 위해 과도한 열을 제거했을 수도 있다.
2016년, 마크 할렛(Mark Hallett)과 매튜 웨델(Mathew Wedel)은 처음으로 ''B. altithorax''를 이러한 구조가 어떻게 형성되었을 수 있는지에 대한 예시로 사용하여 용각류의 전체 기낭 시스템을 재구성했다. 그들의 재구성에서 큰 복부 기낭은 골반과 외부 허파 측면 사이에 위치했다. 조류와 마찬가지로 세 개의 작은 낭이 가슴강의 아래쪽에서 펌핑 과정을 도왔는데, 뒤쪽에는 후흉 기낭, 중간에는 전흉 기낭, 앞쪽에는 쇄골 기낭이 있으며, 크기가 점차 감소했다. 경추 기낭은 앞쪽 허파 위에 어깨뼈 아래에 위치했다. 기낭은 관을 통해 척추뼈와 연결되었다. 돌기는 척추뼈 벽에 움푹 들어간 부분을 형성하는 다양한 와와 측면공동을 채웠다. 이들은 다시 뼈 내부의 유연하지 않은 기포와 연결되었다.
2. 2. 생태
최대 25m, 무게 45ton,[106] 키 15m로, 용각류 중에서도 큰 편에 속한다. 몸무게는 과거 80ton 정도로 추정되었으나, 기낭이 있었다는 연구 이후 추정치가 줄었다. 키가 크고 목이 긴 모습 때문에 세계적으로 가장 널리 알려진 공룡 중 하나이다. 육식공룡 케라토사우루스와 알로사우루스와 공존했고, 어린 개체는 이들의 먹이가 되기도 했지만, 다 자란 성체는 천적이 없었을 것으로 추정된다. 한때 물속에서 살았다는 수생설이 주장되기도 하였으나, 신체 구조가 수생 생활에 적합하지 않음이 밝혀지면서 기각되었다.
''브라키오사우루스''는 북아메리카 서부의 모리슨 층에서만 발견된다(아프리카 종이 재분류된 이후).[40] 모리슨 층은 뚜렷한 우기와 건기가 있는 반건조 환경으로 해석되며,[85][86] 평평한 범람원이 존재했다.[85] 모리슨 층에는 ''아파토사우루스'', ''바로사우루스'', ''카마라사우루스'', ''디플로도쿠스'', ''할로칸토사우루스'', ''수퍼사우루스'' 등 여러 다른 용각류 속이 존재했으며, 신체 비율과 먹이 적응이 달랐다.[41][87][88] ''브라키오사우루스''는 모리슨 층 용각류 중에서 덜 흔한 종이었다. 2003년 200곳이 넘는 화석 산지를 조사한 존 포스터는 이 속의 표본 12개를 보고했는데, 이는 ''바로사우루스''(13) 및 ''할로칸토사우루스''(12)와 비슷하지만, ''아파토사우루스''(112), ''카마라사우루스''(179) 및 ''디플로도쿠스''(98)보다는 훨씬 적은 수치이다.[41] ''브라키오사우루스'' 화석은 광대한 모리슨 층의 하부-중간 부분(지층 구역 2–4)에서만 발견되며, 약 1억 5400만 년에서 1억 5300만 년 전으로 거슬러 올라간다.[89] 반면 다른 많은 종류의 용각류는 층 전체에서 발견되었다.[41] 와이오밍에서 보고된 큰 발(북아메리카에서 브라키오사우루스과의 가장 북쪽 출현지)이 만약 ''브라키오사우루스''의 것이라면, 이 속은 넓은 위도 범위를 차지했을 것이다. 브라키오사우루스과는 넓은 왕관 모양의 이빨로 질긴 식물을 처리할 수 있었고, 따라서 디플로도쿠스과보다 더 넓은 범위의 식생 지역을 차지했을 수 있다. 브라키오사우루스과와 이빨 형태가 유사한 카마라사우루스과 역시 널리 퍼져 있었으며 계절에 따라 이동한 것으로 알려져 있으므로, 이는 브라키오사우루스과에도 해당될 수 있다.[27]
모리슨 층에서 알려진 다른 공룡으로는 육식성 수각류 ''코파리온'', ''스토케소사우루스'', ''오르니톨레스테스'', ''케라토사우루스'', ''알로사우루스'', ''토르보사우루스'', ''사우로파가낙스''와 초식성 조반류 ''캄프토사우루스'', ''드리오사우루스'', ''오트니엘리아'', ''가르고일레오사우루스'', ''스테고사우루스''가 있다.[90] ''알로사우루스''는 수각류 표본의 70~75%를 차지했으며 모리슨 먹이 사슬의 최상위 영양 단계에 있었다.[91] ''케라토사우루스''는 ''브라키오사우루스''의 더 작은 개체를 포함한 큰 용각류를 공격하는 데 특화되었을 수 있다. 이 고생물 환경을 공유한 다른 척추동물로는 조기어류, 개구리, 도롱뇽, ''도르세토켈리스''와 같은 거북, 이빨도마뱀, 도마뱀, ''호플로수쿠스''와 같은 육상 및 수생 악어형류, ''하르팍토그나투스''와 ''메사닥틸루스''와 같은 여러 종의 익룡이 있다. 이매패류와 수생 달팽이 껍질도 흔하다. 이 시대의 식물상은 녹조류, 곰팡이, 이끼, 속새, 소철, 은행나무, 그리고 여러 과의 구과식물 화석으로 밝혀졌다. 식생은 고비고사리와 고사리가 있는 강변 숲에서 사바나에 이르기까지 다양했으며, 브라키필룸과 같은 ''아라우카리아''와 유사한 구과식물과 관목이 있었다.[92]
체장(전장)은 약 25m, 체고(두정고)는 약 16m에 달하며, 최근까지 가장 키가 큰 공룡으로 여겨졌다. 과거에는 체중이 80ton 이상으로 가장 무거운 공룡이며, 그 체중을 지탱하기 위해 연못이나 호수 등 물 속에서 살았다고 추측되기도 했지만, 횡격막을 가지고 있지 않아 수압으로 폐가 짓눌려 호흡을 할 수 없다는 것이 밝혀져 이 설은 부정되었다. 용각류 등의 대형 공룡에게는 조류가 갖는 것과 마찬가지로 기낭에 의해 몸을 가볍게 하는 구조가 있었다고 생각되며, 현재는 23ton 정도[104]라는 설도 있으며, 크게 잡아도 50ton 정도라고 추정된다.
과거에는 두개골의 모양에서 현생 코끼리처럼 근육 조직으로 만들어진 코가 있었을 가능성이 지적되었다. 브라키오사우루스는 용각류의 다른 종과는 달리 앞다리가 뒷다리보다 길고, 어깨가 뒤쪽으로 기울어져 있어서 더 높은 곳의 식물을 먹을 수 있었다. 최근 연구에서는 목을 수직으로 뻗으면 혈액 순환에 문제가 생겨 기린처럼 수직에 가까운 각도로 목을 들어 올릴 수 없었으며, 어깨의 기울기를 연장한 선상, 즉 비스듬한 정도의 앞쪽 각도였을 것이라는 견해도 있다. 이러한 비율 때문에 영화 《쥬라기 공원》에서처럼 뒷다리와 꼬리만으로 일어섰을 가능성은 없다고 여겨진다. 머리 부분도 특징적인데, 부풀어 오른 콧구멍을 가지고 있으며, 머리 가장 높은 곳에 콧구멍이 있다(이 콧구멍 위치는 과거 수중 생활설의 근거 중 하나로 여겨지기도 했다). 머리는 작고 뇌의 크기도 200g 정도이며, 씹는 힘이 그다지 강하지 않았기 때문에 나뭇가지나 잎을 통째로 삼켜서 먹었던 것으로 보인다. 사회성이 있어서 무리 생활을 했던 것으로 생각되며, 특히 밤이나 이동 시에는 어린 개체가 중심이 되어 어른 개체가 이를 둘러싸서 보호했다고 여겨진다.
알로사우루스나 케라토사우루스 등 다른 많은 공룡들과 공존했다.[105]
3. 발견
1900년 미국 콜로라도주 그랜드 리버 계곡 모리슨 지층에서 화석이 처음 발견되었다.[2] 그러나 이 화석은 불완전한 몇 가지 골격이나 뼛조각에 지나지 않았다. 제1차 세계 대전 당시 동아프리카 탄자니아에서 독일 과학자들에 의해 완전한 골격이 발견되었으나, 이후 탄자니아에서 발견된 종류는 기라파티탄이라는 별개의 종으로 밝혀졌다. 대중에게 알려진 브라키오사우루스의 모습은 코뼈가 좀 더 높은 기라파티탄의 모습이다. 1950년대 포르투갈에서도 이 공룡과 비슷한 화석이 발견되었지만 2003년 루소티탄이라는 별개의 속으로 재동정되었다.

브라키오사우루스(Brachiosaurus) 속은 1900년 콜로라도강 계곡의 콜로라도주 프루이타 근처에서 발견된 부분적인 사지 골격을 기반으로 한다.[2] 이 표본은 홀로타입으로 선언되었으며, 모리슨 층의 브러시 베이슨 층에서 발견되어 후기 킴메리지안 시대(약 1억 5400만 년에서 1억 5300만 년 전)의 것이다.[3] 미국의 고생물학자 엘머 S. 리그스(Elmer S. Riggs)와 그의 동료들이 시카고의 필드 자연사 박물관(Field Columbian Museum, 현재의 필드 자연사 박물관)에서 발견했으며,[57] 현재 FMNHP25107로 분류되어 있다.[40]
리그스와 동료들은 콜로라도주 그랜드 정션의 치과의사 스탠튼 메릴 브래드버리와의 서신 교환 결과로 이 지역에서 활동했다. 1899년 봄, 리그스는 서부 콜로라도주의 시장들에게 편지를 보내 철도 종착역에서 유타주 북동부로 이어지는 경로에 대해 문의했는데, 그곳에서 시신세 포유류 화석을 찾기를 희망했다. 그는 아마추어 수집가이자 웨스턴 콜로라도 과학 아카데미의 회장이었던 브래드버리로부터 1885년부터 그랜드 정션 근처에서 공룡 뼈가 수집되었다는 소식을 들었다.[2] 리그스는 회의적이었지만, 그의 상사이자 지질학 큐레이터인 올리버 커밍스 패링턴은 큰 용각류 골격을 수집품에 추가하기를 열망했고, 박물관 경영진에게 500달러를 원정대에 투자하도록 설득했다. 1900년 6월 20일에 도착하여 버려진 고트 랜치에 야영지를 세웠다. 리그스의 현장 조수 해럴드 윌리엄 멘케는 7월 4일에 FMNHP25107의 상완골을 발견하고,[57][4] "지금까지 가장 큰 것!"이라고 외쳤다. 리그스는 처음에는 심하게 보존된 ''브론토사우루스''(Brontosaurus) 표본으로 여겼고, ''모로사우루스''(Morosaurus) 골격 발굴을 우선시했다. 7월 26일, 채석장 13번에서 상완골로 돌아갔는데, 그 크기가 엄청나 리그스는 가장 큰 육상 동물을 발견했음을 확신했다.
리그스 채석장 13번은 리그스 언덕으로 알려진 작은 언덕에 있으며, 오늘날 기념패가 세워져 있다. 리그스 언덕에서 더 많은 브라키오사우루스 화석이 보고되었지만, 다른 화석 발견물은 훼손되었다.[4][5] 표본 발굴 동안, 리그스는 상완골의 길이 때문에 굴곡된 대퇴골로 오인했으며, 같은 골격의 실제 대퇴골이 발견되었을 때 오류가 확인되었다. 1904년 리그스는 "갈비뼈의 특이한 크기가 아니었다면, 이 표본은 아파토사우루스(Apatosaur)로 간주되어 가치가 없을 정도로 제대로 보존되지 않았을 것이다."라고 언급했다. 실험실에서 화석 재료를 준비한 후에야 뼈가 상완골로 인식되었다.[61] 발굴은 많은 방문객을 끌어들여 작업을 지연시키고 멘케에게 뼈를 지키도록 했다. 8월 17일, 마지막 뼈가 석고로 싸여졌다. 10일간의 정찰 여행 후, 원정대는 그랜드 정션으로 돌아와 모든 화석을 철도역으로 운송하기 위해 팀과 마차를 고용했다. 38개의 상자에 짐을 싸는 데 한 주가 걸렸으며, 무게는 였다. 9월 10일, 리그스는 시카고로 기차를 타고 출발하여 15일에 도착했다. 철도 회사는 승객과 화물을 무료로 운송했는데, 이는 홍보 제스처였다.
홀로타입 골격은 오른쪽 상완골(위팔뼈), 오른쪽 대퇴골(넙다리뼈), 오른쪽 장골(엉덩이뼈), 오른쪽 오훼돌기(어깨뼈), 엉치뼈(엉덩이의 융합된 척추), 마지막 7개의 흉추(몸통)와 꼬리뼈 2개, 여러 개의 갈비뼈로 구성된다.[57][40][6] 리그스는 오훼돌기를 왼쪽에서 나온 것으로 묘사했지만,[57][61][6] 재연구 결과 오른쪽 오훼돌기임이 밝혀졌다.[40] 발견 당시, 상완골의 아래쪽 끝, 엉치뼈의 아래쪽, 장골 및 보존된 꼬리 척추는 공기에 노출되어 날씨에 의해 부분적으로 손상되었다. 척추는 원래 위치에서 약간만 벗어났고, 위쪽 면이 아래쪽을 향하도록 발견되었다. 갈비뼈, 상완골, 오훼돌기는 척추의 왼쪽으로 이동했는데, 이는 물의 흐름에 의해 운반되었음을 나타낸다. ''디플로도쿠스''(Diplodocus)의 고립된 장골과 주변 암석의 조성 변화는 척추에 부딪힌 것으로 보인다. 표본은 미세한 점토에 묻혀 낮은 에너지 조건을 나타내는 반면, 조약돌과 사암으로 구성된 거친 퇴적물 층으로 7번째 전(前)엉치 척추에서 잘려 퇴적되었는데, 이는 더 강한 물의 흐름에서 퇴적되었음을 나타낸다. 리그스는 1904년에 골격의 앞부분이 물에 휩쓸려 갔고 뒷부분은 이미 퇴적물에 덮여 보존되었다고 제안했다.
리그스는 1901년에 새로운 발견에 대한 보고서를 발표했지만, 새로운 공룡의 이름을 발표하지 않았다.[6] 1903년, 모식종 ''브라키오사우루스 알티토락스''(Brachiosaurus altithorax)를 명명했다.[57] 속명은 "팔"을 의미하는 고대 그리스어 ''브라키온''/βραχίων과 "도마뱀"을 의미하는 ''사우로스''/σαυροςgrc에서 유래했다.[57] 종명은 라틴어 ''알투스''(altus, "깊은")와 그리스어 ''토락스''/θώραξ("흉갑, 갑옷, 코르셋")에서 유래되었으며, 가슴 부분이 깊고 넓기 때문에 선택되었다.[7] 라틴어 ''토락스''는 그리스어에서 유래되었으며 몸의 가슴에 대한 과학적 명칭이 되었다. 리그스의 1901년 및 1903년 논문 제목은 이 표본이 "가장 큰 공룡"임을 강조했다.[57][6] 리그스는 1903년 출판물을 1904년 단행본에 더 자세한 설명과 함께 따랐다.[61]
홀로타입 준비는 필드 박물관을 위해 리그스가 수집한 직후인 1900년 가을에 시작되었다. 먼저 사지 요소를 처리했다. 1904년 겨울, 등과 엉덩이의 척추는 제임스 B. 애보트와 C.T. 클라인에 의해 준비되었다. 각 뼈의 준비가 끝나면, 필드 박물관의 첫 위치인 세계 컬럼비아 박람회 미술관 35관의 유리 케이스에 전시되었다. 모든 뼈는 ''아파토사우루스''(Apatosaurus)가 공개된 1908년까지 35관에 단독으로 전시되었으며, 이는 채석장 12번에서 리그스가 발견한 표본으로, FMNH P25112로 분류되고 ''브론토사우루스''(Brontosaurus) 표본으로 확인되었다. 골격의 20%만 회수되었기 때문에 브라키오사우루스의 마운트는 시도되지 않았다. 1993년, 홀로타입 뼈가 틀에 부어져 주조되었고, 누락된 뼈는 베를린 자연사 박물관의 ''브라키오사우루스 브란카이''(Brachiosaurus brancai, 현재는 ''지라파티탄''(Giraffatitan))의 재료를 기반으로 조각되었다. 이 플라스틱 골격은 1994년에 필드 박물관 스탠리 필드 홀 북쪽 끝에 전시되었다. 홀로타입의 실제 뼈는 마운트된 캐스트 양쪽의 유리 케이스에 전시되었다. 1999년, ''티라노사우루스''(Tyrannosaurus) 골격 "수"를 위해 오헤어 국제공항 유나이티드 항공 터미널 1의 B Concourse로 옮겨졌다. 동시에, 필드 박물관은 골격의 두 번째 플라스틱 캐스트를 마운트하여 박물관 외부 NW 테라스에 전시했다.
1988년 그레고리 S. 폴은 ''B. brancai'' 골격의 새로운 재구성을 발표, ''B. altithorax''와의 비율 차이를 강조했다. 척추의 변이 방식이 가장 큰 차이점이었다. 아프리카 자료는 척추 길이가 균일하지만, ''B. altithorax''는 길이가 크게 달랐다. 폴은 두 종의 사지 및 골반 요소가 유사하여 아속 수준에서만 구분해야 한다고 제안했다. 즉, ''Brachiosaurus (Brachiosaurus) altithorax''와 ''Brachiosaurus (Giraffatitan) brancai''로 구분해야 한다는 것이다. 1991년 조지 올셰프스키는 척추 변이를 언급하며 ''Giraffatitan''을 완전한 속 수준으로 격상시켰다. 1991년에서 2009년 사이 ''Giraffatitan''은 다른 연구자들에 의해 거의 무시되었다.
2009년 테일러의 연구 결과, ''B. altithorax''와 텐다구루 자료 사이에 상당한 차이가 발견되었다. 26개의 골학적 특징을 발견했는데, 이는 ''디플로도쿠스''와 ''바로사우루스'' 사이의 차이보다 컸으며, 아프리카 자료를 ''Giraffatitan brancai''로, 자체 속(''Giraffatitan'')에 배치해야 한다고 주장했다. 두 속의 중요한 차이점은 몸 모양으로, 브라키오사우루스는 등뼈 계열이 23% 더 길고, 꼬리가 20~25% 더 길고 키가 크다. 이 분리는 2010년 다니엘 추레에 의해 거부되었지만, 2012년부터 대부분의 연구에서 ''Giraffatitan''을 인정했다.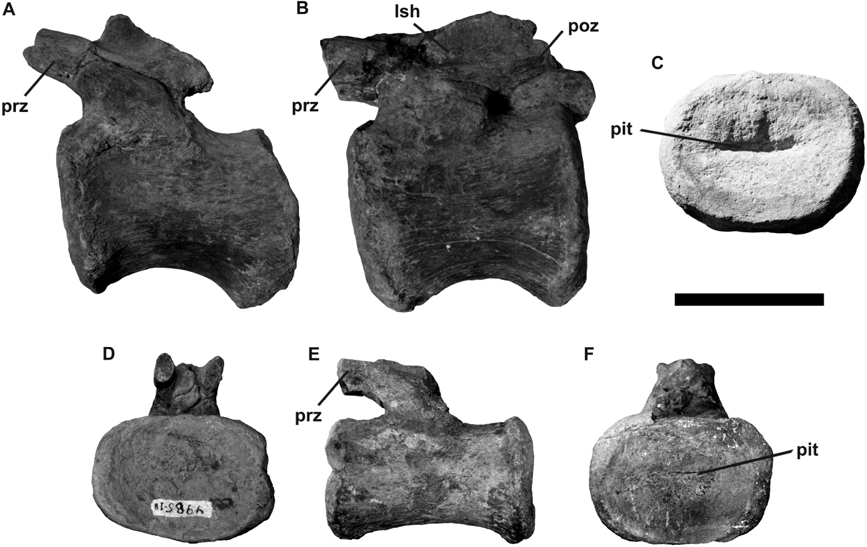
1947년, 포르투갈 아탈라이아에서 티토니아 시기 지층에서 브라키오사우루스과 화석이 발견되었다. 알베르-펠릭스 드 라파렌트와 조르주 즈비셰프스키는 1957년에 화석들을 ''Brachiosaurus atalaiensis'' 종으로 명명했다.[35] 폴 업처치, 바레트, 피터 도슨은 2004년 ''The Dinosauria''에서 아직 명명되지 않은 브라키오사우루스과 속으로 분류하며 의문을 제기했다. 2004년 책 출판 직전, 미겔 텔레스 안투네스와 옥타비오 마테우스는 2003년에 자체 속인 ''Lusotitan''으로 분류했다. 드 라파렌트와 즈비셰프스키는 화석들을 묘사했지만, 모식 표본을 지정하지 않았다. 안투네스와 마테우스는 부분적인 골반 이후 골격(MIGM4978, 4798, 4801–4810, 4938, 4944, 4950, 4952, 4958, 4964–4966, 4981–4982, 4985, 8807, 8793–87934)을 렉토타입으로 선정했다. 이 표본에는 28개의 척추뼈, 시브론, 갈비뼈, 가능한 어깨뼈, 상완골, 팔뚝뼈, 부분적인 왼쪽 골반, 아래 다리뼈, 오른쪽 발목 일부가 포함된다. 낮은 신경 가시, 상완골의 두드러진 삼각근 능선, 길쭉한 상완골, 위쪽으로 기울어진 장골의 긴 축은 ''Lusotitan''이 브라키오사우루스과임을 나타내며, 이는 2013년 분석 등 후속 연구에서 확인되었다.
화석은 로라시아 대륙 서부 지역의 일부였던 현재의 미국에 있는 중생대 쥐라기 후기 및 백악기 전기 지층에서 발굴되었다.
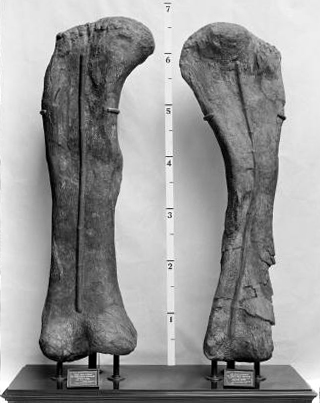
- 그림-1: ''B. altithorax''의 앞다리 뼈(''cf.'' 상완골) 화석과 발견자 엘머 S. 릭스(1900년 촬영).
- 그림-2: 미국시카고의 필드 자연사 박물관에 있는 연구실에서 ''B. altithorax''의 화석을 배경으로 작업하는 엘머 S. 릭스와 H. W. 멘케.
3. 1. 한국과의 연관성
2001년 미국 와이오밍주에서 보존율이 80%에 이르는 브라키오사우루스 화석이 발견되었는데, 이 발굴 작업에는 대한민국 연구팀이 공동으로 참여했다.[1] 이후 화석들은 국내로 옮겨져 보존 처리 및 조립되었으며, 이 진품 화석과 전신 골격은 현재 한국자연사박물관에서 전시되고 있다.[1]4. 분류

북아메리카에서 추가로 발견된 ''브라키오사우루스'' 재료는 흔하지 않으며 몇 개의 뼈로 구성되어 있다. 현재까지 모식 표본 재료와 겹치는 경우에만 이 속에 명확하게 귀속될 수 있으며, 두개골, 목, 앞쪽 등 부위, 또는 원위 사지나 발의 어떤 요소에 대한 언급도 잠정적으로 남아있다. 그럼에도 불구하고, 콜로라도,[40][12][13][14] 오클라호마,[40][15] 유타,[40][12] 그리고 와이오밍[40][41]에서 재료가 기술되었으며, 여러 다른 지역에서 아직 기술되지 않은 재료가 언급되었다.[3][40]
1883년, 미국 고생물학자 오트니엘 찰스 마시를 위해 화석 수집가였던 농부 마샬 파커 펠치는 가든 파크, 콜로라도 근처의 펠치 채석장 1에서 용각류 두개골을 발견했다고 보고했다. 두개골은 1m 경추와 근처의 옅은 노란색 사암에서 발견되었으며, 수집 시도가 실패하여 파괴되었다. 두개골은 YPM 1986으로 분류되었고, 피바디 자연사 박물관에 있는 마시에게 보내졌으며, 마시는 이를 1891년 ''브론토사우루스''의 골격 복원에 포함시켰다.[1][16][17] 펠치가 수집한 표본의 대부분은 마시의 사후인 1899년에 국립 자연사 박물관으로 보내졌으며, 여기에는 두개골도 포함되었고, 그 후 USNM 5730으로 분류되었다.[20][18][19]
1975년, 미국 고생물학자 잭 맥킨토시와 데이비드 버먼은 펠치 채석장의 두개골이 "일반적인 ''카마라사우루스'' 유형"임을 발견하는 동시에 그 근처에서 발견된 척추가 ''브라키오사우루스''에 속한다고 제안했다.[19]
1998년, 케네스 카펜터와 버지니아 티드웰은 펠치 채석장 두개골을 묘사하고, 공식적으로 ''브라키오사우루스'' sp. (불확실한 종)에 할당했다. 그들은 두개골이 나중에 ''지라파티탄''으로 알려진 ''B. brancai''와 유사하다는 것을 기반으로 ''브라키오사우루스''에 할당했다.[20][21] 2019년, 미국 고생물학자 마이클 D. 디에믹과 매튜 T. 카라노는 펠치 채석장 두개골을 재검토하여 ''카마라사우루스'' sp.에 할당되었던 이빨을 두개골에 속하지 않는다고 결론지었다. 그들은 또한 두개골을 불특정 종이 아닌 ''B. altithorax'' 자체에 할당하는 것이 가장 경제적이라고 생각했다.[22]

1987년 젠슨에 의해 간략하게 묘사된 여러 추가 표본 중 하나인 상완골 USNM 21903은 서부 콜로라도의 포터 크릭 채석장에서 1943년경 우라늄 탐사자인 비비안과 다니엘 존스에 의해 발견되어 스미소니언 협회에 기증되었다.[40][12][25] 2009년 테일러에 따르면, 2.13m의 큰 크기에도 불구하고 ''브라키오사우루스''에 명확하게 언급할 수 없다. 젠슨은 1971년과 1975년 포터 크릭 지역에서 직접 일하면서, 중간 등 척추뼈, 불완전한 왼쪽 장골, 왼쪽 요골 및 오른쪽 중수골을 포함하는 분리된 표본 BYU 4744를 발굴했다. 2009년 테일러에 따르면, 이 표본은 모식 표본과 겹치는 한 ''B. altithorax''에 확실하게 언급될 수 있다.[40]
2012년, 호세 카르발리도와 동료들은 길이가 약 2m인 작은 어린 개체의 거의 완전한 후두개 골격을 보고했다. "토니"라는 별명이 붙고 SMA 0009로 분류된 이 표본은 와이오밍주 북중부의 비그혼 분지의 모리슨 층에서 유래되었다. 원래 디플로도쿠스과에 속한다고 생각했지만, 나중에는 브라키오사우루스과로 재해석되었으며, 아마도 ''B. altithorax''에 속했을 것이다.[26] 2018년, 지금까지 발견된 가장 큰 용각류 발이 와이오밍주 웨스턴 카운티의 블랙 힐스에서 보고되었다. 비록 ''브라키오사우루스''에 속할 가능성이 있지만, 저자들은 조심스럽게 이를 불확실한 브라키오사우루스과로 분류했다.[27] 그러나 이 두 표본을 각 분류군에 할당하는 것은 2019년 디에믹과 카라노에 의해 나중에 의문이 제기되었다.[1]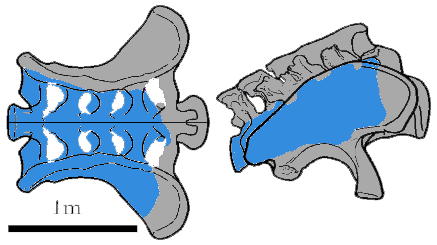
1958년 프랑스 석유 지질학자 F. 누가레드는 동부 알제리의 사하라 사막에서 조각난 브라키오사우루스과 화석을 발견했다고 보고했다.[37] 이를 바탕으로, 알베르-펠릭스 드 라파렌은 1960년에 해당 종을 ''브라키오사우루스 누가레디''(Brachiosaurus nougaredi)로 명명하고 묘사했다.[38] 최근 연구에서는 이를 초기 백악기 후기 알비안 시대에 속하는 것으로 간주한다.[50]
''B. nougaredi''는 2004년 별개의, 명명되지 않은 브라키오사우루스과 속을 나타내는 것으로 여겨졌지만,[50] 2013년 필립 D. 매니언과 그의 동료들의 분석에 따르면 유해가 서로 멀리 떨어진 곳에서 수집되었기 때문에 여러 종에 속할 수 있다고 밝혀졌다.[39] 중수골은 불확실한 티타노사우루스형류에 속하는 것으로 결론지어졌다. 천골은 2013년에 분실된 것으로 보고되었다. 이는 국립 자연사 박물관의 소장품에서 다시 발견될 때까지 분석되지 않았고, 불확실한 용각류를 나타내는 것으로 잠정적으로 간주되었다. 1960년 추정된 원래 전체 길이는 1.3m였으며, 이는 ''B. altithorax''의 0.91m에 비해 큰 것이다.[38]
1903년, 엘머 S. 릭스는 ''브라키오사우루스''를 용각류의 분명한 구성원으로 간주했다. 속의 유효성을 결정하기 위해 그는 이전에 명명된 속인 ''카마라사우루스'', ''아파토사우루스'', ''아틀란토사우루스'', 그리고 ''암피코일리아스''와 비교했다.[57] 1904년, 리그스는 ''브라키오사우루스''가 ''하플로칸토사우루스''와 더 밀접한 관련이 있다고 결정했다. 리그스는 다른 분류군과의 차이점이 별도의 과, 브라키오사우루스과(Brachiosauridae)를 명명할 만큼 충분히 중요하다고 생각했으며, 이 중 ''브라키오사우루스''는 이름의 유래가 된 속이다.[61]
1990년대 이후, 컴퓨터 기반의 분지학 분석은 브라키오사우루스과의 유효성에 의문을 제기했다. 1997년, 레오나르도 살가도는 전통적으로 브라키오사우루스과로 간주되는 종들이 더 큰 그룹인 티타노사우루스형류의 줄기에서 파생된 것이며, 별도의 자체 분기가 아니라는 분석을 발표했다.[66]
그 이후로 많은 분지학 분석은 적어도 일부 속이 브라키오사우루스과에 할당될 수 있으며, 이 그룹이 티타노사우루스형류 내의 기본 분기임을 시사했다.[68]
2012년 Michael D. D'Emic이 발표한 분지도는 다음과 같다:[69]
테일러의 2009년 연구에 따르면, ''B. 알티토락스''는 다른 브라키오사우루스과와 함께 고전적인 특징인 위팔뼈가 대퇴골만큼 거의 길거나 (상완골 길이 대 대퇴골 길이 비율이 최소 0.9) 공유한다. 또 다른 공유 형질은 매우 평평한 대퇴골 몸통으로, 가로 폭이 앞뒤 폭의 1.85배 이상이다.[40]
최초의 발견은 미국의 고생물학자 엘머 S. 릭스에 의해 이루어졌다.
속명 ''Brachiosaurus''는 βραχιων(''brachion'') "팔" + σαυρος(''sauros'') "도마뱀"의 합성어이다.
4. 1. 종
1914년, 독일 고생물학자 베르너 야넨슈는 독일령 동아프리카(현재의 탄자니아) 텐다구루 지층에서 발굴된 화석들과 ''B. altithorax'' 사이의 차이점과 공통점을 바탕으로 두 종을 명명했다. 더 크고 완전한 표본은 ''Brachiosaurus brancai'', 더 작고 알려진 바가 적은 종은 ''Brachiosaurus fraasi''였다.[31]
베를린 자연사 박물관에는 ''B. brancai''로 분류된 자료가 풍부했으며, 이 중 일부는 제2차 세계 대전 중에 파괴되었다.[40]
야넨슈는 텐다구루에서 발굴된 "골격 S"(Skelett S)를 기반으로 ''B. brancai''에 대한 설명을 썼지만,[31] 나중에 이것이 부분적인 두 개체, SI과 SII로 구성되어 있다는 것을 깨달았다.[32] 1935년에 SI(현재 MB.R.2180)을 렉토타입으로 지정했다.[33][34]
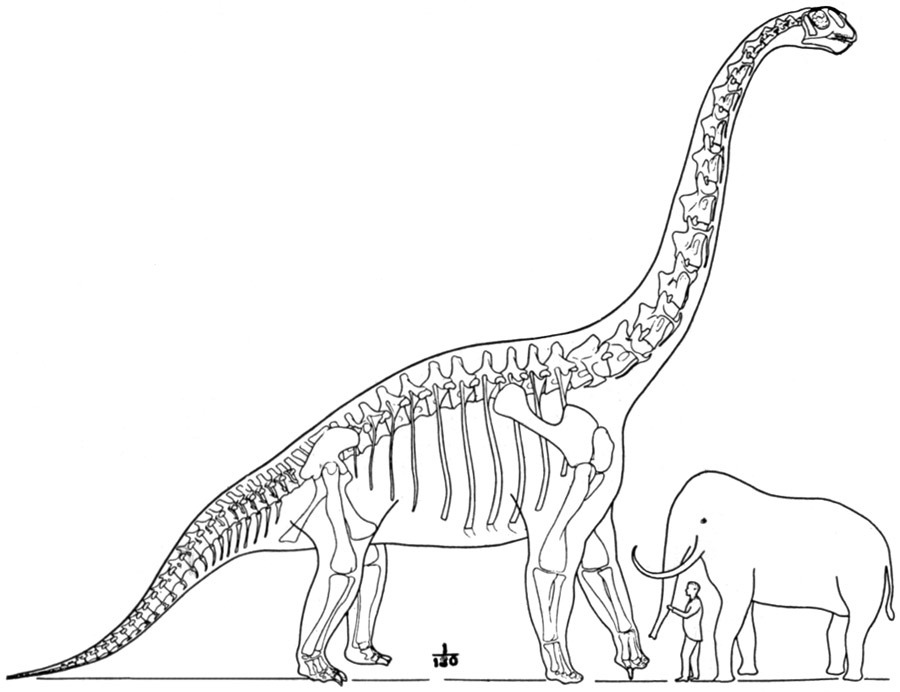
1988년 그레고리 S. 폴은 ''B. brancai'' 골격의 새로운 재구성을 발표하여, 이 골격과 ''B. altithorax'' 사이의 비율 차이를 강조했다. 1991년 조지 올셰프스키는 척추의 변이를 언급하며 ''Giraffatitan''을 완전한 속 수준으로 격상시켰다.[24]
2009년 테일러는 ''B. altithorax''와 텐다구루 자료 사이에 두 종 모두에서 알려진 모든 요소에 상당한 차이가 있다는 것을 발견했다. 26개의 뚜렷한 골학적(뼈 기반) 특징을 발견했는데, 이는 ''디플로도쿠스''와 ''바로사우루스'' 사이의 차이보다 더 컸으며, 따라서 아프리카 자료를 ''Giraffatitan brancai''로, 자체 속(''Giraffatitan'')에 배치해야 한다고 주장했다.[40]
1957년, 알베르-펠릭스 드 라파렌트와 조르주 즈비셰프스키는 포르투갈의 아탈라이아에서 티토니아 시기에 속하는 지층에서 발견된 브라키오사우루스과 화석을 ''Brachiosaurus atalaiensis''로 명명했다.[35] 2003년에 미겔 텔레스 안투네스와 옥타비오 마테우스는 이 종을 자체 속인 ''Lusotitan''으로 분류하였다.[36]
1960년, 알베르-펠릭스 드 라파렌은 동부 알제리의 사하라 사막에서 발견된 화석을 바탕으로 ''Brachiosaurus nougaredi''를 명명하고 묘사했다.[38]
''B. nougaredi''는 2004년 별개의, 명명되지 않은 브라키오사우루스과 속을 나타내는 것으로 여겨졌지만,[50] 2013년 분석에 따르면 유해가 서로 멀리 떨어진 곳에서 수집되었기 때문에 여러 종에 속할 수 있다고 밝혀졌다.[39]
속명 ''Brachiosaurus''는 βραχιων(''brachion'') "팔" + σαυρος(''sauros'') "도마뱀"의 합성어이다.
| 종 | 명명자 | 비고 |
|---|---|---|
| Brachiosaurus altithorax | 엘머 S. 릭스 | |
| Brachiosaurus atalaiensis | de Lapparent et Zbyszewski | 최근 연구에서는 티타노사우루스형류로 분류된다는 설도 있다. |
| Brachiosaurus nougaredi | de Lapparent |
5. 고생물학

대부분의 ''브라키오사우루스 알티토락스''(Brachiosaurus altithorax) 크기 추정치는 브라키오사우루스과 ''지라파티탄''(Giraffatitan, 이전 ''B. brancai'')을 기반으로 하는데, ''지라파티탄''은 ''브라키오사우루스''보다 완전한 화석 자료가 많다. 두 종은 유해가 비교적 많이 발견된 가장 큰 브라키오사우루스과 공룡이다. 북아메리카 ''브라키오사우루스''는 모식표본(가장 완전한 표본)이 아성체로 보여 불확실한 요소가 있는데, 이는 어깨뼈인 오훼돌기와 견갑골 사이 융합되지 않은 봉합으로 나타난다.[40]
모식 표본 질량은 28.3MT~46.9MT 범위로 추정되었다.[40][41][58][42][45] 벤슨 등은 최대 56MT~58MT을 제시했지만,[43][44] 큰 오차 범위와 정밀도 부족으로 의문이 제기되었다.[45] ''브라키오사우루스'' 길이는 20~22미터,[58][46] ,[42][76] 높이는 ,[76] .[58][47]로 추정되었다.
''지라파티탄'' 골격(MB.R.2181) 사지 뼈는 ''브라키오사우루스'' 모식 표본과 크기가 유사했지만, 비례 차이로 ''브라키오사우루스''보다 다소 가벼웠다. ''지라파티탄''은 31.5MT,[33][58] 39.5MT,[48] ,[49] 23.3MT,[40] .로 추정되었다. ''지라파티탄'' 표본 MB.R.2181 역시 최대 크기를 반영하지 않을 가능성이 있는데, 종아리뼈(표본 HMXV2)가 MB.R.2181보다 13% 더 길기 때문이다.[40]
''브라키오사우루스''는 작은 두개골, 긴 목, 높은 타원체 단면 큰 몸통, 길고 근육질 꼬리, 가늘고 기둥 모양 사지를 가진 네 발 동물이다.[50] 허파꽈리와 연결된 큰 기낭이 목과 몸통에 존재, 뼈 흡수를 통해 척추, 갈비뼈를 침범하여 몸 밀도를 크게 감소시켰다.[51][52] 목은 모식 표본에 보존되지 않았지만, ''지라파티탄''은 용각류 기준으로도 매우 길었으며, 13개 길쭉한 경추로 구성되었다.[53] 목은 약간 S자형이며, 아래, 위쪽은 구부러지고 가운데는 곧게 뻗었다.[54] ''브라키오사우루스''는 ''지라파티탄''과 길쭉한 경늑골(목 갈비뼈)을 공유했을 것이며, 목 아랫부분을 따라 뻗어 나가 여러 이전 척추를 겹치게 했다. 이 뼈 막대들은 목 근육 끝에 부착, 근육이 몸통에 가깝게 위치하면서 목 원위 부분을 작동시켜 가볍게 했다.[54][55]
''브라키오사우루스'', ''지라파티탄''은 3, 5번째 흉추(등) 척추 사이 작은 어깨 혹이 있었을 것이며, 옆, 위로 향하는 척추 돌기가 길어져 목 근육 부착을 위한 추가 표면을 제공했다.[56] 갈비뼈는 다른 용각류보다 깊었다.[57] 상완골(위팔뼈), 넙적다리뼈(넓적다리뼈) 길이
5. 1. 목 자세 논쟁
브라키오사우루스과의 목 자세를 둘러싼 논쟁은 계속되고 있으며, 추정치는 거의 수직에서 수평 방향까지 다양하다.[82] 1999년까지는 용각류의 목이 거의 수직이라는 주장이 일반적이었으나, 스티븐스와 패리시는 용각류의 목이 S자형으로 구부러진 자세를 유지할 만큼 유연하지 않으며, 수평으로 유지되었다고 주장했다.[56][72]이러한 연구를 반영하여, 다양한 신문들은 필드 박물관의 ''브라키오사우루스'' 복원 모형이 위로 향하게 구부러진 목을 가지고 있다고 비판했다. 그러나 박물관 고생물학자인 올리비에 리펠과 크리스토퍼 브로츄는 1999년에 긴 앞다리와 위로 경사진 척추를 언급하며 수직에 가까운 자세를 옹호했다. 그들은 또한 근육 부착을 위한 가장 발달된 신경 가시가 어깨 거들 부위에 위치하여 기린과 같은 자세로 목을 들어 올릴 수 있게 해주었을 것이라고 주장했다. 덧붙여 이러한 자세는 목을 낮추는 것보다 적은 에너지를 필요로 했을 것이고, 척추 사이의 디스크는 목을 낮춘 상태에서 장기간 동안 발생한 압력에 대응할 수 없었을 것이라고 주장했다 (물을 마시기 위해 목을 낮추는 것은 가능했을 것이다).[73]
최근의 몇몇 연구들 또한 더 위로 향한 목을 옹호했다. 크리스티안과 젬스키(2007)는 ''지라파티탄''의 목 중간 부분이 60~70도 기울어져 있었고, 수평 자세는 짧은 시간 동안만 유지될 수 있다고 추정했다.[54]
브라키오사우루스과 공룡이 머리를 심장 위로 높이 들었을 경우, 심혈관 계통에 큰 부담이 되었을 것이다. ''브라키오사우루스''의 심장은 뇌에 혈액을 공급하기 위해 기린 혈압의 두 배로 혈액을 펌프질해야 했을 것이며, 무게는 400kg에 달했을 것으로 추정된다.[74] 머리와 심장 사이의 거리는 목의 S자형 곡선에 의해 2m 이상 줄어들었을 것이다.[54] 또한, 목은 이동 중에 20도 정도 낮아졌을 수 있다.[54] ''지라파티탄''의 속귀를 연구한 쿵가 & 키르쉬(2001)는 브라키오사우루스과가 먹이를 먹는 동안 등쪽-배쪽 방향보다 측면 방향으로 목을 더 자주 움직였을 것이라고 결론지었다.[54][75]
5. 2. 식성
''Brachiosaurus''영어는 높이 솟아 있는 잎을 먹는 높이 먹는 동물이었을 것으로 추정된다. 목을 수직에 가깝게 세우지 않았더라도 머리 높이는 땅 위 9m 이상이었을 수 있다.[41][76] 주로 5m 이상 높이의 잎을 먹었을 것이지만, 3m에서 5m 사이의 낮은 곳에서도 먹이를 먹었을 가능성도 있다.[76]식단은 은행나무, 침엽수, 고사리류, 큰 소철류 등으로 구성되었을 가능성이 높으며, 2007년 연구에 따르면 하루에 200kg에서 400kg의 식물 물질을 섭취한 것으로 추정된다.[76] ''Brachiosaurus''영어의 먹이 섭취는 위아래로 간단한 턱 운동을 포함했다.[77] 다른 용각류와 마찬가지로, 추가적인 구강 처리 없이 식물 물질을 삼키고, 음식 처리를 위해 후장 발효에 의존했을 것이다.[82] 이빨은 다소 숟가락 모양이며 끌과 같았다. 이러한 이빨은 선택적이지 않은 잘게 물어뜯기에 최적화되어 있으며,[77] 상대적으로 넓은 턱은 많은 양의 식물 재료를 잘라낼 수 있었다. 40톤의 ''Brachiosaurus''영어가 반 톤의 사료가 필요하더라도, 머리의 정상적인 자르기 동작만으로 식단 요구 사항을 충족할 수 있었을 것이다. 하루에 16시간 동안 먹이를 먹고, 한 번에 10분의 1에서 3분의 2 킬로그램을 물어뜯으며, 분당 1~6번 물어뜯는다면, 일일 식사량은 체중의 약 1.5%에 달하며, 현대 코끼리의 요구량과 비슷했을 것이다.
''Brachiosaurus''영어는 다른 많은 용각류 종과 함께 서식지인 모리슨에서 살았기 때문에, 더 높은 곳에서 먹이를 먹기 위한 전문화는 틈새 분할 시스템의 일부였으며, 다양한 분류군은 서로 직접적인 경쟁을 피했다. 전형적인 먹이 나무는 세쿼이아덴드론과 유사했을 것이다. 그러한 키 큰 침엽수가 모리슨에서 비교적 드물었다는 사실은 ''Brachiosaurus''영어가 텐다가루에서 가장 풍부한 용각류 중 하나였던 관련 종인 ''Giraffatitan''영어보다 생태계에서 훨씬 덜 흔했던 이유를 설명할 수 있다. ''Brachiosaurus''영어는 팔이 짧고 어깨가 낮아 ''Giraffatitan''영어만큼 높은 곳에서 먹는 데 잘 적응하지 못했다.
''Brachiosaurus''영어는 뒷다리로 서서 꼬리를 지면 지지대로 사용하여 먹이를 먹을 수 있다는 주장이 제기되었다.[58] 그러나 하인리히 맬리슨(Heinrich Mallison)의 용각류의 뒷다리 서기 능력에 대한 상세한 물리적 모델링 기반 분석에 따르면, ''Brachiosaurus''영어는 특이한 몸 모양과 사지 길이 비율로 인해 뒷다리로 서는 데 매우 부적합했다. 몸의 중심이 앞으로 향해 있어서 안정성에 문제가 생기고, 직립 자세를 유지하기 위해 엉덩이에 무리한 힘이 필요했을 것이다. 또한 다른 용각류는 뒷다리로 서는 것이 먹이 섭취 높이를 3배나 증가시켰을 수 있지만, ''Brachiosaurus''영어는 33% 증가했을 뿐이다.[78]
''Brachiosaurus''영어의 목이 아래로 움직일 수 있었기 때문에, 똑바로 서 있는 상태에서 발 높이의 열린 물에 닿을 수 있었을 것이다. 현대 기린은 앞다리를 벌려 입을 상대적으로 수평 위치로 낮춰 물을 더 쉽게 삼킨다. ''Brachiosaurus''영어가 이러한 방식으로 안정적인 자세를 취할 수 있었을 가능성은 낮으며, 이로 인해 동물이 호수를 냇물 표면에 코를 거의 수직으로 꽂아야 했을 것이다.
브라키오사우루스는 용각류의 다른 종과는 달리 앞다리가 뒷다리보다 길고, 어깨가 뒤쪽으로 기울어져 있어서 더 높은 곳의 식물을 먹을 수 있었다. 머리 부분도 특징적인데, 부풀어 오른 콧구멍을 가지고 있으며, 머리 가장 높은 곳에 콧구멍이 있다. 머리는 작고 뇌의 크기도 200g 정도이며, 씹는 힘이 그다지 강하지 않았기 때문에 나뭇가지나 잎을 통째로 삼켜서 먹었던 것으로 보인다. 사회성도 있어서 무리 생활을 했던 것으로 생각되며, 특히 밤이나 이동 시에는 어린 개체가 중심이 되어 어른 개체가 이를 둘러싸서 보호했다고 여겨진다.
5. 3. 콧구멍 위치
브라키오사우루스와 같은 신용각류의 뼈로 된 콧구멍은 크고 머리 꼭대기에 위치했다. 전통적으로, 용각류의 살이 많은 콧구멍은 머리 꼭대기, 대략 뼈로 된 콧구멍 뒤쪽에 위치했을 것으로 생각되었는데, 이는 이 동물이 수생 생활을 하며 물속에 잠겼을 때 큰 콧구멍을 스노클로 사용했을 것이라는 잘못된 생각 때문이었다. 미국의 고생물학자 로렌스 M. 위트머는 2001년에 이러한 재구성을 거부하며, 모든 살아있는 척추동물 육상 동물은 바깥쪽 살이 많은 콧구멍이 뼈로 된 콧구멍 앞에 위치해 있다는 점을 지적했다. 이러한 용각류의 살이 많은 콧구멍은 훨씬 더 앞쪽, 즉 콧구멍 앞쪽, 콧구멍 뼈 앞쪽으로 멀리 뻗어 있는 우묵한 곳에 위치했을 것이다.[79]
체르카스는 브라키오사우루스과 코의 독특한 기능에 대해 추측했으며, 피부 자국이 있는 머리가 발견되지 않는 한 콧구멍의 위치를 결정할 결정적인 방법이 없다고 지적했다. 그는 확장된 콧구멍이 동물이 냄새를 맡는 능력과 관련된 조직을 위한 공간을 만들어, 적절한 식물을 냄새 맡는 데 도움이 되었을 것이라고 제안했다. 그는 또한 현대 파충류에서 둥글고 확대되며 위로 솟아오른 콧구멍 뼈의 존재는 살이 많은 뿔과 혹 모양의 돌출부와 연관될 수 있으며, 코가 큰 ''브라키오사우루스''와 다른 용각류는 생물학적 장식 코 볏을 가졌을 수 있다고 언급했다.[80]
''브라키오사우루스''를 포함한 용각류가 뼈로 된 콧구멍 위치를 근거로 코끼리처럼 위쪽으로 더 멀리 뻗기 위해 코를 가졌을 수 있다는 주장이 제기되었다. 파비앙 놀과 동료들은 2006년에 디플로도쿠스와 ''카마라사우루스''에 대해 이를 반박하며, 뇌의 안면 신경 구멍이 작다는 것을 발견했다. 따라서 안면 신경은 코끼리처럼 커지지 않았는데, 코끼리의 경우 정교한 코 근육을 작동시키는 데 관여한다. 그러나 놀과 동료들은 또한 ''기라파티탄''의 안면 신경이 더 크다는 점을 언급하며, 따라서 이 속에서 코끼리 코의 가능성을 배제할 수 없다고 보았다.[81]
과거에는 두개골의 모양에서 현생 코끼리처럼 근육 조직으로 만들어진 코가 있었을 가능성이 지적되었다. 머리 부분도 특징적인데, 부풀어 오른 콧구멍을 가지고 있으며, 머리 가장 높은 곳에 콧구멍이 있다(이 콧구멍 위치는 과거 수중 생활설의 근거 중 하나로 여겨지기도 했다).
6. 대중문화

''브라키오사우루스''는 가장 상징적인 공룡 중 하나로 불리지만, 가장 대중적인 묘사는 아프리카 종인 ''B. brancai''를 기반으로 하며, 이후 자체 속인 ''Giraffatitan''으로 옮겨졌다.[40] 1991년에는 소행성대 소행성 9954 브라키오사우루스가 이 속을 기리기 위해 명명되었다.[98][99] ''브라키오사우루스''는 1993년 영화 ''쥬라기 공원''에서 처음으로 컴퓨터로 생성된 공룡으로 등장했다.[102] 이 효과는 당시 획기적인 것으로 여겨졌으며, 영화 속 등장인물들이 공룡을 처음 봤을 때 느낀 경외감은 관객들에게도 그대로 전달되었다.[100][101] 영화 속 ''브라키오사우루스''의 움직임은 기린의 걸음걸이에 코끼리의 몸집을 결합하여 만들어졌다. 영화 후반부에서는 ''브라키오사우루스''가 인간 등장인물과 상호 작용할 때 애니매트로닉스 머리와 목을 사용했다.[102] ''쥬라기 공원''에 사용된 ''브라키오사우루스''의 디지털 모델은 나중에 1997년 스페셜 에디션 영화 ''스타워즈 에피소드 4: 새로운 희망''의 론토 모델의 시작점이 되었다.[103]
참조
[1]
논문
Redescription of Brachiosaurid Sauropod Dinosaur Material From the Upper Jurassic Morrison Formation, Colorado, USA
2020-04
[2]
서적
Dinosaurs: The Encyclopedia
https://archive.org/[...]
McFarland & Company
[3]
서적
Vertebrate Paleontology in Utah
Utah Geological Survey
[4]
서적
Paleontology and Geology of the Dinosaur Triangle
Museum of Western Colorado
[5]
논문
Geology and artesian water supply of the Grand Junction area, Colorado
https://pubs.er.usgs[...]
[6]
논문
The largest known dinosaur
https://archive.org/[...]
[7]
웹사이트
θώραξ
https://www.perseus.[...]
Perseus Digital Library
2018-04-06
[8]
논문
A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)
[9]
간행물
Expect Awe-Struck Travelers
http://fieldmuseum.o[...]
The Field Museum
1999-11-26
[10]
웹사이트
Captions from Selected Historical Photographs (caption number GN89396_52c)
http://www.fieldmuse[...]
The Field Museum Photo Archives
2009-08-27
[11]
웹사이트
Oldengate Bridge
http://www.wdwthemep[...]
2019-03-06
[12]
논문
New brachiosaur material from the Late Jurassic of Utah and Colorado
[13]
서적
The Continental Jurassic: Transactions of the Continental Jurassic Symposium
Museum of Northern Arizona Bulletin
[14]
서적
Western Association of Vertebrate Paleontologists and Southwest Paleontological Symposium – Proceedings 2001
Mesa Southwest Museum Bulletin
[15]
논문
First occurrence of ''Brachiosaurus'' (Dinosauria, Sauropoda) from the Upper Jurassic Morrison Formation of Oklahoma
https://sauroposeido[...]
[16]
웹사이트
Paleobiology Collections Search
https://collections.[...]
2022-03-11
[17]
웹사이트
cf. Brachiosaurus sp
https://www.si.edu/o[...]
2022-03-11
[18]
논문
Restoration of ''Triceratops''
http://marsh.dinodb.[...]
[19]
논문
Description of the palate and lower jaw of the sauropod dinosaur ''Diplodocus'' (Reptilia: Saurischia) with remarks on the nature of the skull of ''Apatosaurus''
[20]
논문
Preliminary description of a ''Brachiosaurus'' skull from Felch Quarry 1, Garden Park, Colorado
[21]
서적
The Continental Jurassic: Transactions of the Continental Jurassic Symposium
Museum of Northern Arizona Bulletin
[22]
논문
Redescription of brachiosaurid sauropod dinosaur material from the Upper Jurassic Morrison Formation, Colorado, USA
2019
[23]
논문
Three new sauropod dinosaurs from the Upper Jurassic of Colorado
[24]
논문
A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia
http://www.miketaylo[...]
2018-04-14
[25]
논문
Late Jurassic North American brachiosaurids
[26]
논문
New information on a juvenile sauropod specimen from the Morrison Formation and the reassessment of its systematic position
http://doc.rero.ch/r[...]
[27]
논문
The real Bigfoot: a pes from Wyoming, USA is the largest sauropod pes ever reported and the northernmost occurrence of brachiosaurids in the Upper Jurassic Morrison Formation
2018
[28]
논문
Die Wirbelsäule von ''Brachiosaurus brancai''
[29]
논문
Die Gliedmaßen und Gliedmaßengürtel der Sauropoden der Tendaguru-Schichten
http://www.miketaylo[...]
[30]
서적
African dinosaurs unearthed: The Tendaguru Expeditions
Indiana University Press
[31]
논문
Übersicht über der Wirbeltierfauna der Tendaguru-Schichten nebst einer kurzen Charakterisierung der neu aufgefuhrten Arten von Sauropoden
https://people.ohio.[...]
[32]
논문
Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition
[33]
논문
Correction: A re-evaluation of ''Brachiosaurus altithorax'' Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914)"
[34]
논문
Die Schädel der Sauropoden ''Brachiosaurus'', ''Barosaurus'' und ''Dicraeosaurus'' aus den Tendaguru-Schichten Deutsch-Ostafrikas
http://www.miketaylo[...]
[35]
논문
Les dinosauriens du Portugal
http://paleoglot.org[...]
2018-03-12
[36]
논문
Dinosaurs of Portugal
2003
[37]
논문
Nouvelles découvertes de Vertébrés dans les séries continentales au Nord d'Edjelch (Sahara central)
[38]
논문
Les dinosauriens du "continental intercalaire" du Sahara central"
http://www.paleoglot[...]
2010-09-25
[39]
논문
Osteology of the Late Jurassic Portuguese sauropod dinosaur ''Lusotitan atalaiensis'' (Macronaria) and the evolutionary history of basal titanosauriforms
http://docentes.fct.[...]
[40]
논문
A re-evaluation of ''Brachiosaurus altithorax'' Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from ''Giraffatitan brancai'' (Janensh 1914)
http://www.miketaylo[...]
[41]
서적
Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain region, U.S.A
http://econtent.unm.[...]
New Mexico Museum of Natural History and Science
[42]
논문
A new method to calculate allometric length-mass relationships of dinosaurs
2001-03-26
[43]
논문
Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage
[44]
논문
Cope's rule and the adaptive landscape of dinosaur body size evolution
2018
[45]
논문
The accuracy and precision of body mass estimation in non-avian dinosaurs
2020
[46]
서적
The Princeton Field Guide to Dinosaurs
Princeton: Princeton University Press
[47]
서적
Biology of the sauropod dinosaurs
Indiana University Press
2011
[48]
논문
Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs
[49]
논문
A new body mass estimation of ''Brachiosaurus brancai'' Janensch, 1914 mounted and exhibited at the Museum of Natural History (Berlin, Germany)
[50]
서적
The Dinosauria, Second Edition
https://archive.org/[...]
Univ of California Press, Berkeley
[51]
논문
Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs
http://doc.rero.ch/r[...]
[52]
논문
The evolution of vertebral pneumaticity in sauropod dinosaurs
http://doc.rero.ch/r[...]
[53]
논문
Why sauropods had long necks; and why giraffes have short necks
2013
[54]
논문
Reconstruction of the cervical skeleton posture of ''Brachiosaurus brancai'' Janensch, 1914 by an analysis of the intervertebral stress along the neck and a comparison with the results of different approaches
[55]
논문
Histology shows that elongated neck ribs in sauropod dinosaurs are ossified tendons
2012
[56]
논문
Nuchal ligament reconstructions in diplodocid sauropods support horizontal neck feeding postures
2016
[57]
논문
"''Brachiosaurus altithorax'', the largest known dinosaur"
https://zenodo.org/r[...]
[58]
논문
The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, ''Giraffatitan'', and a comparison of the world's largest dinosaurs
http://gspauldino.co[...]
[59]
서적
The sauropods: evolution and paleobiology
University of California Press
2005
[60]
논문
British Museum East Africa Expedition: Account of the work done in 1930
[61]
논문
Structure and relationships of opisthocoelian dinosaurs. Part II. The Brachiosauridae
http://www.thermopwy[...]
[62]
서적
The Dinosauria
University of California Press
[63]
논문
Osteology, paleobiology, and relationships of the sauropod dinosaur ''Sauroposeidon''
[64]
논문
Studies of the sauropod dinosaur ''Astrodon'' Leidy
[65]
논문
Comments on ''Chubutisaurus insignis'' del Corro (Saurischia, Sauropoda)
1993
[66]
논문
Evolution of titanosaurid sauropods. I: phylogenetic analysis based on the postcranial evidence
[67]
논문
First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition
[68]
논문
The illusory evidence for Asian Brachiosauridae: new material of ''Erketu ellisoni'' and a phylogenetic appraisal of basal Titanosauriformes
http://digitallibrar[...]
[69]
논문
The early evolution of titanosauriform sauropod dinosaurs
https://deepblue.lib[...]
[70]
논문
Tipsy punters: sauropod dinosaur pneumaticity, buoyancy and aquatic habits
[71]
논문
Three new sauropod dinosaurs from the Upper Jurassic of Colorado
https://scholarsarch[...]
[72]
논문
Neck posture and feeding habits of two Jurassic sauropod dinosaurs
[73]
논문
Paleontologists defend dinosaur mount
http://libsysdigi.li[...]
[74]
서적
Dinosaurs: A Concise Natural History
Cambridge University Press
[75]
논문
Von Hochleistungsherzen und wackeligen Hälsen
[76]
서적
Jurassic West: The Dinosaurs of the Morrison Formation and Their World
Indiana University Press
[77]
서적
The Sauropods: Evolution and Paleobiology
https://archive.org/[...]
University of California
[78]
간행물
"Rearing Giants{{snd}}kinetic-dynamic modeling of sauropod bipedal and tripodal poses"
Indiana University Press
[79]
논문
Nostril position in dinosaurs and other vertebrates and its significance for nasal function
[80]
서적
Dinosaurs: a Global View
https://archive.org/[...]
Dragons' World
[81]
논문
Paleoneurological evidence against a proboscis in the sauropod dinosaur ''Diplodocus''
[82]
논문
Biology of the sauropod dinosaurs: the evolution of gigantism
[83]
서적
An Odyssey in Time: Dinosaurs of North America
NorthWord Press
[84]
논문
New information on a juvenile sauropod specimen from the Morrison Formation and the reassessment of its systematic position
http://doc.rero.ch/r[...]
[85]
서적
An Odyssey in Time: Dinosaurs of North America
NorthWord Press
[86]
논문
The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation
http://doc.rero.ch/r[...]
[87]
간행물
"Appendix"
Indiana University Press
[88]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[89]
간행물
"Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A."
Utah Geological Survey
[90]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[91]
서적
Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A
New Mexico Museum of Natural History and Science
[92]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation
New Mexico Museum of Natural History and Science
[93]
웹사이트
Brachiosaurus Replica {{!}} Artwork {{!}} Chicago Park District
https://www.chicagop[...]
2024-10-07
[94]
웹사이트
Field's Big Dinosaur Departing for O'Hare
https://www.chicagot[...]
2024-10-07
[95]
웹사이트
Replica of Dinosaur Fossil Gives O'Hare Passengers Monstrous Welcome
https://www.chicagot[...]
2024-10-07
[96]
웹사이트
Tuesday is Dino-day at Chicago’s O’Hare Airport
https://www.usatoday[...]
2024-10-07
[97]
웹사이트
The Most Unique Things to Do and See at O'Hare International Airport, Including a Dinosaur
https://www.thrillis[...]
2024-10-07
[98]
웹사이트
JPL Small-Body Database Browser: 9954 Brachiosaurus (1991 GX7)
http://ssd.jpl.nasa.[...]
NASA
2007-04-28
[99]
웹사이트
Minor Planet Names: Alphabetical List
http://www.cfa.harva[...]
Smithsonian Astrophysical Observatory
2007-02-10
[100]
웹사이트
The Jurassic Park Period: How CGI Dinosaurs Transformed Film Forever
https://www.theatlan[...]
2018-07-20
[101]
논문
The WOW Factor
https://books.google[...]
[102]
서적
The Making of Jurassic Park
Boxtree Ltd
[103]
웹사이트
Ronto
https://www.starwars[...]
Star Wars.com
2009-01-13
[104]
뉴스
2億年前の大型恐竜、本当は重くなかった 英米研究
https://www.afpbb.co[...]
[105]
웹사이트
JWE2ツールボックス 恐竜データべース ブラキオサウルス
https://www.paleo.gg[...]
2024-10-01
[106]
저널
The accuracy and precision of body mass estimation in non-avian dinosaurs
https://onlinelibrar[...]
2020
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com