메기목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
메기목은 골단어상목에 속하는 어류 분류군으로, 형태학적 다양성에도 불구하고 분자적 증거에 의해 단계통군을 형성하는 것으로 밝혀졌다. 약 36개의 과에 3,093종이 있으며, 척추동물 목 중 두 번째 또는 세 번째로 다양한 분류군이다. 메기는 남아메리카를 중심으로 곤드와나 기원을 가지며, 분류 체계는 지속적으로 변화하고 있다. 메기목 어류는 식용, 관상용, 낚시 대상으로 이용되며, 일부 종은 유해종으로 생태계에 문제를 일으키기도 한다. 또한 일본 신화에서는 지진을 일으키는 존재로 묘사되기도 한다.
더 읽어볼만한 페이지
- 메기목 - 네마토게니스 이네르게니스
네마토게니스 이네르게니스는 메기목의 계통 분류를 다루며, 로리카리아아목, 메기아목 등 하위 분류와 각 분류군에 속하는 과들을 상세히 제시한다. - 메기목 - 동자개
동자개는 메기목 동자개과에 속하는 민물고기로, 울음소리 때문에 '빠가사리'라고도 불리며 한국을 포함한 동아시아에 분포하고 식용 및 관상어로 이용되지만 일본에서는 외래 생물로 지정되었다. - 솔 푸드 - 프라이드 치킨
프라이드 치킨은 스코틀랜드와 서아프리카 요리의 영향을 받아 미국 남부에서 발전한 닭고기 튀김 요리로, 아프리카계 미국인 가정부들을 통해 대중화되었으며, 패스트푸드 산업 발전과 함께 전 세계로 확산되어 다양한 스타일과 변형 요리가 존재한다. - 솔 푸드 - 우스터셔 소스
우스터셔 소스는 19세기 영국에서 개발되어 상업화된 소스로, 몰트 식초, 주정 식초, 당밀, 설탕, 소금, 멸치, 타마린드 등을 주재료로 하여 독특한 풍미를 내며, 전 세계적으로 다양한 변형이 존재하고 일본에서는 독자적인 소스 문화가 발달했다.
| 메기목 - [생물]에 관한 문서 | |
|---|---|
| 분류 | |
| 학문적 분류 | 조르주 퀴비에, 1817 |
| 유형 종 | 메기 |
| 유형 종 권위자 | 칼 폰 린네, 1758 |
| 특징 | |
 | |
| 이미지 설명 | 검은돌머리 |
| 이미지 크기 | 1.1 |
| 분포 및 생태 | |
| 서식지 | 담수 |
| 특징 | 수염, 야행성 |
| 계통 분류 | |
| 상위 분류군 | 골표상목 |
| 하위 분류 | |
| 현존 과 | 아이릴리과 아키시스과 암블리키피티스과 암필리우스과 안카리아과 아리우스과 아스프레디니스과 아스트로블레푸스과 아우케니프테루스과 아우스트로글라니스과 바그리다이과 칼리크티스과 케톱시스과 차카과 클라리아스과 클라로테스과 크라노글라니스과 말라프테루루스과 모코쿠스과 네마토게니스과 팡가시우스과 피멜로두스과 플로토수스과 슈도피멜로두스과 실베이다이과 스콜로플락스과 실루루스과 시소리다이과 트리코미크테루스과 incertae sedis: 코노린초스 |
| 멸종 과 | 안디니크티스과† 바흐만니아과† 힙시도리스과† |
| 학명 | |
| 학명 | Siluriformes |
| 로마자 표기 | Sillureumi |
| 어원 | |
| 어원 | Silurus (메기 속) + forma (모양) |
| 화석 기록 | |
| 화석 범위 | 캄파니아기 ~ 현재 |
2. 분류
분자적 증거에 따르면, 모든 메기는 단계통군을 형성한다.[7] 메기는 골표상목에 속하며, 잉어목, 카라신목, 곤어린쿠스목, 김노투스목을 포함한다. 김노투스목을 메기목의 하목으로 분류하기도 하지만, 널리 받아들여지지는 않는다. 현재 메기목은 김노투스목의 자매군으로 여겨지나, 최근 분자적 증거로 인해 논쟁이 있다.[8]
약 36개의 현존하는 메기 과와 약 3,093종의 현존하는 종이 기술되었다.[9] 이는 메기목을 두 번째 또는 세 번째로 다양한 척추동물 목으로 만든다. 척추동물 20마리 중 1마리가 메기이다.[10]
메기는 남아메리카를 중심으로 한 곤드와나 기원으로 여겨진다. 가장 초기의 확실한 구성원은 백악기 후기 캄파니안에서 마스트리히트절 시기에 아메리카 대륙에 살았던 안디니크티스과, ''Vorhisia vulpes'', ''아리우스'' 등이다.[1][11][13]
메기 분류는 빠르게 변화하고 있다. 2007년과 2008년 논문에서 ''Horabagrus'', ''Phreatobius'', ''Conorhynchos''는 현재 메기 과에 분류되지 않았다.[9] 2005년 6월, 연구자들은 Lacantuniidae라는 새로운 메기 과를 명명했는데, 이는 지난 70년 동안 구별된 세 번째 새로운 물고기 과였다. Lacantuniidae의 새로운 종인 ''Lacantunia enigmatica''는 멕시코 치아파스 주의 라칸툰 강에서 발견되었다.[21]
메기목의 상위 계통발생은 주로 분자 계통발생 연구로 인해 최근 여러 차례 변화를 겪었다. 대부분의 연구는 메기가 Diplomystidae 과, Loricarioidea, Siluroidei의 세 주요 계통으로 배열된다는 데 동의하지만, 이들 계통 간의 관계는 논쟁적이었다.[22][23][24][25][26] 형태학적 데이터에 따르면, Diplomystidae는 일반적으로 가장 초기에 분기되는 메기 계통으로 간주되며, Loricarioidei와 Siluroidei의 자매군이다.[25][26][27] 반면 분자적 증거는 일반적으로 Loricarioidei가 가장 초기에 분기되는 메기 계통이며, Diplomystidae와 Siluroidei를 포함하는 분기도의 자매로 나타난다.[7][22][23][28]
다음은 서로 다른 저자에 의한 과 관계 목록이다. Lacantuniidae는 Claroteidae의 자매로 위치하는 최근 증거를 기반으로 Sullivan 체계에 포함되어 있다.[30]
2. 1. 개요
메기목은 2006년 기준으로 2,800종 이상이 기록되어 있으며, 어류의 목 중에서 농어목(약 1만 종), 잉어목(약 3,200종) 다음으로 세 번째로 큰 무리이다. 현재 존재하는 어류 2만 8000종의 약 10%, 담수어종(1만 2400종)만 보면 그 20%가 메기목 어종이다.[90] 유속이 느린 하천, 호수에서 동굴, 산악 지대의 급류까지, 전 세계의 모든 육수에 널리 분포하며, 하구, 기수역 및 연안 부근에서 사는 해산종도 포함된다. 1994년(약 2,400종)부터 10여 년 만에 400종 이상이 새로 기재되는 등, 분류가 확대되는 경향이 계속되고 있다.일반적인 메기류의 공통적인 형태학적 특징은 납작하고 찌그러진 큰 머리와 넓은 입, 그리고 감각 기관으로 발달한 긴 수염이다. 세계 여러 지역에서 예로부터 중요한 어업 자원으로 이용되었고, 양식도 활발하게 이루어지고 있다. 최근에는 중・대형종이 취미나 스포츠로서의 낚시 대상이 되는 것 외에, 코리도라스, 시노돈티스, 플레코 등 관상어로 친숙한 종류도 많아, 세계 각지의 수족관 및 개인 수조에서 사육되고 있다. 한편, 이식된 외래 메기가 고유 생태계에 영향을 미치는 등의 문제도 최근 각지에서 발생하고 있다.
가장 오래된 메기목 어류 화석은 백악기 후기(약 6500만 년 전) 지층에서 발견되었다. 화석종은 오스트레일리아 대륙을 제외한 6개 대륙에서 발견되었으며, 남극 대륙에서는 시신세부터 올리고세(5500만~2400만 년 전) 지층에서 보고되었다.
일본에서는 고비와호층군(현재 미에현우에노 분지 부근)의 3~400만 년 전 지층에서 메기과 어류 화석이 발견되었다. 이보다 훨씬 이전, 가가와현 사누키층군의 신제의 지층 (약 1500만 년 전)에서는 세계에서 가장 오래된 메기과 어류 화석이 발견되고 있다.
가장 오래된 화석이 남아메리카에서 출토되었고, 원시적인 특징을 남기는 디플로뮤스테스과나 케토프시스과 무리가 남아메리카에 분포하는 점 등으로 인해, 메기목의 기원은 남미로 여겨지는 경우가 많다. 메기목의 자매군인 전기뱀장어목이 남아메리카에만 서식하는 것도 남미 기원설의 유력한 증거로 여겨진다.
2. 2. 과 목록
| 과 | 설명 |
|---|---|
| 아키시스과 (Akysidae) | 동남아시아에 분포하며, 등지느러미 가시가 강하다. |
| 퉁가리과 (Amblycipitidae) | 남아시아에서 동아시아(일본 포함) 하천에 분포한다. 참가자치는 일본 고유종이다. |
| 암필리우스과 (Amphiliidae) | 아프리카 열대 지방에 널리 분포하는 소형 어종으로, 급류를 거슬러 바위를 오를 수 있다. |
| 앙카리우스과 (Anchariidae) | |
| † Andinichthyidae | |
| 바다동자개과 (Ariidae) | |
| 밴조메기과 (Aspredinidae) | 남아메리카에 분포하며, 머리가 심하게 종편되어 밴조와 유사한 형태를 띤다. |
| 아스트로블렙푸스과 (Astroblepidae) | 파나마와 남아메리카에 분포하며, 산악 지대 급류에 적응하여 입에 원형 흡반을 가지고 있다. |
| 아우케닙테루스과 (Auchenipteridae) | 파나마 및 남아메리카 열대 지역에 분포하며, 수컷은 교접기를 가지고 체내 수정을 한다. |
| 아우스트로글라니스과 (Austroglanididae) | 아프리카 남부에 서식한다. |
| 동자개과 (Bagridae) | |
| 칼리크티스과 (Callichthyidae) | 파나마 및 남아메리카에 분포하며, 체표면이 2열 골판으로 덮여 있다. 코리도라스가 이 과에 속한다. |
| 케톱시스과 (Cetopsidae) | 남아메리카에 분포하며, 칸디루로 총칭되는 공격적인 성질의 종류를 포함한다. |
| 차카과 (Chacidae) | 인도 동부에서 보르네오섬에 걸쳐 분포하며, 머리가 크고 평평하며 입이 넓어 개구리 같은 외관을 가진다. |
| 공기호흡메기과 (Clariidae) | 대부분 아프리카, 일부 아시아에 분포하며, 아가미에서 발달한 공기 호흡 기관을 가진다. 걸어다니는 메기가 이 과에 속한다. |
| 클라로테스과 (Claroteidae) | 아프리카에 분포한다. |
| 크라노글라니스과 (Cranoglanididae) | 중국, 베트남 등 아시아 대하에 서식한다. |
| 디플로미스테스과 (Diplomystidae) | 칠레, 아르헨티나 등 남아메리카 남부에 분포하며, 메기목에서 가장 원시적인 특징을 보인다. |
| 가시메기과 (Doradidae) | 남아메리카 (특히 브라질, 페루, 기아나)에 분포하며, 가슴지느러미나 부레를 사용하여 발성한다. |
| 에레티스테스과 (Erethistidae) | 남아시아에 분포한다. |
| 헵타프테루스과 (Heptapteridae) | 멕시코에서 남아메리카에 걸쳐 분포한다. |
| 공기주머니메기과 (Heteropneustidae) | 파키스탄부터 태국에 걸친 남부~동남아시아에 분포하며, 아가미에서 몸 뒤쪽으로 뻗은 공기 주머니를 가진다. |
| 호라바그루스과 (Horabagridae) | |
| † Hypsidoridae | |
| 붕메기과 (Ictaluridae) | 북아메리카(캐나다 남부부터 과테말라)에 분포하며, 채널 메기가 이 과에 속한다. |
| 치아파스메기과 (Lacantuniidae) | |
| 로리카리아과 (Loricariidae) | 중앙 아메리카에서 남아메리카에 걸쳐 분포하며, 플레코, 오토싱클루스가 이 과에 속한다. |
| 전기메기과 (Malapteruridae) | 나일강 등 아프리카 열대 지역에 분포하며, 발전 기관을 가지고 있다. |
| 모코쿠스과 (Mochokidae) | 아프리카에 분포하며, 큰 지방 지느러미를 특징으로 한다. 시노돈티스가 이 과에 속한다. |
| 네마토게니스과 (Nematogenyidae 또는 Nematogenyiidae) | 칠레 중부에 서식한다. |
| 메콩메기과 (Pangasiidae) | 파키스탄에서 보르네오섬에 걸쳐 남아시아에 분포하며, 메콩자이언트메기가 이 과에 속한다. |
| 붉은꼬리메기과 (Pimelodidae) | 파나마에서 남아메리카에 걸쳐 분포하며, 붉은꼬리메기가 이 과에 속한다. |
| 쏠종개과 (Plotosidae) | 인도양에서 서부 태평양에 걸쳐 분포하는 해산종과 오스트레일리아, 뉴기니 섬 담수종이 있다. |
| 프세우도피멜로두스과 (Pseudopimelodidae) | 남아메리카에 분포한다. |
| 유리메기과 (Schilbeidae) | 아프리카와 남아시아에 분포한다. |
| 스콜로플락스과 (Scoloplacidae) | 남아메리카에 분포하며, 초소형 종을 포함한다. |
| 메기과 (Siluridae) | 유럽에서 아시아에 걸쳐 분포하며, 메기, 비와호오오나마즈가 이 과에 속한다. |
| 시소르과 (Sisoridae) | 서아시아에서 동남아시아, 터키에서 보르네오 섬에 걸쳐 분포한다. |
| 흡혈메기과 (Trichomycteridae) | 중앙 아메리카 일부에서 남아메리카 전역에 분포하며, 칸디루가 이 과에 속한다. |
2. 3. 계통 분류
메기목의 계통 분류는 2023년 현재 다음과 같다.[92][93][94]| 하위 과 | |
|---|---|
다음은 설리반(Sullivan) 등과 디오고(Diogo)와 펭(Peng)의 연구에 기초한 계통 분류이다.[95][96]
분자적 증거에 따르면, 모든 메기는 단계통군을 형성한다.[7] 메기는 골표상목에 속하며, 여기에는 잉어목, 카라신목, 곤어린쿠스목 및 김노투스목이 포함된다. 일부에서는 김노투스목을 메기목의 하목으로 분류하지만, 널리 받아들여지지 않는다. 현재 메기목은 김노투스목의 자매군이라고 하지만, 최근 분자적 증거로 인해 논쟁이 있다.[8]
약 36개의 현존하는 메기 과가 있으며, 약 3,093종의 현존하는 종이 기술되었다.[9] 이는 메기목을 두 번째 또는 세 번째로 다양한 척추동물 목으로 만든다. 20마리의 척추동물 중 1마리가 메기이다.[10]
메기는 남아메리카를 중심으로 한 곤드와나 기원을 가진 것으로 여겨진다. 가장 초기의 확실한 구성원은 백악기 후기의 캄파니안에서 마스트리히트절에 이르는 시기에 아메리카 대륙에 살았으며, 안디니크티스과, ''Vorhisia vulpes'' 및 아마도 ''아리우스''가 포함된다.[1][11][13]
메기의 분류는 빠르게 변화하고 있다. 2007년과 2008년 논문에서, ''Horabagrus'', ''Phreatobius'', ''Conorhynchos''는 현재의 어떤 메기 과에도 분류되지 않았다.[9]
2005년 6월, 연구자들은 메기의 새로운 과인 Lacantuniidae를 명명했는데, 이는 지난 70년 동안 구별된 세 번째 새로운 물고기 과일 뿐이다. Lacantuniidae의 새로운 종인 ''Lacantunia enigmatica''는 멕시코 치아파스 주의 라칸툰 강에서 발견되었다.[21]
메기목의 상위 계통발생은 최근 여러 차례의 변화를 겪었으며, 이는 주로 분자 계통발생 연구로 인해 발생했다. 대부분의 연구는 메기가 세 개의 주요 계통 (Diplomystidae 과, Loricarioidei, Siluroidei)으로 배열된다는 데 동의하지만, 이들 계통 간의 관계는 논쟁적인 지점이었다.[22][23][24][25][26] 형태학적 데이터에 따르면, Diplomystidae는 일반적으로 가장 초기에 분기되는 메기 계통으로 간주되며, 다른 두 계통인 Loricarioidei와 Siluroidei의 자매군이다.[25][26][27] 분자적 증거는 일반적으로 하목 Loricarioidei가 가장 초기에 분기되는 메기 계통으로 나타나며, Diplomystidae와 Siluroidei를 포함하는 분기도의 자매로 나타난다.[7][22][23][28]
다음은 서로 다른 저자에 의한 과 관계의 목록이다. Lacantuniidae는 Claroteidae의 자매로 위치하는 최근 증거를 기반으로 Sullivan 체계에 포함되어 있다.[30]
2017년에 기반한 현존하는 메기목의 계통 발생 및 Nelson, Grande & Wilson 2016에 기반한 멸종된 과[32]는 다음과 같다.















미할당 과는 다음과 같다.
- Bachmanniidae†
- Scoloplacidae (Loricarioidei)
- Akysidae (Sisoroidea)
- Amblycipitidae (Sisoroidea)
- Anchariidae (Arioidea)
- Ariidae (Arioidea)
- Amphiliidae (거대한 아프리카 메기)
- Austroglanididae (Arioidea)
- Chacidae (Siluroidei)
- ''Conorhynchos'' (Pimelodoidea)
- Cranoglanididae (Ictaluroidea)
- Heteropneustidae (Clarioidea)
- Horabagridae (Sisoroidea)
- Kryptoglanidae (Siluroidei)
- Lacantuniidae (거대한 아프리카 메기)
- Malapteruridae (거대한 아프리카 메기)
- Phreatobiidae (Pimelodoidea)
- ''Rita'' (Sisoroidea)
- Schilbeidae (거대한 아프리카 메기)
3. 형태
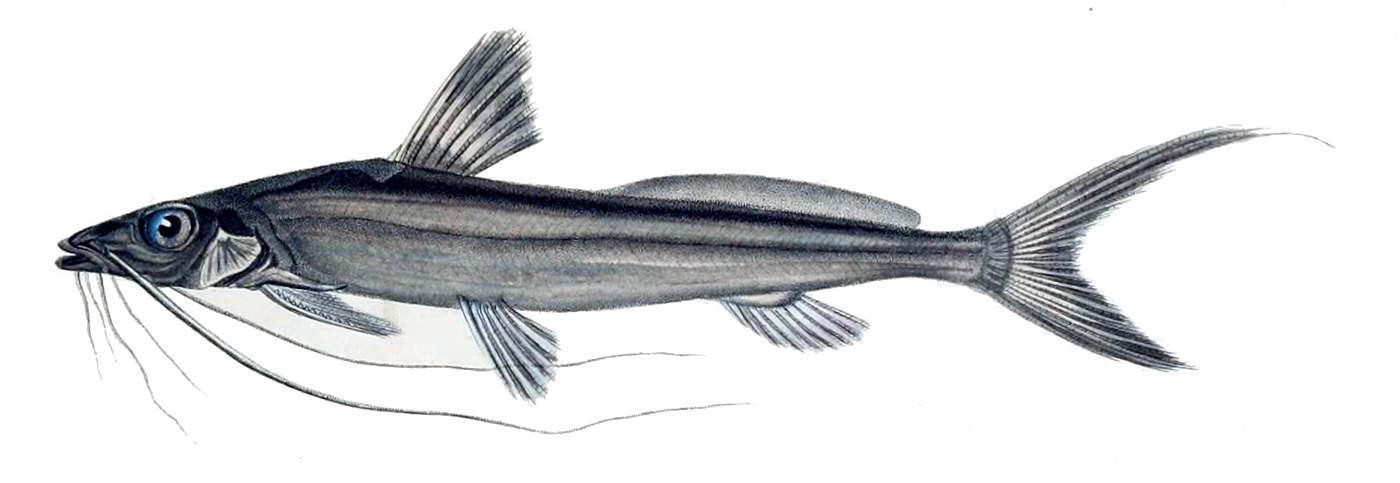
메기류는 납작하고 찌그러진 큰 머리와 넓은 입, 발달된 긴 수염을 가진 것이 특징이다.[90] 몸 표면은 비늘 없이 매끄럽지만, 로리카리아상과와 드라스상과의 일부는 딱딱한 골판이 기와처럼 배열되어 있다. 몸은 좌우로 납작하지만(측편), 머리 부분은 상하로 눌린 듯 납작하다(종편).
코, 위턱, 아래턱 등 머리 부분에 최대 4쌍의 수염이 있는 것이 메기목 어류의 큰 특징이다. 수염에는 미뢰 등 감각 기관이 발달하여 이동 및 먹이 탐색에 활용된다. 야행성이거나 탁한 물에 사는 종류는 눈이 퇴화되어 작아 수염에 의존하는 정도가 크다. 헤테로프네우스테스과와 히레나마즈과는 공기 호흡을 위한 상새 기관을 가지고 있어 물 밖에서도 생존할 수 있다.
메기목에서 가장 큰 종은 메기과의 유럽 메기( ''Silurus glanis'')로, 대부분 몸길이 1.7m 정도이지만 조건이 좋으면 2m를 넘고 최대 2.78m에 달한다는 보고도 있다. 팡가시우스과(메콩자이언트메기 등)와 피메로두스과(피라히바 등)도 매우 크게 성장하지만, 대부분의 메기는 몸길이 12cm 정도의 소형이다. 스콜로플락스과에는 몸길이 2cm 정도로 성숙하는 초소형 종도 있다.[90]
지느러미에는 메기류 특유의 특징이 나타난다. 대부분의 메기는 등지느러미와 가슴지느러미에 "가시"(극조와는 다름)를 가진다. 이 가시는 강하고 날카로우며, 독샘을 갖는 경우도 있어 방어 수단이 된다. 등지느러미 가시 앞에는 작은 가시 모양의 구조가 있어, 이 작은 가시로 큰 가시를 고정할 수 있다.
대부분의 메기류는 지방 지느러미를 갖는다. 지방 지느러미는 등지느러미 뒤쪽, 꼬리지느러미 근처에 위치하는 육질의 지느러미로, 연어목, 카라신목 등에서도 볼 수 있다. 보통 지느러미 줄기가 없지만, 역메기과의 일부는 지느러미 줄기를 갖는다. 메기목과 함께 담수어의 큰 무리를 이루는 잉어목에는 지방 지느러미가 없어, 두 그룹을 구별하는 중요한 특징 중 하나가 된다. 메기류의 꼬리지느러미 주 지느러미 줄기는 18개 이하이며, 많은 종에서 17개이지만, 아스플레도과(10개 이하) 등 더 적은 경우도 있다. 꼬리지느러미 골격은 6개의 분할된 하미골을 갖는 것부터 완전히 유합된 종류까지 다양하며, 과 이하의 분류 형질로 이용된다.
메기목은 잉어목, 카라신목 등과 함께 골표상목에 속하며, 이 그룹은 웨버 기관을 갖는다는 공통점이 있다. 웨버 기관은 척추에서 변화한 4개의 골편으로 구성되어 내이와 부레를 연결하여 뇌에 소리를 전달한다. 야행성으로 작은 눈을 가진 메기류에게는 수염과 함께 중요한 감각 기관이다. 부레는 구형 또는 타원형이며, 바닥에 가만히 있어 유영을 잘 하지 않는 생활 방식을 반영하여 다소 퇴화되었으며, 캡슐 형태로 변형된 척추골에 의해 완전히 둘러싸이는 종류도 있다.
머리가 평평한 것은 골격상의 특징에도 영향을 미친다. 상악골에 이빨이 없고, 상악 골격 자체가 전반적으로 퇴화한 것이 특징이며, 대부분의 과에서는 주상악골이 골편화되어 있다. 이 골편화된 주상악골은 수염을 움직이기 위한 근육의 기점이 된다. 머리나 아가미를 구성하는 뼈 중에서 하새개골, 접속골, 기설골이 없다. 중익상골은 퇴화되었고, 전새개골과 간새개골도 비교적 작다. 두정골과 후측두골은 대부분 불분명하며, 각각 상후두골, 상의쇄골과 유합된 것으로 보인다. 익상골, 구개골, 서골에 이빨을 갖는다. 척추골 수는 15~100개 이상으로 과에 따라 다르다. 다른 진골어류와 달리, 미설골은 개체 발생 초기에 힘줄과 유합하여 특수한 골화를 한다.
3. 1. 외부 형태
대부분의 메기들은 저서 동물을 먹고 산다. 일반적으로 부력이 낮아, 작은 공기주머니와 무겁고 뼈가 많은 머리 때문에 떠오르기보다는 가라앉는 경향이 있다.[33] 메기는 다양한 체형을 가지고 있지만, 대부분은 저서 생활을 위해 배가 평평한 원통형 몸체를 가지고 있다.[33]납작한 머리는 바닥을 파고들 수 있게 해줄 뿐만 아니라 유체역학적 날개 역할을 할 수도 있다. 어떤 메기들은 입이 크게 벌어지고 절치가 없다. 메기는 일반적으로 먹이를 물거나 자르기보다는 흡입하거나 삼키는 방식으로 먹이를 섭취한다.[33] 그러나 일부 과, 특히 Loricariidae와 Astroblepidae는 빠르게 흐르는 물체에 부착할 수 있는 흡반을 가지고 있다. 메기는 또한 상악골이 수염을 지지하도록 축소되어 있다. 이것은 잉어와 같은 다른 물고기처럼 입을 내밀 수 없다는 것을 의미한다.[33]

메기는 최대 4쌍의 수염 - 코, 위턱(입의 양쪽), 2쌍의 턱 수염 - 을 가질 수 있지만, 종에 따라 수염이 없을 수도 있다. 메기의 수염은 항상 쌍으로 나타난다. 많은 대형 메기는 또한 몸 전체에 화학 수용체를 가지고 있어, 만지는 모든 것을 "맛"보고 물속의 모든 화학 물질을 "냄새"를 맡는다. "메기에서 미각은 먹이를 찾고 위치를 파악하는 데 중요한 역할을 한다".[46] 수염과 화학 수용체가 먹이를 감지하는 데 더 중요하기 때문에 메기의 눈은 일반적으로 작다. 다른 골단어류와 마찬가지로 웨버 기관의 존재가 특징이다.[8] 발달된 웨버 기관과 축소된 공기주머니는 청각과 소리 생성을 향상시킨다.[33]
메기는 비늘이 없다. 몸은 종종 매끄럽다. 일부 종에서는 점액으로 덮인 피부가 피부 호흡에 사용되며, 물고기는 피부를 통해 숨을 쉰다.[33] 일부 메기에서는 피부가 골판이라고 하는 뼈판으로 덮여 있다. 어떤 형태의 몸 보호는 목 안에서 다양한 방식으로 나타난다. Loricarioidea와 아시아 속 ''Sisor''에서 갑옷은 주로 하나 이상의 열의 자유 피부판으로 구성된다. 유사한 판은 ''Lithodoras''의 큰 표본에서 발견된다. 이 판은 척추 돌기에 의해 지지될 수 있으며, scoloplacids와 ''Sisor''에서 그렇지만, 돌기는 판에 융합되거나 외부 갑옷을 형성하지 않는다. 반대로, Doumeinae 아과(과 Amphiliidae)와 hoplomyzontines(Aspredinidae)에서는 갑옷은 판을 형성하는 확장된 척추 돌기만으로 형성된다. 마지막으로, doradids, ''Sisor'', hoplomyzontines의 측면 갑옷은 등쪽과 배쪽 판을 가진 비대해진 측선 골편으로 구성된다.[47]
Malapteruridae(전기 메기)를 제외한 모든 메기는 등지느러미와 가슴 지느러미에 강하고 속이 빈 뼈로 된 날카로운 가시 모양의 광선을 가지고 있다. 방어 수단으로, 이 가시는 밖으로 튀어나오도록 고정되어 심각한 상처를 입힐 수 있다.[10] 많은 메기 종에서 이 지느러미 광선은 물고기가 자극을 받으면 쏘는 단백질을 전달하는 데 사용될 수 있다.[48] 모든 메기 종의 절반이 이런 방식으로 독성이 있을 수 있으며, 메기목은 독성 종의 수가 가장 많은 척추 동물 목이 된다.[49] 이 독은 가시를 덮고 있는 표피 조직의 샘 세포에 의해 생성된다.[8] Plotosidae과 ''Heteropneustes'' 속의 구성원에서는 이 단백질이 너무 강력해서 쏘인 사람이 입원할 수 있다. ''Plotosus lineatus''에서는 쏘임이 치명적일 수 있다.[8] 등지느러미와 가슴 지느러미 가시는 실루리폼의 가장 눈에 띄는 특징 중 두 가지이며, 다른 물고기 그룹의 특징과 다르다.[50] 가시가 분류학적 및 계통 발생학적 연구에 널리 사용됨에도 불구하고, 가시 명칭에 대한 일관성이 부족하여 정보를 효과적으로 사용하는 데 어려움을 겪어왔으며, 이 문제를 해결하기 위해 2022년에 메기 가시의 설명 해부학에 대한 일반적인 표준이 제안되었다.[50]
어린 메기는 대부분의 물고기와 마찬가지로, 더 크고 성숙한 개체에 비해 머리, 눈, 후방 중앙 지느러미가 비교적 크다. 이러한 어린 메기는 특히 지느러미나 몸 모양이 매우 변형된 종의 경우, 쉽게 과에 속하게 할 수 있다. 어떤 경우에는 속을 식별할 수 있다. 대부분의 메기에 대해 알려진 바와 같이, 입과 지느러미 위치, 지느러미 모양, 수염 길이와 같이 종의 특징인 경우가 많으며, 어린 개체와 성체 사이에 거의 차이가 없다. 많은 종의 경우, 색소 패턴도 어린 개체와 성체에서 유사하다. 따라서 어린 메기는 일반적으로 성체 형태와 유사하며 뚜렷한 어린 시절의 특수화 없이 부드럽게 발달한다. 이에 대한 예외는 어린 개체가 난황낭을 어린 단계까지 유지하는 ariid 메기, 그리고 길쭉한 수염과 지느러미 필라멘트 또는 착색 패턴을 가질 수 있는 많은 pimelodids이다.[51]
성적 이형성은 모든 메기 과의 약 절반에서 보고된다.[52] 항문 지느러미를 교접기로 변형시키는 것(내부 수정의 경우)과 생식 장치의 부속 구조(내부 및 외부 수정 모두)는 11개의 다른 과에 속하는 종에서 설명되었다.[53]

메기목 어류는 경골어류 내에서 크기 범위가 가장 넓은 분류군 중 하나이다.[33] 많은 메기의 최대 길이는 12cm 미만이다.[8] Aspredinidae와 Trichomycteridae에 속하는 일부 가장 작은 종은 단 1cm에서 성적으로 성숙한다.[10]
웰스 메기 (''Silurus glanis'')와 훨씬 작은 친척인 아리스토텔레스 메기는 유럽에 서식하는 유일한 메기이다. 전자는 유럽 전역에 분포하며 후자는 그리스에 국한된다. 신화와 문학은 놀라운 크기의 웰스 메기를 기록하고 있지만 과학적으로 증명되지는 않았다. 이 종의 일반적인 크기는 약 1.2m ~ 1.6m이며, 2m를 초과하는 물고기는 드물다. 그러나 길이 2.5m 이상, 무게 100kg을 초과하는 것으로 알려져 있다. 2009년 7월, 88kg의 메기가 스페인 에브로 강에서 11세 영국 여학생에게 잡혔다.[54]
북아메리카에서 Ictalurus furcatus (블루 메기) 중 가장 큰 개체는 2010년 7월 20일 미주리 강에서 잡혔으며, 무게는 59kg이었다. 역대 최대의 플랫헤드 메기 (''Pylodictis olivaris'')는 캔자스주 인디펜던스에서 잡혔으며, 무게는 56kg이었다.
이 기록들은 2005년 5월 1일 태국 북부에서 잡혀 약 2개월 후에 언론에 보도된 메콩 자이언트 메기에 비하면 초라하다. 이 메기의 무게는 293kg이었다. 이는 태국 관리들이 1981년부터 기록을 시작한 이후 잡힌 가장 큰 메콩 자이언트 메기이다.[55] 또한 아시아에서는 제레미 웨이드가 인도-네팔 국경의 칼리 강에서 세 번의 치명적인 인간 공격 이후 75.5kg짜리 군치를 잡았다. 웨이드는 이 물고기가 18세 소년뿐만 아니라 물소까지 잡아먹을 정도로 훨씬 더 컸을 것이라고 생각했다.
피라이바 (''Brachyplatystoma filamentosum)''는 예외적으로 크게 성장할 수 있으며 아마존 분지에 서식한다. 수많은 어획 사례를 보면 200kg까지 성장할 수 있다. 이 물고기에게 삼켜져 사망한 사건이 이 지역에서 보고되었다.

메기류의 체표면에는 비늘이 없고, 일반적으로 매끄럽다. 몸은 점액으로 덮여 미끈거리는 경우가 많지만, 로리카리아상과와 드라스상과의 일부에서는 딱딱한 골판이 기와처럼 열을 지어 있다. 몸은 전체적으로 좌우로 납작하지만(측편), 머리 부분은 상하로 압박된 듯 납작하며(종편), 메기목 어류의 외형을 특징짓고 있다.
코·위턱·아래턱 등 머리 부분에 최대 4쌍의 수염을 갖는 것이 메기목 어류의 큰 특징이다. 입 수염에는 미뢰 등 감각 기관이 발달하여 이동 및 먹이를 찾을 때 수염을 최대한 활용한다. 야간에 활동하는 것과 시야 불량의 탁한 수역에 사는 종류에서는 눈이 퇴화되어 작은 경우가 많아 입 수염에 의존하는 정도가 크다. 헤테로프네우스테스과·히레나마즈과는 공기 호흡을 위한 특수한 기관(상새 기관)을 가지고 있어 물 밖에서도 어느 정도 생존이 가능하다.
메기목 중 가장 큰 물고기는 메기과의 유럽 메기(''Silurus glanis'')이다. 이 종의 대부분은 체장 1.7m 정도로 조건이 좋으면 체장 2m를 넘고 최대 2.78m에 달한다는 보고가 있다.
팡가시우스과(메콩자이언트메기 등)·피메로두스과(피라히바 등)의 종류도 매우 커지지만, 전체를 보면 이들은 예외이며, 대부분의 종류는 체장 12cm 정도의 소형 물고기이다. 스콜로플락스과에는 체장 2cm 정도로 성숙하는 초소형 종이 소속되어 있다. 2005년 내셔널 지오그래픽 뉴스에서 2.7m의 메콩자이언트메기가 어획되었다는 보고가 있다.
어류의 중요한 분류 형질인 지느러미에도 메기류 특유의 특징이 보인다. 대부분의 메기는 등지느러미와 가슴지느러미의 선두에 1개의 "가시"(소위 극조와는 다름)를 가지고 있다. 이 가시는 종종 강인하고 날카로우며, 종류에 따라 독샘을 갖는 경우도 있는 등, 외적에 대한 강력한 방어 수단이 된다. 등지느러미 가시 앞에는 더욱 작은 가시 모양의 구조가 있으며, 이 작은 가시를 사용하여 긴 가시를 기계적으로 고정할 수 있다. 강한 가시를 세운 상태로 고정되면 포식자가 삼키는 것은 쉽지 않다.
대부분의 메기류는 지방 지느러미를 갖는다. 지방 지느러미는 등지느러미의 후방, 꼬리지느러미 근처에 위치하는 육질의 지느러미로, 연어목·카라신목 등 여러 그룹에서 볼 수 있다. 지방 지느러미는 지느러미 줄기를 결여하는 것이 보통이지만, 역메기과의 일부 종류는 지방 지느러미에도 지느러미 줄기를 갖는다. 담수어의 일군으로서 메기목과 쌍벽을 이루는 잉어목의 종류에는 지방 지느러미가 없고, 양자의 중요한 감별점 중 하나가 되고 있다. 메기류의 꼬리지느러미의 주 지느러미 줄기는 18개 이하이며, 많은 종에서는 17개이지만, 아스플레도과(10개 이하) 등 더욱 적은 경우도 있다. 꼬리지느러미의 골격은 6개의 분할된 하미골을 갖는 것에서, 완전히 유합된 종류까지 다양하며, 과 이하의 분류 형질로 이용된다.
3. 2. 내부 형태
대부분의 메기들은 저서 동물을 먹고 산다. 일반적으로 부력이 낮아, 작은 공기주머니와 무겁고 뼈가 많은 머리 때문에 떠오르기보다는 가라앉는 경향이 있다.[33] 메기는 다양한 체형을 가지고 있지만, 대부분은 저서 생활을 위해 배가 평평한 원통형 몸체를 가지고 있다.[33]납작한 머리는 바닥을 파고들 수 있게 해줄 뿐만 아니라 유체역학적 날개 역할을 할 수도 있다. 어떤 메기들은 입이 크게 벌어지고 절치가 없다. 메기는 일반적으로 먹이를 물거나 자르기보다는 흡입하거나 삼키는 방식으로 먹이를 섭취한다.[33] 그러나 일부 과, 특히 Loricariidae와 Astroblepidae는 빠르게 흐르는 물체에 부착할 수 있는 흡반을 가지고 있다. 메기는 또한 상악골이 수염을 지지하도록 축소되어 잉어와 같은 다른 물고기처럼 입을 내밀 수 없다.[33]
메기는 최대 4쌍의 수염(코, 위턱(입의 양쪽), 2쌍의 턱 수염)을 가질 수 있지만, 종에 따라 수염이 없을 수도 있다. 메기의 수염은 항상 쌍으로 나타난다. 많은 대형 메기는 또한 몸 전체에 화학 수용체를 가지고 있어, 만지는 모든 것을 "맛"보고 물속의 모든 화학 물질을 "냄새"맡는다. 메기에서 미각은 먹이를 찾고 위치를 파악하는 데 중요한 역할을 한다.[46] 수염과 화학 수용체가 먹이를 감지하는 데 더 중요하기 때문에 메기의 눈은 일반적으로 작다. 다른 골단어류와 마찬가지로 웨버 기관의 존재가 특징이다.[8] 발달된 웨버 기관과 축소된 공기주머니는 청각과 소리 생성을 향상시킨다.[33]
메기는 비늘이 없다. 몸은 종종 매끄럽다. 일부 종에서는 점액으로 덮인 피부가 피부 호흡에 사용되며, 물고기는 피부를 통해 숨을 쉰다.[33] 일부 메기에서는 피부가 골판이라고 하는 뼈판으로 덮여 있다. 몸 보호는 목 안에서 다양한 방식으로 나타난다. Loricarioidea와 아시아 속 ''Sisor''에서 갑옷은 주로 하나 이상의 열의 자유 피부판으로 구성된다. 유사한 판은 ''Lithodoras''의 큰 표본에서 발견된다. 이 판은 척추 돌기에 의해 지지될 수 있으며, scoloplacids와 ''Sisor''에서 그렇지만, 돌기는 판에 융합되거나 외부 갑옷을 형성하지 않는다. 반대로, Doumeinae 아과(과 Amphiliidae)와 hoplomyzontines(Aspredinidae)에서는 갑옷은 판을 형성하는 확장된 척추 돌기만으로 형성된다. 마지막으로, doradids, ''Sisor'', hoplomyzontines의 측면 갑옷은 등쪽과 배쪽 판을 가진 비대해진 측선 골편으로 구성된다.[47]
Malapteruridae(전기 메기)를 제외한 모든 메기는 등지느러미와 가슴 지느러미에 강하고 속이 빈 뼈로 된 날카로운 가시 모양의 광선을 가지고 있다. 방어 수단으로, 이 가시는 밖으로 튀어나오도록 고정되어 심각한 상처를 입힐 수 있다.[10] 많은 메기 종에서 이 지느러미 광선은 물고기가 자극을 받으면 쏘는 단백질을 전달하는 데 사용될 수 있다.[48] 모든 메기 종의 절반이 이런 방식으로 독성이 있을 수 있으며, 메기목은 독성 종의 수가 가장 많은 척추 동물 목이 된다.[49] 이 독은 가시를 덮고 있는 표피 조직의 샘 세포에 의해 생성된다.[8] Plotosidae과 ''Heteropneustes'' 속의 구성원에서는 이 단백질이 너무 강력해서 쏘인 사람이 입원할 수 있다. ''Plotosus lineatus''에서는 쏘임이 치명적일 수 있다.[8] 등지느러미와 가슴 지느러미 가시는 실루리폼의 가장 눈에 띄는 특징 중 두 가지이며, 다른 물고기 그룹의 특징과 다르다.[50] 가시가 분류학적 및 계통 발생학적 연구에 널리 사용됨에도 불구하고, 가시 명칭에 대한 일관성이 부족하여 정보를 효과적으로 사용하는 데 어려움을 겪어왔으며, 이 문제를 해결하기 위해 2022년에 메기 가시의 설명 해부학에 대한 일반적인 표준이 제안되었다.[50]
어린 메기는 대부분의 물고기와 마찬가지로, 더 크고 성숙한 개체에 비해 머리, 눈, 후방 중앙 지느러미가 비교적 크다. 특히 지느러미나 몸 모양이 매우 변형된 종의 경우, 쉽게 과에 속하게 할 수 있다. 어떤 경우에는 속을 식별할 수 있다. 대부분의 메기에 대해 알려진 바와 같이, 입과 지느러미 위치, 지느러미 모양, 수염 길이와 같이 종의 특징인 경우가 많으며, 어린 개체와 성체 사이에 거의 차이가 없다. 많은 종의 경우, 색소 패턴도 어린 개체와 성체에서 유사하다. 따라서 어린 메기는 일반적으로 성체 형태와 유사하며 뚜렷한 어린 시절의 특수화 없이 부드럽게 발달한다. 이에 대한 예외는 어린 개체가 난황낭을 어린 단계까지 유지하는 ariid 메기, 그리고 길쭉한 수염과 지느러미 필라멘트 또는 착색 패턴을 가질 수 있는 많은 pimelodids이다.[51]
성적 이형성은 모든 메기 과의 약 절반에서 보고된다.[52] 항문 지느러미를 교접기로 변형시키는 것(내부 수정의 경우)과 생식 장치의 부속 구조(내부 및 외부 수정 모두)는 11개의 다른 과에 속하는 종에서 설명되었다.[53]
메기목 어류는 경골어류 내에서 크기 범위가 가장 넓은 분류군 중 하나이다.[33] 많은 메기의 최대 길이는 12cm 미만이다.[8] Aspredinidae와 Trichomycteridae에 속하는 일부 가장 작은 종은 단 1cm에서 성적으로 성숙한다.[10]

많은 메기류에서 "상완돌기"는 가슴 지느러미 기저부 바로 위에 있는 가슴띠에서 뒤쪽으로 뻗어 있는 뼈 돌기이다. 이는 피부 아래에 위치하며, 피부를 해부하거나 바늘로 찔러 그 윤곽을 확인할 수 있다.[56]
메기류의 망막은 단일 원추 세포와 큰 간상 세포로 구성되어 있다. 많은 메기류는 휘판을 가지고 있는데, 이는 광자 포착을 향상시키고 저조도 감도를 높이는 데 도움이 될 수 있다. 대부분의 경골어류에 존재하지만 이중 원추는 메기류에는 없다.[57]
메기류의 고환 해부학적 구성은 메기류 과에 따라 다르지만, 대부분은 가장자리 고환을 나타낸다: ictaluridae, claridae, auchenipteridae, doradidae, pimelodidae, pseudopimelodidae.[58] 일부 실루리폼류 종의 고환에서는 정자 발생 두부와 분비 미부와 같은 기관 및 구조가 관찰되며, 미부에서 정낭이 존재한다.[59] 총 가장자리 수와 길이는 종 사이의 미부 및 두부 부분에서 다르다.[58] 미부의 가장자리는 세관을 나타낼 수 있으며, 그 내강은 분비물과 정자로 채워져 있다.[58] 정자낭은 세르톨리 세포의 세포질 확장에서 형성되며; 낭벽이 파괴됨으로써 정자의 방출이 허용된다.[58]
정낭의 발생은 크기, 육안 형태 및 기능에 있어서 종간 변이가 있음에도 불구하고, 수정 방식과 관련이 없다. 그들은 전형적으로 쌍을 이루고, 다중 실로 되어 있으며, 정관과 연결되어 있으며, 분비 및 저장 기능을 하는 것으로 보고되었다. 정낭 분비물은 스테로이드와 스테로이드 글루쿠로니드를 포함할 수 있으며, 호르몬 및 페로몬 기능을 하지만, 주로 점액 단백질, 산성 뮤코다당류 및 인지질로 구성되는 것으로 보인다.[53]
물고기 난소는 두 가지 유형일 수 있다 - 체강 난소 또는 낭 난소. 첫 번째 유형에서는 난모세포가 직접 체강으로 방출된 다음 제거된다. 두 번째 유형에서는 난모세포가 난관을 통해 외부로 전달된다.[59] 많은 메기류는 낭 난소 유형이며, 여기에는 ''프세우도플라티스토마 코르루스칸스'', ''P. 파시아텀'', ''로피오실루루스 알렉산드리'', ''로리카리아 렌티기노사''가 있다.[58][59]
메기목은 잉어목이나 카라신목 등과 함께 골표상목으로 불리는 그룹에 속하며, 이 무리에 공통적으로 나타나는 특징으로는 웨버 기관을 갖는다는 점이 있다. 이 기관은 척추에서 변화한 4개의 골편으로 구성되어 있으며, 내이와 부레를 연결하여 뇌에 소리를 전달하는 역할을 한다. 야행성으로 작은 눈을 가진 메기류에게는, 수염과 함께 중요한 감각 기관이다. 부레의 형태는 구형~타원형이며, 바닥에 가만히 있어 유영을 잘 하지 않는 생활 방식을 반영하여 다소 퇴화되었으며, 캡슐 형태로 변형된 척추골에 의해 완전히 둘러싸이는 종류도 있다.
머리가 평평한 것은 골격상의 특징에도 영향을 미친다. 상악골에 이빨이 없고, 상악의 골격 자체가 전반적으로 퇴화한 것이 현저한 특징이며, 대부분의 과에서는 주상악골이 골편화되어 있다. 이 골편화된 주상악골은 수염을 움직이기 위한 근육의 기점이 된다. 머리나 아가미를 구성하는 뼈 중에서, 하새개골·접속골·기설골을 결여한다. 중익상골은 퇴화되었고, 전새개골·간새개골도 비교적 작다. 또한, 두정골과 후측두골은 대부분 불분명하며, 각각 상후두골, 상의쇄골과 유합된 것으로 보인다. 익상골·구개골·서골에 이빨을 갖는다. 척추골의 수는 15~100개 이상으로 과에 따라 다르다. 다른 진골어류와는 달리, 미설골은 개체 발생 초기에 힘줄과 유합하여 특수한 골화를 한다.
4. 생태
메기목 어류는 기본적으로 저서성이며 활발하게 헤엄치는 경우는 적고, 물 밑을 기어 다니듯 천천히 헤엄치는 경우가 많다. 대부분 야행성이며, 입수염을 활발하게 움직여 주변을 탐색하며 이동한다. 로리카리아과 등 물살이 빠른 계류에 분포하는 과에서는 입이 흡반 모양으로 변형되어 있어, 바위 등에 붙어 격류를 헤쳐 나간다.
메기목 어류 중에는 체내에 독을 포함하거나, 독샘을 통해 체외로 독액을 분비하는 종류가 알려져 있다. 독액은 등지느러미와 가슴지느러미(대부분은 후자)의 가시에서 분비된다. 대부분의 독 메기류에게 이러한 가시는 방어 수단이며, 적극적으로 공격하는 경우는 드물다. 인도에 분포하는 레드캣(''Heteropneustes fossiles'', 헤테로프네우스테스과) 등 일부 종류에 한해 독가시를 사용하여 다른 물고기나 인간을 공격하는 습성을 가지고 있는 것으로 알려져 있다. 일본 연안에도 분포하는 해산종인 쏠종개(''Plotosus lineatus'', 쏠종개과)가 가진 독은 강력하여, 쏘일 경우 사망에 이를 수도 있다.
드라스과나 기기과 등의 메기류에는 가슴지느러미 가시나 부레의 진동을 이용하여 소리를 낼 수 있는 종류가 있다. 일본을 포함한 동아시아에 분포하는 기기(''Pelteobagrus nudiceps'')는 가슴지느러미 가시를 사용하여 내는 위협 소리가 이름의 유래가 되었다. 이러한 메기류는 어획될 때에도 발음하기 때문에, 외부 적에 대한 경고의 역할을 하는 것으로 보이지만, 동료 간의 전달 수단으로 사용되는지는 불명확하다.
전기메기과는 발전을 하는 어류로 알려져 있다. 특히 전기메기는 최대 400V를 넘는 전압을 낼 수 있으며, 이는 어류로서는 전기뱀장어(최대 600V)에 이어 높은 발전력이다. 전기메기과뿐만 아니라 대부분의 메기목 어류는 체표(특히 입과 머리 부분)에 전기장을 감지하는 수용기가 있어, 수중에서의 전기장 변화에 민감하게 반응한다. 메기류의 전기 수용기는 연골어류가 가진 로렌치니 기관과 비슷한 병 모양을 하고 있지만, 동모 유무 등 세포 수준에서의 미세 형태에 차이가 있다. 야행성 종류가 많은 메기류에게는, 수염이나 웨버 기관과 함께 중요한 감각기로서 기능하는 것으로 보인다.
4. 1. 서식지
메기는 주로 느리게 움직이는 물속의 어두운 강바닥에 산다. 주로 어두운 곳에 살기 때문에 침침한 색깔을 띠고 있는데, 글래스캣피시라는 메기는 투명한 색깔을 띠어 몸을 보호한다. 또한, 메기들은 동굴, 바다, 또는 땅밑에까지 산다. 환경을 거의 가리지 않고 살 수 있는 능력 덕분에 메기와 그 친척들은 민물고기의 4분의 3 정도를 차지한다.[8]현존하는 메기목 어류는 남극을 제외한 모든 대륙의 내륙 또는 연안 해역에 서식한다. 한때 모든 대륙에 서식했었다.[8] 열대 열대 남아메리카, 아시아, 아프리카에서 가장 다양한 종류가 서식하며, 북아메리카와 유럽에 각각 한 과(科)가 있다.[10] 모든 메기목 어류 종의 절반 이상이 아메리카 대륙에 서식한다. 마다가스카르, 호주, 뉴기니에서 담수 서식지에 진입한 유일한 골단어류이다.[33]
대부분 얕은 물이나 흐르는 물에 서식하지만, 담수/기수 환경에서도 발견된다.[33] 최소 8개 과의 대표 어종이 저서생물(지하에 서식)이며, 3개 과는 동굴 생물(동굴에 서식)이다.[34][35] Phreatobius cisternarum는 대수층 서식지에서 지하 생활하는 것으로 알려져 있다.[36] Ariidae, Plotosidae, Aspredinidae, Bagridae의 몇몇 종은 바닷물에서 발견된다.[37][38]
4. 2. 먹이
메기류는 기본적으로 저서성이며 활발하게 헤엄치는 경우는 적고, 물 밑을 기어 다니듯 천천히 헤엄치는 경우가 많다. 대부분 야행성이며, 입수염을 활발하게 움직여 주변을 탐색하며 이동한다. 로리카리아과 등 물살이 빠른 계류에 분포하는 과에서는 입이 흡반 모양으로 변형되어 있어, 바위 등에 붙어 격류를 헤쳐 나간다.4. 3. 번식
메기목 어류는 종에 따라 다양한 번식 방법을 가진다. 대부분의 메기류는 일반적인 체외수정을 하지만, 아우케니프테루스과 무리는 수컷이 교접기를 가지고 교미를 통해 체내수정을 한다. 코리도라스아과 일부에서는 암컷이 정자를 삼켜 장관을 경유하여 체내수정을 하는 특이한 번식 방법이 알려져 있다.어미가 알이나 치어를 보호하는 습성을 가진 종류도 많다. 수저의 모래를 크레이터 모양으로 판 둥지를 만드는 종류, 칼릭티스아과 등 수면에 거품 둥지를 만드는 종류 등이 있다. 또한, 해산인 하마기기과의 무리는 수컷이 알을 입 안에 물고 보호하는 마우스브루더로서의 육아를 한다.
아프리카의 탕가니카 호에 서식하는 거꾸로 메기과의 일종(''Synodontis multipunctatus'')은 시클리드(농어목) 등 다른 마우스브루더에게 알을 맡겨 치어를 키우는 탁란을 하는 매우 드문 습성을 가진 어류이다. 맡겨진 메기 치어는 숙주인 시클리드의 알보다 빨리 부화하고 성장하여, 최종적으로 치어를 다 먹어 치운다. 이처럼 숙주에게 이익이 없는 뻐꾸기형 탁란을 하는 어류는 이 종 외에는 알려져 있지 않다.
말라위 호에 사는 기기과의 칸팡고(''Bagrus meridionalis'')는 부화 후에 암컷 어미가 다량의 미수정란을 낳아 치어에게 먹이는 습성이 있다. ''S. multipunctatus''의 예와는 반대로, 칸팡고의 둥지에는 많은 시클리드류가 치어를 맡기러 온다.
4. 4. 발성 및 전기 발생
메기목 어류는 다양한 종류의 소리를 낼 수 있으며, 서로 다른 음높이와 속도의 소리를 구별하는 데 사용되는 발달된 청각 수용 능력을 가지고 있다. 또한 소리의 기원과 방향을 감지할 수 있다.[60] 이는 특히 대립 행동과 고통 행동에서 매우 중요한 어류 통신 메커니즘이다. 메기목 어류는 의사소통을 위해 다양한 소리를 낼 수 있으며, 크게 드럼 소리와 마찰음 소리로 분류할 수 있다. 메기류의 소리 신호 변동성은 소리가 생성되는 메커니즘, 결과적인 소리의 기능, 크기, 성별, 나이와 같은 생리적 차이 등 몇 가지 요인에 의해 달라진다.[61]드럼 소리를 만들기 위해 메기목 어류는 부레를 사용하는 간접적인 진동 메커니즘을 사용한다. 이 어류에서 음파 근육은 탄성 스프링이라고도 알려진 Mulleri의 가지에 삽입된다. 음파 근육은 탄성 스프링을 앞으로 당기고 부레를 확장시킨다. 근육이 이완되면 스프링의 장력으로 부레가 원래 위치로 빠르게 돌아가 소리가 발생한다.[62]
메기목 어류는 또한 가슴 지느러미에 소리 생성 메커니즘을 가지고 있다. 메기목 어류의 많은 종은 가시라고 불리는 강화된 첫 번째 가슴 지느러미 줄기를 가지고 있으며, 이는 큰 외전근과 내전 근육에 의해 움직일 수 있다. 메기목 어류의 가시 기저부에는 일련의 능선이 있으며, 가시는 일상적인 움직임 동안 어류의 골반대 홈 안에서 일반적으로 미끄러진다. 그러나 가시의 능선을 골반대 홈에 누르면 일련의 짧은 맥동이 생성된다.[60][62] 이 움직임은 빗의 이빨을 따라 손가락을 움직이는 것과 유사하며, 결과적으로 일련의 날카로운 두드림이 생성된다.[61]
소리 생성 메커니즘은 성별에 따라 종종 다르다. 일부 메기목 어류의 경우 가슴 지느러미는 비슷한 길이의 암컷보다 수컷에서 더 길며, 생성되는 소리의 특성 차이도 관찰되었다.[62] Maria Clara Amorim의 연구에 따르면, 같은 메기목 어류목에 속하는 과들 간의 비교에서 과 및 종 특이적인 발성 패턴이 나타났다. 세 종의 ''코리도라스'' 메기목 어류의 구애 행동 동안, 모든 수컷은 알 수정 전에 능동적으로 마찰음 소리를 생성했으며, 종의 소리는 펄스 수와 소리 지속 시간에서 달랐다.[63]
메기목 어류의 소리 생성은 싸움 및 경고 신호와 관련될 수 있다. Kaatz의 연구에 따르면, 방해(예: 경고) 및 대립 행동에 대한 소리는 유의미한 차이가 없었으며, 이는 고통 소리가 대립 소리 생성의 변화를 샘플링하는 데 사용될 수 있음을 시사한다.[63] 그러나 몇 종의 열대 메기목 어류를 비교한 결과, 일부 어류는 고통스러운 조건에서 드럼 소리보다 더 높은 강도의 마찰음 소리를 생성했다.[64] 드럼 소리와 마찰음 소리의 비율 차이는 다른 크기의 드럼 근육과 가슴 가시와 같은 형태적 제약에 따라 다르다. 이러한 제약으로 인해 일부 어류는 특정 소리를 생성하지 못할 수도 있다. 여러 종의 메기목 어류에서 공격적인 소리 생성은 은신처 방어 중이나 다른 어류의 위협 동안 발생한다. 더 구체적으로, 긴 수염 메기목 어류에서 드럼 소리는 위협 신호로 사용되고 마찰음은 방어 신호로 사용된다. Kaatz는 14과의 83종의 메기목 어류를 조사하여 메기목 어류가 방해 상황에서 더 많은 마찰음 소리를, 동종 간의 갈등 상황에서 더 많은 부레 소리를 생성한다는 것을 밝혀냈다.[64]
메기목 어류 중에는 체내에 독을 포함하거나, 독샘을 통해 체외로 독액을 분비하는 종류가 알려져 있다. 독액은 등지느러미와 가슴지느러미(대부분은 후자)의 가시에서 분비된다. 대부분의 독 메기류에게 이러한 가시는 방어 수단이며, 적극적으로 공격하는 경우는 드물다. 인도에 분포하는 레드캣(''Heteropneustes fossiles'', 헤테로프네우스테스과) 등 일부 종류에 한해 독가시를 사용하여 다른 물고기나 인간을 공격하는 습성을 가지고 있는 것으로 알려져 있다. 일본 연안에도 분포하는 해산종인 쏠종개(''Plotosus lineatus'', 쏠종개과)가 가진 독은 강력하여, 쏘일 경우 사망에 이를 수도 있다.
드라스과나 기기과 등의 메기류에는 가슴지느러미 가시나 부레의 진동을 이용하여 소리를 낼 수 있는 종류가 있다. 일본을 포함한 동아시아에 분포하는 기기(''Pelteobagrus nudiceps'')는 가슴지느러미 가시를 사용하여 내는 위협 소리가 이름의 유래가 되었다. 이러한 메기류는 어획될 때에도 발음하기 때문에, 외부 적에 대한 경고의 역할을 하는 것으로 보이지만, 동료 간의 전달 수단으로 사용되는지는 불명확하다.
전기메기과는 발전을 하는 어류로 알려져 있다. 특히 전기메기는 최대 400V를 넘는 전압을 낼 수 있으며, 이는 어류로서는 전기뱀장어(최대 600V)에 이어 높은 발전력이다. 전기메기과에 한정되지 않고 메기목 어류의 대부분은 체표(특히 입과 머리 부분)에 전기장을 감지하는 수용기가 있어, 수중에서의 전기장 변화에 민감하게 반응하는 것으로 생각된다. 메기류의 전기 수용기는 연골어류가 가진 로렌치니 기관과 비슷한 병 모양을 하고 있지만, 동모 유무 등 세포 수준에서의 미세 형태에 차이가 있다. 야행성 종류가 많은 메기류에게는, 수염이나 웨버 기관과 함께 중요한 감각기로서 기능하는 것으로 보인다.
'''전기메기과'''는 2속 19종을 포함하며, 나일강 등 아프리카의 열대 지역에 분포한다. 잘 발달된 발전 기관을 갖는 것이 최대 특징이다. 발전 기관은 몸 앞부분의 근육을 기원으로 한다. 입수염은 3쌍이다. 등지느러미가 없고, 가슴지느러미에는 가시가 없다. 기름지느러미는 몸 뒤쪽에 위치하며, 꼬리지느러미는 둥근 모양이다. 부레는 몸 뒤쪽으로 길게 뻗어 있으며, 2개 또는 3개의 작은 방으로 나뉜다.
- 전기메기속 ''Malapterurus''
- ''Paradoxoglanis'' 속
5. 인간과의 관계
메기목 어류는 납작하고 찌그러진 큰 머리와 넓은 입, 그리고 발달한 긴 수염을 가진 것이 특징이며, 전 세계의 다양한 환경에 분포한다. 이들은 식용, 관상용, 낚시 등 여러 방면에서 인간과 밀접한 관계를 맺고 있다.
메기는 중요한 어업 자원으로서 오랫동안 이용되어 왔으며, 양식 또한 활발하게 이루어지고 있다. 특히 아시아와 아프리카 지역을 중심으로 양식 생산량이 크게 증가하고 있다. 국제 연합 식량 농업 기구(FAO)의 통계에 따르면, 1950년대에는 남짓이었던 세계 메기목 어류의 총 어획량은 1990년대 후반에는 을 넘어섰고, 2006년에는 에 달했다.[88]
최근에는 코리도라스, 시노돈티스, 플레코 등 다양한 종류가 수족관에서 관상어로 인기를 얻고 있으며, 스포츠 낚시의 대상어로도 각광받고 있다.
하지만, 이식된 외래 메기가 고유 생태계에 영향을 미치는 문제도 발생하고 있다. 워킹캣피시는 국제 자연 보전 연맹(IUCN)이 지정한 "세계의 침략적 외래종 100" 중 하나로, 세계 각지에 정착하여 생태계를 교란시키고 있다.[85] 일본에서도 채널 메기 등이 외래생물법에 따라 규제 대상이 되고 있다.[86]
일본 신화에서는 지진을 일으키는 거대한 메기 나마즈에 대한 이야기가 전해져 내려온다.
5. 1. 식용
메기목 어류는 전 세계적으로 식용으로 널리 이용되며, 다양한 요리법으로 소비된다.메기는 따뜻한 기후에서 양식하기 쉬워, 저렴하고 안전한 식품으로 공급된다. 미국 양식 메기의 약 60%는 미시시피주 벨조니 반경 100km 이내에서 재배된다.[65] 채널 메기( ''Ictalurus punctatus'')는 연간 4.5억달러 규모의 양식 산업을 뒷받침한다.[10] 주요 생산지는 미국 남부 (미시시피주, 앨라배마주, 아칸소주 등)이다.[66]
내륙 탱크나 채널에서 사육된 메기는 폐기물과 질병이 억제되어 야생으로 확산되지 않으므로 일반적으로 환경에 안전하다고 평가받는다.[67]
아시아에서는 많은 메기 종이 식량으로 중요하며, 공기 호흡 메기(Clariidae)와 상어 메기(Pangasiidae) 종은 아프리카와 아시아에서 대규모로 양식된다. 베트남에서 특정 상어 메기 종( ''Pangasius bocourti'')의 수출은 미국 메기 산업의 압력을 받았다. 2003년, 미국 의회는 수입된 생선을 메기로 표기하는 것을 금지하는 법안을 통과시켰다.[68] 그 결과, 베트남은 미국에서 판매되는 자국 제품에 "바사 생선"으로 표기하고 있다. 트레이더 조는 베트남산 ''Pangasius hypophthalmus''의 냉동 필레를 "스트라이퍼"로 표기했다.[69]

메기는 수천 년 동안 아프리카, 아시아, 유럽, 북아메리카에서 식용으로 널리 잡히고 양식되어 왔다. 품질과 풍미에 대한 평가는 다양하며, 일부는 메기를 훌륭하다고 평가하는 반면, 다른 이들은 물기가 많고 맛이 없다고 평가한다.[70] 메기에는 비타민 D가 많이 함유되어 있다.[71] 양식 메기는 오메가-3 지방산 수치가 낮고 오메가-6 지방산 비율이 높다.[72]
중앙 유럽에서 메기는 진미로 여겨져 축제일과 명절에 즐겨 먹었다. 유럽과 아프리카에서 미국으로 이주한 사람들은 이러한 전통을 가져왔고, 미국 남부에서는 메기가 인기 있는 음식이다.
미국에서 가장 흔하게 먹는 종은 채널 메기와 블루 메기인데, 둘 다 야생에서 흔하고 양식도 늘고 있다. 양식 메기는 미국의 식단에서 주식이 되었고, 로널드 레이건 대통령은 1987년 6월 25일을 "양식 메기의 가치"를 인정하는 전국 메기 데이로 선포했다.[73]
메기는 다양한 방식으로 먹는다. 유럽에서는 잉어와 비슷하게 요리하지만, 미국에서는 옥수수 가루를 묻혀 튀겨 먹는 것이 일반적이다.[70]

인도네시아에서는 메기를 보통 ''와룽''이라는 노점에서 튀기거나 구워 채소, 삼발(매운 렐리시 또는 소스)과 함께 먹으며, ''나시 우둑''(전통 코코넛 쌀)과 함께 먹는다. 이 요리는 pecel leleid 또는 pecak leleid라고 불린다. Leleid는 메기를 뜻하는 인도네시아어 단어이다. 생선을 삼발과 함께 돌 절구로 가볍게 으깬 경우 lele penyetid(으깬 메기)라고도 부른다.
말레이시아에서 메기는 ''이칸 켈리''(ikan keli)라고 불리며 향신료와 함께 튀기거나 구워 타마린드와 태국 고추 그레이비와 함께 먹으며, 찐 밥과 함께 먹기도 한다.
방글라데시와 인도의 오디샤, 서벵골, 아삼 주에서는 메기(현지에서는 ''마구르''(magur)라고 알려짐)를 우기 동안 선호하는 진미로 먹는다. 인도의 케랄라 주에서는 ''테두''(thedu) 또는 ''에타''(etta)로 알려진 현지 메기가 인기가 있다.
헝가리에서는 메기를 종종 파프리카 소스(Harcsapaprikás)로 요리하는데, 이는 헝가리 요리의 전형적인 요리이다. 전통적으로 커드 치즈(túrós csusza)를 듬뿍 바른 파스타와 함께 제공된다.
미얀마(구 버마)에서는 메기를 레몬그라스, 생강, 마늘, 후추, 바나나 줄기, 양파 및 기타 현지 재료로 요리한 전통 국수 생선 수프인 ''모힝가''에 사용한다.

베트남 메기(속 ''Pangasius'')는 미국에서 메기로 판매될 수 없으므로, ''스와이'' 또는 ''바사''로 불린다.[74] Ictaluridae과에 속하는 물고기만이 미국에서 메기로 판매될 수 있다.[75][76] 영국에서는 베트남 메기를 "베트남 리버 코블러"로 판매하기도 하지만, 바사로 더 흔하게 판매한다.[77]
나이지리아에서는 메기를 종종 다양한 스튜로 요리한다. 특히 "메기 페퍼 수프"는 전국에서 즐겨 먹는 진미이다.[78]
유대교의 코셔 식사 규정에 따르면, 생선은 코셔가 되려면 지느러미와 비늘이 있어야 한다.[79] 메기는 비늘이 없으므로 코셔가 아니다.[80]
메기목 어류는 세계 각지에서 오래전부터 식용어로 이용되어 왔다. 어획 대상이 되는 것은 아메리카메기과(북아메리카), 메기과·팡가시우스과(아시아), 그리고 실꼬리치과(아시아·아프리카)에 속하는 중·대형종이 많으며, 이들 담수산 종은 양식도 각 지역에서 활발하게 이루어지고 있다.
국제 연합 식량 농업 기구(FAO)의 통계[88]에 따르면, 세계 메기목 어류의 총 어획량은 다음과 같이 변화하였다.
| 연도 | 1950년대 | 1990년대 후반 | 2000년 | 2006년 |
|---|---|---|---|---|
| 총 어획량 | 남짓 | 이상 |
지역별 어획량 변화는 다음과 같다.
| 지역 | 2000년~2006년 변화 |
|---|---|
| 아시아 | 약 3배 증가 ( → ) |
| 북남미 | 대 유지 |
| 유럽 | 대 유지 |
2006년 총 어획량 중 양식 메기가 을 차지한다. 아시아·아프리카 양 지역에서의 어획량 증가 역시 내수면에서의 양식업 발달에 의해 뒷받침되고 있다.
아시아에서의 담수 메기류 양식 수확량 변화는 다음과 같다.
| 연도 | 1990년 | 2000년 | 2006년 |
|---|---|---|---|
| 수확량 |
아프리카에서는 2000년에 미만이었던 어획량이 2006년에는 으로 10배 이상 증가했다. 생산액 면에서도 비슷한 성장이 보이고 있으며, 아시아 지역에서는 2006년에 16억달러에 달했다(2000년 시점에서 3억달러).
해산 메기류로는 갯장어과의 어획량이 비교적 많으며, 아시아 지역에서의 수확량은 1990년대 이후 거의 지속적으로 대를 유지하고 있다. 융중치과는 동 지역·동 시대에 ~의 어획량에 머물고 있다.
항해기과 에는 21속 150종이 있으며, 대부분 해산종이다. 열대에서 온대에 걸쳐 따뜻한 바다에 널리 분포하며, 담수·기수역으로 진출하는 종류도 많다. 입수염은 대부분 3쌍(드물게 2쌍)이다. 기름지느러미, 두 갈래로 갈라지는 꼬리지느러미를 가진다. 머리와 등지느러미 근처에 골판을 가진 종류도 있다. 등지느러미·가슴지느러미에 가시를 가진다.
항해기과는 메기목 중에서는 몇 안 되는 해산 그룹이다. 활발하게 헤엄쳐 다니는 물고기가 많으며, 대부분 수컷이 알을 입 안에서 보호하는 구강 부화를 한다.
- 항해기속
- 다른 20속
판가시우스과 는 3속 28종을 포함하며, 파키스탄에서 보르네오섬에 걸쳐 남아시아를 중심으로 분포한다. 메콩자이언트메기는 식물성 먹이를 먹는 대형종으로, 몸길이 3m, 몸무게 300kg에 달하는 경우도 있다.
몸은 옆으로 납작하며, 입수염은 2쌍으로 짧다. 기름지느러미는 작다. 등지느러미는 몸의 앞쪽에 있으며, 가시를 갖는다. 식용으로 사용되는 중·대형종을 포함하며, 활발하게 헤엄치는 것이 많다.
- ''Pangasius'' 속
- 다른 2속
5. 2. 관상용
수족관에서 인기 있는 메기 종류로는 코리도라스, 플레코스토머스, 거꾸로메기 등이 있다. 코리도라스와 플레코스토머스는 이끼를 먹는 것으로 알려져 수조 관리에 도움이 된다고 생각되지만, 실제로는 별도의 먹이를 주어야 한다.[65] 플레코스토머스는 최대 70cm까지 자랄 수 있어 큰 수조가 필요하다.thumb
수족관에서는 수백 종의 메기가 관상어로 인기가 있다. '''칼리크티스과(Callichthyidae)'''의 코리도라스속 메기들과 '''로리카리아과'''의 플레코로 통칭되는 메기들이 대표적이다. 그 외에도 반조 메기, 토킹 메기, 긴 수염 메기, 시노돈티스속 메기 등이 관상어로 사육된다.
5. 3. 낚시
메기는 낚시꾼들에게 인기 있는 어종이다. 특히 대형 메기는 큰 몸집과 강력한 힘으로 낚시꾼들에게 짜릿한 손맛을 제공한다.유럽에서는 웰스 메기와 아리스토텔레스 메기가 낚시 대상 어종이다. 웰스 메기는 1.2m 정도가 일반적이지만, 2.5m 이상, 무게 100kg이 넘는 대형 개체도 기록된 바 있다. 2009년에는 스페인 에브로 강에서 88kg의 웰스 메기가 잡히기도 했다.[54]
북아메리카에서는 블루 메기(Ictalurus furcatus)와 플랫헤드 메기(Pylodictis olivaris)가 인기 있다. 2010년 미주리 강에서 잡힌 블루 메기는 59kg, 캔자스주 인디펜던스에서 잡힌 플랫헤드 메기는 56kg에 달했다.
아시아에서는 메콩 자이언트 메기와 군치(Bagarius yarrelli)가 유명하다. 2005년 태국 북부에서 잡힌 메콩 자이언트 메기는 293kg으로 기록되었다.[55] 인도-네팔 국경의 칼리 강에서는 제레미 웨이드가 75.5kg짜리 군치를 잡았는데, 이 물고기는 사람이나 물소를 공격할 정도로 크게 자라는 것으로 알려져 있다.
남아메리카의 아마존 분지에 서식하는 피라이바(Brachyplatystoma filamentosum)는 200kg까지 성장하는 대형 메기이다.
대부분의 메기는 인간에게 무해하지만, 줄무늬 장어 메기(Plotosus lineatus) 등 일부 종은 독을 가지고 있어 주의해야 한다.[83]
국제 게임 피시 협회(IGFA)에 따르면, 1998년 5월 19일 미국 캔자스주 엘크 시티 저수지에서 켄 폴리가 낚은 퉁가리의 무게는 55.79kg이었다.[84]
5. 4. 유해종
퉁가리속의 대표 종들은 스포츠와 식량 자원을 얻기 위해 유럽에 도입되었지만, 유럽산 미국 메기들은 원산지에서만큼의 크기로 성장하지 못했으며, 유럽 고유 동물상에 대한 생태적 압박만 가중시켰다.[10] 걸어다니는 메기 또한 플로리다의 담수 지역에 도입되었으며, 탐욕스러운 이 메기는 그곳에서 주요 외래 해충이 되었다. 플랫헤드 메기(''Pylodictis olivaris'') 또한 대서양 사면 배수구역에서 북미의 유해종이다.[10] 플레코속 (''Pterygoplichthys'') 종들은 수족관에서 기르던 사람들이 방생하여, 전 세계 많은 따뜻한 물에서 야생화된 개체군을 형성했다.[41][42][43][44][45]양식 목적으로 유입된 식용 메기나, 사육을 포기한 외래 관상용 메기가 자연계에 정착하여 문제가 되고 있는 사례가 세계 각지에서 알려져 있다. 담수산 메기(특히 중~대형종)의 대부분은 서식 환경에서의 먹이 사슬 상위에 위치하는 경우가 많아, 토착 수생 생물을 멸종시키는 등 생태계에 악영향을 미치는 것이 우려된다. 메기과의 워킹캣피시(클라리아스 바트라쿠스, ''Clarias batrachus'')는 본래 동남아시아에 분포하는 열대성 메기이지만, 현재는 미국 남부나 하와이 등 세계 여러 지역에 귀화해 있다. 이 종은 호흡을 통해 공기 호흡이 가능하며, 육상에서 이동하는 특성이 있어 쉽게 분포를 넓힐 수 있으며, 국제 자연 보전 연맹(IUCN)이 지정하는 "세계의 침략적 외래종 100" 중 하나로 선정되었다.[85]
외국산 메기의 정착은 일본에서도 문제가 되고 있다. 1981년, 가스미가우라에 식용 목적으로 도입된 북아메리카 원산의 채널 메기(아메리카메기, ''Ictalurus punctatus'')는 1994년 이후 급격히 개체수를 늘리고 있다. 이 종은 체장 1m를 넘는 대형 포식어이며 다른 천적이 없고, 외래생물법에 따른 특정 외래 생물로 규제의 대상이 되고 있다.[86] 또한, 유럽메기(메기목 중 최대종)・워킹캣피시의 정착이 우려되는 외에, 얼룩로리카리아는 이미 오키나와에 정착이 확인되었다. 이들 모두 토착 어류와의 경쟁이 우려되며, 이들 3종은 일본 환경성이 지정하는 주의 외래생물 리스트에 게재되어 있다.[87]
5. 5. 문화
일본 신토 신화에서 자연 현상은 가미에 의해 발생한다고 여겨진다. 지진은 나마즈라고 불리는 거대한 메기에 의해 발생한다고 알려져 있다. 지진과 관련된 다른 가미도 존재한다. 교토에서는 보통 뱀장어이지만, 1855년 에도 지진 이후에는 鯰絵|나마즈에일본어("메기 그림")가 인쇄되어 16세기 오쓰에부터 알려진 메기 가미의 인기가 높아졌다.[81] 한 메기 그림에서는 아마테라스의 신성한 흰 말이 지진을 일으키는 메기를 쓰러뜨리는 모습이 묘사되어 있다.[82]참조
[1]
논문
Phylogenetic classification of living and fossil ray-finned fishes (Actinopterygii)
2024-04-18
[2]
논문
A saharan fossil and the dawn of Neotropical armoured catfishes in Gondwana
[3]
FishBase
[4]
논문
Phylogenetic Relationships of Five Asian Schilbid Genera Including ''Clupisoma'' (Siluriformes: Schilbeidae)
[5]
웹사이트
Catfish Varieties
http://animal-world.[...]
[6]
웹사이트
How Nocturnal Catfish Stalk Their Prey
http://www.scientifi[...]
2001-06-06
[7]
논문
A phylogenetic analysis of the major groups of catfish (Teleostei: Siluriformes) using rag1 and rag2 nuclear gene sequences
[8]
서적
Fishes of the World
John Wiley & Sons, Inc
[9]
논문
Checklist of catfish, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types
http://mapress.com/z[...]
2009-06-22
[10]
웹사이트
Siluriformes
http://tolweb.org/tr[...]
Tree of Life Web Project
2007-04-18
[11]
논문
Upper Cretaceous teleostean otoliths from the Severn Formation (Maastrichtian) of Maryland, USA, with an unusual occurrence of Siluriformes and Beryciformes and the oldest Atlantic coast Gadiformes
2021-09-01
[12]
서적
The Fossil Record 2
Chapman & Hall
[13]
간행물
Evolutionary Histories of Freshwater Fishes
http://dx.doi.org/10[...]
Elsevier
2024-05-08
[14]
논문
Comment on “A Saharan fossil and the dawn of Neotropical armoured catfishes in Gondwana” by Brito et al
[15]
논문
A reply to a comment on Brito et al., 2024, A Saharan fossil and the dawn of the Neotropical armoured catfishes in Gondwana by Britz, Pinion, Kubicek and Conway
[16]
웹사이트
Catfish Families
http://silurus.acnat[...]
All Catfish Species Inventory
2007-04-28
[17]
FishBase
[18]
ITIS
2016-09-10
[19]
논문
Revision of the endemic Malagasy catfish family Anchariidae (Teleostei: Siluriformes), with descriptions of a new genus and three new species
http://www.pfeil-ver[...]
[20]
논문
Neotropical catfish diversity: an historical perspective
[21]
논문
''Lacantunia enigmatica'' (Teleostei: Siluriformes) a new and phylogenetically puzzling freshwater fish from Mesoamerica
http://biostor.org/r[...]
2009-06-22
[22]
논문
Genome-wide interrogation advances resolution of recalcitrant groups in the tree of life
2017-01-13
[23]
논문
Evolutionary Origin and Early Biogeography of Otophysan Fishes (Ostariophysi: Teleostei)
2013-04-09
[24]
논문
Back to the roots: Reducing evolutionary rate heterogeneity among sequences gives support for the early morphological hypothesis of the root of Siluriformes (Teleostei: Ostariophysi)
2018-10
[25]
논문
Phylogeny, origin and biogeography of catfishes: support for a Pangean origin of 'modern teleosts' and reexamination of some Mesozoic Pangean connections between the Gondwanan and Laurasian supercontinents
2004-11-01
[26]
서적
The origin of higher clades : osteology, myology, phylogeny and evolution of bony fishes and the rise of tetrapods
Science Publishers
2007
[27]
논문
GONORYNCHIFORMES AND OSTARIOPHYSAN RELATIONSHIPS: A COMPREHENSIVE REVIEW - Edited by T. Grande, F. J. Poyato-Ariza and R. Diogo
2011-04
[28]
논문
Evolutionary history of Otophysi (Teleostei), a major clade of the modern freshwater fishes: Pangaean origin and Mesozoic radiation
2011-06-22
[29]
논문
LSX: automated reduction of gene-specific lineage evolutionary rate heterogeneity for multi-gene phylogeny inference
Springer Science and Business Media LLC
2019-08-13
[30]
논문
Discovery of African roots for the Mesoamerican Chiapas catfish, ''Lacantunia enigmatica'', requires an ancient intercontinental passage
https://webspace.ute[...]
2009-06-22
[31]
논문
Phylogenetic classification of bony fishes
[32]
서적
Fishes of the World
John Wiley & Sons
[33]
논문
Alternative life-history strategies of catfishes
[34]
논문
Morphological Adaptations of the Texas Blind Catfishes ''Trogloglanis pattersoni'' and ''Satan eurystomus'' (Siluriformes: Ictaluridae) to Their Underground Environment
[35]
논문
Mexican blindcats genus ''Prietella'' (Siluriformes: Ictaluridae): an overview of recent explorations
[36]
FishBase
[37]
서적
Brackish Water Fishes
TFH
[38]
서적
Brackish Water Fishes
Aqualog
[39]
웹사이트
Texas Dept. Wildlife
http://www.tpwd.stat[...]
2012-01-03
[40]
웹사이트
Catfish
https://www.friendso[...]
2023-06-14
[41]
논문
The South American Suckermouth Armored Catfish, Pterygoplichthys anisitsi (Pisces: Loricaridae), in Texas, with Comments on Foreign Fish Introductions in the American Southwest
[42]
논문
Amazon Sailfin Catfish, Pterygoplichthys pardalis (Castelnau, 1855) (Loricariidae), Another Exotic Species Established in Southeastern Mexico
[43]
논문
New Philippine record of South American sailfin catfishes (Pisces: Loricaridae)
http://www.mapress.c[...]
2009-06-22
[44]
논문
The South American Sailfin Armored Catfish, Liposarcus multiradiatus (Hancock), a New Exotic Established in Puerto Rican Fresh Waters
http://academic.uprm[...]
2009-06-22
[45]
논문
Size Structure, Reproductive Phenology, and Sex Ratio of an Exotic Armored Catfish (Liposarcus multiradiatus) in the Kaoping River of Southern Taiwan
http://zoolstud.sini[...]
2009-06-22
[46]
서적
Chemical senses, chemical signals, and feeding behavior in fishes
https://books.google[...]
[47]
논문
Micromyzon akamai, gen. et sp. nov., a small and eyeless banjo catfish (Siluriformes: Aspredinidae) from the river channels of the lower Amazon basin
[48]
웹사이트
Channel Catfish
http://www.fcps.k12.[...]
Fairfax County Public Schools
2006-12-02
[49]
논문
Diversity, phylogenetic distribution, and origins of venomous catfishes
2009-12-04
[50]
논문
A standardized terminology of spines in the order Siluriformes (Actinopterygii: Ostariophysi)
https://doi.org/10.1[...]
2022-02-10
[51]
논문
First description of small juveniles of the primitive catfish Diplomystes (Siluriformes: Diplomystidae)
https://mansfield.os[...]
2023-03-27
[52]
논문
Synodontis acanthoperca, a new species from the Ogôoué River system, Gabon with comments on spiny ornamentation and sexual dimorphism in mochokid catfishes (Siluriformes: Mochokidae)
http://www.mapress.c[...]
2009-06-22
[53]
논문
Variation of male reproductive apparatus in relation to fertilization modalities in the catfish families Auchenipteridae and Callichthyidae (Teleostei: Siluriformes)
[54]
뉴스
Schoolgirl nets 9ft monster fish
https://www.telegrap[...]
2009-07-15
[55]
뉴스
Grizzly Bear-Size Catfish Caught in Thailand
https://web.archive.[...]
National Geographic News
2005-06-29
[56]
웹사이트
Term : humeral process
http://filaman.ifm-g[...]
FishBase
[57]
논문
The eyes of suckermouth armoured catfish (Loricariidae, subfamily Hypostomus): pupil response, lenticular longitudinal spherical aberration and retinal topography
http://jeb.biologist[...]
The Journal of Experimental Biology
2002-11-15
[58]
논문
Reproductive apparatus and gametogenesis of Lophiosilurus alexandri Steindachner (Pisces, Teleostei, Siluriformes)
[59]
논문
Reproduction of the surubim catfish (Pisces, Pimelodidae) in the São Francisco River, Pirapora Region, Minas Gerais, Brazil
[60]
논문
Sounds and Sound Production in Fishes
[61]
논문
Variation in Stridulatory Sound Production in the Channel Catfish, Ictalurus punctatus
[62]
논문
Sound-Generating Mechanisms in Fishes: A Unique Diversity in Vertebrates
[63]
논문
Diversity of Sound Production in Fish
[64]
논문
Agonistic Behavior and Acoustic Communication
[65]
웹사이트
Pond Culture of Channel Catfish in the North Central Region
http://www.extension[...]
North Central Regional Aquaculture Center
1993-10
[66]
웹사이트
Catfish Production
http://usda.mannlib.[...]
2017-07-21
[67]
웹사이트
Economy of Scales
https://web.archive.[...]
Stanford University
[68]
뉴스
'Catfish' bred in Asia move up on U.S. food chain
http://www.latimes.c[...]
2006-11-28
[69]
뉴스
Catfish imports not slowing
https://web.archive.[...]
Northwest Arkansas News
2006-01-27
[70]
서적
Simply Fish
Faver & Faber
[71]
웹사이트
Vitamin D and Healthy Bones
http://www.health.st[...]
New York State Department of Health
2007-07-13
[72]
웹사이트
Fatty Fish Not Equal in Good Fats
http://www.fitfuncti[...]
Reuters
2012-03-21
[73]
웹사이트
The Rehab Archipelago Forced Labor and Other Abuses in Drug Detention Centers in Southern Vietnam
https://www.hrw.org/[...]
2011-09-07
[74]
웹사이트
Union Fish Company – Basa/Swai Details
http://www.unionfish[...]
2007-11-11
[75]
웹사이트
United States Code, Title 21, section 321d. Market names for catfish and ginseng
https://www.law.corn[...]
2020-10-28
[75]
웹사이트
United States Code, Title 21, section 343 (t). Misbranded food
https://www.law.corn[...]
2017-05-09
[76]
법원판결
Piazza's Seafood World, LLC v. Odom
https://scholar.goog[...]
2023-09-23
[77]
웹사이트
Fish Labelling (Amendment) (England) Regulations 2006
http://www.legislati[...]
Committee on Toxicity
2007-05-26
[78]
뉴스
How to cook the irresistible catfish pepper soup
http://www.pulse.ng/[...]
2018-05-14
[79]
웹사이트
Kosher Spirit: Fins and Scales
https://www.ok.org/a[...]
2022-11-29
[80]
웹사이트
Channel Catfish
https://fisheries.ta[...]
2019-11-14
[81]
서적
The Mythical Creatures Bible
Sterling
2009
[82]
서적
Seismic Japan: The Long History and Continuing Legacy of the Ansei Edo Earthquake
University of Hawaii Press
2013
[83]
FishBase
2014-11
[84]
웹사이트
IGFA World Records
http://wrec.igfa.org[...]
International Game Fish Association
2015-11-01
[85]
서적
決定版 日本の外来生物
平凡社
2008-04-21
[86]
웹사이트
特定外来生物の解説・チャネルキャットフィッシュ
https://warp.ndl.go.[...]
環境省
2008-10-14
[87]
웹사이트
注意すべき外来生物リスト・魚類
https://warp.ndl.go.[...]
環境省
2008-10-14
[88]
웹사이트
Fisheries and Aquaculture Department
http://www.fao.org/f[...]
FAO
2008-11-11
[89]
저널
Lacantunia enigmatica (Teleostei: Siluriformes) a new and phylogenetically puzzling freshwater fish from Mesoamerica
[90]
웹사이트
All Catfish Species Inventory
http://silurus.acnat[...]
ACSI
2008-10-24
[91]
서적
海南島淡水魚類譜
海南海軍特務部政務局
[92]
웹사이트
Phylogenetic Classification of Bony Fishes – Version 4
https://sites.google[...]
2016
[93]
웹사이트
Study Resolves 50-Year Dispute of Teleost Fishes Ancestral Lineage
https://www.fau.edu/[...]
[94]
웹사이트
Genome structures resolve the early diversification of teleost fishes
https://hal.science/[...]
[95]
저널
A phylogenetic analysis of the major groups of catfishes (Teleostei: Siluriformes) using rag1 and rag2 nuclear gene sequences
[96]
서적
State of the Art of Siluriform Higher-level Phylogeny
Science Publishers
2010
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com