수정 (생물학)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
수정은 생물학에서 생식 세포인 정자와 난자가 결합하여 새로운 개체를 형성하는 과정이다. 이 과정은 감수 분열을 통해 생식 세포가 형성되고, 정자의 형성 및 난자의 성숙을 거쳐 일어난다. 수정은 체외 수정과 체내 수정으로 구분되며, 식물에서는 화분관을 통해 정자가 난세포에 도달하여 수정이 이루어진다. 동물에서는 정자가 난자에 접근하여 수정막을 형성하고, 정핵과 난핵이 융합하여 수정란을 생성한다. 수정은 유전적 재조합을 통해 유전적 다양성을 증가시키며, 단성생식, 타가수정, 자가수정 등 다양한 생식 현상과 관련된다. 인간의 수정은 난관에서 일어나며, 수정란은 자궁에 착상하여 임신을 시작한다.
더 읽어볼만한 페이지
- 꽃가루받이 - 과일
과일은 식용 가능한 열매를 맺는 목본성 식물을 넓게, 과실 수확 목적의 과일나무를 좁게 의미하며, 형태, 생육 환경 등에 따라 나뉘고 단과, 집합과, 다화과 등으로 구분되며, 인류는 오래전부터 재배 및 섭취해왔고 한국에서는 과일 소비를 장려하기 위해 '과일의 날'을 지정했다. - 꽃가루받이 - 자웅이체
자웅이체는 생물이 암컷과 수컷의 성을 뚜렷하게 구분하는 생식 전략으로, 다양한 생물 계통에서 수렴 진화를 통해 나타나며 동물 종의 약 95%에서 관찰된다. - 생식력 - 성교
성교는 생물학적 생식 과정의 한 형태로서, 암수의 생식기 접촉을 통해 정자가 난자를 수정하는 행위를 의미하며, 쾌락이나 애정 표현의 수단으로도 활용되고, 사회문화적 요인에 따라 다양한 형태로 이루어진다. - 생식력 - 출생률
출생률은 특정 기간 동안의 출생아 수를 나타내는 지표이며, 사회, 경제, 문화, 정책 등 다양한 요인에 영향을 받아 세계적으로 감소 추세에 있고, 한국은 심각한 저출산 문제를 겪고 있다. - 생식 - 성교
성교는 생물학적 생식 과정의 한 형태로서, 암수의 생식기 접촉을 통해 정자가 난자를 수정하는 행위를 의미하며, 쾌락이나 애정 표현의 수단으로도 활용되고, 사회문화적 요인에 따라 다양한 형태로 이루어진다. - 생식 - 난태생
난태생은 어미의 몸 안에서 알을 부화시켜 새끼를 낳는 생식 방식으로, 난황낭 난태생과 모체영양 난태생으로 구분되며 난생과 태생의 중간 단계로 여겨진다.
| 수정 (생물학) | |
|---|---|
| 개요 | |
| 설명 | 수정은 성별이 다른 생식 세포가 성적 생식 과정에서 결합하여 접합체를 형성하는 것이다. |
| 관련 용어 | 수정 임신 생식 |
| 상세 내용 | |
| 정의 | 수정은 성별이 다른 생식 세포가 성적 생식 과정에서 결합하여 접합체를 형성하는 과정이다. |
| 생물학적 과정 | 배우자 부착 배우자 융합 |
| 관련 세포 | 정자 난자 |
2. 생식 세포의 형성
생식 세포는 감수 분열을 통해 형성되며, 이 과정에서 유전적 다양성이 확보된다. 감수 분열은 각 부모로부터 유전자를 무작위로 섞이게 하여, 각 생식 세포가 유전적으로 독특하게 만든다.[6] 수정 시 부모의 염색체가 결합하여 새로운 조합을 생성한다.
사람의 경우, 비성염색체만 고려했을 때 (2²²)² = 17.6x1012개의 서로 다른 접합자가 가능하다. 염색체 교차가 일어나지 않는다고 가정해도 이 정도의 다양성이 확보되며, X염색체와 Y염색체는 교차가 일어나지 않아 계산에서 제외된다. 미토콘드리아 DNA는 모계로부터만 유전된다.
2. 1. 감수 분열

감수 분열(수가 줄어듦)은 생식 세포 형성 시 일어나는 세포 분열 방식이다. 난자와 정자의 염색체 수는 체세포가 가지고 있는 염색체 수 46개의 절반인 23개이며, 생식 세포의 염색체를 n, 체세포의 염색체를 2n으로 표시한다. 생식 세포가 형성될 때는 염색체 수가 반감되므로, 이와 같은 분열을 '감수 분열'이라고 한다. 감수 분열에서는 2회의 핵분열이 연속해서 일어난다.
감수 분열은 세포가 2회 연속 분열을 함으로써 이루어지는데, 보통 제1분열에서는 염색체 수가 반감되고, 제2분열에서는 염색체 수에 변화가 없다. 제1분열의 전기에는 상동 염색체가 접합하여 굵은 2가 염색체가 되고, 그 각 염색체는 다시 세로로 갈라져서 4개의 염색분체(4분 염색체)가 된다. 2가 염색체는 중기에 적도면에 배열되고, 후기에는 접합면에서 갈라져 양극으로 끌려가는데, 이 때 염색체 수가 반감된다. 양극으로 끌려온 염색체(n) 주위에는 전기에 소실되었던 핵막이 다시 생기며, 반수의 염색체를 갖는 2개의 딸세포가 생긴다. 이 두 딸세포는 곧 제2분열을 하게 된다. 제2분열에서는 각 염색체가 2분되어 염색체 수가 n인 딸세포 4개가 된다.
수정의 진화는 감수분열의 기원과 관련이 있으며, 둘 다 유성생식의 일부로 진핵생물에서 유래했다. 한 가설은 감수 분열이 유사분열에서 유래했다고 주장한다.[6] 감수 분열은 각 부모가 기여하는 유전자의 무작위 분리를 초래한다.
2. 2. 정자의 형성
정원 세포는 생식소에서 세포 분열을 반복하여 수가 증가한다. 정원 세포는 분열을 중지하고 성숙하여 제1정모 세포가 된다. 제1정모 세포는 감수 분열(제1분열)을 하여 2개의 제2정모 세포가 되고, 제2정모 세포는 다시 분열(제2분열)을 하여 4개의 정세포가 된다. 이들 정세포는 분화하여 운동 능력이 있는 정자가 된다.[1]사람이나 개의 정자는 올챙이 모양으로, 머리부·중편·꼬리부로 구별할 수 있다. 머리부는 골지체에서 유래하는 첨체와 핵으로 되어 있고, 중편에는 주로 미토콘드리아가 있어서 정자의 운동에 필요한 에너지를 제공한다. 꼬리부는 중편의 앞 끝에 있던 중심체가 변형된 편모로서, 이 편모의 중심에는 두 다발, 주변에는 아홉 다발의 섬유가 있다.[1]

정자의 모양은 사람의 경우처럼 올챙이 모양인 것이 잘 알려져 있는데, 이와 달리 쥐의 정자는 머리부가 낫 모양이고, 오리너구리 등 단공류의 정자는 선충 모양, 도롱뇽은 트리파노소마 모양의 정자를 지니고 있다. 이 밖에도, 갑각류의 별사탕 모양, 곤충의 화살촉 모양, 흡충류의 실모양, 선충류의 별사탕·오똑이 모양 등 여러 가지 모양의 정자가 있다. 또 곤충류와 연체동물에서는 모양과 크기가 다른 2종류의 정자를 낳는 일이 있는데, 이러한 정자를 '이형 정자'라고 부른다.[1]
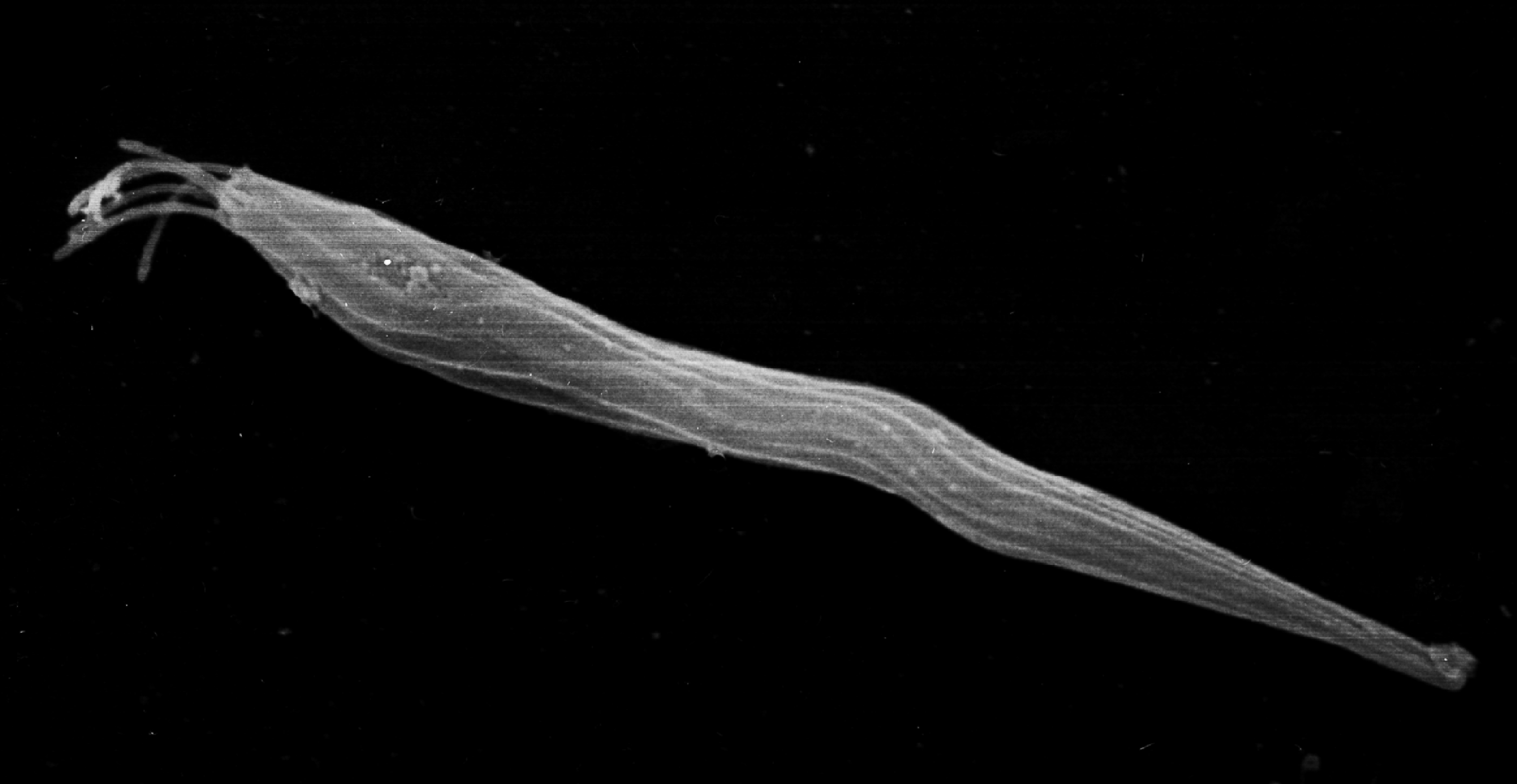
2. 3. 난자의 형성
정자 형성과 마찬가지로, 난원 세포는 난소에서 분열을 반복한 다음, 분열을 중지하고 세포질 속에 영양분인 난황을 축적하여 비대해진다. 이 과정은 정자 형성에 비하여 매우 두드러지게 나타나므로 '성장기'라고 한다. 성장기 후, 충분히 영양을 축적한 세포는 제1난모 세포가 된다. 제1난모 세포는 부등 분열(제1분열)에 의해, 모세포 세포질의 대부분을 차지하는 1개의 제2난모 세포와 극히 소량의 세포질을 포함하는 1개의 제1극체가 된다. 이어서 제2난모 세포는 제2분열을 하여 난자(난세포)와 극체(제2극체)로 되고, 제1극체도 분열을 하여 2개의 극체가 된다. 그러므로 1개의 제1난모 세포는 감수 분열을 하여 1개의 난자와 3개의 극체를 만드는 셈이다.그러나 제1극체가 분열하지 않는 종류도 있는데, 이 때는 1개의 난자와 2개의 극체가 생기게 된다. 분열에 의해 생성된 극체는 발생 능력이 없어 얼마 후에 소실되므로, 결국 1개의 제1난모 세포에서 1개의 난자가 생긴다. 이러한 과정을 거쳐 형성된 난자는 장래의 배 발생에 필요한 난황을 다량 함유하고 있어서, 정자에 비하면 매우 크다. 그러나 태반을 지니는 포유류의 배는 모체에서 영양분이 공급되므로 난자가 비교적 작은데, 그래도 정자에 비하면 훨씬 큰 편이다. 예를 들면, 고양이의 정자는 길이가 약 40μm인데 비해, 난자의 지름은 약 130μm이다. 또한, 조류나 파충류의 알은 매우 크며, 특히 타조의 알은 긴지름이 20cm에 달한다. 그 밖에 상어나 가오리 등의 연골어류를 비롯하여 경골어류의 알에도 큰 것이 많다. 무척추동물의 알은 몸의 크기에 비하여 큰 것이 많은데, 특히 극피동물과 연체동물의 알에는 지름이 수 mm에 달하는 것도 있다.
3. 수정의 과정
수정은 정자와 난자가 만나 하나의 수정란을 형성하는 과정이다.
고대 아리스토텔레스는 암수의 체액이 융합하여 새로운 개체가 형성되며, 형태와 기능이 점진적으로 나타난다고 보았다. 이를 후생설이라 불렀다.[4]
1784년, 스팔란차니는 개구리 실험을 통해 접합자 형성에 암컷의 난자와 수컷의 정자가 모두 필요함을 밝혔다.[5] 1827년, 카를 에른스트 폰 베어는 포유류의 난자를 최초로 관찰했다.[4] 1876년, 오스카 헤르트비히는 성게의 정자와 난자 핵융합을 설명했다.[5]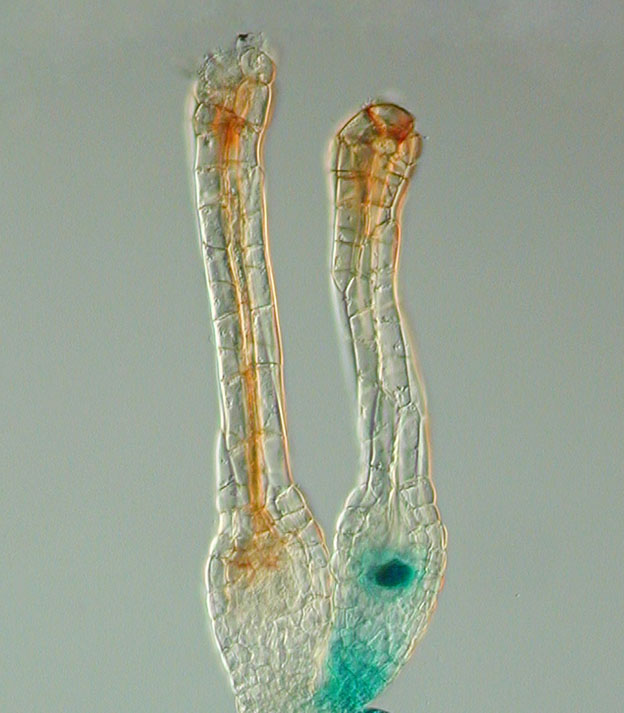
식물의 경우, 선태식물과 양치식물은 장정기 내에서 수정이 일어난다. 종자식물은 수컷 배우체가 화분 알갱이 내에서 형성되며, 수분 후 화분관이 자라 배주의 주공을 통해 정자가 이동하여 난세포와 수정한다. 속씨식물에서는 두 개의 정세포가 방출되어 두 번의 수정이 일어나는 이중 수정이 일어난다.[7]
수정 후 씨방은 부풀어 과일이 된다.[10] 꽃가루관 성장은 영양 세포질에 의해 조절되며, 꽃가루관은 암술 조직을 분해하며 자라고, 이때 생성된 영양분을 이용한다. 성장하는 꽃가루관에는 3개의 핵(2개의 정핵, 1개의 관핵)이 존재한다.[12] 정자는 상호 연결되어 있으며, 큰 정자는 관핵에도 연결되어 '수컷 생식단위'를 형성한다.[13]
피자식물의 이중 수정은 두 정자가 배낭의 두 세포를 수정하는 과정이다. 하나의 정자는 난세포와 수정하여 이배체 접합자를 형성하고, 다른 정자는 두 개의 반수체 극핵과 융합하여 삼배체 배젖을 형성한다.[7] Nuphar polysepala와 같은 원시 꽃피는 식물은 이배체 배젖을 가지기도 한다.[14]
수박과 같은 식물은 정상적인 과일 형성을 위해 많은 꽃가루가 필요하다.
동물 수정 연구는 성게와 생쥐를 대상으로 활발히 진행되었으며, 정자와 난자의 만남, 단일 정자 침입 기작 등을 연구한다. 수정은 화학주성, 정자 활성화/첨체 반응, 정자/난자 부착의 3단계로 이루어진다.
정자는 난자의 표면으로 유인되며, 난자와 정자가 놓인 경우 매우 빠르게 난자 표면에 모인다. 정자가 난자 표면에 침입하면, 침입 지점을 중심으로 난자 표면에서 투명한 '''수정막'''이 솟아오른다. 수정막은 다른 정자의 추가 침입을 막는다.
일반적으로 난자에 침입하는 정자는 하나뿐이다. 침입한 정자의 핵은 난핵과 융합하여 수정을 완료한다. 여러 정자가 침입하더라도 난핵과 융합하는 것은 하나뿐이며, 나머지는 흡수되거나 난할을 비정상적으로 만든다.
수정 후, 난할이 시작되며, 성게 등에서는 수정막이 배를 감싸는 막이 되고, 배가 이 막에서 탈출하는 것을 부화라고 한다.
유전학적으로 수정은 배우자에서 유래하는 정핵과 난핵의 융합 현상이다. 발생학적으로는 정자가 난자에 접촉하는 시점부터 난자의 변화가 시작되므로, 이 시점을 수정의 순간으로 볼 수도 있다. 실제로 난자 표면에 화학적 자극을 주는 것만으로 발생이 시작되는 경우도 있으며, 수정에서 정자의 역할은 난자 발생의 방아쇠를 당기는 것이라는 견해도 있다.
3. 1. 체외 수정과 체내 수정
체외 수정은 몸 밖에서 난자와 정자가 수정하는 것이고, 체내 수정은 암컷의 몸 속에서 수정하는 것이다. 굴이나 모시조개처럼 무척추동물 중에는 체외 수정을 하는 종류가 많은데, 여름에 수온이 높아지면 알과 정자를 물 속에 방출하여 물 속에서 수정이 이루어진다. 이러한 체외 수정은 강장동물, 유형동물, 환형동물의 다모류, 연체동물의 부족류, 극피동물 등에서 널리 볼 수 있다.[50]그러나 절지동물의 곤충·거미 및 환형동물의 지렁이 등 육지에 사는 동물과 회충 등의 기생 동물은 일반적으로 체내 수정을 한다. 권패류나 편형동물은 수생(水生)인데도 체내 수정을 한다. 문어·오징어 등의 두족류와 게·새우 등의 갑각류는 교미 기관을 통해 교미를 하지만, 정자가 암컷의 체내에 들어가 '정협(精莢)'이라는 자루 속에 들어가 수란관 입구 부근에 부착했다가 암컷이 산란할 때 정협이 터지면서 정자가 나와 물 속에서 수정이 이루어지기 때문에 실제 수정은 체외에서 이루어진다.[50]
척추동물 중 원구류·경골어류·양서류의 대부분은 체외 수정을 하며, 파충류·조류는 일반적으로 체내 수정을 한다. 개구리는 교미에 가까운 행동을 하나, 수정은 체외에서 한다. 어류 중 가오리나 상어 등의 판새류와 볼락·망상어·거피 등의 난태생인 종류에서는 교미가 행해진다. 뱀이나 도마뱀의 일부 종류와 박쥐 등은 정자가 암컷의 체내에 수개월에서 수년 동안이나 수정 능력을 지닌 채로 보존되므로 한 번의 교미만으로도 몇 번의 생식기에 걸쳐 수정란을 낳을 수 있다.[50]
동물에서 수정에 이르는 과정은 각 개체의 관계에 따라 체외수정과 체내수정으로 구분된다. 체외수정은 각 개체가 난자와 정자를 체외로 방출하여 그곳에서 수정이 이루어지는 것이고, 체내수정은 난자가 체내에 남아 그곳에 정자가 도달하여 수정이 이루어지는 것이다.
포유류는 교미(copulation)를 통해 체내 수정을 한다.[22] 수컷이 사정(ejaculation)을 하면, 많은 정자가 질 수축을 통해 질 상부로 이동하고, 자궁경부(cervix)를 지나 자궁(uterus) 전체를 가로질러 난자를 만난다.
3. 2. 수정의 메커니즘
식물의 정자는 사과산이나 설탕과 같은 물질에 대하여 양성 주화성을 나타내어, 그것에 의하여 난자에 접근하기도 한다. 하지만 대부분의 동물은 난자의 표면이나 젤리층과 같은 부속 구조에 정자를 활성화시키는 물질이 있어서 그것에 의하여 정자가 접근한다. 극피동물을 비롯한 많은 해산 무척추동물은 난자에서 정자를 응집시키는 물질, 즉 '수정소'를 바닷물 속으로 방출한다.[52]수정소는 단백질로서, 정자 표면에 존재하는 단백질과 특이하게 결합하여, 종류를 구별하고 수정이 이루어지게 하는 데 어떤 작용을 한다. 어류의 알은 대개 두꺼운 난막으로 둘러싸여 있는데, 정자는 난막에 있는 작은 구멍인 난문(卵門)을 통하여 수정한다. 즉, 정자의 운동을 활발하게 하는 물질이 난문부에 있어서, 정자가 이 난문 근처에 오면 갑자기 운동이 활발해져서 난자 속으로 침입하게 된다. 포유류는 수란관의 섬모 운동과 정자의 운동으로 수란관 상단까지 헤엄쳐 가서 난자와 수정한다. 이 밖에 편형동물, 환형동물의 거머리, 회충·요충·십이지장충 등의 구충에서도 특이하게 정자가 난자에 접근한다. 특히, 편형동물 와충강에 속하는 삼기장류와 봉장류는 교미할 때 수컷의 음경이 암컷의 체벽을 뚫고 들어가 피하의 결합 조직 속으로 정자를 방출하는데, 이들의 정사는 수정낭에 모여 축적된다. 또, 거머리류는 교미할 때 정협이 암컷의 몸 표면에 부착하고 얼마 후 그 부분의 피부가 붕괴하여 그곳으로부터 정자가 몸속으로 들어간다. 이러한 현상은 구충에서도 잘 알려져 있다.[52]
한편, 어떤 종류의 정자는 난자 주위에 존재하는 젤리층과 투명층을 용해하는 효소를 지니고 있다. 이 정자가 난자에 접근하여, 특히 젤리층에 접하게 되면 정자의 첨체에서 가느다란 실 모양의 첨체 돌기가 나와 난막에 부착하여, 마침내 난막이 녹아 난자 안으로 정자가 침입해 들어가는데, 이것을 '첨체 반응'이라고 한다. 이때, 난자에 도달한 정자가 방출하는 항정소는 나머지 정자들의 접근을 방해하여 다른 정자의 침입을 막는 구실을 한다. 불가사리류의 경우, 정자가 난막에 도달하면 난막의 표면이 솟아올라 수정 돌기가 형성되는데, 특히 첨체 돌기가 붙는 부분을 수정구라고 하며, 정자는 이 수정구를 통하여 난자 속으로 들어간다.[52]
수정구를 통하여 정자의 머리부가 난자 속으로 들어가면, 얇은 수정막이 형성되어 난자 전체를 덮는다. 성게와 송사리의 알에서 관찰한 수정막의 형성 과정은 다음과 같다.[53]
- 미수정란의 정자가 접촉한 부분에서 알의 표면을 따라 자극이 전해진다.
- 이 자극에 의해 난자의 표면 바로 밑에 존재하는 표층포(表層胞)라 불리는 과립이 파괴된다.
- 이때, 과립 속에 포함되어 있던 콜로이드 모양의 물질이 알의 원형질막과 난막 사이에 방출된다.
- 콜로이드의 친수성에 의해 물이 들어오면, 그 압력으로 난막을 밀어올리게 된다.
- 이때 밀어올려진 막을 '수정막'이라 하며, 난자의 표면과 수정막 가까이의 공간을 '위난강'이라 한다.
- 수정막은 얼마 후 굳어져서 난자를 보호할 뿐만 아니라 정자가 방출한 항정소와 함께 다른 정자의 침입을 막는 구실을 한다.
- 난자 속으로 들어간 정자의 머리부는 형체가 불확실해지고 포상(胞狀)을 이루어 '웅성 전핵(雄性前核)'이 된다.
- 난핵은 일반적으로 감수분열의 제2분열 도중에 분열이 중지되는데, 정자가 침입하면 분열은 다시 진행되어 제2극체가 성숙된다. 이것을 '자성 전핵(雌性前核)'이라고 한다.
- 웅성 전핵과 자성 전핵은 그 둘레에 성상체를 형성한다.
- 정핵은 난핵 쪽으로 이동을 하여 핵융합이 일어나는데, 이로써 수정은 끝난다.
핵융합이 일어난 '합핵'은 얼마 후 세포 분열을 반복하는 난할을 시작하므로, 이 시기부터 수정란의 발생이 진행된다.[53]
미수정란에 인위적으로 자극을 주어 단독으로 발생시키는 것을, 자연 상태에서의 단위 생식에 대하여 '인공 단위 발생' 또는 '인공 단위 생식'이라고 한다. 19세기 후반에, 누에의 미수정란을 마찰시키거나 산으로 처리하면, 배(胚)까지 발생이 진행된다는 사실이 보고되었으며, 그 후에도 인공 단위 발생의 예는 많이 알려졌다.[54]
수정의 기전은 성게와 생쥐를 대상으로 광범위하게 연구되어 왔다. 이 연구는 정자와 적절한 난자가 서로 어떻게 찾아내는지, 그리고 어떻게 단 하나의 정자만이 난자에 들어가 그 내용물을 전달하는지에 대한 질문을 다룬다. 종 특이성을 보장하는 수정에는 세 가지 단계가 있다.
# 화학주성
# 정자 활성화/첨체 반응
# 정자/난자 부착
정자는 화학주성이라는 리간드/수용체 상호작용을 통해 난자를 찾는다. 레색트(Resact)는 ''A. punctulata''의 난황막에서 정제된 14개의 아미노산으로 구성된 펩타이드로, 정자의 이동을 유인한다.
난자를 찾은 후, 정자는 정자 활성화라는 과정을 통해 젤리층을 통과한다. 또 다른 리간드/수용체 상호작용에서 난자의 올리고당 성분이 정자의 수용체에 결합하여 활성화시키고 수정소 반응을 일으킨다. 정자의 수정소 소포는 세포막과 융합되어 방출된다. 이 과정에서 바인딘(bindin)과 같이 수정소 소포막에 결합된 분자가 정자 표면에 노출된다. 이러한 내용물은 젤리층과 결국 난막을 소화한다. 수정소 소포의 방출 외에도, 정자 머리에 가는 돌기인 수정돌기를 형성하기 위해 액틴의 폭발적인 중합이 일어난다.
정자는 난막의 수용체 사이의 또 다른 리간드 반응을 통해 난자에 결합한다. 정자 표면 단백질인 바인딘(bindin)은 EBR1로 확인된 난막의 수용체에 결합한다.
정자와 난자의 세포막 융합은 바인딘(bindin)에 의해 매개될 가능성이 높다. 접촉 지점에서 융합은 수정 원추의 형성을 유발한다.
일반적으로 정자는 난자의 표면으로 유인된다. 연속된 액체 속에 난자와 정자가 놓인 경우, 정자는 매우 빠르게 난자의 표면에 모인다. 정자가 난자의 표면으로 침입하면, 그 직후에 침입 지점을 중심으로 난자의 표면에서 투명한 막이 솟아오른다. 이 막을 '''수정막'''이라고 한다. 수정막이 생기면 다른 정자는 난자의 표면에서 멀어진다.
난자에 침입하는 정자의 수는 일반적으로 하나뿐이다. 침입한 정자의 핵은 난핵에 접근하고, 결국 두 핵이 융합하여 수정이 완료된다. 여러 개가 침입하더라도 난핵과 융합할 수 있는 것은 하나뿐이므로, 나머지 정자는 갈 곳이 없다. 난자에 흡수되는 경우도 있지만, 성게에서는 여러 개의 정자가 침입하면 난할이 비정상적으로 되는 것이 알려져 있다. 보통은 여러 개의 정자가 침입하는 일은 없다. 이것은 수정막의 작용이라고 생각되기도 했지만, 반드시 그렇지 않다는 설도 있다.
수정 후, 바로 난할이 시작되는 경우가 많다. 성게 등에서는 수정막은 그대로 배를 감싸는 막이 되고, 거기서 탈출하는 것을 부화라고 한다.
유전학적으로 보면, 수정은 배우자로부터 유래하는 현상이며, 정핵과 난핵의 융합을 수정의 순간으로 보는 것이 옳다. 발생학적으로 보면, 새로운 개체의 핵이 형성되는 것이 두 핵의 융합이므로, 그것을 수정의 순간으로 볼 수도 있지만, 정자가 난자에 접촉한 시점에서 난자의 변화가 시작되므로, 그것을 수정의 순간으로 볼 수도 있다. 실제로 난자의 표면에 화학적인 자극을 준 것만으로 발생이 시작되는 예도 있으며, 그런 면에서 보면 수정에서 정자의 역할은 난자의 발생의 방아쇠를 당기는 것이라는 견해도 성립한다.
4. 다양한 생물에서의 수정
척추동물은 출산 방식에 따라 내부 수정 또는 외부 수정을 사용한다. 난생 동물은 두꺼운 칼슘 껍질이나 가죽 껍질을 가진 알을 낳기 때문에 정자가 알의 두꺼운 보호층을 통과할 필요 없이 내부 수정을 통해 번식한다. 난태생 및 태생 동물 또한 내부 수정을 사용한다. 일부 양서류처럼 포접을 통해 번식하는 경우도 내부 수정을 사용할 수 있다. 내부 수정은 배우자 낭비를 최소화하고, 개별 알 수정 가능성을 높이며, 알 보호 기간을 연장하고, 선택적 수정이 가능하다는 장점이 있다. 많은 암컷은 장기간 정자를 저장하여 원하는 시기에 알을 수정할 수 있다.
반면 얇은 막이나 막이 없는 알을 낳는 난생 동물은 외부 수정 방법을 사용하며, 이들은 난황생으로 불리기도 한다.[21] 외부 수정은 접촉을 최소화하여 질병 전염 위험을 줄이고 유전적 변이가 더 크다는 장점이 있다.
4. 1. 식물의 수정
식물 수정에 관여하는 배우자는 정자(수컷)와 난세포(암컷)이다. 다양한 식물 그룹은 수컷과 암컷 배우체가 생성한 배우자가 만나 수정되는 방법이 다르다. 선태식물과 양치식물 지상 식물에서 정자와 난세포의 수정은 장정기 내에서 일어난다.[7] 종자식물에서 수컷 배우체는 화분 알갱이 내에서 형성된다. 수분 후, 화분 알갱이가 발아하고, 화분관이 자라 배주의 주공이라고 하는 작은 구멍을 통해 침투한다. 정자는 화분관을 통해 화분에서 배주로 이동하여 난세포가 수정된다. 속씨식물에서는 두 개의 정세포가 화분관에서 방출되고, 두 번째 정세포와 배주의 중심 세포(두 번째 암 배우자)가 관여하는 두 번째 수정 사건이 발생한다.[7]
동물 정자와 달리 대부분의 종자식물의 정자는 운동성이 없으며, 정자를 난자에 전달하고 방출하는 데 꽃가루관에 의존한다.[8] 꽃가루관은 암술머리를 관통하여 씨방에 도달하기 전에 암술대의 세포외 기질을 통해 신장한다. 그런 다음 화탁 근처에서 배주의 주피(배주 벽의 개구부)를 뚫고 꽃가루관이 배낭으로 "터져" 정자를 방출한다.[9] 꽃가루관의 성장은 암술의 화학적 신호에 의존하는 것으로 여겨져 왔지만, 이러한 메커니즘은 1995년까지 거의 알려지지 않았다. 담배 식물에서 수행된 연구는 꽃가루관의 성장을 촉진하는 TTS 단백질이라는 당단백질 계열을 밝혀냈다.[9] 무당 성분 배지와 정제된 TTS 단백질이 포함된 배지에서 모두 꽃가루관이 자랐다. 그러나 TTS 배지에서는 무당 배지보다 3배 빠른 속도로 자랐다.[9] TTS 단백질은 반생체 내 수분된 암술의 여러 위치에 놓였고, 꽃가루관이 단백질 쪽으로 즉시 신장하는 것이 관찰되었다. TTS 단백질을 생성할 수 없는 형질전환 식물은 꽃가루관 성장이 느리고 생식력이 감소했다.[9]
애기장대에서 정자를 방출하기 위한 꽃가루관의 파열은 암배우체에서 오는 신호에 의존하는 것으로 나타났다. 밑씨에 존재하는 FER 단백질 키나아제라고 불리는 특정 단백질은 활성산소종이라고 불리는 매우 반응성이 높은 산소 유도체의 생성을 조절한다. GFP를 통해 ROS 수치는 밑씨가 꽃가루관에 가장 수용적인 꽃의 단계에서 가장 높고, 발달 과정과 수정 후에는 가장 낮은 것으로 나타났다.[8] 높은 농도의 ROS는 꽃가루관에서 칼슘 이온 채널을 활성화시켜 이러한 채널이 다량의 칼슘 이온을 흡수하게 한다. 이러한 칼슘 흡수량의 증가는 꽃가루관이 파열되어 정자를 밑씨에 방출하게 한다.[8] 식물에 디페닐 아이오도늄 클로라이드(DPI)를 공급하는 암술 영양 공급 분석에서 애기장대의 ROS 농도가 억제되어 꽃가루관 파열이 억제되었다.[8]
수정이 된 후, 씨방은 부풀어 오르기 시작하여 과일이 된다.[10] 여러 개의 씨앗이 있는 과일의 경우, 각 배주와의 수정을 위해서는 여러 개의 꽃가루가 필요하다. 꽃가루관의 성장은 영양(또는 관) 세포질에 의해 조절된다. 꽃가루관이 암술머리와 암술대를 따라 자라면서 가수분해 효소가 분비되어 암술 조직을 분해하고, 분해된 조직은 꽃가루관이 자라는 데 필요한 영양분으로 사용된다. 씨방으로 향하는 꽃가루관의 성장 동안, 생식핵은 두 개의 별개의 정핵(염색체의 반수체 수)으로 분열한다.[11] 따라서 성장하는 꽃가루관에는 세 개의 별개의 핵, 즉 두 개의 정핵과 한 개의 관핵이 포함되어 있다.[12] 정자는 상호 연결되어 있으며 이형이며, 많은 식물에서 큰 정자는 관핵에도 연결되어 있으며, 상호 연결된 정자와 관핵은 "수컷 생식단위"를 형성한다.[13]
이중 수정은 피자식물(꽃피는 식물)에서 일어나는 과정으로, 각 꽃가루관의 두 정자가 배주 내부에 있는 암 배우체(때로는 배낭이라고도 함)의 두 세포를 수정한다. 꽃가루관이 배우체에 들어가면 꽃가루관 핵은 붕괴되고 두 개의 정세포가 방출된다. 두 정세포 중 하나는 난세포(주피 근처 배우체 하단에 위치)를 수정하여 이배체(2n) 접합자를 형성한다. 이것이 수정이 실제로 일어나는 시점이며, 수분과 수정은 별개의 과정이다. 다른 정세포의 핵은 배우체 중앙에 있는 두 개의 반수체 극핵(중앙 세포에 포함됨)과 융합한다. 그 결과 생성된 세포는 삼배체(3n)이다. 이 삼배체 세포는 유사분열을 통해 분열하여 종자 내부의 영양분이 풍부한 조직인 배젖을 형성한다.[7] 배젖에 기여하는 두 개의 중앙 세포 모계 핵(극핵)은 난세포를 생성한 단일 감수 분열 생성물에서 유사분열에 의해 발생한다. 따라서 삼배체 배젖의 유전적 구성에 대한 모계 기여는 배의 기여의 두 배이다.
꽃피는 식물의 한 원시 종인 Nuphar polysepala는 두 개가 아닌 하나의 모계 핵과 정자가 융합하여 형성된 이배체 배젖을 가지고 있다. 피자식물 계통의 초기 발달 단계에서 이러한 번식 방식의 중복이 일어나 7세포/8핵 암 배우체와 2:1의 모계 대 부계 게놈 비율을 가진 삼배체 배젖이 생성된 것으로 여겨진다.[14]
많은 식물에서 과육의 발달은 수정된 배주의 비율에 비례한다. 예를 들어 수박의 경우, 정상적인 크기와 모양의 과일을 만들려면 약 1,000개의 꽃가루가 암술머리의 세 개의 조각에 고르게 퍼져야 한다.
타가수분 또는 타가수정과 자가수분은 서로 다른 이점과 비용을 가진 다른 전략을 나타낸다. 식물 종의 약 48.7%는 암수딴그루 또는 자가불화합성 의무 타가수분 식물이다.[15] 또한, 약 42%의 꽃피는 식물이 자연에서 혼합 교배 시스템을 나타내는 것으로 추정된다.[16]
가장 일반적인 유형의 혼합 교배 시스템에서 개별 식물은 단일 유형의 꽃을 생산하며, 열매에는 자가수정된 것, 타가수정된 것 또는 혼합된 자손 유형이 포함될 수 있다. 타가수정에서 자가수정으로의 전환은 식물에서 가장 흔한 진화적 전환이며, 많은 독립적인 계통에서 반복적으로 발생했다.[17] 꽃피는 식물의 약 10~15%는 주로 자가수정한다.[17]
수분 매개체 또는 배우자가 드문 환경에서는 자가수정이 생식 확보의 이점을 제공한다.[17] 따라서 자가수정은 개척 능력을 향상시킬 수 있다. 어떤 종에서는 자가수정이 여러 세대에 걸쳐 지속되었다. 애기장대속의 붉은애기장대는 5만 년에서 10만 년 전에 자가화합성이 된 자가수정 종이다.[18] 애기장대는 야생에서 타가수정 비율이 0.3% 미만인 주로 자가수정하는 식물이다.[19] 한 연구에 따르면 자가수정은 애기장대에서 약 100만 년 전 또는 그 이상에 진화한 것으로 제시되었다.[20] 오랫동안 자가수정해 온 식물에서 해로운 돌연변이의 가림과 유전적 변이성의 생성은 빈번하지 않으므로 여러 세대에 걸쳐 감수분열 기구를 유지하기에 충분한 이점을 제공할 가능성이 적습니다. 따라서 자연에서 자가수정은 감수분열이 없는 무성 생식 형태로 대체될 것으로 예상할 수 있으며, 이는 비용이 덜 들 것이다. 그러나 오랫동안 자가수정해 온 식물에서 감수분열과 자가수정이 생식 형태로 지속되는 것은 각 세대에서 감수분열에 의해 제공되는 생식 세포 형성 중 DNA 손상의 효율적인 재조합 복구의 직접적인 이점과 관련이 있을 수 있다.
종자식물은 화분이 암술의 주두에 닿는 것을 수정과 동일시하는 경우가 있지만, 이는 수분이라고 한다. 화분에서 화분관이 자라 배주에 도달하고, 배주 안에 있는 난세포와 화분관 내의 정핵(정세포)이 융합하는 것이 수정이다. 화분관이 난세포에 정확하게 도달하는 것은, 배낭에 있는 조세포가 유도하기 때문이다. 트리니아라는 식물을 사용하여 이 사실을 발견한 나고야 대학 교수 히가시야마 테츠야 등의 논문은 2001년 사이언스에 게재되었고, 사진이 표지를 장식했다. 이후에도 히가시야마 등은 연구를 계속하여, 조세포가 내놓는 시스테인이 풍부한 2종류의 저분자량 단백질에 의해 화분관이 유인되고 있다는 것을 알아내고, 이 단백질을 “루어 1”, “루어 2”라고 명명했다.(2009년, 이 연구 결과의 사진은 Nature의 표지에 채택되었다.)[49] 또한, 속씨식물은 이중수정이라고 불리는 독특한 수정 형식을 한다.
4. 2. 동물의 수정
포유류는 교미를 통해 체내 수정을 한다.[22] 수컷이 사정을 하면, 많은 정자가 질 수축을 통해 질 상부로 이동하고, 자궁경부를 지나 자궁 전체를 가로질러 난자를 만난다. 수정이 일어나는 경우, 암컷은 일반적으로 교미 전 몇 시간에서 며칠 후까지 지속되는 기간 동안 배란을 한다.정자는 질에 침착되면 수정이 불가능하며, 느린 직선 운동 패턴을 보인다. 이러한 운동성과 근육 수축은 자궁과 난관으로의 정자 수송을 가능하게 한다.[23] 암컷 생식기관 내에는 pH 기울기가 있어 질 개구부 근처의 pH는 난관(약 8)보다 낮다(약 5).[24] CatSper라고 하는 정자 특이적 pH 감응성 칼슘 수송 단백질은 생식 기관으로 더 이동함에 따라 정자 세포의 칼슘 투과성을 증가시킨다. 세포 내 칼슘 유입은 정자 능동화와 과활성화에 기여하여 난모세포에 접근함에 따라 더욱 격렬하고 빠른 비선형 운동 패턴을 유발한다. 능동화된 정자와 난모세포는 난관의 팽대부에서 만나 상호 작용한다. 유주성, 온도주성, 화학주성은 정자 이동의 마지막 단계에서 난자를 향해 정자를 유도하는 것으로 알려진 메커니즘이다.[25] 정자는 난관과 팽대부 사이의 약 2°C 온도 기울기에 반응하며,[26] 토끼와 사람의 난모세포를 둘러싸고 있는 난구세포에서 나오는 신호로서 프로게스테론의 화학주성 기울기가 확인되었다.[27] 능동화되고 과활성화된 정자는 이러한 기울기에 반응하여 행동을 변화시키고 난구세포-난모세포 복합체를 향해 이동한다. 포르밀 메트-류-페(fMLF)와 같은 다른 화학주성 신호도 정자를 유도할 수 있다.[28]
난자를 둘러싸고 있는 두꺼운 세포외 기질층인 투명대는 정자를 결합한다. 투명대에 있는 당단백질인 ZP3는 사람에서 난자/정자 부착에 중요한 역할을 한다. 수용체인 갈락토실전이효소(GalT)는 ZP3의 N-아세틸글루코사민 잔기에 결합하며 정자와 결합하고 과립반응을 활성화하는 데 중요하다.
포유류에서 정자가 GalT에 결합하면 과립반응이 시작된다. 이 과정은 난모세포 주위의 막에 있는 히알루론산 기질을 분해하는 히알루로니다제를 방출한다. 또한 난모세포 근처에서 헤파린 유사 글리코사미노글리칸(GAGs)이 방출되어 과립반응을 촉진한다.[29] 그 다음 난모세포 세포막과 정자 사이의 융합이 일어나 정자 세포핵, 전형적인 중심체, 그리고 편모에 부착되어 있지만 미토콘드리아는 아닌 비정형적인 중심체가 난모세포로 들어간다.[30] 단백질 CD9는 이러한 융합을 매개할 가능성이 높다. 난자는 단일 정자 세포와 융합하면 자신을 "활성화"시키고 그에 따라 다른 정자와의 융합을 방지하도록 세포막을 변경한다.
이 과정은 궁극적으로 이배체 세포인 접합자를 형성하게 된다. 접합자는 분열하여 배반포를 형성하고, 자궁에 들어가면 자궁내막에 착상하여 임신을 시작한다. 자궁 벽이 아닌 곳에서의 배아 착상은 어머니를 죽일 수 있는 자궁외임신을 초래한다.
토끼와 같은 동물에서는 교미가 뇌하수체 호르몬인 성선자극호르몬의 방출을 자극하여 배란을 유도하며, 이는 임신 가능성을 크게 높인다.
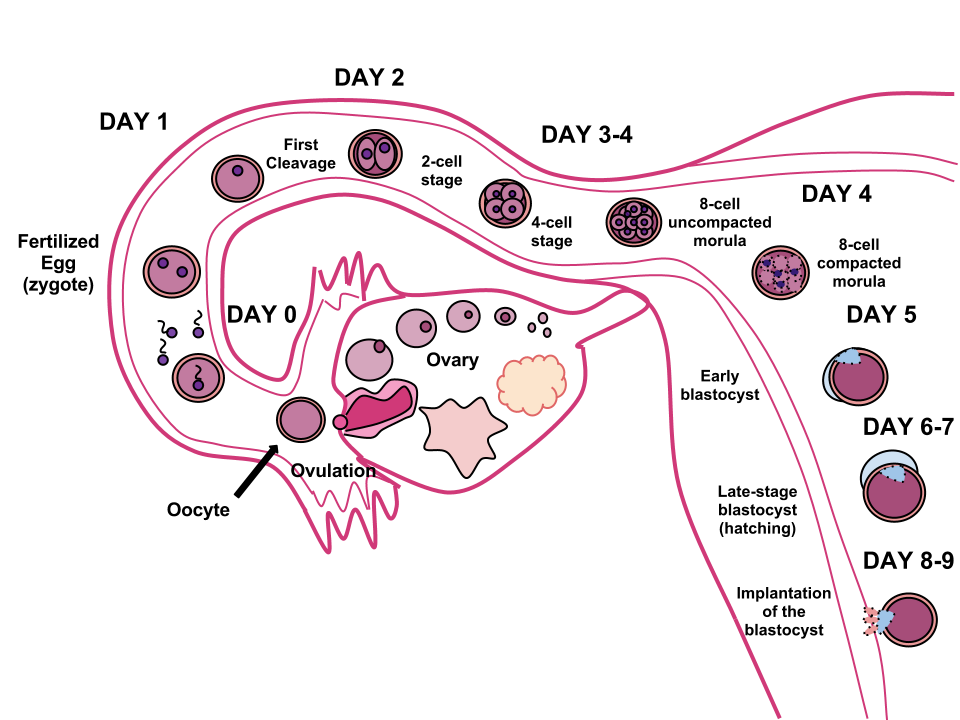
인간의 수정은 인간의 난자와 정자가 결합하는 과정으로, 일반적으로 난관팽대부에서 일어나며, 단일 세포인 접합자를 생성한다. 이는 유전적으로 독특한 생명체의 발생에서 최초 단계이며, 배아 발생을 시작한다.[31]
일반적으로 '수태'라는 용어는 "수정 또는 착상 또는 둘 모두를 포함하는 임신 과정"을 의미한다.[32] 이 용어의 사용은 임신 시작에 대한 의미론적 논쟁의 주제가 되는데, 일반적으로 낙태 논쟁의 맥락에서 발생한다.
정자는 난자의 표면으로 유인된다. 연속된 액체 속에 난자와 정자가 놓인 경우, 정자는 매우 빠르게 난자의 표면에 모인다. 정자가 난자의 표면으로 침입하면, 그 직후에 침입 지점을 중심으로 난자의 표면에서 투명한 막이 솟아오른다. 이 막을 '''수정막'''이라고 한다. 수정막이 생기면 다른 정자는 난자의 표면에서 멀어진다.
난자에 침입하는 정자의 수는 일반적으로 하나뿐이다. 침입한 정자의 핵은 난핵에 접근하고, 결국 두 핵이 융합하여 수정이 완료된다. 여러 개가 침입하더라도 난핵과 융합할 수 있는 것은 하나뿐이므로, 나머지 정자는 갈 곳이 없다. 난자에 흡수되는 경우도 있지만, 성게에서는 여러 개의 정자가 침입하면 난할이 비정상적으로 되는 것이 알려져 있다. 보통은 여러 개의 정자가 침입하는 일은 없다. 이것은 수정막의 작용이라고 생각되기도 했지만, 반드시 그렇지 않다는 설도 있다.
유전학적으로 보면, 수정은 배우자로부터 유래하는 현상이며, 정핵과 난핵의 융합을 수정의 순간으로 보는 것이 옳다. 발생학적으로 보면, 새로운 개체의 핵이 형성되는 것이 두 핵의 융합이므로, 그것을 수정의 순간으로 볼 수도 있지만, 정자가 난자에 접촉한 시점에서 난자의 변화가 시작되므로, 그것을 수정의 순간으로 볼 수도 있다. 실제로 난자의 표면에 화학적인 자극을 준 것만으로 발생이 시작되는 예도 있으며, 그런 면에서 보면 수정에서 정자의 역할은 난자의 발생의 방아쇠를 당기는 것이라는 견해도 성립한다.
동물에서 수정에 이르는 과정은 각 개체의 관계에 따라 체외수정과 체내수정으로 구분된다. 전자는 각 개체가 난자와 정자를 체외로 방출하여 그곳에서 수정이 이루어지는 것이고, 후자는 난자가 체내에 남아 그곳에 정자가 도달하여 수정이 이루어지는 것이다.
인간은 난자 1개체에 정자 1개체가 침입함으로써 수정이 성립하지만, 조류는 1개체의 난자에 여러 개의 정자가 침입해야 수정이 성립한다. 단, 조류의 경우 이것은 배의 성장에 정자에 포함된 유전 물질과는 별개의 물질이 대량으로 필요하기 때문이며, 유전 물질 자체는 정자 1개체분이면 충분하다.[48]
사정 한 번에 포함된 정액 속 정자 수는 일반적으로 1억~4억 정도이지만, 질 내 사정 시 자궁경부에 도달하기 전에 약 99%가 사멸하고, 자궁까지 도달하는 것은 약 수천~수십만 개, 배란기에 난자 바로 앞까지 도달하는 것은 약 수십~수백 개이다. 또한, 질에서 수정 장소인 난관까지는 약 수십 분~수 시간이 걸리는 것으로 생각되지만, 수 분 만에 도달했다는 사례도 확인되고 있다.
4. 3. 균류의 수정
많은 균류(키트리드균 제외)에서 수정은 두 단계 과정으로 진행된다. 먼저 두 배우자 세포의 세포질이 융합하는데(세포질융합), 이를 통해 다핵성인 이핵체 또는 다핵체 세포가 생성된다. 이 세포는 이핵체 또는 다핵체 균사를 생성하기 위해 분열될 수 있다. 수정의 두 번째 단계는 핵융합으로, 핵이 융합하여 이배체 접합자를 형성하는 과정이다.키트리드균류에서는 동물과 식물처럼 배우자의 융합을 통해 단일 단계에서 수정이 일어난다.
4. 4. 원생생물의 수정
원생생물의 수정 과정에는 세 가지 유형이 있다.[37]- 배우자합체(gametogamy)
- 자가수정(autogamy)[38][39]
- 배우체합체(gamontogamy)
일부 육상 식물과 마찬가지로 조류는 세대교번을 한다. 일부 조류는 이형세대(isomorphic)로, 포자체(2n)와 배우체(n)가 형태적으로 동일하다. 조류의 생식이 난형생식(oogamous)으로 설명될 때, 암수 배우자는 형태적으로 다르며, 암배우자는 큰 비운동성 난자이고, 수배우자는 편모가 하나인 운동성 배우자이다. 접합(syngamy) 과정을 통해 이들은 새로운 접합자를 형성하여 다시 포자체 세대를 재생한다.
5. 인간의 수정
인간의 수정은 난자와 정자가 난관의 팽대부에서 결합하여 단일 세포인 접합자를 생성하는 과정이다.[31] 이는 유전적으로 독특한 생명체 발생의 최초 단계이며, 배아 발생을 시작한다.[31] 과학자들은 19세기에 인간 수정의 역학을 발견했다.
포유류는 교미(copulation)를 통해 체내 수정을 한다.[22] 수컷이 사정(ejaculation)을 하면, 많은 정자가 질 수축을 통해 질 상부로 이동하고, 자궁경부를 지나 자궁 전체를 가로질러 난자를 만난다. 수정이 일어나는 경우, 암컷은 일반적으로 교미 전 몇 시간에서 며칠 후까지 지속되는 기간 동안 배란(ovulation)을 한다.
능동화(capacitation)된 정자와 난모세포는 난관의 팽대부에서 만나 상호 작용한다.[25] 정자는 난관과 팽대부 사이의 약 2°C 기울기에 반응하며,[26] 난구세포에서 나오는 신호로서 프로게스테론의 화학주성 기울기가 확인되었다.[27]
난자를 둘러싸고 있는 두꺼운 세포외 기질층인 투명대는 정자를 결합한다. 투명대에 있는 당단백질인 ZP3는 사람에서 난자/정자 부착에 중요한 역할을 한다. 수용체인 갈락토실전이효소(GalT)는 ZP3의 N-아세틸글루코사민 잔기에 결합하며 정자와 결합하고 과립반응을 활성화하는 데 중요하다.
포유류에서 정자가 GalT에 결합하면 과립반응(acrosome reaction)이 시작된다. 이 과정은 난모세포 주위의 막에 있는 히알루론산 기질을 분해하는 히알루로니다제를 방출한다. 그 다음 난모세포 세포막과 정자 사이의 융합이 일어나 정자 세포핵, 전형적인 중심체, 그리고 편모에 부착되어 있지만 미토콘드리아는 아닌 비정형적인 중심체가 난모세포로 들어간다.[30] 난자는 단일 정자 세포와 융합하면 자신을 "활성화(egg activation)"시키고 그에 따라 다른 정자와의 융합을 방지하도록 세포막을 변경한다.
이 과정은 궁극적으로 이배체 세포인 접합자를 형성하게 된다. 접합자는 분열하여 배반포를 형성하고, 자궁에 들어가면 자궁내막에 착상하여 임신을 시작한다.
정자는 난자의 표면으로 유인된다. 연속된 액체 속에 난자와 정자가 놓인 경우, 정자는 매우 빠르게 난자의 표면에 모인다. 정자가 난자의 표면으로 침입하면, 그 직후에 침입 지점을 중심으로 난자의 표면에서 투명한 막이 솟아오른다. 이 막을 '''수정막'''이라고 한다. 수정막이 생기면 다른 정자는 난자의 표면에서 멀어진다.
난자에 침입하는 정자의 수는 일반적으로 하나뿐이다. 침입한 정자의 핵은 난핵에 접근하고, 결국 두 핵이 융합하여 수정이 완료된다. 여러 개가 침입하더라도 난핵과 융합할 수 있는 것은 하나뿐이므로, 나머지 정자는 갈 곳이 없다.
유전학적으로 보면, 수정은 배우자로부터 유래하는 현상이며, 정핵과 난핵의 융합을 수정의 순간으로 보는 것이 옳다. 발생학적으로 보면, 새로운 개체의 핵이 형성되는 것이 두 핵의 융합이므로, 그것을 수정의 순간으로 볼 수도 있지만, 정자가 난자에 접촉한 시점에서 난자의 변화가 시작되므로, 그것을 수정의 순간으로 볼 수도 있다.
사정(射精) 한 번에 포함된 정액(精液) 속 정자 수는 일반적으로 1억~4억 정도이지만, 질(腟) 내 사정 시 자궁경부에 도달하기 전에 약 99%가 사멸하고, 자궁까지 도달하는 것은 약 수천~수십만 개, 배란기(排卵期)에 난자(卵子) 바로 앞까지 도달하는 것은 약 수십~수백 개이다. 또한, 질에서 수정 장소인 난관까지는 약 수십 분~수 시간이 걸리는 것으로 생각되지만, 수 분 만에 도달했다는 사례도 확인되고 있다.
배란기에는 난자가 난포에서 나와(배란) 나팔관을 따라 자궁으로 이동하며, 수정되면 착상하고, 수정되지 않으면 수 시간~24시간 이내에 퇴화한다. 한편, 정자의 수정 능력 유지 기간은 수 시간에서 3일, 최대 7일(자궁경관, 자궁 내, 나팔관 내)로 알려져 있으며, 이를 고려하면 정자의 수정 가능 기간인 배란 전 7일과 난자의 수정 가능 기간인 배란 후 1일을 합쳐, 배란 전후 8일이 한 번의 월경 주기에서 수정이 가능한 기간이 된다.
배란은 생리 시작 14일 전후에 일어나지만, 배란 시기는 여러 원인으로 쉽게 변동하기 때문에 임신 가능 시기를 예측하는 것은 어렵다.
인간의 쌍둥이는 하나의 난자에 정자가 두 개 들어갔기 때문이라는 속설이 있지만, 이는 잘못된 것이다.
6. 수정과 관련된 기타 생식 현상
수정은 감수분열의 기원과 관련되며, 둘 다 유성생식의 일부로 진핵생물에서 유래했다.[6] 감수분열은 각 부모가 기여하는 유전자의 무작위 분리를 초래하며, 수정 시 부모의 염색체가 결합한다. 유전학적으로 수정은 배우자로부터 유래하는 현상이며, 정핵과 난핵의 융합을 수정의 순간으로 본다.
일반적으로 유성생식을 하는 생물도 단성생식을 통해 번식할 수 있는데, 이는 수정되지 않은 암컷 배우자가 생존 가능한 자손을 생성하는 과정이다. 2004년 코노 토모히로가 이끄는 일본 연구팀은 두 마리 쥐의 난자를 융합하여 정상적인 쥐를 탄생시키는 데 성공했다.[43]
타가수정은 한 개체의 난세포가 다른 개체의 정자와 수정하는 것을 말하며,[46] 근교약세를 피하는 것이 주요 이점이다. 찰스 다윈은 서로 다른 환경에서 자란 두 개체의 결합으로 나온 자손이 자가수정을 통해 나온 자손보다 여러 면에서 큰 이점을 가진다고 밝혔다.[46]
자가수정은 식물과 편형동물 같은 자웅동체 생물에서 일어나는 현상으로, 한 개체의 두 배우자가 융합하는 것을 말한다. 수분 매개체가 드문 환경에서는 자가수정이 생식에 유리하다.[17]
6. 1. 유전적 재조합
수정의 진화는 감수분열의 기원과 관련이 있으며, 둘 다 유성생식의 일부로 진핵생물에서 유래했다.[6] 감수분열은 각 부모가 기여하는 유전자의 무작위 분리를 초래한다. 각 부모는 유전자의 일부를 제외하고는 일반적으로 동일한 유전자를 가지므로, 각 생식세포는 유전적으로 독특하다. 수정 시 부모의 염색체가 결합한다. 인간의 경우, 비성염색체에 대해 (2²²)² = 17.6x1012개의 염색체적으로 다른 접합자가 가능하며, 염색체 교차가 발생하지 않는다고 가정하더라도 가능하다. 교차가 한 번 발생하면 평균적으로 (4²²)² = 309x1024개의 유전적으로 다른 접합자가 각 부부에 대해 가능하며, 이는 교차 사건이 각 염색체의 대부분 지점에서 발생할 수 있다는 점을 고려하지 않은 것이다. X염색체와 Y염색체는 교차 사건이 발생하지 않으므로 계산에서 제외된다.[6] 미토콘드리아 DNA는 모계 부모로부터만 상속된다.유전학적으로 보면, 수정은 배우자로부터 유래하는 현상이며, 정핵과 난핵의 융합을 수정의 순간으로 본다.
6. 2. 단성생식
일반적으로 유성생식을 하는 생물도 단성생식을 통해 번식할 수 있다. 단성생식은 수정되지 않은 암컷 배우자가 생존 가능한 자손을 생성하는 과정이다. 이 자손은 어미의 복제일 수도 있고, 어떤 경우에는 유전적으로 어미와 다르지만 어미의 DNA 일부만을 물려받을 수도 있다. 단성생식은 많은 식물과 동물에서 일어나며, 다른 생물에서는 난자에 화학적 또는 전기적 자극을 가하여 유도될 수 있다. 2004년, 코노 토모히로가 이끄는 일본 연구팀은 정상적으로 단성생식을 막는 특정 단백질을 차단하여 두 마리 쥐의 난자를 융합하는 데 457번의 시도 끝에 성공했다. 그 결과 생성된 배아는 정상적으로 쥐로 발달했다.[43]다소 특이한 형태의 생식에는 다음과 같은 것들이 있다.[44][45]
- 단성생식: 정자가 수정이나 배우자합체 없이 난자가 발생하도록 자극한다. 정자는 난자에 들어갈 수 있다.
6. 3. 타가수정과 자가수정
타가수정(또는 타가수분)은 한 개체의 난세포가 다른 개체의 정자와 수정하는 것을 말한다.[46] 식물 종의 약 48.7%는 암수딴그루 또는 자가불화합성으로, 타가수분을 통해서만 번식이 가능하다.[15] 또한, 약 42%의 꽃피는 식물은 자연에서 타가수정과 자가수정을 혼합하는 교배 시스템을 가진다.[16]타가수정의 주요 이점은 근교약세를 피하는 것이다. 찰스 다윈은 그의 저서 ''식물계에서의 타가수정과 자가수정의 효과''에서 서로 다른 환경에서 자란 두 개체의 결합으로 나온 자손은 자가수정을 통해 나온 자손보다 키, 무게, 활력, 생식력에서 큰 이점을 가진다고 밝혔다.[46]
자가수정(또는 자가수분)은 식물과 편형동물과 같은 자웅동체 생물에서 일어나는 현상으로, 한 개체의 두 배우자가 융합하는 것을 말한다. 꽃피는 식물의 약 10~15%는 주로 자가수정을 한다.[17] 수분 매개체가 드문 환경에서는 자가수정이 생식에 유리하여 개척 능력을 향상시킬 수 있다.[17] ''애기장대속''의 ''붉은애기장대''는 5만 년에서 10만 년 전에 자가수정을 하는 종으로 진화했다.[18] ''애기장대''는 야생에서 타가수정 비율이 0.3% 미만인 주로 자가수정하는 식물이다.[19] 한 연구에 따르면 자가수정은 ''애기장대''에서 약 100만 년 전 또는 그 이상에 진화했다.[20]
참조
[1]
웹사이트
impregnation
https://www.oxfordle[...]
[2]
논문
The cell biology of fertilization: Gamete attachment and fusion
https://rupress.org/[...]
2021-01-14
[3]
웹사이트
Fertilization
https://www.merriam-[...]
2018-07-10
[4]
서적
Integrated History and Philosophy of Science
2017
[5]
서적
Sperm Biology
2009
[6]
논문
The evolution of meiosis from mitosis
2009-01
[7]
서적
Fertilization in Higher Plants.
Springer
1999
[8]
논문
Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis
2014
[9]
논문
A floral transmitting tissue-specific glycoprotein attracts pollen tubes and stimulates their growth
1995
[10]
서적
Biology: facts & practice for A level
https://archive.org/[...]
Oxford University Press
2001
[11]
서적
Handbook of plant science
John Wiley
2007
[12]
서적
Biology today
https://archive.org/[...]
CRM
1975
[13]
서적
Double fertilization: embryo and endosperm development in flowering plant
https://archive.org/[...]
Springer-Verlag
2006
[14]
논문
Modularity of the Angiosperm Female Gametophyte and Its Bearing on the Early Evolution of Endosperm in Flowering Plants
2003
[15]
논문
The distribution of plant mating systems: study bias against obligately outcrossing species
2006
[16]
논문
The evolutionary enigma of mixed mating systems in plants: Occurrence, theoretical explanations, and empirical evidence
2005
[17]
논문
Evolutionary consequences of self-fertilization in plants
2013
[18]
논문
Genomic Identification of Founding Haplotypes Reveals the History of the Selfing Species Capsella rubella
2013
[19]
논문
Population genetic structure and outcrossing rate of Arabidopsis thaliana (L.) Heynh
1989
[20]
논문
The evolution of selfing in Arabidopsis thaliana
2007
[21]
서적
Les stratégies de reproduction des animaux
Dunod
2001
[22]
서적
Advances in the Study of Behavior
https://books.google[...]
Academic Press
2020-04-19
[23]
논문
Sperm transport in the female reproductive tract
2006
[24]
논문
In vivo oxygen, temperature and pH dynamics in the female reproductive tract and their importance in human conception: a systematic review
2018-01-01
[25]
논문
Oviduct: Roles in fertilization and early embryo development
2016
[26]
논문
Sperm thermotaxis
https://zenodo.org/r[...]
2006
[27]
논문
Molecular Mechanism for Human Sperm Chemotaxis Mediated by Progesterone
2009
[28]
논문
Evidence for the presence of specific receptors for N-formyl chemotactic peptides on human spermatozoa
1986
[29]
논문
Glycosaminoglycans in Ewe Reproductive Tracts and Their Influence on Acrosome Reactions in Bovine Spermatozoa in Vitro
1986-09-01
[30]
논문
A novel atypical sperm centriole is functional during human fertilization
2018
[31]
웹사이트
Zygote {{!}} Definition, Development, Example, & Facts {{!}} Britannica
https://www.britanni[...]
2023-01-14
[32]
웹사이트
Conception
https://www.merriam-[...]
2019-01-27
[33]
서적
The Developing Human: Clinically Oriented Embryology
https://archive.org/[...]
W. B. Saunders Company
2003
[34]
서적
Field Guide to the Dragonflies of Britain and Europe
British Wildlife Publishing
2006
[35]
서적
The Birder's Bug Book
Harvard University Press
1998
[36]
웹사이트
Beekeeping in Africa: Colony life and social organization
http://www.fao.org/d[...]
FAO
[37]
서적
Fertilization in Protozoa and Metazoan Animals: Cellular and Molecular Aspects
Springer Science & Business Media
2000
[38]
문서
Autogamy
[39]
웹사이트
Autogamy | biology
https://www.britanni[...]
[40]
논문
The Role of Sperm Centrioles in Human Reproduction – The Known and the Unknown
2019-10-01
[41]
논문
A novel atypical sperm centriole is functional during human fertilization
2018-12
[42]
논문
Parental genome unification is highly error-prone in mammalian embryos
2021-05
[43]
논문
Birth of parthenogenetic mice that can develop to adulthood
2004
[43]
잡지
The End of Males? Mouse Made to Reproduce Without Sperm
http://news.national[...]
2004-04-21
[44]
논문
Meiosis and Its Deviations in Polyploid Animals
2013
[45]
논문
Simultaneous Mendelian and clonal genome transmission in a sexually reproducing, all-triploid vertebrate
2011
[46]
서적
The effects of cross and self fertilisation in the vegetable kingdom
https://www.gutenber[...]
J. Murray
1876
[47]
논문
Why have sex? The population genetics of sex and recombination
2006
[48]
방송
사이언스ZERO
2015-02-08
[49]
서적
三訂版 フォトサイエンス 生物図録
数研出版
2016
[50]
백과사전
수정의 구조 - 체외 수정과 체내 수정
글로벌 세계 대백과사전
2017-09-29
[51]
백과사전
수정의 구조 - 수정의 과정
글로벌 세계 대백과사전
2017-09-29
[52]
백과사전
수정의 구조 - 정자의 접근과 침입
글로벌 세계 대백과사전
2017-09-29
[53]
백과사전
수정의 구조 - 난자의 수정막 형성과 핵의 합체
글로벌 세계 대백과사전
2017-09-29
[54]
백과사전
수정의 구조 - 인공 단위 발생
글로벌 세계 대백과사전
2017-09-29
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com