관박쥐류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
관박쥐류는 굽은코박쥐과에 속하는 박쥐들을 통칭하며, 굽은코박쥐속에 속하는 다양한 종을 포함한다. 1799년 처음 기술되었으며, 코에 잎 모양의 돌출부인 코잎을 가지고 있어 "관박쥐"라는 이름이 붙었다. 관박쥐류는 작거나 중간 크기의 소형박쥐로, 반향정위를 통해 먹이를 찾고, 곤충, 거미 등을 섭취한다. 주로 구열대구에 분포하며, 동굴, 건물 등 다양한 서식지에서 발견된다. 일부 종은 식용 또는 약용으로 사용되기도 하며, IUCN 적색 목록에 따라 보존 상태가 평가되고 있다.
더 읽어볼만한 페이지
- 관박쥐과 - 달링관박쥐
달링관박쥐는 1905년 안데르센에 의해 처음 기재되었고 몸길이 4.5~5.5cm의 관박쥐과 박쥐로, 아프리카 남부 지역에 분포하며, 어두운 갈색 등과 연한 갈색 배, 넓은 날개폭을 가진 여러 아종으로 분류되는 종이다. - 관박쥐과 - 마엔델레오관박쥐
- 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 관박쥐류 - [생물]에 관한 문서 | |
|---|---|
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 포유강 |
| 하강 | 진수하강 |
| 상목 | 로라시아상목 |
| 목 | 박쥐목 |
| 아목 | 음박쥐아목 |
| 과 | 관박쥐과 (Rhinolophidae) |
| 과 명명자 | Gray, 1821 |
| 속 | 관박쥐속 (Rhinolophus) |
| 속 명명자 | Lacépède, 1799 |
| 하위 분류 | 종 (본문 참조) |
| 생물학적 분류 | |
| 학명 | Rhinolophidae |
| 명명자 | Gray, 1825 |
| 상위 분류 | Gray, 1825 |
| 속 | Rhinolophus |
| 속 명명자 | Lacépède, 1799 |
| 하위 분류 | 종 |
| 모식종 | Vespertilio ferrum-equinum |
| 모식종 명명자 | Schreber, 1774 |
| 특징 | |
| 무게 | 4–28 g |
| 몸통 길이 | 30–75 mm |
| 앞팔 길이 | 35–110 mm |
| 이미지 | |
 | |
| 기타 | |
| 영어 이름 | horseshoe bat |
2. 분류 및 진화
2002년 박쥐목 계통 분류에 따르면, 관박쥐류는 관박쥐과(Rhinolophidae)에 속하며 잎코박쥐과와 함께 틈새얼굴박쥐과, 위흡혈박쥐과, 생쥐꼬리박쥐류, 키티돼지코박쥐과, 대꼬리박쥐과와 가까운 관계를 가진다.[63]
가와다 외(2018)의 연구에 따르면 굽은코박쥐과에는 굽은코박쥐속(관박쥐속)이 있으며, 여기에는 뾰족코잎코박쥐, 콩고굽은코박쥐, 나카굽은코박쥐 등 다양한 종이 포함된다.
2. 1. 분류 역사
1799년 프랑스 박물학자 베르나르 제르맹 드 라세페드가 ''Rhinolophus''를 처음 속으로 기술했다. 처음에는 현존하는 모든 관박쥐와 현재 Hipposideros(둥근잎박쥐)에 속하는 종까지 ''Rhinolophus''에 포함되었다.[1] 초기에는 ''Rhinolophus''가 애기박쥐과(Vespertilionidae)에 속했다. 1825년, 영국의 동물학자 존 에드워드 그레이는 애기박쥐과를 Rhinolophina라는 아과로 세분화했다.[2] 1836년, 영국의 동물학자 토마스 벨은 관박쥐를 별도의 과(Rhinolophidae)로 처음 인정했다.[3] 벨이 Rhinolophidae의 권위자로 인정받기도 하지만,[4] 일반적으로 그레이(1825)를 권위자로 본다.[3][5] 관박쥐는 상과(Superfamily) Rhinolophoidea에 속하며, 꼬마귀과(Craseonycteridae), 잎코박쥐과(Hipposideridae), 큰귀잎코박쥐과(Megadermatidae), 코주머니박쥐과(Rhinonycteridae), 관코박쥐과(Rhinopomatidae)와 함께 분류된다.[6][10]''Rhinolophus''를 다른 속으로 나누려는 시도가 있었다. 1816년, 영국의 동물학자 윌리엄 엘포드 리치는 ''Phyllorhina''라는 속명을 제안했고, 그레이는 1847년에 ''Aquias'', 1866년에 ''Phyllotis''를 제안했으며, 독일의 박물학자 빌헬름 페터스는 1867년에 ''Coelophyllus''를 제안했다. 1876년, 아일랜드의 동물학자 조지 에드워드 돕슨은 모든 아시아 관박쥐를 다시 ''Rhinolophus''로 되돌렸고, 잎코박쥐류를 위한 아과 Phyllorhininae와 관박쥐류를 위한 Rhinolophinae를 추가로 제안했다. 미국의 동물학자 게릿 스미스 밀러 주니어는 1907년에 잎코박쥐류를 관박쥐류에서 더 세분화하여 Hipposideridae를 별도의 과로 인정했다.[1] 일부 저자는 2000년대 초반까지 ''Hipposideros''와 관련된 속을 Rhinolophidae의 일부로 간주했지만,[44] 현재는 별도의 과로 인식되는 경우가 많다.[17][7] Rhinolophidae와 Hipposideridae로 분리된 후, ''Rhinolophus''에 대한 추가적인 분할이 제안되었으며, 1934년에는 ''Rhinolphyllotis'', 1951년에는 ''Rhinomegalophus''가 제안되었지만, 두 속 모두 다시 ''Rhinolophus''로 되돌려졌다.[1]
덴마크의 포유류학자 크누드 안데르센은 1905년에 ''Rhinolophus''에 대한 종 그룹을 처음 제안했다. 종 그룹은 진화적 관계를 반영하기 위해 종을 묶는 방법이다. 그는 ''R. simplex''(현재 ''R. megaphyllus''), ''R. lepidus'', ''R. midas''(현재 ''R. hipposideros''), ''R. philippinensis'', ''R. macrotis'', ''R. arcuatus''의 6개 종 그룹을 인식했다. 새로운 그룹이 추가되고, 새로운 종이 기술되며, 종 간의 관계가 수정됨에 따라 종은 그룹 간에 자주 재배열되었다.[1] 2003년 Csorba와 동료들은 15개의 종 그룹을 제시했다.[1][8] 다양한 아속도 제안되었으며, 2003년 Csorba ''et al''.에 의해 ''Aquias'', ''Phyllorhina'', ''Rhinolophus'', ''Indorhinolophus'', ''Coelophyllus'', ''Rhinophyllotis''의 6개가 나열되었다.[1] 비공식적으로 관박쥐류는 주로 아프리카 계통과 동양 계통의 두 개의 주요 분지군으로 나눌 수 있다.[44]
2. 2. 진화 역사
관박쥐속(''Rhinolophus'')의 가장 최근 공통 조상은 약 3,400만~4,000만 년 전인 에오세에 히포시데리드 계통에서 분화되었다.[11][44] 화석 관박쥐는 유럽(초기~중기 중신세, 초기 올리고세), 호주(중신세), 아프리카(중신세 및 후기 플라이오세)에서 발견된다.[9] 관박쥐의 생물지리학은 제대로 알려져 있지 않다. 다양한 연구에서 이 과가 유럽, 아시아 또는 아프리카에서 기원했다고 주장한다. 2010년 연구는 올리고세 동안 아프리카와 동양 계통의 급속한 진화적 방산과 함께 이 과의 아시아 또는 동양 기원을 지지했다.[44] 2019년 연구에 따르면 유라시아 종인 ''R. xinanzhongguoensis''와 ''R. nippon''이 다른 유라시아 종보다 아프리카 종과 더 밀접하게 관련되어 있어, 관박쥐과가 아시아 및 아프리카 열대 지역과 복잡한 생물지리학적 관계를 가질 수 있음을 시사했다.[11]2016년 미토콘드리아 DNA 및 핵 DNA를 사용한 연구에 따르면 관박쥐는 Yinpterochiroptera 내에 있으며 히포시데리과의 자매 분류군으로 분류되었다.[10]
관박쥐과는 현존하는 유일한 속인 ''Rhinolophus''로 대표된다. 관박쥐과와 관박쥐속 모두 단계통군(공통 조상의 모든 후손을 포함)으로 확인되었다. 2019년 현재, ''Rhinolophus''에는 106종의 종이 기술되어 있으며, 이는 ''Myotis'' 다음으로 두 번째로 다양한 박쥐 속이다. ''Rhinolophus''는 아프리카 열대 지역에서 표본이 부족할 수 있으며, 한 유전자 연구에서는 이 지역에 최대 12종의 숨겨진 종이 있을 수 있다고 추정했다. 또한, 완전한 종으로 인식된 일부 분류군에서 유전자 분기가 거의 발견되지 않았다. ''Rhinolophus kahuzi''는 루웬조리 관박쥐(''R. ruwenzorii'')의 동의어일 수 있으며, ''R. gorongosae'' 또는 ''R. rhodesiae''는 부시펠트 관박쥐(''R. simulator'')의 동의어일 수 있다. 또한 스미더스 관박쥐(''R. smithersi''), 코헨 관박쥐(''R. cohenae''), 마운트 마부 관박쥐(''R. mabuensis'')는 모두 힐데브란트 관박쥐(''R. hildebrandtii'')와 유전자 분기가 거의 없다. 전자의 세 종을 완전한 종으로 인식하면 힐데브란트 관박쥐가 측계통군이 된다.[11]
관박쥐과의 두 번째 속은 멸종된 ''Palaeonycteris''이며, 대표 종은 ''Palaeonycteris robustus''이다.[12] ''Palaeonycteris robustus''는 초기 중신세에 살았으며, 화석 유해는 프랑스 생제랑르퓌이에서 발견되었다.[13][14]

3. 형태
관박쥐류는 작거나 중간 크기의 소형박쥐로, 머리와 몸통 길이는 35mm, 팔뚝 길이는 30mm이다. 작은관박쥐는 4g, 큰관박쥐는 16.5g 정도의 무게가 나간다. 털 색깔은 검은색에서 붉은 갈색, 밝은 주황색-빨간색까지 다양하며, 몸 아랫부분은 등 털보다 옅다. 대부분 길고 부드러운 털을 가지고 있지만, 솜털관박쥐와 작은솜털관박쥐는 매우 길고 솜털 같은 털을 가지고 있다는 점에서 특이하다.[15][9]
대부분의 박쥐와 마찬가지로 가슴에 두 개의 유선이 있다. 성체 암컷은 복부에 젖꼭지 모양의 돌기 두 개(음부 젖꼭지 또는 가짜 젖꼭지)를 가지는데, 이는 유선에 연결되어 있지 않다. Hipposideridae, Craseonycteridae, Megadermatidae, Rhinopomatidae를 포함한 몇몇 다른 박쥐 과만 음부 젖꼭지를 가지고 있으며, 이는 새끼가 부착되는 지점 역할을 한다.[16] 일부 관박쥐 종에서는 수컷이 각 겨드랑이에 가짜 젖꼭지를 가지기도 한다.[17]
모든 관박쥐류는 코에 크고 잎 모양의 돌출부인 코잎을 가지고 있다.[17] 코잎은 종을 식별하는 데 중요하며, 여러 부분으로 구성된다. 코잎의 앞부분은 말굽을 닮았으며, 말굽이라고 불린다. 이 때문에 "관박쥐"라는 일반적인 이름이 붙었다.[17] 창은 삼각형 모양으로 뾰족하고 주머니 모양이며 박쥐의 눈 사이를 위로 향한다.[18] 안장은 콧구멍 뒤에서 솟아 머리에서 수직으로 뻗어 나오는 평평하고 능선 모양의 구조이다.[18]
귀는 크고 잎 모양이며, 길이만큼 넓고 귓불이 없다. 귀의 대이주는 눈에 띈다. 눈은 매우 작다.[17] 관박쥐류의 전형적인 치식은 1.1.2.32.1.3.3이지만, 중간 아래쪽 소구치와 앞쪽 위쪽 소구치는 종종 없다.[1] 새끼는 자궁 내에서 젖니를 잃고 몸 안으로 흡수된다.[15][19] 영구적인 송곳니 4개가 맹출된 상태로 태어나 어미에게 매달릴 수 있는데,[19] 이는 대부분의 박쥐과와는 다르다.
말굽박쥐는 눈이 매우 작고 시야가 코잎에 의해 제한되므로 시력이 중요한 감각은 아닐 것이다. 대신, 반향정위를 사용하여 탐색하며, 박쥐 그룹 중 가장 정교한 반향정위를 사용한다.[22] 콧구멍을 통해 소리를 내어 반향정위를 하며, 주파수 변조 대신 정주파수 반향정위(단일 주파수 반향정위)를 사용한다.[24] 높은 듀티 사이클을 가지고 있어 발신할 때 30% 이상 소리를 생성하며, 이는 크기에 따라 먹이를 구별하는 데 도움이 된다.[22] 코잎은 포물 반사기처럼 작용하여 생성된 소리를 목표로 하면서 동시에 일부 소리로부터 귀를 보호한다.[17]
말굽박쥐는 잘 발달된 달팽이관을 가지고 있으며,[17] 도플러 효과에 의해 이동된 메아리를 감지하여 소리를 동시에 생성하고 수신할 수 있다.[1] 반향정위 주파수가 높은 종은 귀 길이가 짧아지는 경향이 있으며,[23] 반향정위 동안 귀는 서로 독립적으로 움직이고, 머리는 위아래 또는 좌우로 움직인다.[17]
3. 1. 외형
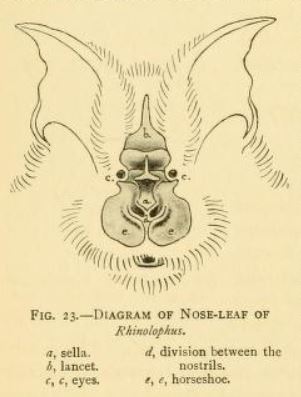
관박쥐류는 작거나 중간 크기의 소형박쥐로 여겨진다.[17] 머리와 몸통 길이는 35mm이며, 팔뚝 길이는 30mm이다. 더 작은 종 중 하나인 작은관박쥐는 4g의 무게가 나가고, 더 큰 종 중 하나인 큰관박쥐는 16.5g의 무게가 나간다. 털 색깔은 종에 따라 매우 다양하며 검은색에서 붉은 갈색, 밝은 주황색-빨간색까지 다양하다.[15][9] 몸 아랫부분은 등 털보다 더 옅다.[15] 대부분의 종은 길고 부드러운 털을 가지고 있지만, 솜털관박쥐와 작은솜털관박쥐는 매우 길고 솜털 같은 털을 가지고 있다는 점에서 특이하다.[9]
대부분의 박쥐와 마찬가지로 관박쥐는 가슴에 두 개의 유선을 가지고 있다. 성체 암컷은 또한 복부에 젖꼭지 모양의 돌기 두 개를 가지고 있는데, 이를 음부 젖꼭지 또는 가짜 젖꼭지라고 부르며, 이는 유선에 연결되어 있지 않다. Hipposideridae, Craseonycteridae, Megadermatidae, Rhinopomatidae를 포함한 몇몇 다른 박쥐 과만 음부 젖꼭지를 가지고 있으며, 이는 새끼가 부착되는 지점 역할을 한다.[16] 일부 관박쥐 종에서는 수컷이 각 겨드랑이에 가짜 젖꼭지를 가지고 있다.[17]
3. 2. 코잎(비엽)
모든 관박쥐류는 코에 크고 잎 모양의 돌출부를 가지고 있으며, 이를 코잎이라고 부른다.[17] 코잎은 종을 식별하는 데 중요하며 여러 부분으로 구성된다.[18] 코잎의 앞부분은 말굽을 닮았으며, 말굽이라고 불리며, 이 때문에 "관박쥐"라는 일반적인 이름을 얻었다.[17] 말굽은 윗입술 위에 있으며 얇고 평평하다. 창은 삼각형 모양으로 뾰족하고 주머니 모양이며 박쥐의 눈 사이를 위로 향한다.[18] 안장은 콧구멍 뒤에서 솟아 머리에서 수직으로 뻗어 나오는 평평하고 능선 모양의 구조이다.[18]
3. 3. 귀와 이빨
모든 관박쥐류는 코에 크고 잎 모양의 돌출부를 가지고 있으며, 이를 코잎이라고 부른다.[17] 코잎은 종을 식별하는 데 중요하며 여러 부분으로 구성된다.[18] 귀는 크고 잎 모양이며, 길이만큼 넓고 귓불이 없다. 귀의 대이주는 눈에 띈다. 눈은 매우 작다.[17] 관박쥐류의 전형적인 치식은 1.1.2.32.1.3.3이지만, 중간 아래쪽 소구치와 앞쪽 위쪽 소구치(입 앞쪽의 소구치)가 종종 없다.[1] 새끼는 자궁 내에서 젖니를 잃으며,[15] 치아는 몸 안으로 흡수된다.[19] 영구적인 송곳니 4개가 맹출된 상태로 태어나 어미에게 매달릴 수 있는데,[19] 이는 대부분의 박쥐과와는 다르다. 대부분의 박쥐과 신생아는 적어도 일부 젖니를 가지고 태어나며, 이는 영구적인 치아로 빠르게 대체된다.[20]
3. 4. 기타

말굽박쥐는 눈이 매우 작고 시야가 큰 코잎에 의해 제한되므로 시력이 매우 중요한 감각은 아닐 것이다. 대신, 그들은 반향정위를 사용하여 탐색하며,[9] 박쥐 그룹 중 가장 정교한 반향정위를 사용한다.[22] 콧구멍을 통해 소리를 내어 반향정위를 한다. 일부 박쥐는 주파수 변조 반향정위를 사용하지만 말굽박쥐는 정주파수 반향정위(단일 주파수 반향정위라고도 함)를 사용한다.[24] 이들은 높은 듀티 사이클을 가지고 있으며, 이는 개체가 발신할 때 30% 이상 소리를 생성한다는 것을 의미한다. 높은 듀티, 정주파수 반향정위의 사용은 크기에 따라 먹이를 구별하는 데 도움이 된다. 이러한 반향정위 특성은 잎이 무성한 어수선한 환경에서 움직이는 먹이를 찾는 박쥐에게 전형적이다.[22]
말굽박쥐는 박쥐에 비해 특히 높은 주파수로 반향정위를 하며, 체구에 비해 히포시데리드보다 높지는 않으며, 대부분의 반향정위 에너지를 두 번째 배음에 집중시킨다. 왕 말굽박쥐(''R. rex'')와 대이 말굽박쥐(''R. philippensis'')는 두 번째 배음이 아닌 첫 번째 배음에 에너지를 집중시키는 예외적인 종이다.[23] 이들의 고도로 주름진 코잎은 소리의 방출을 집중시켜 환경 혼란의 영향을 줄이는 데 도움이 될 수 있다.[24] 일반적으로 코잎은 포물 반사기처럼 작용하여 생성된 소리를 목표로 하면서 동시에 일부 소리로부터 귀를 보호한다.[17]
말굽박쥐는 잘 발달된 달팽이관으로 인해 정교한 청각 감각을 가지고 있으며,[17] 도플러 효과에 의해 이동된 메아리를 감지할 수 있다. 이를 통해 소리를 동시에 생성하고 수신할 수 있다.[1] 말굽박쥐 내에서 귀 길이와 반향정위 주파수 사이에는 음의 관계가 있다. 반향정위 주파수가 높은 종은 귀 길이가 짧아지는 경향이 있다.[23] 반향정위 동안 귀는 가족의 특징인 "깜박임" 동작으로 서로 독립적으로 움직일 수 있으며, 머리는 동시에 위아래 또는 좌우로 움직인다.[17]
4. 생태
관박쥐류는 곤충을 먹지만, 거미와 같은 다른 절지동물도 먹는다.[1] 먹이 활동은 보통 지상 5m 위에서 일어난다.[9] 저녁박쥐와 달리 관박쥐는 요막을 사용하여 먹이를 잡지 않는다. 큰관박쥐는 날개 끝에서 손가락뼈를 구부려 먹이를 잡은 다음 입으로 옮기는 것으로 기록되어 있다.[17][26] 대부분의 관박쥐는 야행성이며 밤에 사냥하지만, 블라이스 관박쥐는 티오만 섬에서 낮 동안 먹이 활동을 하는 것으로 알려져 있다. 이는 섬에 주행성 조류(낮에 활동하는 새) 포식자가 부족하기 때문인 것으로 추정된다.[27]
관박쥐는 작고 둥근 날개 끝, 낮은 날개 하중(체질량에 비해 날개가 크다는 의미), 높은 캠버를 가지고 있어 민첩하게 느린 속도에서도 빠르고 좁은 회전을 할 수 있다.[25] 모든 박쥐에 비해 관박쥐의 날개 길이는 체형에 비해 일반적이며, 가로 세로 비는 평균이거나 평균보다 낮다. 루펠 관박쥐, 힐데브란트 관박쥐, 랜더 관박쥐, 스위니 관박쥐와 같은 일부 종은 특히 넓은 총 날개 면적을 가지고 있지만, 대부분의 관박쥐 종은 평균적인 날개 면적을 가지고 있다.[25]
4. 1. 반향정위 및 청각
관박쥐는 곤충을 먹지만, 거미와 같은 다른 절지동물도 섭취하며, 두 가지 주요한 먹이 활동 전략을 사용한다. 첫 번째 전략은 나무와 덤불 사이를 천천히 낮은 고도로 비행하는 것이다. 이 전략을 사용하는 일부 종은 먹이 위를 맴돌며 기찰하여 기질에서 먹이를 잡아낼 수 있다. 다른 전략은 횃대 먹이 활동으로 알려져 있는데, 개체는 먹이 활동 횃대에 앉아 먹이가 지나가기를 기다린 다음, 먹이를 잡기 위해 날아간다.[1] 먹이 활동은 보통 지상 5m 위에서 일어난다.[9] 저녁박쥐는 요막에서 먹이를 잡아 입으로 옮길 수 있지만, 관박쥐는 요막을 사용하여 먹이를 잡지 않는다. 적어도 한 종인 큰관박쥐는 날개 끝에서 손가락뼈를 구부려 먹이를 잡은 다음 입으로 옮기는 것으로 기록되어 있다.[17][26] 대부분의 관박쥐는 야행성이며 밤에 사냥하지만, 블라이스 관박쥐는 티오만 섬에서 낮 동안 먹이 활동을 하는 것으로 알려져 있다. 이는 섬에 주행성 조류(낮에 활동하는 새) 포식자가 부족하기 때문인 것으로 추정된다.[27]관박쥐는 특히 작고 둥근 날개 끝, 낮은 날개 하중(체질량에 비해 날개가 크다는 의미), 높은 캠버를 가지고 있다. 이러한 요소들은 민첩성을 증가시켜 느린 속도에서도 빠르고 좁은 회전을 할 수 있게 해준다.[25] 모든 박쥐에 비해 관박쥐의 날개 길이는 체형에 비해 일반적이며, 날개 길이와 날개 면적을 관련시키는 가로 세로 비는 평균이거나 평균보다 낮다. 루펠 관박쥐, 힐데브란트 관박쥐, 랜더 관박쥐, 스위니 관박쥐와 같은 일부 종은 특히 넓은 총 날개 면적을 가지고 있지만, 대부분의 관박쥐 종은 평균적인 날개 면적을 가지고 있다.[25]
4. 2. 먹이 및 사냥
관박쥐는 곤충을 먹지만, 거미와 같은 다른 절지동물도 섭취하며, 두 가지 주요한 먹이 활동 전략을 사용한다. 첫 번째 전략은 나무와 덤불 사이를 천천히 낮은 고도로 비행하는 것이다. 이 전략을 사용하는 일부 종은 먹이 위를 맴돌며 기찰하여 기질에서 먹이를 잡아낼 수 있다. 다른 전략은 횃대 먹이 활동으로 알려져 있는데, 개체는 먹이 활동 횃대에 앉아 먹이가 지나가기를 기다린 다음, 먹이를 잡기 위해 날아간다.[1] 먹이 활동은 보통 지상 5m 위에서 일어난다.[9] 저녁박쥐는 요막에서 먹이를 잡아 입으로 옮길 수 있지만, 관박쥐는 요막을 사용하여 먹이를 잡지 않는다. 적어도 한 종인 큰관박쥐는 날개 끝에서 손가락뼈를 구부려 먹이를 잡은 다음 입으로 옮기는 것으로 기록되어 있다.[17][26] 대부분의 관박쥐는 야행성이며 밤에 사냥하지만, 블라이스 관박쥐는 티오만 섬에서 낮 동안 먹이 활동을 하는 것으로 알려져 있다. 이는 섬에 주행성 조류(낮에 활동하는 새) 포식자가 부족하기 때문인 것으로 추정된다.[27]특히 작고 둥근 날개 끝, 낮은 날개 하중(체질량에 비해 날개가 크다는 의미), 높은 캠버를 가지고 있다. 이러한 요소들은 민첩성을 증가시켜 느린 속도에서도 빠르고 좁은 회전을 할 수 있게 해준다.[25]
4. 3. 번식 및 수명
관박쥐류의 짝짓기 체계는 잘 알려져 있지 않다. 2000년 연구에 따르면, 관박쥐류 중 약 4%만이 짝짓기 체계에 대한 정보가 발표되었으며, 이는 큰꼬리박쥐과와 함께 종 다양성에 비해 가장 적은 관심을 받은 것이다. 큰관박쥐는 수컷이 영역을 설정하고 방어하여 여러 암컷을 유인하는 일부다처제 짝짓기 체계를 가지고 있는 것으로 보인다. 그러나 ''Rhinolophus sedulus''는 일부일처제로 여겨지는 몇 안 되는 박쥐 종 중 하나이다 (2000년 기준으로 17종의 박쥐만이 일부일처제로 인정받고 있다).[28] 온대 종의 일부는 가을에 연간 번식기를 가지는 반면, 다른 종은 봄에 짝짓기를 한다.[15]많은 관박쥐 종은 암컷 정자 저장을 통해 수정이 지연되는 적응을 보인다. 이는 특히 온대 종에서 흔하다. 동면하는 종에서 정자 저장 시기는 동면 시기와 일치한다.[1] 랜더 관박쥐와 같은 다른 종은 배아 휴지를 가지는데, 이는 교미 직후 수정이 일어나지만 접합자가 오랜 기간 동안 자궁벽에 착상하지 않는 것을 의미한다.[17] 큰 관박쥐는 배아 발달이 지연되는 적응을 보이는데, 이는 암컷이 무활동에 들어갈 경우 배아의 성장이 조건부로 지연된다는 것을 의미한다. 이로 인해 수정과 출산 사이의 간격이 2~3개월로 달라진다.[29]
임신 기간은 약 7주이며, 한 마리의 새끼(유충)가 태어난다. 개체는 2세가 되면 성적으로 성숙해진다. 수명은 일반적으로 6~7년을 넘지 않지만, 일부 개체는 매우 오래 살 수 있다. 큰 관박쥐 개체는 한 번 띠를 부착한 후 30년 후에 다시 발견되기도 했다.[15]
4. 4. 행동 및 사회 시스템
관박쥐는 곤충을 먹지만, 거미와 같은 다른 절지동물도 섭취하며, 두 가지 주요한 먹이 활동 전략을 사용한다. 첫 번째 전략은 나무와 덤불 사이를 천천히 낮은 고도로 비행하는 것이다. 이 전략을 사용하는 일부 종은 먹이 위를 맴돌며 기찰하여 기질에서 먹이를 잡아낼 수 있다. 다른 전략은 횃대 먹이 활동으로 알려져 있는데, 개체는 먹이 활동 횃대에 앉아 먹이가 지나가기를 기다린 다음, 먹이를 잡기 위해 날아간다.[1] 먹이 활동은 보통 지상 5m 위에서 일어난다.[9] 저녁박쥐는 요막에서 먹이를 잡아 입으로 옮길 수 있지만, 관박쥐는 요막을 사용하여 먹이를 잡지 않는다. 적어도 한 종인 큰관박쥐는 날개 끝에서 손가락뼈를 구부려 먹이를 잡은 다음 입으로 옮기는 것으로 기록되어 있다.[17][26] 대부분의 관박쥐는 야행성이며 밤에 사냥하지만, 블라이스 관박쥐는 티오만 섬에서 낮 동안 먹이 활동을 하는 것으로 알려져 있다. 이는 섬에 주행성 조류(낮에 활동하는 새) 포식자가 부족하기 때문인 것으로 추정된다.[27]특히 작고 둥근 날개 끝, 낮은 날개 하중(체질량에 비해 날개가 크다는 의미), 높은 캠버를 가지고 있다. 이러한 요소들은 민첩성을 증가시켜 느린 속도에서도 빠르고 좁은 회전을 할 수 있게 해준다.[25] 모든 박쥐에 비해 관박쥐의 날개 길이는 체형에 비해 일반적이며, 날개 길이와 날개 면적을 관련시키는 가로 세로 비는 평균이거나 평균보다 낮다. 루펠 관박쥐, 힐데브란트 관박쥐, 랜더 관박쥐, 스위니 관박쥐와 같은 일부 종은 특히 넓은 총 날개 면적을 가지고 있지만, 대부분의 관박쥐 종은 평균적인 날개 면적을 가지고 있다.[25]
4. 5. 포식자 및 기생충
관박쥐는 곤충을 먹지만, 거미와 같은 다른 절지동물도 먹이로 삼으며, 두 가지 주요 사냥 전략을 사용한다. 첫 번째는 나무와 덤불 사이를 천천히 낮은 고도로 비행하는 것이다. 이 전략을 사용하는 일부 종은 먹이 위를 맴돌며 기찰하여 기질에서 먹이를 잡아내기도 한다. 두 번째는 횃대 먹이 활동으로, 먹이 활동 횃대에 앉아 먹이가 지나가기를 기다렸다가 날아가 잡는 방식이다.[1] 먹이 활동은 주로 지상 5m 위에서 이루어진다.[9] 저녁박쥐와 달리 관박쥐는 요막을 사용하여 먹이를 잡지 않는다. 큰관박쥐는 날개 끝 손가락뼈를 구부려 먹이를 잡은 다음 입으로 옮기는 것으로 알려져 있다.[17][26] 대부분의 관박쥐는 야행성이지만, 블라이스 관박쥐는 티오만 섬에서 낮 동안 먹이 활동을 하는 것으로 알려져 있는데, 이는 섬에 낮에 활동하는 새인 포식자가 없기 때문으로 추정된다.[27]5. 인간과의 관계
관박쥐는 코로나바이러스의 근원으로서 공중 보건 및 동물원성 질병에 특히 관심을 받고 있다.
5. 1. 질병 매개체
관박쥐는 코로나바이러스의 근원으로서 공중 보건 및 동물원성 질병에 특히 중요하다.2002–2004년 SARS 발생 이후, 여러 동물 종이 사인이 되는 코로나바이러스인 SARS-CoV의 자연 숙주로 조사되었다. 2003년부터 2018년까지 관박쥐에서 47개의 SARS 관련 코로나바이러스가 발견되었다.[47] 2019년에는 중국 우한의 수산 시장이 SARS-CoV-2 발생과 연관되었다. SARS-CoV-2의 유전자 분석 결과, 관박쥐에서 발견된 바이러스와 매우 유사했다.[46]
SARS 발생 이후, 작은관박쥐는 혈청 반응 양성이었고, 큰관박쥐는 바이러스에 대해서만 양성 반응을 보였으며, 큰귀관박쥐, 중국관박쥐, 피어슨관박쥐는 혈청 반응 및 바이러스 양성 반응을 보였다.[47][48] 박쥐의 바이러스는 SARS-CoV와 88–92% 유사했다.[49] SARS 유사 코로나바이러스의 종내 다양성은 상동 재조합에 의해 ''Rhinolophus sinicus''에서 발생한 것으로 보인다.[50] ''R. sinicus''는 인간 SARS-CoV의 직접적인 조상을 가지고 있었을 것이다. 관박쥐가 SARS 관련 코로나바이러스의 자연 숙주인 것으로 보이지만, 인간은 바이러스의 중간 숙주로 확인된 감염된 사향고양이와 접촉하여 감염되었을 가능성이 높다.[49]
2003년부터 2018년까지 박쥐에서 47개의 SARS 관련 코로나바이러스가 검출되었으며, 그 중 45개는 관박쥐에서 검출되었다.
[47]
SARS-CoV-2가 검출된 우한 시장에서는 96%가 중간관박쥐에서 분리된 바이러스와 유사했다. SARS-CoV-2의 진화적 기원에 대한 연구[51]는 박쥐가 SARS-CoV-2의 자연 숙주임을 나타낸다. 중간 숙주가 관련되었을 수 있지만, 바이러스가 어떻게 인간에게 전염되었는지는 불분명하다. 한때 말레이판골린으로 여겨졌지만,[52] 2020년 7월 발표된 논문에서는 판골린에서 인간으로의 전염 증거가 발견되지 않았다.[51]
관박쥐류는 오르토레오바이러스류, 플라비바이러스류 및 한타바이러스류와도 관련이 있다. 이들은 포유류 오르토레오바이러스(MRV)에 대해 양성 반응을 보였다. 붉은관박쥐는 인도 남부에서 알려진 진드기 매개 바이러스성 출혈열인 카야사누르 숲병에 대해 혈청 양성 반응을 보였다. 카야사누르 숲병은 감염된 진드기에 물려 인간에게 전염되며, 치사율은 2~10%이다.[54] 롱취안 바이러스는 한타바이러스의 일종으로, 중간관박쥐, 중국붉은관박쥐, 일본작은관박쥐에서 검출되었다.[55]
5. 2. 식용 및 약용
소형박쥐는 과일박쥐만큼 많이 사냥되지 않는다. 식충성 박쥐 종의 8%만이 식용으로 사냥되는 반면, 구세계 열대 지역의 모든 과일박쥐 종의 절반이 사냥된다. 말굽박쥐는 특히 사하라 이남 아프리카에서 식용으로 사냥된다. 아프리카에서 사냥되는 종에는 할시온 말굽박쥐 (''R. alcyone''), 기니 말굽박쥐 (''R. guineensis''), 힐 말굽박쥐 (''R. hilli''), 힐스 말굽박쥐 (''R. hillorum''), 맥클로드 말굽박쥐 (''R. maclaudi''), 루웬조리 말굽박쥐, 숲 말굽박쥐 (''R. silvestris''), 지아마 말굽박쥐 (''R. ziama'') 등이 있다.[56] 동남아시아에서는 마샬 말굽박쥐 (''R. marshalli'')가 미얀마에서, 대형 적갈색 말굽박쥐 (''R. rufus'')가 필리핀에서 섭취된다.[56]인도 북동부의 아오 나가족은 말굽박쥐의 살을 천식 치료에 사용하는 것으로 알려져 있다. 생태 인류학자 윌 툴라다르-더글러스는 네팔의 네와르족이 다른 종들과 함께 "거의 확실하게" 말굽박쥐를 사용하여 ''치카 라파 와사''("박쥐 기름")를 준비한다고 말했다. 죽은 박쥐는 말아서 겨자유가 들어 있는 밀봉된 항아리에 넣는다. 기름은 독특하고 불쾌한 냄새가 나면 준비된다. 박쥐 기름의 전통적인 약용 용도에는 귀에 들어가 뇌를 갉아먹는 지네로 알려진 "귀 벌레"를 제거하는 것이 포함된다. 이는 아마도 편두통에 대한 전통적인 설명일 것이다. 또한 탈모와 부분 마비의 치료제로도 사용된다.[57] 세네갈에서는 말굽박쥐가 정신 질환 치료를 위한 약초에 사용된다는 일화적인 보고가 있으며, 베트남에서는 한 제약 회사가 매년 50ton의 말굽박쥐 구아노를 약용으로 사용한다고 보고했다.[58]
5. 3. 보존
2023년 기준으로 IUCN은 94종의 관박쥐류를 평가했다. 이들은 다음과 같은 IUCN 상태를 가지고 있다.[59]| 상태 | 종 수 |
|---|---|
| 위급종 | 1종 (힐 관박쥐) |
| 위기종 | 13종 |
| 취약종 | 5종 |
| 준위협종 | 9종 |
| 관심대상종 | 51종 |
| 정보부족 | 15종 |
모든 동굴에 서식하는 박쥐와 마찬가지로, 동굴에 서식하는 관박쥐류는 동굴 서식지 교란에 취약하다. 교란에는 박쥐 구아노 채취, 석회암 채석, 동굴 관광 등이 포함될 수 있다.[45]
6. 하위 종
| 이름 | 학명 |
|---|---|
| 뾰족관박쥐 | R. acuminatus |
| 애덤관박쥐 | R. adami |
| 중간관박쥐 | R. affinis |
| 할퀴온관박쥐 | R. alcyone |
| 궁형관박쥐 | R. arcuatus |
| 베돔관박쥐 | R. beddomei |
| 블라시우스관박쥐 | R. blasii |
| 부하라관박쥐 | R. bocharicus |
| 보르네오관박쥐 | R. borneensis |
| 카누트관박쥐 | R. canuti |
| 케이프관박쥐 | R. capensis |
| 술라웨시관박쥐 | R. celebensis |
| 키우키관박쥐 | R. chiewkweeae |
| 조프루아관박쥐 | R. clivosus |
| 페테르스관박쥐 | R. coelophyllus |
| 안다만관박쥐 | R. cognatus |
| 코언관박쥐 | R. cohenae |
| 볼록관박쥐 | R. convexus |
| 작은일본관박쥐 | R. cornutus |
| 크리그관박쥐 | R. creaghi |
| 달링관박쥐 | R. darlingi |
| 데켄관박쥐 | R. deckenii |
| 덴트관박쥐 | R. denti |
| 엘러퀀트관박쥐 | R. eloquens |
| 지중해관박쥐 | R. euryale |
| 넓은귀관박쥐 | R. euryotis |
| 관박쥐 | R. ferrumequinum |
| 포르모사양털관박쥐 | R. formosae |
| 프랜시스양털관박쥐 | R. francisi |
| 루펠관박쥐 | R. fumigatus |
| 기니관박쥐 | R. guineensis |
| 힐데브란트관박쥐 | R. hildebrandtii |
| 힐관박쥐 | R. hilli |
| 고지대관박쥐 | R. hillorum |
| 작은관박쥐 | R. hipposideros |
| 호라체크관박쥐 | R. horaceki |
| 화난관박쥐 | R. huananus |
| 이마이즈미관박쥐 | R. imaizumii |
| 필리핀숲관박쥐 | R. inops |
| 카후지관박쥐 | R. kahuzi |
| 섬관박쥐 | R. keyensis |
| 랜더관박쥐 | R. landeri |
| 블라이스관박쥐 | R. lepidus |
| 말레이시아양털관박쥐 | R. luctoides |
| 양털관박쥐 | R. luctus |
| 마부산관박쥐 | R. mabuensis |
| 매클라우드관박쥐 | R. maclaudi |
| 큰귀관박쥐 | R. macrotis |
| 마두라관박쥐 | R. madurensis |
| 마엔델레오관박쥐 | R. maendeleo |
| 말라얀관박쥐 | R. malayanus |
| 마셜관박쥐 | R. marshalli |
| 동부관박쥐 | R. megaphyllus |
| 미헤이관박쥐 | R. mehelyi |
| 인도차이나갈색관박쥐 | R. microglobosus |
| 인도관박쥐 | R. mitratus |
| 포르모사작은관박쥐 | R. monoceros |
| 티모르관박쥐 | R. montanus |
| 산악관박쥐 | R. monticolus |
| 모잠비크관박쥐 | R. mossambicus |
| 아남바스관박쥐 | R. nereis |
| 오스굿관박쥐 | R. osgoodi |
| 부레관박쥐 | R. paradoxolophus |
| 피어슨관박쥐 | R. pearsonii |
| 필리핀관박쥐 | R. philippinensis |
| 꼬마관박쥐 | R. pusillus |
| 왕관박쥐 | R. rex |
| 반도관박쥐 | R. robinsoni |
| 적갈색관박쥐 | R. rouxii |
| 큰적갈색관박쥐 | R. rufus |
| 루웬조리관박쥐 | R. ruwenzorii |
| 사케지관박쥐 | R. sakejiensis |
| 슈니츨러관박쥐 | R. schnitzleri |
| 작은양털관박쥐 | R. sedulus |
| 샤멜관박쥐 | R. shameli |
| 쇼트리지관박쥐 | R. shortridgei |
| 태국잎코박쥐 | R. siamensis |
| 숲관박쥐 | R. silvestris |
| 부시벨트관박쥐 | R. simulator |
| 중국적갈색관박쥐 | R. sinicus |
| 스미더스관박쥐 | R. smithersi |
| 작은갈색관박쥐 | R. stheno |
| 작은네팔관박쥐 | R. subbadius |
| 작은적갈색관박쥐 | R. subrufus |
| 스위니관박쥐 | R. swinnyi |
| 태국관박쥐 | R. thailandensis |
| 토마스관박쥐 | R. thomasi |
| 삼엽형관박쥐 | R. trifoliatus |
| 노랑얼굴관박쥐 | R. virgo |
| 윌러드관박쥐 | R. willardi |
| 중국관박쥐 | R. xinanzhongguoensis |
| 돕슨관박쥐 | R. yunanensis |
| 지아마관박쥐 | R. ziama |
7. 계통 분류
| 1 | 2 | 3 | 4 |
|---|---|---|---|
| 큰박쥐과 | 흡반발박쥐과 | 짧은꼬리박쥐과 | 큰귀박쥐과 |
| 대꼬리박쥐과 | 원반날개박쥐과 | 불독박쥐과 | 뭉툭귀박쥐아과 |
| 키티돼지코박쥐과 | 민발톱박쥐과 | 유령얼굴박쥐과 | 큰귀박쥐아과 |
| 생쥐꼬리박쥐과 | 깔때기귀박쥐과 | 주걱박쥐과 | 긴가락박쥐과 |
| 위흡혈박쥐과 | 넓은 의미의 애기박쥐과 | 좁은 의미의 애기박쥐과 | |
| 틈새얼굴박쥐과 | |||
| 관박쥐과 | |||
| 잎코박쥐과 |
가와다 외 (2018)에 따른 계통 분류는 다음과 같다.
- '''굽은코박쥐과'''
- * '''굽은코박쥐속'''
- ** 뾰족코잎코박쥐
- ** 콩고굽은코박쥐
- ** 나카굽은코박쥐
- ** 알키온굽은코박쥐
- ** 아치굽은코박쥐
- ** 베드메굽은코박쥐
- ** 브라시우스굽은코박쥐
- ** 아프가니스탄굽은코박쥐
- ** 보르네오굽은코박쥐
- ** 카누토굽은코박쥐
- ** 케이프굽은코박쥐
- ** 셀레베스굽은코박쥐
- ** 조프로이굽은코박쥐
- ** 피터굽은코박쥐
- ** 안다만굽은코박쥐
- ** 볼록굽은코박쥐
- ** 작은굽은코박쥐
- ** 크리굽은코박쥐
- ** 다링굽은코박쥐
- ** 데켄굽은코박쥐
- ** 덴티굽은코박쥐
- ** 웅변굽은코박쥐
- ** 지중해굽은코박쥐
- ** 큰귀굽은코박쥐
- ** 굽은코박쥐
- ** 타이완굽은코박쥐
- ** 아프리카굽은코박쥐
- ** 기니굽은코박쥐
- ** 힐데브란트굽은코박쥐
- ** 힐굽은코박쥐
- ** 업랜드굽은코박쥐
- ** 작은굽은코박쥐
- ** 이리오모테굽은코박쥐
- ** 미다나오굽은코박쥐
- ** 케이굽은코박쥐
- ** 랜더굽은코박쥐
- ** 브라이스굽은코박쥐
- ** 털굽은코박쥐
- ** 맥클라우드굽은코박쥐
- ** 큰귀굽은코박쥐
- ** 마두라굽은코박쥐
- ** 멘데레오굽은코박쥐
- ** 말레이굽은코박쥐
- ** 마셜굽은코박쥐
- ** 남부굽은코박쥐
- ** 메헬리굽은코박쥐
- ** 헤어밴드굽은코박쥐
- ** 타이완작은굽은코박쥐
- ** 몬태나굽은코박쥐
- ** 네레이스굽은코박쥐
- ** 오스굿굽은코박쥐
- ** 파라독솔로푸스굽은코박쥐
- ** 피어슨굽은코박쥐
- ** 야에야마작은굽은코박쥐
- ** 필리핀굽은코박쥐
- ** 오키나와작은굽은코박쥐
- ** 렉스굽은코박쥐
- ** 로빈슨굽은코박쥐
- ** 루굽은코박쥐
- ** 붉은굽은코박쥐
- ** 알버틴굽은코박쥐
- ** 사케지굽은코박쥐
- ** 세둘루스굽은코박쥐
- ** 샤멜굽은코박쥐
- ** 쇼트리지굽은코박쥐
- ** 시암굽은코박쥐
- ** 실베스트리스굽은코박쥐
- ** 시뮬레이터굽은코박쥐
- ** 중국굽은코박쥐
- ** 스테노굽은코박쥐
- ** 차굽은코박쥐
- ** 붉은털굽은코박쥐
- ** 스윈니굽은코박쥐
- ** 토마스굽은코박쥐
- ** 삼잎굽은코박쥐
- ** 처녀굽은코박쥐
- ** 두손굽은코박쥐
- ** 지아마굽은코박쥐
참조
[1]
서적
Horseshoe Bats of the World: (Chiroptera: Rhinolophidae)
Alana Books
[2]
논문
An attempt at a division of the family Vespertilionidae into groups
https://biodiversity[...]
[3]
서적
Classification of mammals: above the species level
https://books.google[...]
Columbia University Press
[4]
논문
Four New Bat Species (Rhinolophus hildebrandtii Complex) Reflect Plio-Pleistocene Divergence of Dwarfs and Giants across an Afromontane Archipelago
[5]
웹사이트
Family Rhinolophidae
https://www.departme[...]
Bucknell University
2020-07-23
[6]
논문
Integrated fossil and molecular data reconstruct bat echolocation
[7]
웹사이트
Family Hipposideridae
https://www.departme[...]
Bucknell University
2005
[8]
논문
Surprising Genetic diversity in ''Rhinolophus luctus'' (Chiroptera: Rhinolophidae) from peninsular Malaysia: Description of a new species based on genetic and morphological characters
[9]
서적
Grzimek's Animal Life Encyclopedia Mammals II
Gale
[10]
논문
Bat systematics in the light of unconstrained analyses of a comprehensive molecular supermatrix
[11]
논문
Molecular phylogenetics of the African horseshoe bats (Chiroptera: Rhinolophidae): Expanded geographic and taxonomic sampling of the Afrotropics
[12]
논문
A List of the Genera and Families of Mammal
https://books.google[...]
[13]
서적
Catalogue of the Fossil Mammalia in the British Museum, (Natural History): The orders Primates, Chiroptera, Insectivora, Carnivora, and Rodentia
https://www.biodiver[...]
Order of the Trustees
[14]
논문
Phylogenetic analyses of the bat family Rhinolophidae
http://www.faunapara[...]
[15]
서적
Walker's Bats of the World
https://archive.org/[...]
JHU Press
[16]
논문
Morphology, function, and phylogenetic significance of pubic nipples in bats (Mammalia, Chiroptera)
http://digitallibrar[...]
1993
[17]
서적
Mammals of Africa
A&C Black
2013
[18]
서적
Fauna of Australia
https://www.environm[...]
AGPS Canberra
1989
[19]
논문
Dental ontogeny in the Old World leaf-nosed bats (Rhinolophidae, Hipposiderinae)
[20]
서적
Biology of Bats
Academic Press
[21]
논문
Molecular phylogenetics and historical biogeography of Rhinolophus bats
[22]
논문
The evolution of echolocation in bats
[23]
논문
Correlations between call frequency and ear length in bats belonging to the families Rhinolophidae and Hipposideridae
[24]
논문
The furrows of Rhinolophidae revisited
[25]
논문
Ecological morphology and flight in bats (Mammalia; Chiroptera): Wing adaptations, flight performance, foraging strategy and echolocation
[26]
논문
The role of the flight membranes in insect capture by bats
[27]
논문
Into the light: atypical diurnal foraging activity of Blyth's horseshoe bat, ''Rhinolophus lepidus'' (Chiroptera: Rhinolophidae) on Tioman Island, Malaysia
2018-12-19
[28]
서적
Reproductive Biology of Bats
[29]
서적
Mammals of Africa
A&C Black
2013
[30]
논문
Hibernation and daily torpor in Australian mammals
[31]
서적
Bats: From Evolution to Conservation
https://books.google[...]
Oxford University Press
[32]
논문
Hibernation and Torpor in Tropical and Subtropical Bats in Relation to Energetics, Extinctions, and the Evolution of Endothermy
[33]
논문
Bat Predation by Spiders
[34]
논문
Bats as prey of diurnal birds: a global perspective: Predation of bats by diurnal birds
2016
[35]
논문
Bat predation by long-eared Owls in mediterranean and temperate regions of southern Europe
https://digital.csic[...]
[36]
논문
Snake predation on bats in Europe: New cases and a regional assessment
http://real.mtak.hu/[...]
[37]
논문
Curiosity killed the bat: Domestic cats as bat predators
[38]
논문
Presence of humans and domestic cats affects bat behaviour in an urban nursery of greater horseshoe bats (''Rhinolophus ferrumequinum'')
[39]
논문
Variation in ectoparasite load in the Mehely's horseshoe bat, ''Rhinolophus mehelyi'' (Chiroptera: Rhinolophidae) in a nursery colony in western Iran
[40]
논문
Description of a new tick species, ''Ixodes collaris'' n. sp. (Acari: Ixodidae), from bats (Chiroptera: Hipposideridae, Rhinolophidae) in Vietnam
[41]
논문
Distribution and Specificity of Host-Parasite Associations of Fleas (Siphonaptera) in the Central Caucasus
[42]
논문
Gastrointestinal digeneans (Platyhelminthes: Trematoda) of horseshoe and vesper bats (Chiroptera: Rhinolophidae and Vespertilionidae) in Serbia
[43]
논문
First report of ''Potorolepis'' Spassky, 1994 (Eucestoda: Hymenolepididae) from China, with description of a new species in bats (Chiroptera:: Rhinolophidae)
[44]
논문
Molecular phylogenetics and historical biogeography of Rhinolophus bats
[45]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer, Cham
[46]
웹사이트
Novel Coronavirus (2019-nCoV) Situation Report
https://www.who.int/[...]
2020-02-11
[47]
논문
Molecular epidemiology, evolution and phylogeny of SARS coronavirus
[48]
논문
A review of studies on animal reservoirs of the SARS coronavirus
[49]
논문
Review of Bats and SARS
[50]
논문
Intraspecies diversity of SARS-like coronaviruses in Rhinolophus sinicus and its implications for the origin of SARS coronaviruses in humans
[51]
논문
Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic
[52]
논문
COVID-19: A novel zoonotic disease caused by a coronavirus from China: What we know and what we don't
[53]
서적
Bats and Human Health: Ebola, SARS, Rabies and Beyond
https://books.google[...]
John Wiley & Sons
[54]
논문
Kyasanur forest disease: An epidemiological view in India
[55]
논문
Phylogeny and Origins of Hantaviruses Harbored by Bats, Insectivores, and Rodents
[56]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer
[57]
논문
The Use of Bats as Medicine Among the Newars
[58]
논문
Bats as materia medica: An ethnomedical review and implications for conservation
https://www.research[...]
[59]
웹사이트
Taxonomy=Rhinolophidae
https://www.iucnredl[...]
2023-12-28
[60]
문서
世界哺乳類標準和名目録
https://doi.org/10.1[...]
[61]
웹사이트
キクガシラコウモリ
https://kotobank.jp/[...]
[62]
문서
[63]
간행물
http://www.molekular[...]
2002
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com