망형충류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
망형충류는 세포 외부에 위치하지만 세포막으로 둘러싸인 필라멘트를 가지는 미생물 그룹이다. 이들은 세포 소기관인 보스로솜에 의해 형성되며, 단핵 세포로 타원형 모양을 띤다. 망형충류는 해양, 기수역, 담수 등 다양한 환경에 서식하며, 유기물을 분해하는 부생성 생물로서 생태계에서 중요한 역할을 한다. 일부 종은 해조류나 해초에 기생하며, 고도불포화지방산, 카로티노이드, 스쿠알렌 등을 생산하여 인간에게 유용하게 사용되기도 한다. 분류학적으로는 부등편모조류에 속하며, 라비린추라강에 라비린추라목과 야브레츠보카비목 등이 포함된다.
더 읽어볼만한 페이지
- 부등편모조류 - 유사균류
유사균류는 과거 균류로 분류되었으나 진균류와는 다른 계통이며, 난균류, 사카게카비류 등을 포함하고 농작물 피해를 유발하기도 한다. - 부등편모조류 - 역모균류
역모균류는 유주자의 앞쪽에 편모를 가지며 셀룰로스와 키틴으로 된 세포벽, 관상 크리스타를 갖는 미토콘드리아를 가지는 부등편모조류의 일종으로, 해수, 담수, 토양 등 다양한 환경에서 서식하며 다른 생물에 기생하기도 한다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 망형충류 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
 | |
| 학명 | Labyrinthulomycetes |
| 명명자 | Arx, 1970, Dick, 2001 |
| 이명 | Labyrinthulomycota Whittaker, 1969 Labyrinthomorpha Page in Levine et al., 1980 Labyrinthulea Olive, 1975 ex Cavalier-Smith, 1989 |
| 분류 | |
| 하위 계급 | 목 |
| 하위 분류 | 암피트레미다 암피필리다 오블롱기키트리다 라비린툴리다 트라우토키트리다 |
2. 특징
망형충류는 사제노겐 또는 보스로솜이라고 하는 독특한 세포 소기관을 통해 세포질과 연결되는 세포막으로 둘러싸인 필라멘트(filament)를 가진다. 세포는 단핵이며 타원형이고, 비정형 네트워크를 따라 분당 5~150 μm 속도로 움직인다. 망형충류는 세포가 튜브 내부에 갇혀 있고, 트라우스토키트리드는 튜브 측면에 부착되어 있다는 특징이 있다.[24]
망형충류는 균류가 아닌 스트라메노필에 속하는 진핵생물이다. 이들은 엽록체를 잃은 조류 공통 조상으로부터 기원했을 수 있으며, 오메가-3 지방산을 생성하고 유주자에 눈점 기구를 가지는 등 일부 특징은 조상 엽록체로부터 기원했을 가능성이 있다.
2. 1. 형태
망형충류의 필라멘트는 세포 외부에 위치하지만, 세포막으로 둘러싸여 있다. 이들은 사제노겐 또는 보스로솜이라고 하는 독특한 세포 소기관에 의해 형성되어 세포질과 연결된다. 세포는 단핵이며, 전형적으로 타원형이고, 비정형 네트워크를 따라 분당 5~150 μm의 속도로 앞뒤로 움직인다. 망형충류 중에서는 세포가 튜브 내부에 갇혀 있으며, 트라우스토키트리드 중에서는 튜브 측면에 부착되어 있다.[24][5][50][6]영양체(일반적인 상태의 체)는 기본적으로 단세포성이지만, 라비린툴라속은 특이한 망상 변형체양 군체를 형성한다(아래 그림 2; 아래 참조). 소로디프로프리스속의 영양체는 단세포성이지만, 포자 형성 시 세포가 모여 세포성 점균의 누적 자실체와 매우 유사한 자실체를 형성한다.[7][8]

세포는 일반적으로 얇은 (두께 2–3 nm) 판 모양의 '''인편'''이 겹쳐진 외피 구조로 둘러싸여 있다(위 그림 1b). 이 인편은 황산화 다당류(갈락토스, 자일로스, 푸코스 등에 풍부하며, 계통군에 따라 조성이 다르다)로 구성되어 있으며, 골지체에서 형성된다.[5][6][56] 일부 종은 생활환의 한 시기에 인편을 결여한 아메바 모양 세포(amoeboid cell)를 형성한다.[5] 예외적으로, 암피트레마과의 종은 양쪽 끝에 개구부를 갖는 두껍고 느슨한 껍질(로리카)을 형성한다(위 그림 1c).[34][9] 또한 라비린툴라류 중에는 세포외 다당류(exopolysaccharide)를 많이 분비하는 것도 있다.[5] 세포외 다당류는 포도당과 갈락토스를 많이 함유한다.[10]
세포의 한쪽 또는 여러 위치에서, '''외질망'''(외질망[54], 망상 구조[24]; ectoplasmic net, – network, – thread, – filament)이라고 불리는 구조가 생긴다.[3][4][11][5][6][38] 외질망은 종종 분지하는 가근 또는 실 모양 가짜 발과 같은 구조이며, 세포 외피가 없고, 생체막에 싸여 있지만 리보솜과 세포 소기관을 거의 포함하지 않는다.[3][50][6][13] 단, 내부에 종종 소포가 존재하고, 또한 액틴 섬유를 포함한다.[16] 외질망은 일반적으로 세포 표면에 존재하는 '''보스로솜'''(bothrosome; 외질망 형성체[54], 사게노제네토솜 sagenogenetosome, sagenetosome[50], 사게넷[12])이라고 불리는 세포 소기관에서 생긴다.[5][38][13][14] 보스로솜은 소포체가 밀집되어 있고 전자 밀도가 높은 물질이 부착되어 있다. 단 암피필라속 등에서는 외질망이 존재하지만, 보스로솜은 확인되지 않았다.[15] 외질망은 기질에 부착하여, 효소를 분비(세포외 소화)하고 영양물을 흡수한다고 생각된다.[5][6][56][13]
라비린툴라속에서는, 세포에서 생긴 외질망이 세포 전체를 감싸고, 더 나아가 다수의 세포의 외질망이 융합하여 망상 변형체와 같은 구조를 형성한다(위 그림 2).[3][4][5][6][16][56] 따라서 라비린툴라속은 net slime molds 또는 slime nets라고도 불리며,[16][12] 이러한 구조는 망상 변형체(net plasmodium, filoplasmodium)라고도 불렸다.[50] 라비린툴라속의 속명의 어원인 ''labyrinthus''는 라틴어로 "미로"를 의미하며, 이러한 망상 구조의 형태를 나타낸다.[54] 이 망상 구조에서는 다수의 세포가 공통 생체막으로 싸여 있지만, 각 세포는 보스로솜에 의해 구분된다. 각 세포는 이 공통 기질 중에서 액틴·미오신이 관여하는 활주 운동을 한다(100um/분에 달한다).[3][12] 아플라노키트리움속은 기본적으로 단세포성이지만, 라비린툴라속과 유사한 외질망에 따른 활주 운동 능력을 갖는다.[6] 또한 야브레츠보카비과 중에는 생활환의 한 시기에 운동 능력을 갖는 아메바 세포를 형성하는 것도 있다.[16]
세포는 기본적으로 단핵성(핵을 1개만 갖는)이며, 일반적으로 핵에 접하여 중심체가 존재한다.[5] 미토콘드리아의 크리스타는 관 모양.[5][16] 골지체가 존재하며, 또한 핵에 근접하여 활면 소포체에 둘러싸인 리보솜을 결여한 영역(paranuclear body)을 갖는 경우가 많다.[5] 세포질 중에는 종종 다량의 지질 과립이 존재하며, 큰 지질 과립이 세포의 대부분을 차지하는 경우도 있다(오른쪽 그림 3).[5][16] 지질 과립은 투명한 것부터, 카로테노이드를 포함하여 오렌지색에서 적색을 띠는 것까지 있다. 건조 중량당 총 지방산량은 8%에서 82%에 달한다.[5]
라비린툴라류는, 색소체와 관련된 세포 소기관을 갖지 않는다. 예외적으로, 암피트레마과의 종은 녹조의 클로렐라(트레보욱시아조강)를 세포 내 공생시키고 있다.[36]
2. 2. 생식
이분열, 유주자(편모를 가진 포자), 부동포자, 아메바 세포 형성 등에 의한 무성생식이 보고되었다.[3][4][5][16][11] (오른쪽 그림 4). 기본적으로 분열 단심성으로, 외질 네트 이외의 세포 전체가 포자낭이 되고, 세포 외피로 둘러싸여 포자를 형성, 세포 외피가 붕괴되면서 포자가 방출된다.[24][5][16] 포자 방출 후에 포자낭 내에 남은 부분 (원기체 proliferous body)이 성장하여, 다시 영양 세포가 되는 것도 알려져 있다.[11][17] 라비린툴라속에서는, 시스트 형성이 알려져 있다.[50]유주자는 세포 측면 (복면)에서 전후로 뻗어 있는 2개의 부등 편모를 가지며, 앞 편모에는 관상 소모가 부착되어 있다.[5][50] (위 그림 5). 라비린툴라속의 유주자는 안점을 가지며, 주광성을 나타내는 것으로 보고되었다.[24][18] 유주자는, 영양 세포와 마찬가지로 인편으로 덮여 있는 경우가 있다.[16] 많은 스트라메노필의 편모의 기부에 존재하는 나선 구조는 존재하지 않고, 대신 원추형의 구조가 보고되었다.[16][19]
솔로디프로플리스속은 육상 생물이며, 포자 형성 시 세포가 집합하여 세포성 점균에서 보이는 자실체와 매우 유사한 자실체 (누적자실체)를 형성한다.[7][8]
유성생식에 대해서는 밝혀지지 않았다.[16] 라비린툴라속의 유주자 형성 시 제1분열에서 시냅토넴 복합체가 관찰되었으며 (염색체 수는 ''n'' = 9), 이것이 감수 분열인 것으로 시사되었다.[50][20] 다만, 세포질 융합과 핵융합의 시기는 명확하지 않다. 한편, 야브레츠보카비류에서 편모 세포의 합체가 보고되었지만, 그 후의 경과에 대해서는 불명확하다.[21] (오른쪽 그림 4V). 또한 일부 종에서 휴면 포자 형성이 알려져 있지만, 이것이 배우자 합체의 결과인지 여부는 확실하지 않다.[6][13]
2. 3. 생태
라비린툴라류의 알려진 종 대부분은 바다에서 기수역에 걸쳐 생육하며, 고염분 환경에서도 보고되고 있다.[5][16] 열대에서 극지까지 분포하며, 연안역에 많지만, 외양의 심해역에서도 보고되고 있다.[5][6][16] 생물량도 많고, 세토 내해에서는 라비린툴라류의 생물량이 세균 생물량의 3.5–41%에 상당하는 것으로 보고되고 있다.[3] 수저 퇴적물, 맹그로브 숲, 하구 등 일반적으로 유기물이 풍부한 환경에 많으며, 부유 유기물에 부착된 플랑크톤성의 것도 있다.[16] 담수나 육상에 분포하고 있는 것도 있으며, 예를 들어 암피트레마과의 종은 고층 습원에,[34][22] 디플로프리스속은 일반적인 호소에,[22][23] 소로디플로프리스속은 토양에[7] 생육하고 있다. 환경 DNA에 기초한 연구로부터, 담수나 육상역에는 미지의 라비린툴라류가 많이 존재하고 있는 것이 시사되고 있다.[6] 또한 이러한 연구로부터, 지금까지 라비린툴라류의 보고가 없는 혐기적인 퇴적물 등의 환경에도 라비린툴라류가 생육하고 있는 것이 시사되고 있다.[16]기본적으로 흡수 영양 생물 (세포막을 통해 가용성 유기물을 흡수하여 이용하는 생물)이며, 동식물 유해 등 살아있지 않은 유기물을 세포외 소화, 흡수하는 부생성의 것이 많이 알려져 있다.[24][3][5][16][12] 이러한 라비린툴라류는, 특히 해양 생태계에 있어서 분해자로서 중요한 생물군으로 생각되고 있다.[16][25][26] 특히 세균 등에서는 분해가 어려운 육상 식물 유래의 유기물 (셀룰로스 등) 분해에 있어서, 라비린툴라류가 중요한 역할을 담당하고 있는 것이 시사되고 있다.[3][26] 게다가 라비린툴라류는 세균에 비해 대형이기 때문에, 부식물로부터 고차 포식자로 직접 에너지와 물질이 흐르는 경로가 된다고 생각되고 있다 (세균-세균 포식자의 단계를 생략한다).[3][26] 또한 라비린툴라류는, 사체뿐만 아니라 살아있는 미생물을 분해·흡수하는 일도 있다. 배양 하에서는, 세균이나 효모, 사상균, 규조, 사상조, 더 나아가 다른 라비린툴라류를 분해하는 것이 보고되고 있다.[16]
라비린툴라류는, 균주에 따라 아가라아제, 카라기나아제, 알긴산 리아제, 펙티나아제, 크실라나아제, 키티나아제, 셀룰라아제, 아밀라아제, α-글루코시다아제, 프로테아제, 젤라티나아제, 우레아아제, 리파아제, 포스파타아제 등의 분해 효소 활성을 가진다.[5] 또한, 유주자가 펙틴에 대한 주화성을 나타내는 것이 보고되고 있다.[3] 적어도 일부 종은, 매우 견고한 물질인 화분의 스포로폴레닌 분해능을 가진다.[50] 야브레츠보카비류를 야외에서 단리할 때에는, 일반적으로 화분을 먹이로 하는 방법이 이용되고 있다.[12]
라비린툴라류 중에는, 살아있는 생물에 공생하여 영양을 얻는 공생성 (기생성 포함)인 것도 많다고 생각되고 있다.[24][6] 다양한 해조류나 해초의 표면이나 내부에서 라비린툴라류가 발견되고 있으며, 이것들은 공생성 (상리 공생, 편리 공생, 또는 기생)일 가능성이 있다.[16] 라비린툴라속 중에는, 해초의 잘피에 기생하는 종이나, 육생 식물인 잔디에 기생하는 종도 알려져 있다.[27][28] 또한 규조의 세포 내에 외질 네트워크 또는 본체를 침입시키는 기생성의 종도 보고되고 있으며, 이러한 라비린툴라류가 해양에 널리 생육하고 있는 것이 시사되고 있다.[29] 라비린툴라속의 1종이, Arbuscular 균근을 형성하는 균류인 글로무스류의 포자에 기생하고 있는 예도 보고되고 있다.[30] 그 외에도 아메바류에 세포 내 공생하는 예나,[31] 해면, 산호, 성게, 멍게, 무지개송어, 편형동물, 다양한 연체동물 (문어나 오징어, 갯민숭달팽이류, 전복, 이매패류)에 공생 (기생)하고 있는 예가 알려져 있다.[5][16][32][33] 다만 대형 생물에 대한 공생성으로 여겨지는 예 중에는, 실제로는 거기에 부착되어 있는 미생물을 분해·흡수하고 있는 일도 있다.[16]
적어도 일부 종에서는, 세균 등을 섭취하는 식작용능을 가진다는 것이 시사되고 있다.[34][35]
라비린툴라류는 광합성 능력을 가지지 않지만, 암피트레마과의 생물은 녹조류의 클로렐라 (트레보욱시아 조강)를 세포 내 공생시키고 있으며, 아마 공생 조류의 광합성에 의한 유기물을 얻고 있다고 생각되고 있다.[36][37]
라비린툴라류에 기생하는 바이러스 (1가닥 RNA 바이러스, 2가닥 DNA 바이러스)가 발견되고 있다.[3][38] 라비린툴라류는 해양 생태계에 있어서 분해자로서 중요한 요소로 생각되고 있지만, 그 증감에 바이러스가 크게 영향을 미치고 있을 가능성이 있다. 또한 공생성의 라비린툴라류의 경우, 라비린툴라류가 숙주에 대한 바이러스의 매개자가 되고 있을 가능성도 있다.[16]
3. 인간과의 관계
1930년대에 북대서양 양안에서 거머리말 군락이 대규모로 감소하여 어패류 등 수산 자원에 큰 영향을 미쳤다. 이 거머리말 쇠약병의 원인은 라비린툴라속의 1종(Labyrinthula zosterae|라비린툴라 조스테레la)이 기생했기 때문으로 여겨지지만, 그 자세한 관계는 밝혀지지 않았다.[3][50][16][12][39][40] 또한 라비린툴라속의 다른 1종(Labyrinthula terrestris|라비린툴라 테레스트리스la)은 잔디(갯새풀, 가는털수염],
4. 계통 분류
망형충류는 한때 멸종된 균류 문인 Labyrinthulomycota를 구성했다. 이들은 원래 특이한 점균류로 여겨졌지만, 다른 종류와는 크게 유사하지 않다. 유주자의 구조와 유전학적 연구를 통해 이들이 헤테로콘트의 원시적인 집단임이 밝혀졌지만, 분류 및 처리는 다소 불안정하게 남아있다.
미로충류는 균류가 아니라 스트라메노필 내의 단계통군 진핵생물 그룹이다. Bigyra문에 속하며, bicosoecids와 같은 다른 종속 영양 미생물을 포함한다. Stramenopiles의 엽록체가 그들의 최근 공통 조상에서 일어난 내공생 사건의 결과일 수 있다는 점을 고려할 때, bicosoecids와 미로충류는 혼합 영양 조류 공통 조상으로부터 기원했을 수 있으며, 2차 손실로 엽록체를 잃었다.
미로충류의 일부 특징은 조상 엽록체로부터의 기원으로 설명될 수 있다. 일반적으로 엽록체에 존재하는 탈포화 효소를 사용하여 오메가-3 지방산을 생성한다. 미로충의 유주자는 눈점 기구를 가지고 있는데, 이는 광합성 스트라메노필의 눈점과 유사한 막으로 둘러싸인 과립으로 구성되어 있으며, 엽록체 내에 있거나 엽록체에서 파생된 것으로 여겨진다.
Bigyra 내에서, 미로충류는 ''Pseudophyllomitus vesiculosus'' 종과 MAST-4라고 불리는 환경 DNA 계통을 포함하는 Eogyrea의 자매군이다. 이들은 함께 아문 Sagenista를 구성한다.
이 강에는 일반적으로 Labyrinthulales와 Thraustochytriales (ICBN) 또는 Labyrinthulida와 Thraustochytrida (ICZN)의 두 목이 포함되었지만, 최근 다른 분류법이 제안되었다.
라비린추라속은 망상 변형체와 같은 구조를 형성하며, 세포는 그 안에서 활주 운동을 한다. (상기 참조) 이러한 특징으로 인해 점균류 (광의의 변형균) 또는 육질충 (광의의 아메바류)으로 분류되었다.[49][50][11][51][52][53]。 그 특이성으로 인해 원생동물이나 원생생물의 독립된 문 (라비린추라문)으로 분류되기도 했다.[54][51][78]。한편, 야브레츠보카비류는 털곰팡이류와 유사한 체(가근과 같은 구조가 생긴 구형 세포)를 가지면서도, 난균과 유사한 편모 세포를 형성하기 때문에 난균강으로 분류되었다.[5][16][55]。
1970년대 이후 전자 현미경을 이용한 미세 구조학적 연구를 통해 라비린추라속과 야브레츠보카비류가 다양한 특징(비늘이 겹쳐진 외피, 보스토솜에서 생기는 외질 네트 등)을 공유하고, 스트라메노필 (이형털류)의 특징(관상 소모를 갖는 전편모 등)을 가지고 있음이 밝혀졌다. 따라서 라비린추라속과 야브레츠보카비류가 스트라메노필 내에서 하나의 계통군을 구성한다고 생각하게 되었다.[16][56][57][58]。 20세기 말 이후의 분자 계통학적 연구에서도 이러한 생각은 지지받고 있다.[59][60]。 미세 구조 및 분자 계통학적 연구를 통해, 사상 가족을 갖는 아메바류(사상 가조족)로 취급되던 몇몇 원생생물(디플로프리스속, 암피트레마과 등)도 라비린추라류에 속하는 것으로 나타났다.[34][61][62]。
2020년 현재, 이러한 생물을 함께 묶어 '''라비린추라강'''[63] (라비린추라강[64] ;라비린추라균강[11], 라비린추라균강[65] )으로 분류하는 것이 일반적이다. 독립된 문(라비린추라문[63], 라비린추라문[64] [16];라비린추라균문[11], 라비린추라균문[65] [16][76])으로 취급되는 경우도 있다. 한편, 다수의 유전자 염기 서열에 기반한 계통 분석에서는 포식성 편모충인 Pseudophyllomitusla가 라비린추라류와 근연 관계에 있다는 점이 시사되어, 양자를 함께 사게니스타문 또는 아문[66] (Sagenistala)으로 분류하기도 한다[67]。 환경 DNA 연구에서는 실체가 불분명한 몇몇 계통군(MAST-4, 7, 8, 9, 10, 11, 20)이 라비린추라류와 근연 관계에 있음을 시사한다[69]。 스트라메노필 안에는 균류적인 흡수 영양 생물인 그룹으로서 다른 난균류가 존재하지만, 라비린추라류와 난균류는 근연 관계가 아니다.[16][59][60]。
스트라메노필 중에서 이형털조류는 홍조류 유래의 색소체를 가지지만, 라비린추라류와 난균, 오파리나, 비코소에카류 등 그 외의 그룹은 모두 종속영양성이며 색소체를 갖지 않는다. 이형털조류가 갖는 색소체의 기원은 크립토조류나 햅토조류가 갖는 색소체의 기원과 동일하다는 가설이 있으며(크로미스타 가설), 이 가설에서는 라비린추라류의 조상도 홍조류 유래의 색소체를 가지고 있었지만 이를 이차적으로 잃었다고 한다[66]。 그러나 2020년 현재, 라비린추라류가 색소체를 가지고 있었다는 것을 명확히 지지하는 증거는 얻어지지 않았으며, 라비린추라류가 광합성 생물에서 유래했는지 여부는 불분명하다[16]。
라비린추라강 안에는 라비린추라목과 야브레츠보카비목이 인식되고 있다[24][3][59]。 야브레츠보카비류의 몇몇 속(Schizochytriumla, Ulkeniala 등)에 대해 분류학적 재편성이 이루어졌고[11][71][72][70], 그 과정에서 야브레츠보카비류는 단일 계통군이 아니라는 점이 시사되었으며, 일부 속(Aplanochytriumla, Oblongichytriumla)은 야브레츠보카비목에서 제외되었다.[16]。 사상 가조를 갖는 아메바류로 취급되던 몇몇 생물이 라비린추라강을 구성하는 다른 계통군(암피트레마목, 암피필라목)임이 시사되고 있다.[34][62]。 이러한 라비린추라강 내의 분류는 분자계통학적 분석이나 생활환 패턴, 미세 구조, 고도 불포화 지방산 및 카로티노이드의 조성 등에 기반하여 이루어진다[3][5][71][72][73]。 환경 DNA 연구를 통해 라비린추라류 안에 실체가 불분명한 계통군이 다수 존재함이 시사되고 있다[16][74]。
'''라비린추라강 내의 계통 가설의 예'''[16][34][62][73][77][75]
{| class="wikitable" style="text-align: center;"
|-
! '''라비린추라강'''
|-
|
{| class="wikitable" style="text-align: center;"
|-
! "오블롱기키트리움목"
|-
| '''오블롱기키트리움과''': Oblongichytriumla
|-
| Stellarchytriumla
|-
! '''라비린추라목'''
|-
|
| 아플라노키트리움과: Aplanochytriumla |
| 라비린추라과: Labyrinthulala |
|-
! '''야브레츠보카비목'''
|-
| '''야브레츠보카비과''': Aurantiochytriumla, Schizochytriumla, Thraustochytriumla, Ulkeniala etc.
|-
! '''암피트레마목'''
|-
|
| 디플로프리스과: Diplophrysla |
| 암피트레마과: Amphitremala, Archerellala |
|-
! '''암피필라목'''
|-
|
| 암피필라과: Amphifilala, Fibrophrysla |
| 소로디플로프리스과: Sorodiplophrysla |
|}
|}
4. 1. 계통
다음은 부등편모조류의 계통 분류이다.[80][81][82]{| class="wikitable" style="text-align: center;"
|-
! 부등편모조류
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 플라티술쿠스강
|-
|
{| class="wikitable" style="text-align: center;"
|-
! 사게니스타류
|-
|
| Eogyrea |
| 망형충류 |
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 비코소에카류
|-
|
{| class="wikitable" style="text-align: center;"
|-
! 플라시디아류
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 플라시디아강
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 나노모나스강
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 오팔로모나스강
|-
! 오팔리나상강
|-
|
|}
|}
|}
|-
! 기리스타류
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 비기로모나드강
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 난균류
|-
|
{| class="wikitable" style="text-align: center;"
|-
| 역모균류
|-
|
|}
|}
|}
|}
|}
|}
|}
미로충류는 균류가 아니라 Stramenopiles 내의 단계통군 진핵생물 그룹이다. 이들은 Bigyra문에 속하며, bicosoecids와 같은 다른 종속 영양 미생물을 포함한다. Stramenopiles의 엽록체가 그들의 최근 공통 조상에서 일어난 내공생 사건의 결과일 수 있다는 점을 고려할 때, bicosoecids와 미로충류는 혼합 영양 조류 공통 조상으로부터 기원했을 수 있으며, 2차 손실로 엽록체를 잃었다.
미로충류의 일부 특징은 조상 엽록체로부터의 기원으로 설명될 수 있다. 이들은 일반적으로 엽록체에 존재하는 탈포화 효소를 사용하여 오메가-3 지방산을 생성한다. 미로충의 유주자는 눈점 기구를 가지고 있는데, 이는 광합성 스트라메노필의 눈점과 유사한 막으로 둘러싸인 과립으로 구성되어 있으며, 엽록체 내에 있거나 엽록체에서 파생된 것으로 여겨진다.
Bigyra 내에서, 미로충류는 ''Pseudophyllomitus vesiculosus'' 종과 MAST-4라고 불리는 환경 DNA 계통을 포함하는 Eogyrea의 자매군이다. 이들은 함께 아문 Sagenista를 구성한다.
{| class="wikitable" style="text-align: center; font-size:90%;"
|-
! Stramenopiles
|-
|
{| class="wikitable" style="text-align: center;"
|-
| Platysulcea
|-
|
{| class="wikitable" style="text-align: center; font-size: smaller;"
|-
| ''plastid loss''
|-
! Bigyra
|-
|
{| class="wikitable" style="text-align: center;"
|-
! Opalozoa
|-
|
Placidozoa 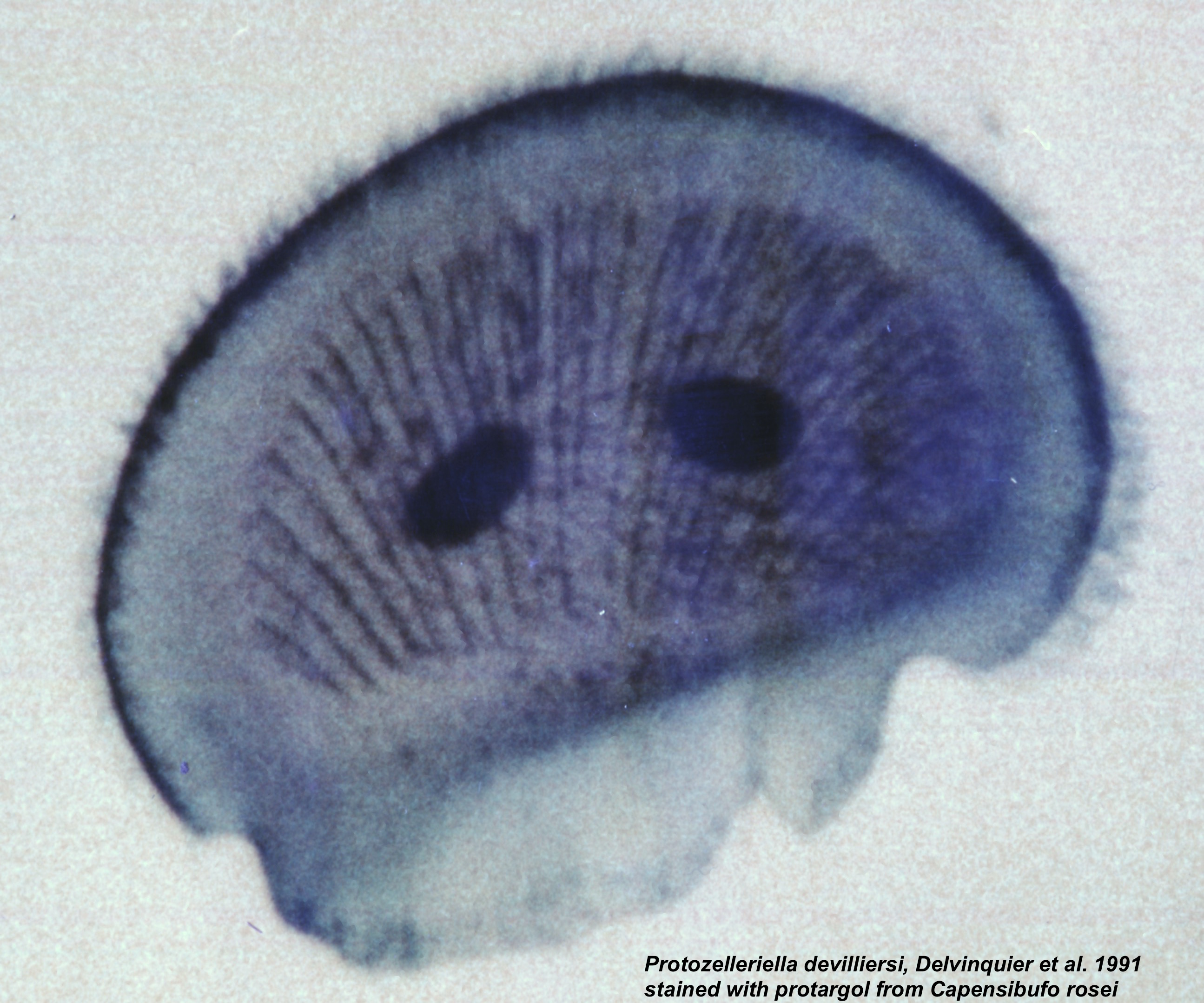 |
Bicosoecida 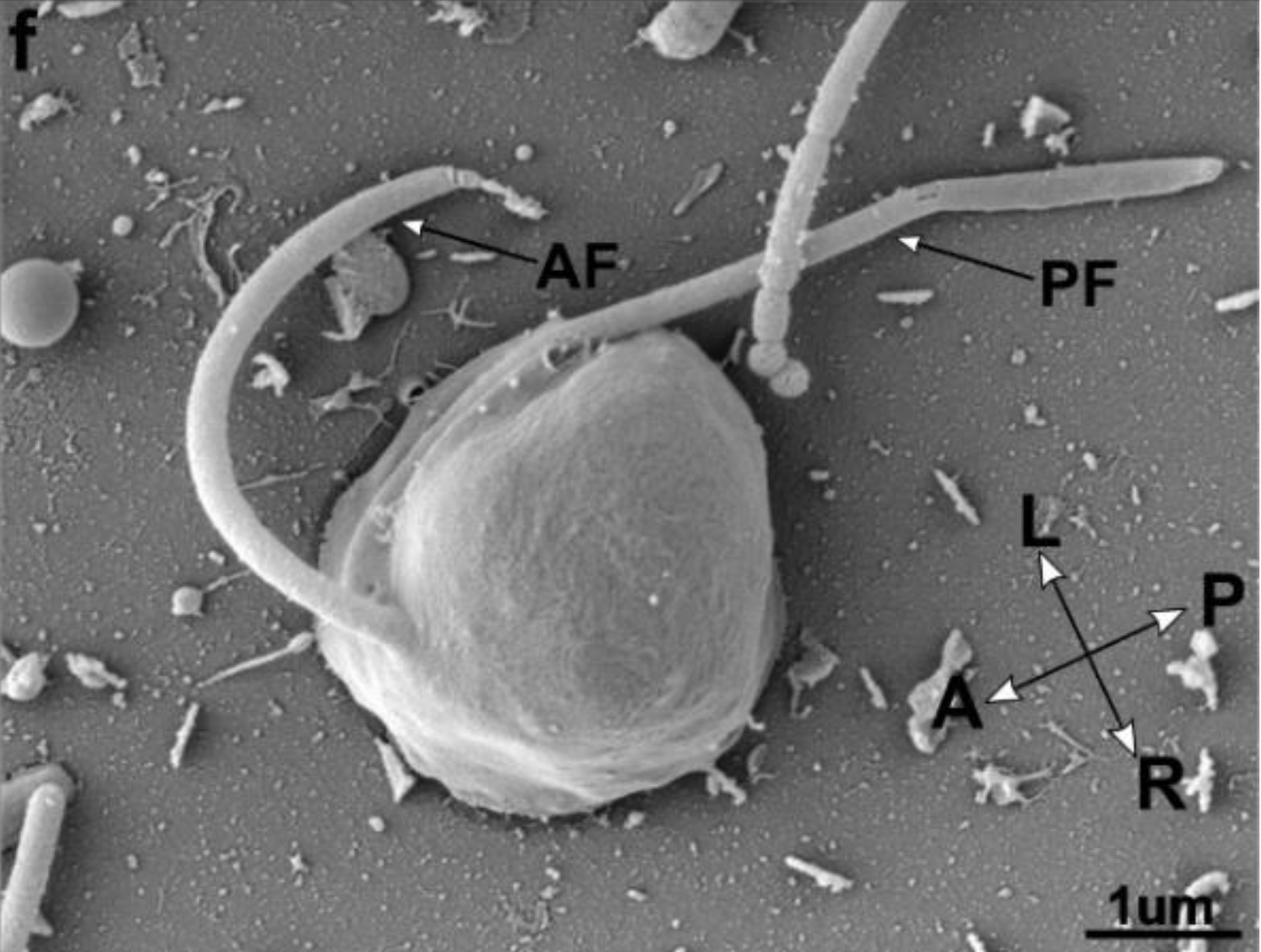 |
|-
! Sagenista
|-
|
| Labyrinthulomycetes |
| Eogyrea |
|}
|-
! Gyrista
|-
|
Ochrophyta  |
| plastid loss |
Pseudofungi 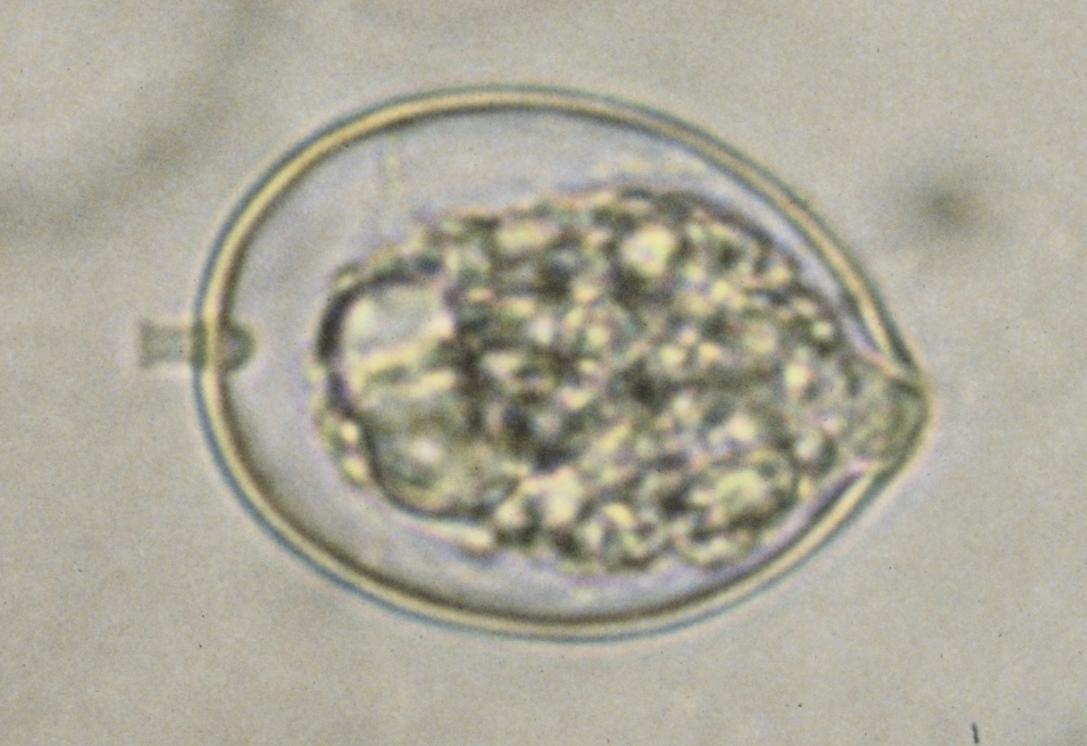 |
|}
|}
|}
4. 2. 분류
망형충류는 오랫동안 균류의 일종으로 알려졌으나, 유주자의 구조와 유전학적 연구를 통해 헤테로콘트의 원시적인 집단으로 밝혀졌다.다음은 부등편모조류의 계통 분류이다.[80][81][82]
{| class="wikitable"
|-
! 계통 분류
|-
|
{| class="wikitable"
|-
| 부등편모조류
{| class="wikitable"
|-
| 플라티술쿠스강
|-
|
{| class="wikitable"
|-
| 사게니스타류
| Eogyrea |
| 망형충류 |
|-
| 비코소에카류
|-
|
{| class="wikitable"
|-
| 플라시디아류
{| class="wikitable"
|-
| 플라시디아강
|-
|
{| class="wikitable"
|-
| 나노모나스강
|-
|
{| class="wikitable"
|-
| 오팔로모나스강
|-
| 오팔리나상강
|}
|}
|}
|-
| 기리스타류
{| class="wikitable"
|-
| 비기로모나드강
|-
|
{| class="wikitable"
|-
| 난균류
|-
|
{| class="wikitable"
|-
| 역모균류
|-
|
|}
|}
|}
|}
|}
|}
망형충류의 과(Family)는 다음과 같다.
- Labyrinthulaceae
- * ''Labyrinthula''
- Thraustochytriaceae
- * ''Aplanochytrium''
- * ''Labyrinthuloides''
- * ''Japonochytrium''
- * ''Schizochytrium''
- * ''Thraustochytrium''
- * ''Ulkenia''
- * ''Diplophryids''
- * ''Diplophrys''
참조
[1]
논문
Ueber den Bau und die Entwicklung der Labyrinthuleen
https://www.biodiver[...]
1867
[2]
웹사이트
ラビリンスラ
https://kotobank.jp/[...]
2020-11-11
[3]
서적
藻類ハンドブック
株式会社エヌ・ティー・エス
[4]
서적
岩波 生物学辞典 第5版
岩波書店
[5]
논문
Taxonomy, ecology and biotechnological applications of thraustochytrids: A review
[6]
서적
THE MYCOTA, volume 7A. Systematics and Evolution Part A
Springer
[7]
논문
"Sorodiplophrys: an unusual sorocarp-producing protist"
[8]
논문
"Sorodiplophrys stercorea: another novel lineage of sorocarpic multicellularity"
[9]
서적
An Illustrated Guide to the Protozoa, Organisms Traditionally Referred to as Protozoa, or Newly Discovered Groups
Blackwell Publishers
[10]
논문
Extracellular polysaccharide production by thraustochytrid protists
[11]
서적
菌類の事典
朝倉書店
[12]
서적
Introductory Mycology
John Wiley & Sons. Inc., New York
[13]
서적
Handbook of Protoctista
Jones and Bartlett
[14]
논문
An ultrastructural study of taxonomically significant characters of the Thraustochytriales and Labyrinthulales
[15]
논문
"Diplophrys marina, a new scale-forming marine protist with labyrinthulid affinities"
[16]
서적
Handbook of the Protists
Springer
[17]
웹사이트
Laby Base
http://syst.bio.kona[...]
甲南大学
2020-11-05
[18]
논문
Zoosporulation in Labyrinthula sp.; an electron microscope study
[19]
논문
A comparison of the flagellar apparatus in Phytophthora, Saprolegnia, Thraustochytrium, and Rhizidiomyces
[20]
논문
Chromosome number of a small protist: accurate determination
[21]
논문
Genomics, biology and phylogeny Aurantiochytrium acetophilum sp. nov.(Thraustrochytriaceae), including first evidence of sexual reproduction
[22]
서적
淡水微生物図鑑 原生生物ビジュアルガイドブック
誠文堂新光社
[23]
서적
プランクトンハンドブック 淡水編
文一総合出版
[24]
서적
バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統
裳華房
[25]
서적
Biology of Marine Fungi. Prog. Mol. Subcell. Biol. 53.
Springer
[26]
논문
海洋環境におけるラビリンチュラ類の生態学的役割
[27]
논문
"Labyrinthula terrestris sp. nov., a new pathogen of turf grass"
[28]
논문
Genetic diversity in Labyrinthula terrestris, a newly emergent plant pathogen, and the discovery of new Labyrinthulid organism
[29]
논문
A widely distributed thraustochytrid parasite of diatoms isolated from the Arctic represents a gen. and sp. nov.
[30]
논문
Labyrinthula inside the spores of a vesicular—arbuscular mycorrhizal fungus
[31]
논문
Living together: The marine amoeba Thecamoeba hilla Schaeffer, 1926 and its endosymbiont Labyrinthula sp
[32]
논문
Some ultrastructural observations of a thraustochytrid (Protoctista, Labyrinthulomycota) from the clam Ruditapes decussatus (Mollusca, Bivalvia)
[33]
논문
Ultrastructural observations on a thraustochytrid fungus parasitic in the gills of squid (Illex illecebrosus Lesueur)
[34]
논문
Amphitremida (Poche, 1913) is a new major, ubiquitous labyrinthulomycete clade
[35]
논문
Bacterivory: a novel dual role for thraustochytrids in the sea
[36]
서적
Algal and Cyanobacteria Symbioses
[37]
논문
One alga to rule them all: unrelated mixotrophic testate amoebae (Amoebozoa, Rhizaria and Stramenopiles) share the same symbiont (Trebouxiophyceae)
[38]
논문
ウイルス研究から見た海産原生生物ラビリンチュラ類の生態学的多様性
[39]
논문
"Labyrinthula zosterae" sp. nov., the causative agent of wasting disease of eelgrass, "Zostera marina"
[40]
논문
Current European "Labyrinthula zosterae" are not virulent and modulate seagrass ("Zostera marina") defense gene expression
[41]
논문
SNP hot-spots in the clam parasite QPX
[42]
논문
Erection of a new genus and species for the pathogen of hard clams ‘Quahog Parasite Unknown’(QPX): "Mucochytrium quahogii" gen. nov., sp. nov.
[43]
논문
Mechanism of infection by "Labyrinthuloides haliotidis" (Protozoa, Labyrinthomorpha), a parasite of abalone ("Haliotis kamtschatka") (Mollusca, Gastropoda)
[44]
논문
Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids
[45]
논문
Recent developments in the commercial production of DHA and EPA rich oils from micro-algae
[46]
웹사이트
なぜlife's製品なのか?
https://www.lifesdha[...]
2020-08-15
[47]
논문
Thraustochytrid as a potential source of carotenoids
[48]
논문
Thraustochytrid "Aurantiochytrium" sp. 18W-13a accummulates high amounts of squalene
[49]
서적
バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統
裳華房
[50]
서적
ウェブスター菌類概論
講談社サイエンティフィク
[51]
서적
バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統
裳華房
[52]
서적
植物系統分類の基礎
北隆館
[53]
웹사이트
A Preliminary Catalogue of the Names of Fungi above the Rank of Order
https://ucjeps.berke[...]
Constancea, University and Jepson Herbaria
2020-10-18
[54]
서적
バイオディバーシティ・シリーズ (5) 無脊椎動物の多様性と系統
裳華房
[55]
서적
植物系統分類の基礎
北隆館
[56]
서적
The Biology of Marine Fungi
Cambridge University Press
[57]
논문
The ultrastructure of holdfasts,“rhizoids”, and “slime tracks” in thraustochytriaceous fungi and "Labyrinthula" spp
[58]
논문
Taxonomic revisions in the marine biflagellate fungi: the ultrastructural evidence
[59]
논문
Molecular phylogeny of labyrinthulids and thraustochytrids based on the sequencing of 18S RNA gene
[60]
논문
Labyrinthulomycetes phylogeny and its implications for the evolutionary loss of chloroplasts and gain of ectoplasmic gliding
[61]
서적
The Chromophyte Algae, Problems and Perspectives
Clarendon Press
[62]
논문
Ultrastructure of "Diplophrys parva", a new small freshwater species, and a revised analysis of Labyrinthulea (Heterokonta)
[63]
서적
岩波 生物学辞典 第5版
岩波書店
[64]
서적
バイオディバーシティ・シリーズ (5) 無脊椎動物の多様性と系統
裳華房
[65]
서적
バイオディバーシティ・シリーズ (4) 菌類・細菌・ウイルスの多様性と系統
裳華房
[66]
논문
Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences
[67]
논문
Taxon-rich multigene phylogenetic analyses resolve the phylogenetic relationship among deep-branching stramenopiles
[68]
논문
Sagenista and Bigyra, two phyla of heterotrophic heterokont chromists
[69]
논문
Revisions to the classification, nomenclature, and diversity of eukaryotes
[70]
논문
Proposal of "Monorhizochytrium globosum" gen. nov., comb. nov.(Stramenopiles, Labyrinthulomycetes) for former "Thraustochytrium globosum" based on morphological features and phylogenetic relationships
[71]
논문
Taxonomic rearrangement of the genus ''Schizochytrium sensu lato'' based on morphology, chemotaxonomic characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): emendation for ''Schizochytrium'' and erection of ''Aurantiochytrium'' and ''Oblongichytrium'' gen. nov
[72]
논문
Taxonomic rearrangement of the genus ''Ulkenia sensu lato'' based on morphology, chemotaxonomical characteristics, and 18S rRNA gene phylogeny (Thraustochytriaceae, Labyrinthulomycetes): Emendation for ''Ulkenia'' and erection of ''Botryochytrium'', ''Parietichytrium'', ''Sicyoidochytrium'' gen. nov.
[73]
논문
''Fibrophrys columna'' gen. nov., sp. nov: A member of the family Amphifilidae
[74]
논문
Reference tree and environmental sequence diversity of Labyrinthulomycetes
[75]
논문
Characterization of three novel species of Labyrinthulomycota isolated from ochre sea stars (''Pisaster ochraceus'')
[76]
웹사이트
The MycoBank engine and related databases
https://www.mycobank[...]
Robert, V., Stegehuis, G. & Stalpers, J.
2020-10-25
[77]
논문
Outline of Fungi and fungus-like taxa
[78]
논문
A Newly Revised Classification of the Protozoa
[79]
서적
Syllabus of Plant Families. A. Engler's Syllabus der Pflanzenfamilien Part 1/1
Borntraeger
[80]
논문
Higher Level Classification of All Living Organisms
[81]
논문
Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes
https://hal.archives[...]
[82]
논문
Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes
2013
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com