반삭동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
반삭동물은 몸이 구문부, 금부, 동부로 나뉘며, 척삭동물의 척삭과 유사한 반삭을 가진 해양 무척추동물이다. 소화계, 개방 순환계를 가지며, 일부 종은 탄산칼슘을 생물화한다. 장새류와 익새류로 분류되며, 극피동물과 함께 척삭동물의 가장 가까운 친척으로 척삭동물의 진화 연구에 중요하다. 반삭동물은 장새류와 익새류로 나뉘며, 두 강은 바다 지렁이 형태를 띤다.
더 읽어볼만한 페이지
- 반삭동물 - 필석
필석은 캄브리아기부터 석탄기 지층에서 화석으로 발견되는 고생대 초기의 멸종된 해양 동물 군체로, 반삭동물로 분류되며, 지질 시대 구분에 중요한 표준 화석으로 활용되어 오르도비스기와 실루리아기 지층 연구에 기여한다. - 반삭동물 - 익새류
익새류는 반삭동물문에 속하며 촉수완으로 여과 섭식을 하는 작은 해양 동물로, 몸은 세 부분으로 나뉘고 군체 생활을 하며 장새류와 관련이 있고 일부 종은 대체 유전 암호를 사용한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.
| 반삭동물 - [생물]에 관한 문서 | |
|---|---|
| 분류 정보 | |
| 계 | 동물계 |
| 계급 없음 | 후구동물 |
| 문 | 반삭동물문 |
| 학명 | Hemichordata |
| 명명자 | Bateson, 1885 |
| 하위 분류 | 강 |
| 하위 분류 | 장새강 익새강 플랑크톤구강 필석강 |
| 일반 정보 | |
| 로마자 표기법 | Hemichordata |
| 발음 (IPA) | /ˌhɛmɪkɔːrˈdeɪtə/ |
| 발음 (재철자) | HEM-ih-kor-DAY-tə |
| 특징 | |
| 서식지 | 해양 |
| 분류군 | 후구동물 |
| 생물학적 특징 | |
| 섭식 방식 | 퇴적물 섭취 |
| 포식자 방어 | 2,4-디브로모페놀 분비 |
| 화석 기록 | |
| 출현 시기 | 미아오링세 (기원전 5억 1,400만 년) ~ 현재 |
| 최초 출현 시기 | 포르투나절 |
| 참고 문헌 | |
| 논문 | Sato, Atsuko; Rickards, RB; Holland, PWH (2008년 12월). "The origins of graptolites and other pterobranchs: a journey from 'Polyzoa'". Lethaia 41 (4): 303–316. doi:10.1111/j.1502-3931.2008.00123.x. Caron, J. B.; Conway Morris, S.; Cameron, C. B. (2013년). "Tubicolous enteropneusts from the Cambrian period". Nature 495 (7442): 503–506. doi:10.1038/nature12017. Giray, Cem; King, G. M. (1997년). "Predator deterrence and 2,4-dibromophenol conservation by the enteropneusts, Saccoglossus bromophenolosus and Protoglossus graveolens". Marine Ecology Progress Series 159: 229–238. doi:10.3354/meps159229. Sato, Atsuko; Bishop, JDD; Holland, PWH (2008년). "Developmental biology of pterobranch hemichordates: history and perspectives". Genesis 46 (11): 587–91. doi:10.1002/dvg.20395. |
| 잡지 | ROM (2020년 가을). "A Cambrian Puzzle: An Ancient, Fossilized Marine Worm Crawls into the Light". ROM Magazine. Toronto: Royal Ontario Museum. |
2. 특징

반삭동물의 몸은 근육 조직으로 이루어져 있으며, 전후 축을 따라 크게 세 부분으로 나뉜다. 앞쪽의 전체(prosome), 중간의 중체(mesosome), 그리고 뒤쪽의 후체(metasome)이다. 각 부분에는 체강이 존재하는데, 앞몸에는 하나, 가운데몸과 뒷몸에는 각각 한 쌍의 진체강이 있다. 이러한 체강 형성 방식은 극피동물과 유사하며, 반삭동물의 토르나리아(tornaria) 유생은 극피동물의 비핀나리아(bipinnaria) 유생과 매우 비슷하다.
몸 안에는 맹낭 형태의 구조물인 반삭(stomochord)이 있는데, 과거에는 척삭동물의 척삭과 관련이 있다고 여겨졌으나, 현재는 상동 관계가 아닌 수렴 진화의 결과일 가능성이 더 높다고 본다.[8]
몸의 구체적인 형태는 그룹에 따라 차이가 있다.
- 장새강 (Enteropneusta, 바다나리벌레): 몸이 벌레 모양이며, 앞쪽의 근육질이고 섬모가 있는 입촉(proboscis), 중간의 깃(collar), 뒤쪽의 몸통(trunk)으로 나뉜다. 입촉은 운동과 먹이 수집 및 운반에 사용된다. 입은 입촉과 깃 사이에 위치한다. 몸통은 가장 긴 부분으로, 인두, 식도, 긴 창자, 항문, 그리고 생식선을 포함한다. 인두에는 아가미틈(새열)이라 불리는 여러 개의 구멍이 있어 호흡과 여과 섭식에 관여하는데, 이는 척삭동물과의 공통된 특징이다. 항문 뒤 꼬리는 바다나리벌레과 Harrimaniidae의 어린 개체에서 관찰된다.[6]
- 익새강 (Pterobranchia, 깃갯나물벌레강): 전체는 운동과 군체(coenecium) 분비에 사용되는 근육질이고 섬모가 있는 두부 방패(cephalic shield)로 특화되어 있다. 중체는 여과 섭식에 사용되는 촉수가 달린 팔(한 쌍 또는 여러 쌍)로 확장된다. 후체(몸통)에는 고리 모양의 소화관과 생식선이 있으며, 무성 생식으로 생성된 다른 개체와 연결되는 수축성 자루로 이어진다. 속 ''Rhabdopleura''에서는 개충(zooid)이 공통 줄기 시스템을 통해 군체의 나머지 부분에 영구적으로 연결된다.
개방순환계를 가지며, 무색투명한 혈액이 순환한다. 심장은 등쪽에 위치한 심낭(pericardium)으로, 직접 혈액을 포함하지는 않고 등쪽 혈관의 박동을 통해 혈액 순환을 돕는다.[12]
신경계는 신경망과 세로 신경으로 구성되어 있으며, 뚜렷한 뇌는 없다.[9][10] 일부 종에서는 발생 초기 단계에 속이 빈 등쪽 신경관이 나타나는데, 이는 척삭동물과 공통 조상으로부터 물려받은 원시적인 특징일 수 있다.[8]
일부 종은 탄산칼슘을 이용하여 생물 광물화를 한다.[11] 대부분 암수딴몸이며, 체외수정을 통해 번식한다.
3. 구조
반삭동물의 몸은 크게 구문부(prosome, 앞몸), 금부(mesosome, 가운데몸), 동부(metasome, 뒷몸)의 세 부분으로 나뉜다.[6] 몸 안에는 척삭동물의 척삭과 유사한 구조로 여겨졌던 반삭(stomochord)이라는 앞창자의 게실이 존재하지만, 이는 상동 관계가 아닌 수렴 진화의 결과일 가능성이 높다.[8] 몸의 각 부분에는 체강이 존재하는데, 앞몸에는 하나, 가운데몸과 뒷몸에는 각각 한 쌍의 진체강을 갖는다. 반삭동물은 장체강동물로서 체강 형성 방식이 극피동물과 유사하며, 유생인 토르나리아 유생 또한 극피동물의 비핀나리아 유생과 매우 비슷하다.
=== 장새류 (Enteropneusta) ===
장새강의 대표적인 예인 도토리 지렁이(Acorn worm)는 몸이 지렁이처럼 길고 유연하며, 주로 얕은 바다의 모래나 진흙 속에서 서식한다. 몸길이는 수 cm에서 2m에 이르기까지 다양하다.
- 구문부(입촉, Proboscis): 몸 앞쪽 끝에 위치하며 근육질이고 섬모가 나 있어, 땅을 파고 이동하거나 먹이 입자를 모아 입으로 운반하는 데 사용된다. 영어권에서는 이 부분이 도토리를 닮았다 하여 'acorn worm'이라 부른다.
- 금부(깃, Collar): 구문부 바로 뒤에 위치하며 짧은 고리 모양이다. 입은 구문부와 금부 사이에 열려 있다.
- 동부(몸통, Trunk): 몸의 가장 긴 부분으로, 앞쪽에는 호흡과 먹이 여과에 관여하는 여러 쌍의 아가미틈(인두열)이 나란히 배열된 인두 부위가 있다. 그 뒤로 식도, 긴 창자, 그리고 몸 뒤쪽 끝의 항문으로 이어지는 소화관이 있다. 생식선도 동부에 위치한다. 일부 종(Harrimaniidae과)의 어린 개체는 항문 뒤에 꼬리가 존재하기도 한다.[6]
=== 익새류 (Pterobranchia) ===
익새류는 대부분 군체를 이루어 생활하며, 자신이 분비한 물질로 만든 관(coenecium) 속에서 산다. 몸길이는 보통 수 mm 정도로 작다.
- 구문부(두부 보호막, Cephalic shield): 근육질의 섬모성 기관으로, 이동하거나 관(coenecium)을 분비하는 데 사용된다.
- 금부(촉수관, Lophophore): 한 쌍(''Rhabdopleura'' 속) 또는 여러 쌍(''Cephalodiscus'' 속)의 촉수가 달린 팔이 뻗어 나와 있어, 물속의 작은 먹이 입자를 걸러 먹는 여과 섭식에 사용된다.
- 동부(몸통, Trunk): 고리 모양의 소화관과 생식선이 들어있다. 몸통 끝은 수축성 줄기(stalk)로 이어지는데, 이 줄기는 무성 생식으로 만들어진 새로운 개체를 연결하는 역할을 한다. ''Cephalodiscus'' 속에서는 새로운 개체가 완전히 자랄 때까지 어미의 줄기에 붙어 있다가 분리되지만, ''Rhabdopleura'' 속에서는 공통된 포복지(stolon) 시스템을 통해 군체의 다른 개체들과 영구적으로 연결된다.
=== 기타 구조적 특징 ===
- 신경계: 일부 종에서는 발생 초기 단계에 속이 빈 신경관이 나타나는데, 이는 척삭동물과 다른 후구동물의 공통 조상이 가졌던 원시적인 특징일 수 있다.[8] 하지만 성체는 명확한 뇌 구조 없이 신경망과 세로 신경으로 구성된 비교적 단순한 신경계를 가진다.[9][10]
- 순환계: 개방혈관계이며, 등쪽에 위치한 심장(심포)이 박동하여 무색투명한 혈액을 순환시킨다.
- 골격: 일부 종은 탄산 칼슘으로 이루어진 구조물을 만들기도 한다.[11]
- 생식: 대부분 암수딴몸이며, 장새류는 체외수정을 하고, 익새류도 체외수정을 할 것으로 추정된다. 익새류는 무성 생식으로 군체를 형성하기도 한다.
4. 발달과 성장
극피동물과 함께 보대동물군(步帶動物群)에 속하는 반삭동물은 무척추동물 중에서 현존하는 척삭동물과 계통발생학적으로 가장 가까운 친척으로 여겨진다.[13] 이러한 이유로 반삭동물은 척삭동물의 진화적 기원을 연구하는 데 중요한 대상이 된다. 반삭동물은 크게 장새류(Enteropneusta)와 익새류(Pterobranchia)의 두 강(綱)으로 나뉘며, 모두 바다 지렁이와 유사한 형태를 가진다.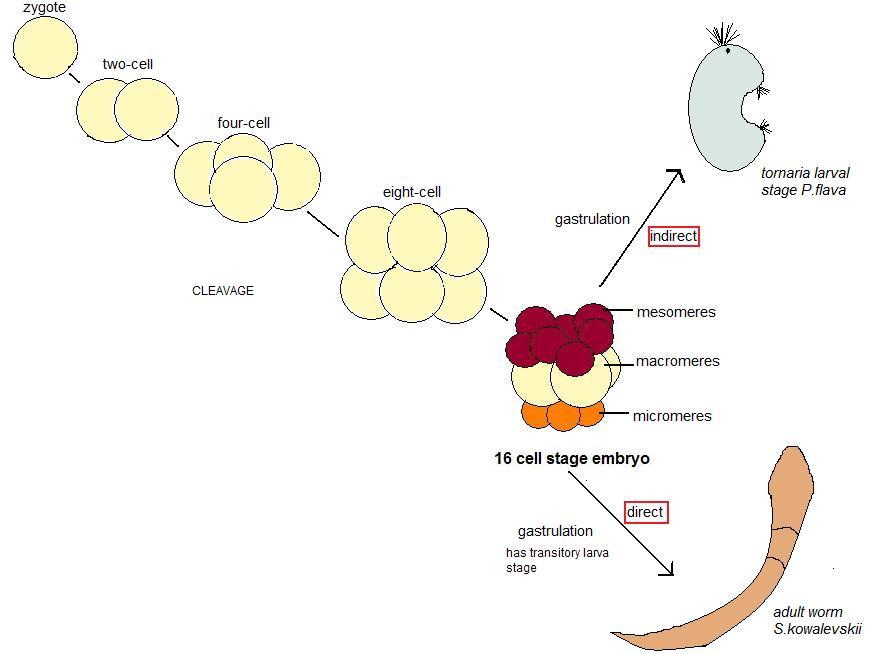
반삭동물의 발생 방식은 크게 두 가지로 나뉜다. 하나는 간접 발생이고 다른 하나는 직접 발생이다.[13] 장새류에서는 이 두 가지 발생 전략이 모두 나타난다. 간접 발생 전략을 따르는 종들은 플랑크톤을 먹고 자라는 자유 유영 단계인 토르나리아 유생(Tornaria larva) 시기를 거친 후 성체로 변태한다.[14] 반면, 직접 발생을 하는 종들은 이러한 유생 단계를 거치지 않고 바로 성체의 형태로 발달한다.
널리 연구되는 반삭동물 종으로는 간접 발생을 하는 ''Ptychodera flava''와 직접 발생을 하는 ''Saccoglossus kowalevskii''가 있다. 반삭동물 발생에 대한 자세한 연구는 주로 직접 발생하는 종들을 통해 이루어졌다. 익새류 중에서는 영국 플리머스와 버뮤다 등지에서 발견되는 ''Rhabdopleura'' 속이 가장 많이 연구되었다.[15][16][17][18]
반삭동물은 척삭과 유사한 구조로 여겨졌던 입맹관(stomochord)과 아가미틈(새열)과 같은 공통 구조 때문에 척삭동물과의 연관성이 제기되어 왔다. 아가미틈의 공유는 두 그룹의 연관성을 지지하지만, 척삭과 입맹관의 상동성에 대해서는 의문이 제기되고 있다. 예를 들어, 척삭 형성에 관여하는 ''Brachyury'' 유전자가 입맹관 형성 시에는 발현되지 않는 등, 두 구조의 상동성을 뒷받침하는 명확한 증거는 부족한 상태이다.
4. 1. Ptychodera flava
''Ptychodera flava''(''P. flava'')는 간접 발생을 하는 반삭동물이다. 초기 난할 패턴은 직접 발생을 하는 ''Saccoglossus kowalevskii''와 유사하다.
''P. flava''의 단세포 수정란에서 시작되는 첫 번째와 두 번째 난할은 등할이며 서로 직교한다. 이 두 분열은 모두 배아의 동물극과 식물극을 통과하여 4개의 난할구를 형성한다. 세 번째 난할 역시 등할이며 적도를 따라 일어나므로, 배아는 동물극과 식물극 각각에 4개의 난할구를 갖게 되어 총 8세포기가 된다.
네 번째 난할은 주로 동물극의 난할구들에서 일어나는데, 이들은 수평으로 등할하여 8개의 동물성 중할구를 형성한다. 반면, 식물극의 4개 난할구는 적도를 따라 부등할하여, 4개의 큰 대할구와 4개의 작은 식물성 소할구를 형성한다. 이 네 번째 분열이 끝나면 배아는 16세포기에 도달하며, 이 배아는 4개의 식물성 소할구, 8개의 동물성 중할구, 그리고 4개의 큰 대할구로 구성된다.
이후 추가적인 분열이 일어나 포배 단계를 완료하고 낭배형성으로 진행된다. 각 할구는 발생 과정에서 다음과 같은 구조로 분화한다.
- 동물성 중할구: 유생 시기의 외배엽을 형성한다. 동물극 할구도 외배엽 형성에 기여하는 것으로 보이지만, 배아마다 정확한 기여도는 다를 수 있다.
- 대할구: 후기 유생 외배엽을 형성한다.
- 식물성 소할구: 내부의 내배엽-중배엽 조직을 만든다.[19]
초기 발생 단계(2세포기 및 4세포기)의 할구를 분리하여 배양해도 각각 완전한 토르나리아 유생으로 발생할 수 있다는 연구 결과가 있다. 이는 이 시기까지는 각 세포의 최종 운명이 완전히 결정되지 않았음을 시사한다.[20] ''P. flava''와 같은 간접 발생형 반삭동물은 척삭동물의 발생 기원을 연구하는 데 중요한 단서를 제공한다.
4. 2. Saccoglossus kowalevskii
''Saccoglossus kowalevskii''는 직접 발생 방식으로 자란다. ''S. kowalevskii''의 알은 원래 타원형이지만, 수정 후에는 구형이 된다.첫 번째 난할은 동물극에서 식물극 방향으로 일어나며, 일반적으로는 등할이지만 부등할이 나타날 수도 있다. 배아가 4세포 단계에 이르는 두 번째 난할 또한 동물극에서 식물극 방향으로 일어나며, 대체로 균등하지만 첫 번째 난할처럼 불균등하게 분열될 수도 있다. 8세포 단계의 난할은 위도 방향(가로 방향)으로 일어나, 4세포 단계의 각 할구가 두 개로 나뉜다. 네 번째 난할은 동물극 쪽 세포에서 먼저 시작되어, 방사대칭이 아닌 8개의 중할구(mesomere|메소미어eng)를 형성한다. 이후 식물극 쪽 4개의 할구가 분열하여 4개의 대할구(macromere|매크로미어eng)와 4개의 매우 작은 미할구(micromere|마이크로미어eng)를 만든다. 다섯 번째 난할은 동물극 쪽 세포에서 먼저, 이후 식물극 쪽 세포에서 일어나 32개의 할구를 형성한다. 여섯 번째 난할도 비슷한 순서로 진행되어 64세포 단계가 되고, 마지막 일곱 번째 난할을 통해 128개의 할구를 가진 포배(blastula)가 형성되면서 난할 과정이 마무리된다.
이 포배는 원장배 형성(gastrulation) 과정을 거치면서 아가미틈(gill slit)을 가진 유충의 기본적인 구조를 갖추게 되며, 최종적으로 성체인 바다 도토리 지렁이로 발생한다.[21][22]
4. 3. 배측-복측 패턴 형성의 유전적 제어
반삭동물의 배측-복측 축 패턴 형성은 척삭동물과 유사한 유전적 메커니즘을 따른다. 많은 유전학적 연구가 척삭동물과의 비교를 통해 이루어졌으며, 반삭동물에서 확인된 유전자 표지 다수가 척삭동물에서도 발견되거나 상동 관계에 있다.특히 장고벌레류의 일종인 ''Saccoglossus kowalevskii''에 대한 연구가 활발히 진행되었다. ''S. kowalevskii''는 척삭동물처럼 배아의 배측(등쪽)에서 BMP 유사 인자를 발현한다. 예를 들어, ''bmp 2/4'' 유전자(초파리의 ''dpp'' 유전자와 상동)는 원장배 형성 시작 시 배아 외배엽 측면에서 발현되기 시작하여, 발생이 진행됨에 따라 배측 중간선으로 발현 영역이 좁아진다. 다만, 항문 뒤 꼬리 부분에서는 발현되지 않는다. BMP 신호를 억제하는 코르딘은 ''S. kowalevskii''의 내배엽에서 발현되어 복측(배쪽) 형성을 유도하는 것으로 보인다.
이러한 주요 조절 인자 외에도 배측-복측 패턴 형성에 관여하는 다른 분자들이 ''S. kowalevskii''에서 발견되었다. 넷린 계열 유전자(넷린 1, 넷린 2)가 그 예이다.[7] 넷린은 척삭동물의 신경계 패턴 형성에 중요한 역할을 한다. 소닉 헤지호그(Shh) 신호 전달 경로 역시 척삭동물 발생에 중요하지만, ''S. kowalevskii''는 단 하나의 헤지호그(hh) 유전자만을 가지며, 이 유전자는 척삭동물에서 일반적으로 발현되는 배측 중간선과는 다른 영역에서 발현되는 것으로 관찰되었다. 이는 반삭동물과 척삭동물 간의 발생 조절 메커니즘에 차이가 있음을 시사한다.
5. 척삭동물과의 관계
반삭동물은 극피동물과 함께 후구동물상문을 형성하며, 현존하는 동물 중 척삭동물과 가장 가까운 근연종으로 여겨진다.[13] 이 때문에 반삭동물은 척삭동물의 발생 및 기원을 연구하는 데 중요한 분류군으로 주목받고 있다.
반삭동물과 척삭동물 사이의 연관성은 인두 부위에 아가미틈(새열)이라는 공통된 구조를 가진다는 점에서 뒷받침된다. 이는 두 동물군이 공통 조상에서 유래했을 가능성을 시사하는 중요한 특징이다.
과거에는 반삭동물의 입 앞쪽에 있는 입 맹관(stomochord) 또는 반삭이라 불리는 구조가 척삭동물의 척삭과 상동기관일 것으로 추정되어 '반삭동물'이라는 이름이 붙었다. 그러나 척삭 형성에 중요한 역할을 하는 것으로 알려진 ''Brachyury'' (Bra) 유전자가 입 맹관 형성 과정에서는 발현되지 않는다는 연구 결과[13] 등, 두 구조의 상동성을 직접적으로 뒷받침하는 증거는 아직 부족하여 이들의 관계에 대해서는 논란이 있다.
척삭동물과 반삭동물의 공통 조상이 어떤 생활 방식을 가졌는지에 대해서는 두 가지 주요 가설이 존재한다.
# 첫 번째 가설은 멍게와 같은 부착성 동물이 공통 조상이며, 이들이 유형성숙(네오테니)과 같은 진화 과정을 통해 장새류나 척추동물 같은 자유 생활 동물을 출현시켰다는 것이다.
# 두 번째 가설은 장새류와 같은 자유 생활성 동물이 공통 조상이며, 이후 익새류나 멍게류와 같은 부착성 동물이 분기했다는 것이다.
최근 18S rDNA 염기서열 분석과 같은 분자계통학적 연구 결과들은 대체로 두 번째 가설, 즉 자유 생활성 동물이 공통 조상이었다는 견해를 지지하는 경향을 보인다.
6. 분류
반삭동물문은 크게 두 개의 강으로 나뉜다: 장새류(Enteropneusta)와 익새류(Pterobranchia)이다.[23][24]
- 장새류: 흔히 '도토리벌레'(acorn worm)라고 불리며, 몸이 길고 유연한 벌레 모양이다. 얕은 바다의 모래나 진흙 속에 서식하며, 몸길이는 수 cm에서 2m에 이르기까지 다양하다. 몸 앞부분이 도토리나 일본식 난간 장식인 보주(擬宝珠)와 비슷하게 생겨 이름 붙여졌다.
- 익새류: 대부분 군체를 이루어 생활하며, 각 개체는 자신이 분비한 관(tube) 속에서 산다. 몸길이는 수 mm 정도로 작고, 몸 중앙에 여러 쌍의 촉수가 달려 있다. 멸종한 필석류(Graptolithina)를 포함한다.[24]

고생대의 중요한 표준화석인 필석류는 익새류와 유사한 구조의 관을 가진 동물군으로, 현재는 익새강의 일부로 분류되기도 한다. 이와 유사한 화석으로는 중국 윈난성의 캄브리아기 초기 지층에서 발견된 윈난조아(Yunnanozoon)가 있으며, 이는 반삭동물에 속하는 것으로 여겨진다.[31]
또한, 유충 단계만 알려진 단일 종(플랑크토스페라)을 기반으로 제안된 플랑크토스페라강(Planctosphaeroidea)이 있다.
6. 1. 계통 발생
반삭동물은 일반적으로 도토리 지렁이라고 불리는 장새류(Enteropneusta)와 필석류(Graptolithina)를 포함하는 익새류(Pterobranchia)의 두 강으로 나뉜다.[24]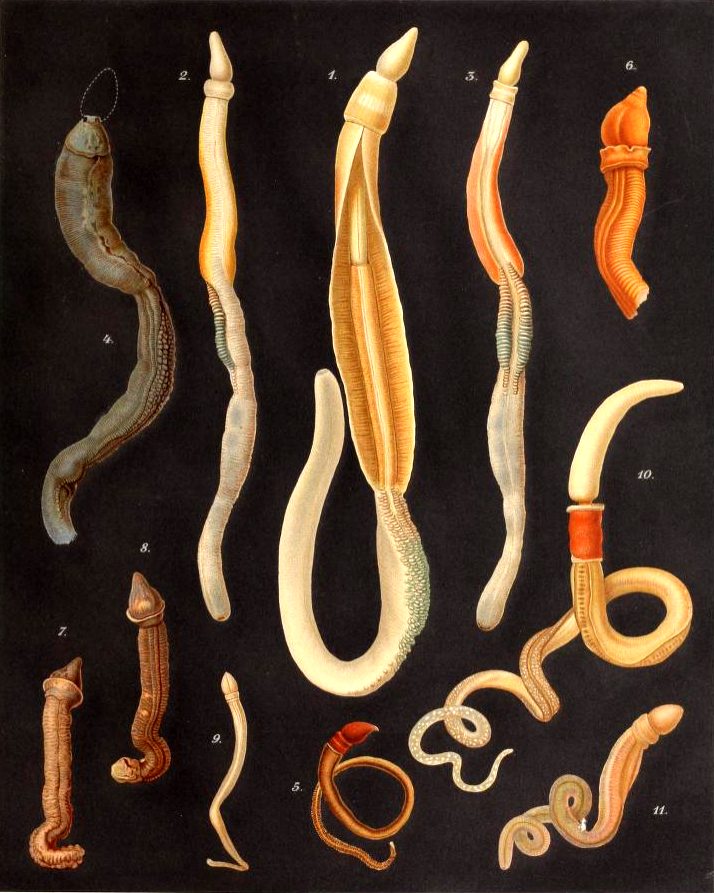
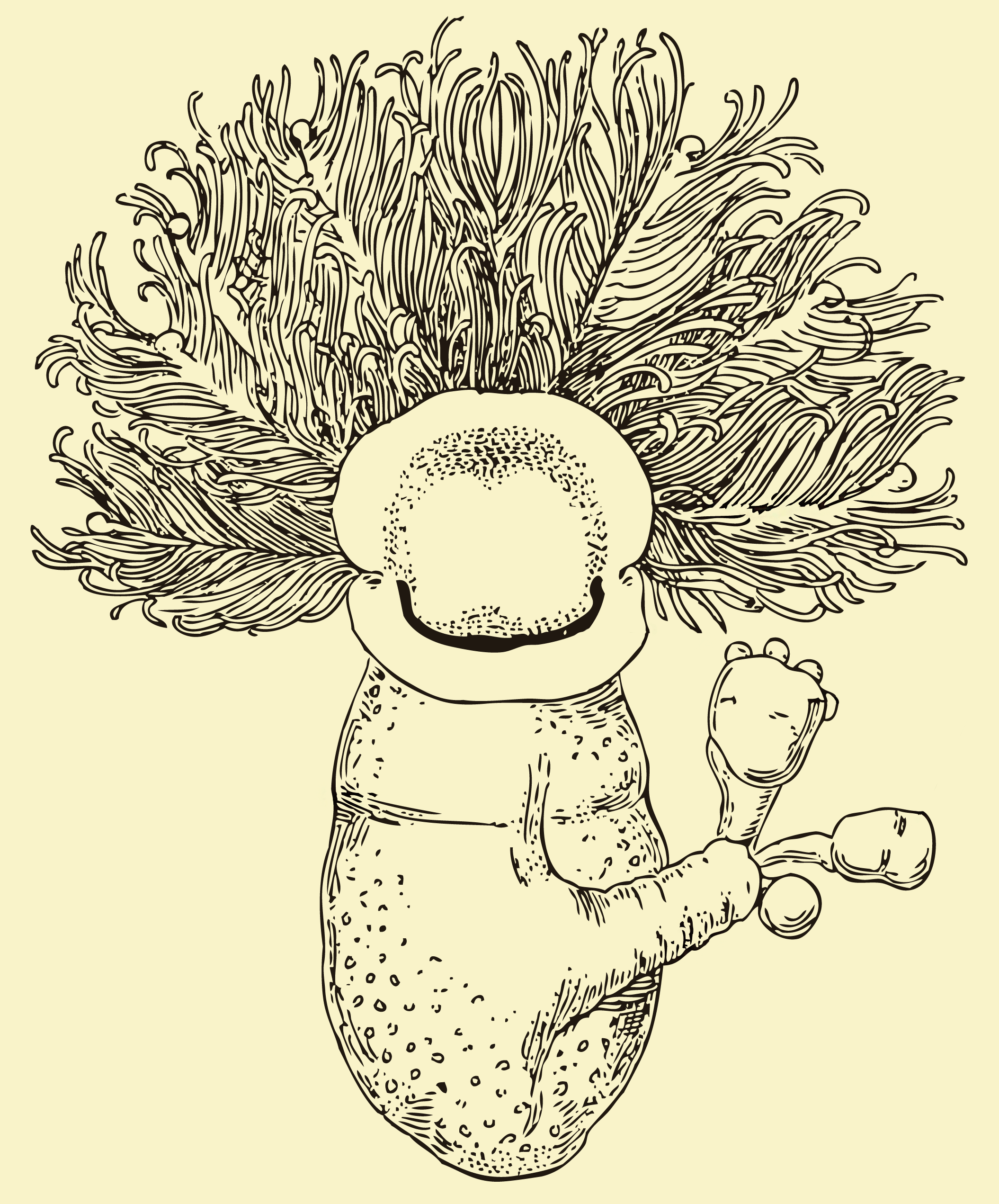
분자 계통학적 연구에 따르면, 반삭동물은 극피동물문과 가장 가까운 자매군을 형성하며, 이 두 문은 암불라크라리아(Ambulacraria)라는 상위 분류군을 이룬다.[28][29][30] 암불라크라리아는 척삭동물과 함께 후구동물 내의 주요 그룹을 구성한다. 따라서 반삭동물은 척삭동물의 발생 기원을 연구하는 데 중요한 동물군으로 여겨진다.[13] 진와충류(Xenoturbellida)가 암불라크라리아 그룹의 기저 분류군일 가능성도 제기된다.
반삭동물 내부의 계통 관계는 16S 및 18S rRNA 염기서열 데이터와 계통유전체학 연구를 통해 밝혀지고 있다.[28][29][30] 이 연구들에 따르면, 익새강(Pterobranchia)은 장새강(Enteropneusta) 내에서 분화된 것으로 나타나, 장새강은 측계통군(paraphyletic group)일 가능성이 있다.
- 장새류 (Enteropneusta): 일부 연구에서는 ''Stereobalanus'' 속이 가장 먼저 분기한 것으로 나타난다. 다른 주요 과로는 Harrimaniidae, Spengeliidae, Torquaratoridae, Ptychoderidae 등이 있다.[28][29][30]
- 익새류 (Pterobranchia): 두흉류(Cephalodiscida)와 필석류(Graptolithina)로 나뉜다. 필석류에는 현존하는 긴목류(Rhabdopleurida)와 멸종한 진필석류(Eugraptolithina)가 포함되며, 진필석류는 다시 수지상류(Dendroidea)와 필석류(Graptoloidea) 등으로 분류된다.[28][29][30] 멸종한 생물인 ''Etacystis''가 반삭동물의 일원이거나 익새류와 밀접한 관련이 있을 가능성도 제기된다.[26]
반삭동물이라는 이름은 과거 척삭동물의 척삭과 상동 기관으로 여겨졌던 입 속의 맹낭(stomochord) 구조 때문에 붙여졌다. 반삭동물과 척삭동물 모두 아가미구멍(gill slit)과 유사한 구조를 가진다는 점에서 두 그룹의 연관성은 인정되지만, 맹낭과 척삭의 상동성에 대해서는 의문이 제기되고 있다. 예를 들어, 척삭 형성에 관여하는 ''Brachyury'' 유전자가 맹낭 형성 시에는 발현되지 않는 등, 두 구조의 상동성을 지지하지 않는 연구 결과들이 있다.
반삭동물과 척삭동물의 기원에 대해서는 두 가지 주요 가설이 있다. 첫 번째는 멍게류와 같은 고착성 동물이 공통 조상이며, 이들이 유형성숙(neoteny)을 통해 창고기나 척추동물과 같은 자유 유영 형태로 진화했다는 설이다. 두 번째는 창고기와 같은 자유 유영 동물이 공통 조상이며, 여기서 고착성 동물들이 분기했다는 설이다. 18S rDNA 염기서열 분석 결과 등은 두 번째 가설, 즉 자유 유영 동물이 공통 조상이라는 설을 지지하는 경향이 있다.
현재까지 약 130종의 반삭동물이 기록되었으며, 특히 심해 탐사를 통해 새로운 종들이 계속 발견되고 있다.[25][27] 유충 단계에서만 알려진 단일 종을 기반으로 제3의 강인 유영구형강(Planctosphaeroidea)이 제안되기도 했다.
참조
[1]
논문
The origins of graptolites and other pterobranchs: a journey from 'Polyzoa'
2008-12
[2]
논문
Tubicolous enteropneusts from the Cambrian period
[3]
논문
Predator deterrence and 2,4-dibromophenol conservation by the enteropneusts, Saccoglossus bromophenolosus and Protoglossus graveolens
[4]
논문
Developmental biology of pterobranch hemichordates: history and perspectives
[5]
잡지
A Cambrian Puzzle: An Ancient, Fossilized Marine Worm Crawls into the Light
Royal Ontario Museum
2020-09-01 # Fall 2020 추정
[6]
논문
The Global Diversity of Hemichordata
[7]
논문
Dorsoventral patterning in hemichordates: insights into early chordate evolution
2006-08-22
[8]
논문
Centralization of the deuterostome nervous system predates chordates
2009-08-11
[9]
논문
Neural nets
[10]
서적
The Brain: Big Bangs, Behaviors, and Beliefs
https://books.google[...]
Yale University Press
2012-04-30
[11]
논문
Biomineral ultrastructure, elemental constitution and genomic analysis of biomineralization-related proteins in hemichordates
[12]
서적
Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth
https://books.google[...]
[13]
논문
Development of Sea Urchins, Ascidians, and Other Invertebrate Deuterostomes: Experimental Approaches
https://archive.org/[...]
[14]
논문
The Spawning and Early Development of the Hawaiian Acorn worm (Hemichordate), Ptycodhera flava
1998-01-01
[15]
논문
Aspects of the reproduction and life cycle of Rhabdopleura compacta (Hemichordata)
[16]
논문
The larva of Rhabdopleura compacta (Hemichordata)
1973-01
[17]
논문
Settlement and metamorphosis of Rhabdopleura normani (Hemichordata: Pterobranchia)
1988-06
[18]
논문
Ultrastructure of adult gonads and development and structure of the larva of Rhabdopleura normani
[19]
논문
Deuterostome evolution: early development in the enteropneust hemichordate, ''Ptychodera flava''
2001-11-01
[20]
논문
The developmental capacities of separated early blastomeres of an enteropneust, ''Saccoglossus kowalevskii''
[21]
논문
Relationships between the egg and larva of Saccoglossus kowalevskii (Enteropneusta): axes and planes; general prospective significance of the early blastomeres
[22]
논문
The normal embryology of saccoglossus kowalevskii (enteropneusta)
1953-05
[23]
논문
Evolution of the chordate body plan: new insights from phylogenetic analyses of deuterostome phyla
2000-04-25
[24]
논문
Zooid morphology and molecular phylogeny of the graptolite ''Rhabdopleura annulata'' (Hemichordata, Pterobranchia) from Heron Island, Australia
[25]
논문
Animal biodiversity: An introduction to higher-level classification and taxonomic richness
http://mapress.com/z[...]
[26]
논문
Etacystis communis, a Fossil of Uncertain Affinities from the Mazon Creek Fauna (Pennsylvanian of Illinois)
http://jpaleontol.ge[...]
1976-11
[27]
논문
The Global Diversity of Hemichordata
[28]
논문
The Global Diversity of Hemichordata
2016-10-04
[29]
논문
Mitogenomics Reveals a Novel Genetic Code in Hemichordata
2019-01-01
[30]
논문
The classification of the Pterobranchia (Cephalodiscida and Graptolithina)
[31]
논문
Yunnantozoon and the ancestry of chordates
http://www.app.pan.p[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com